
Predicting Key Genes and Therapeutic Molecular Modelling to Explain the Association between Porphyromonas gingivalis (P. gingivalis) and Alzheimer’s Disease (AD)

Abstract
:1. Introduction
2. Results
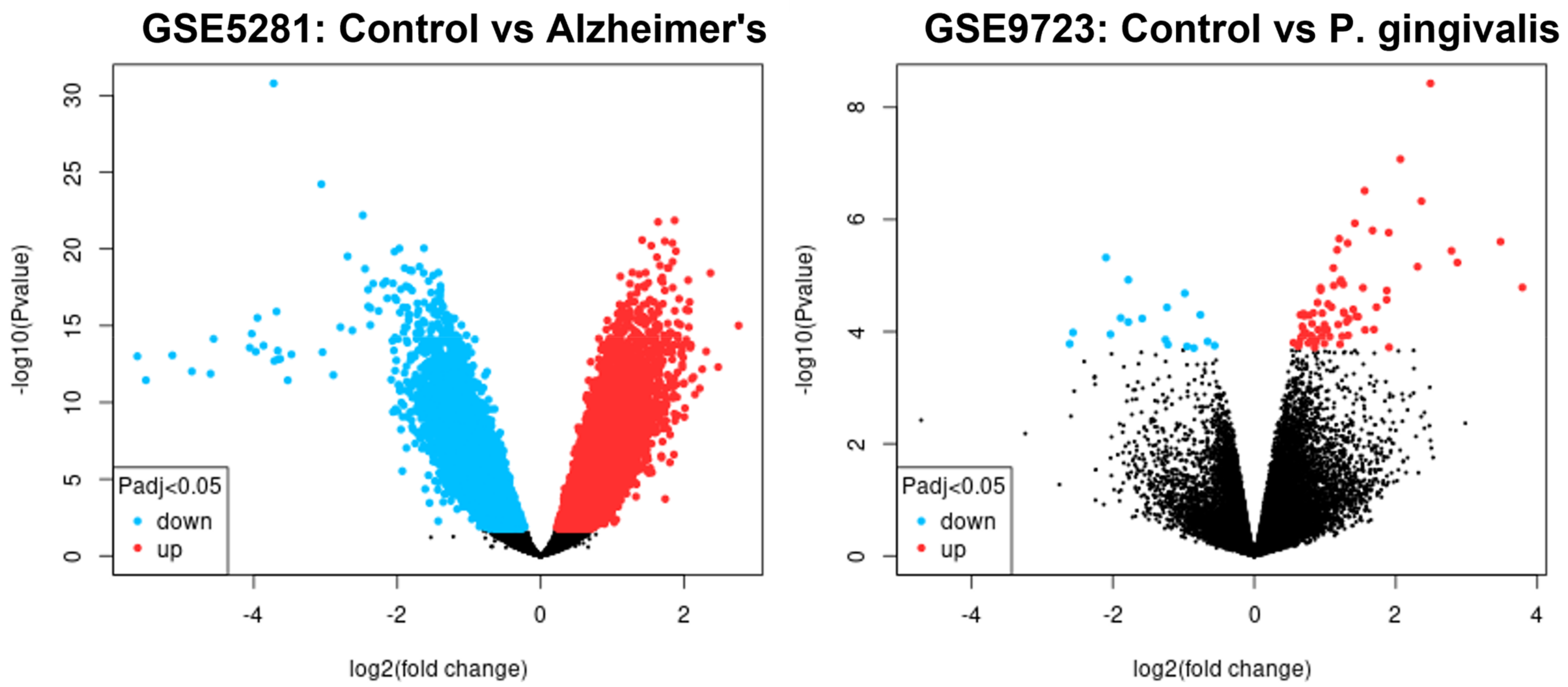
2.1. Data Normalisation
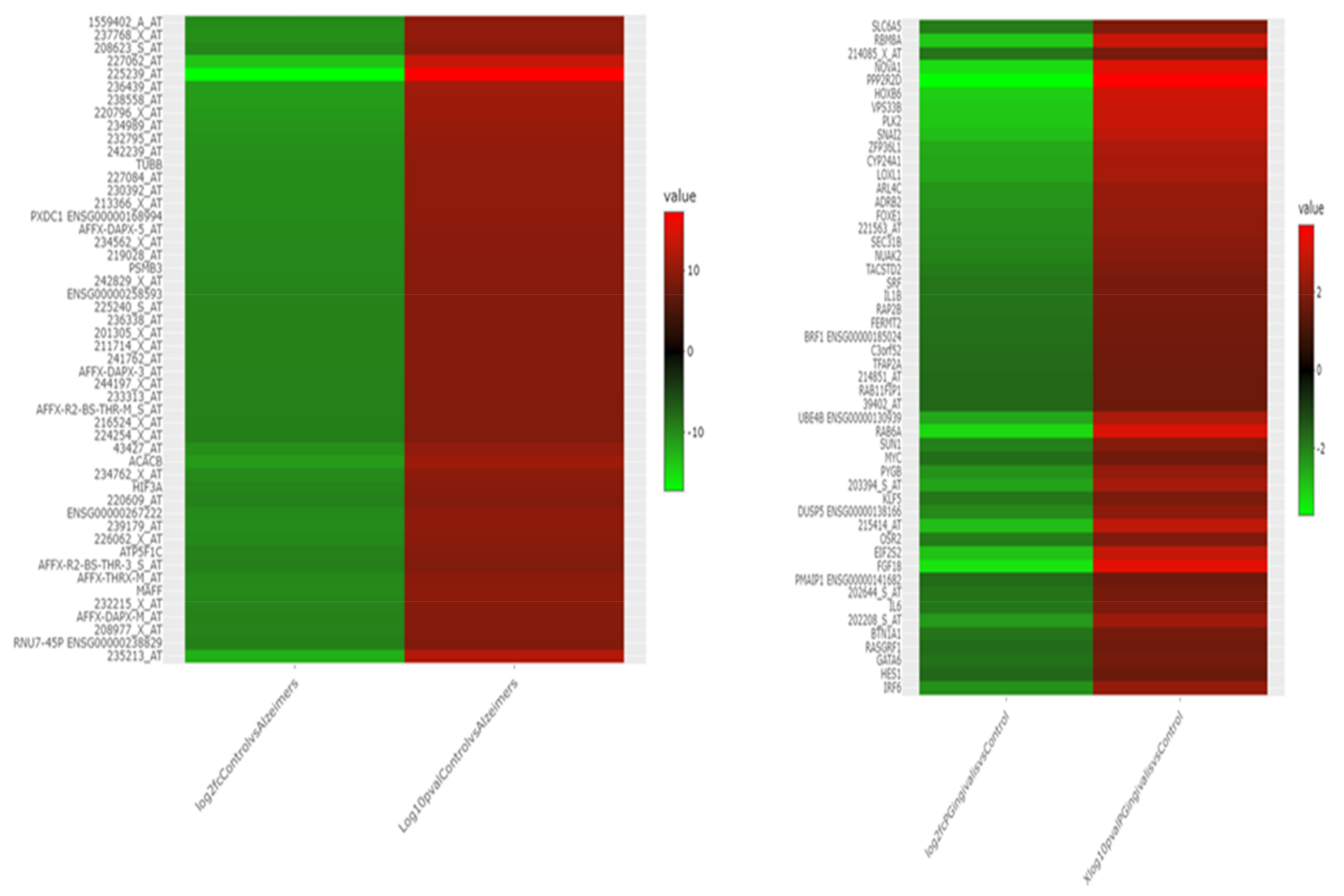
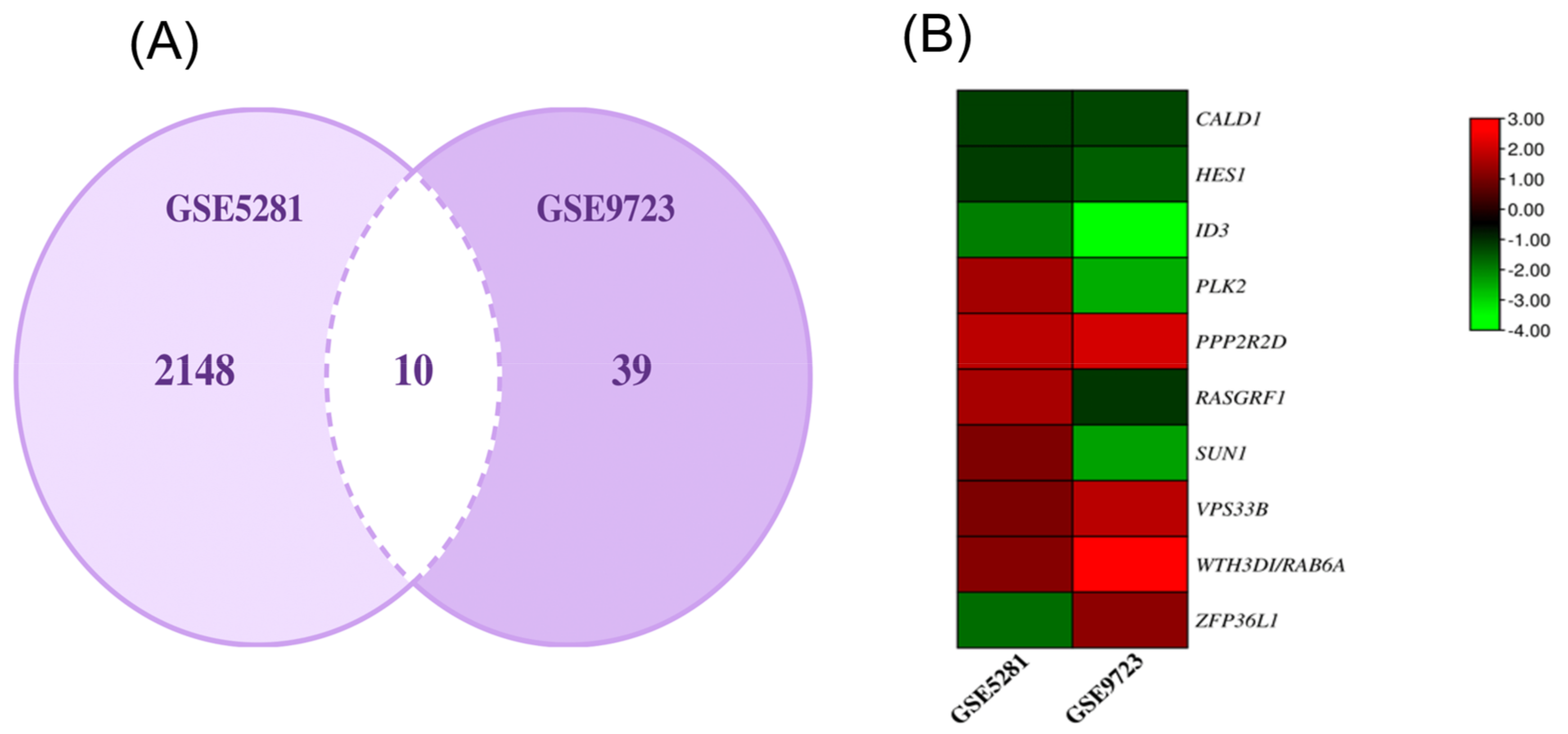
2.2. DEGs Common to AD and P. gingivalis
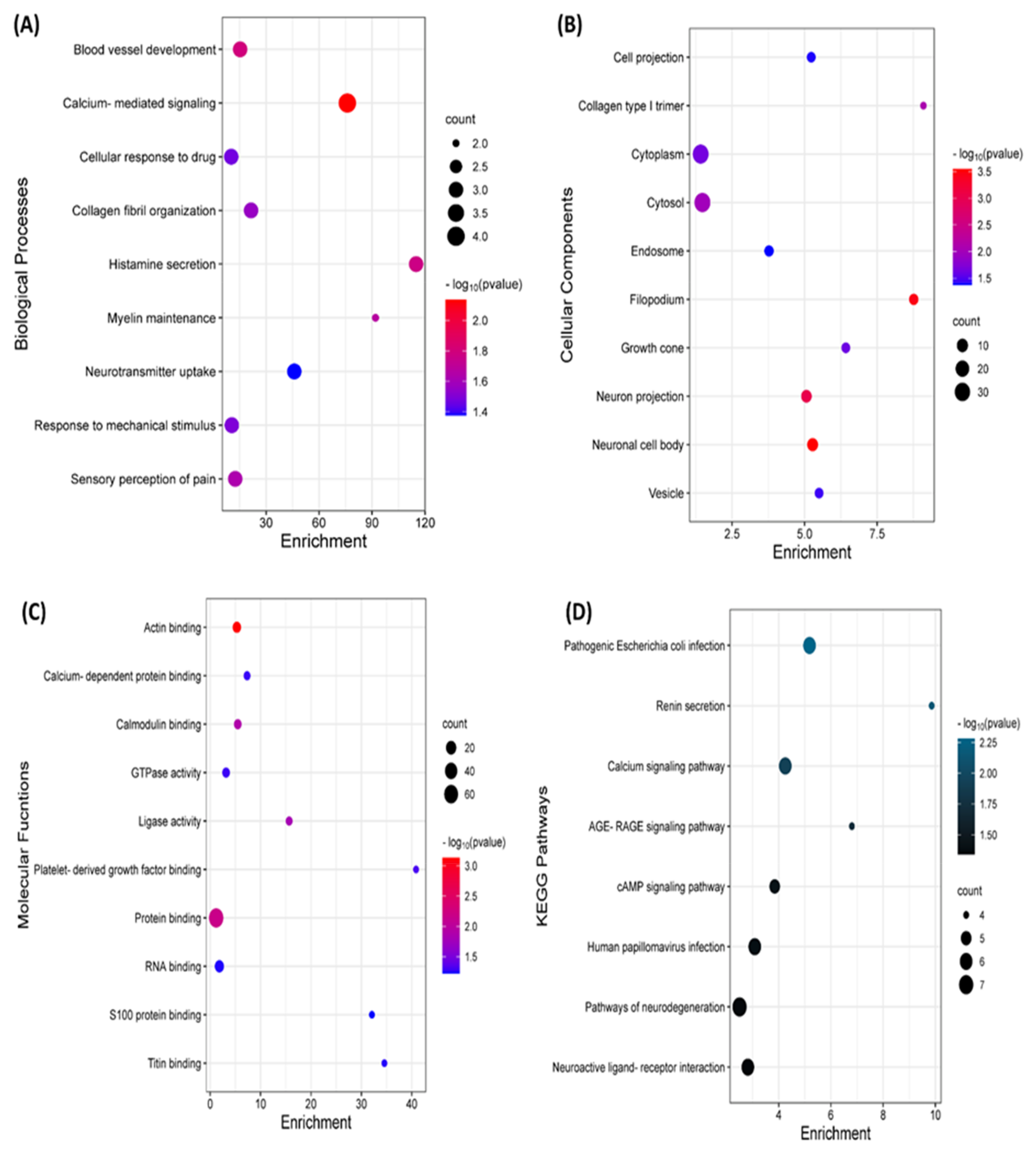
2.3. GO and KEGG Analysis
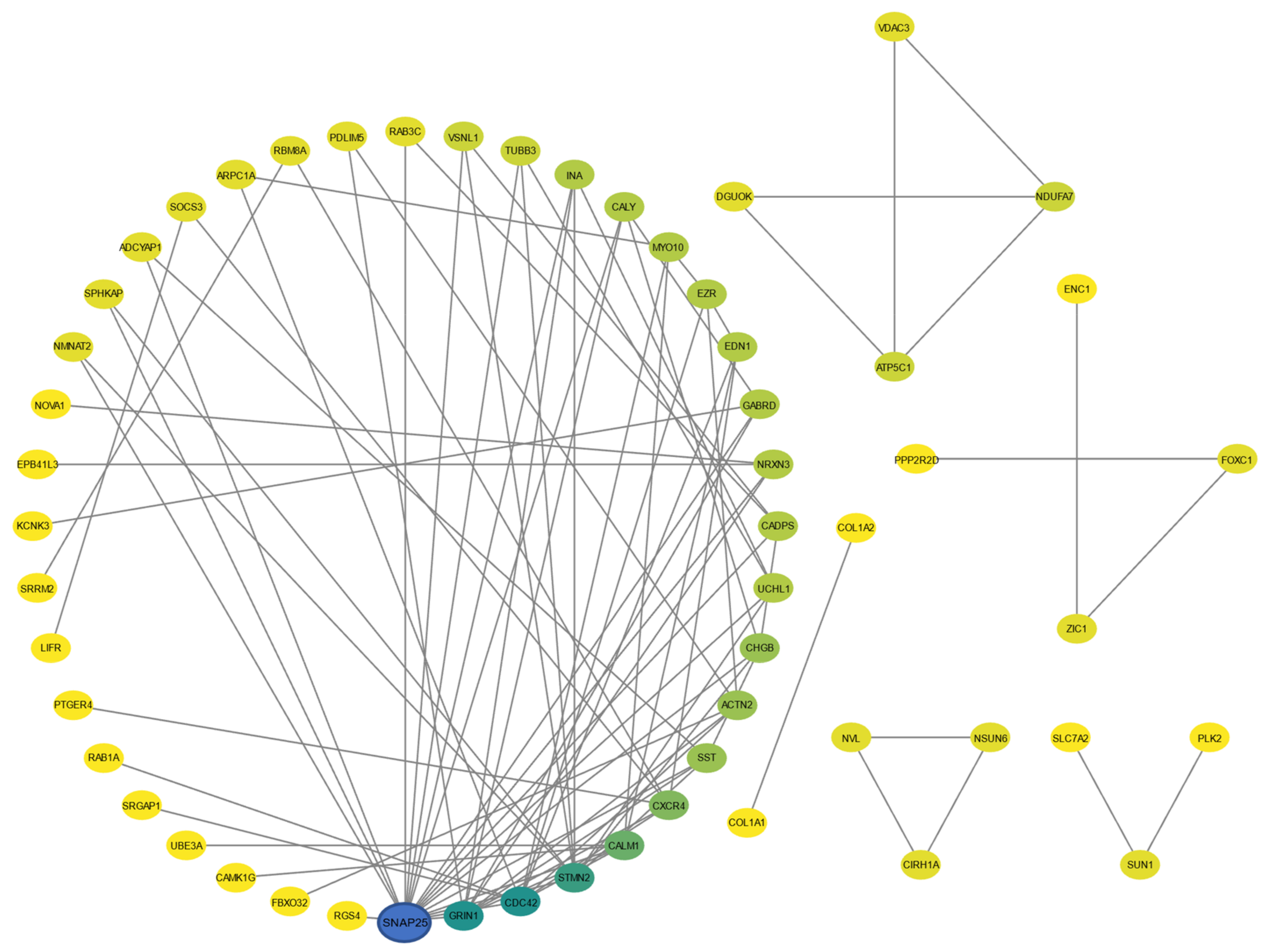
2.4. PPI Network and Hub Gene Selection
2.5. CMap and Molecular Docking
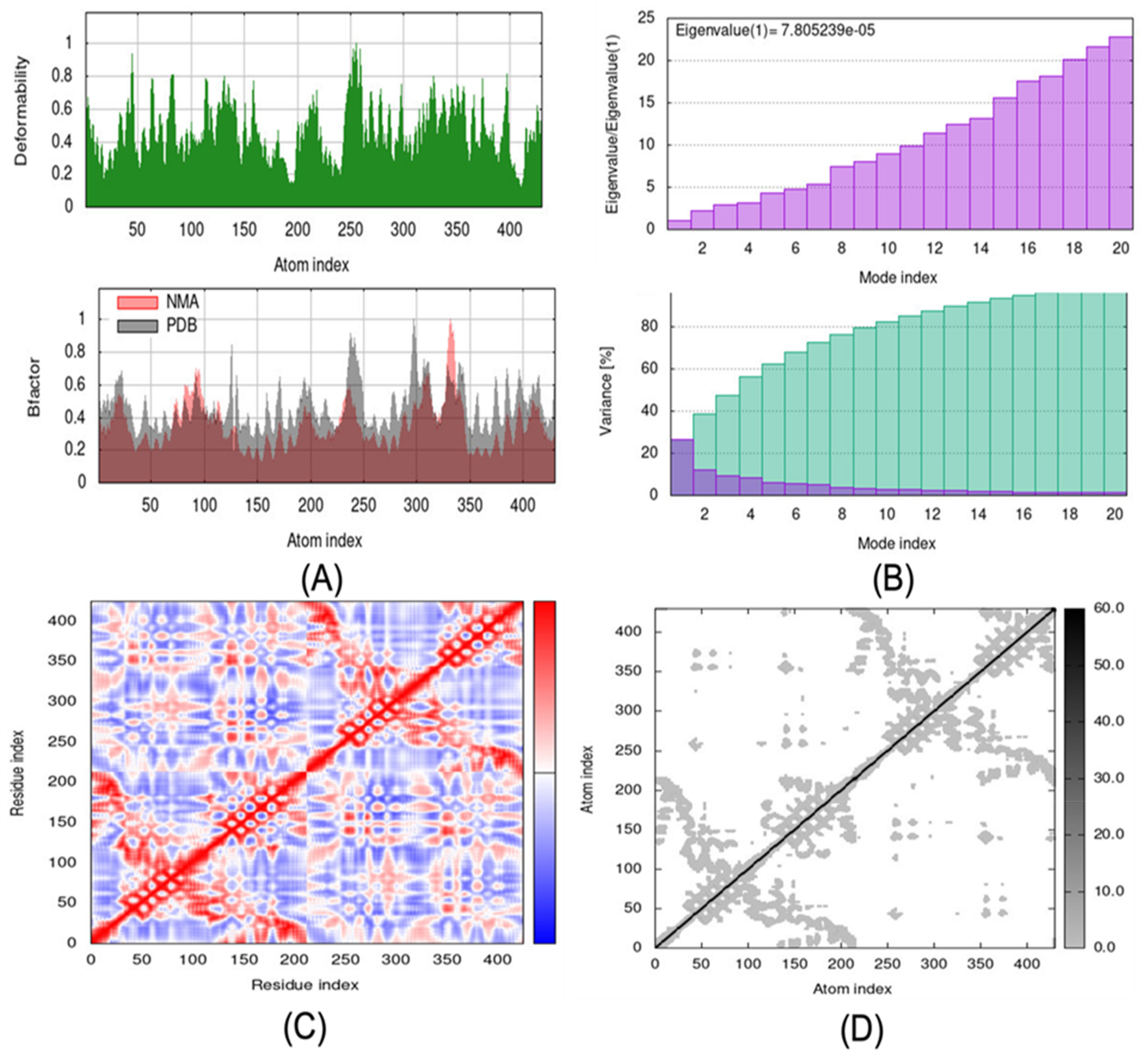
2.6. Molecular Dynamic Simulations
3. Discussion
3.1. Common Genes Found in P.gingivalis and AD
3.2. PPI Network Selected Hub Genes UCHL1, SST, CHGB, CALY, and INA
3.3. Molecular Docking of PLK2 with 11364421 (C28H39N703)
4. Materials and Methods
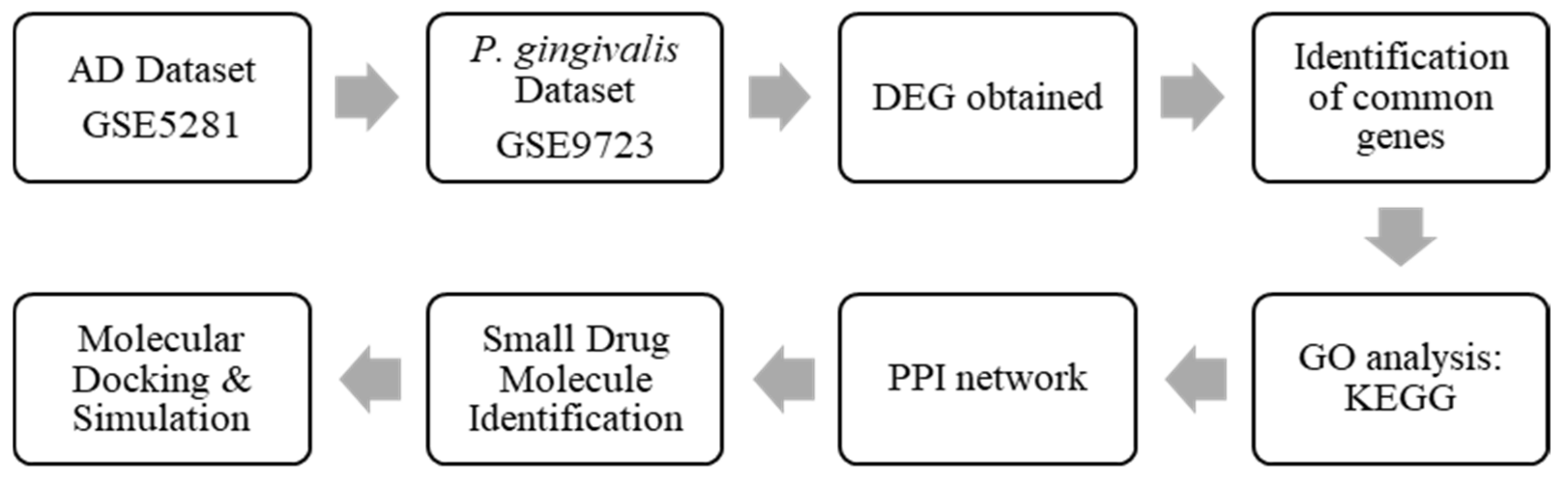
4.1. Pipeline of Research
4.2. Data Acquisition
4.3. DEGs Screening
4.4. Functional Analysis of Common Genes
4.5. Construction of PPI Network
4.6. CMap Analysis, Molecular Docking, and Simulation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ball, S.L.; Holland, A.J.; Watson, P.C.; Huppert, F.A. Theoretical Exploration of the Neural Bases of Behavioural Disinhibition, Apathy and Executive Dysfunction in Preclinical Alzheimer’s Disease in People with Down’s Syndrome: Potential Involvement of Multiple Frontal-Subcortical Neuronal Circuits. J. Intellect. Disabil. Res. 2010, 54, 320–336. [Google Scholar] [CrossRef] [PubMed]
- Jellinger, K.A. Alzheimer’s Disease. Neurobiol. Dis. 2007, 69–82. [Google Scholar] [CrossRef]
- Balolong, M.P.; Mendoza, M.A.F.; Balolong, M.P.; Mendoza, M.A.F. Understanding Oral Diseases: Exploring Opportunities from Filipino Oral Microbiome Research. Dent. Caries 2021. [Google Scholar] [CrossRef]
- Viral Role in Alzheimer’s Disease Discovered|University of Oxford. Available online: https://www.ox.ac.uk/news/2022-08-02-viral-role-alzheimers-disease-discovered (accessed on 25 February 2023).
- Sharma, V.K.; Mehta, V.; Singh, T.G. Alzheimer’s Disorder: Epigenetic Connection and Associated Risk Factors. Curr. Neuropharmacol. 2020, 18, 740. [Google Scholar] [CrossRef] [PubMed]
- Lunnon, K.; Mill, J. Epigenetic Studies in Alzheimer’s Disease: Current Findings, Caveats and Considerations for Future Studies. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. Off. Publ. Int. Soc. Psychiatry Genet. 2013, 162, 789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fülöp, T.; Itzhaki, R.F.; Balin, B.J.; Miklossy, J.; Barron, A.E. Role of Microbes in the Development of Alzheimer’s Disease: State of the Art—An International Symposium Presented at the 2017 IAGG Congress in San Francisco. Front. Genet. 2018, 9, 362. [Google Scholar] [CrossRef] [Green Version]
- Jackson, J.P.; Lindroth, A.M.; Cao, X.; Jacobsen, S.E. Control of CpNpG DNA Methylation by the KRYPTONITE Histone H3 Methyltransferase. Nature 2002, 416, 556–560. [Google Scholar] [CrossRef] [PubMed]
- Balazs, R. Epigenetic Mechanisms in Alzheimer’s Disease. Degener. Neurol. Neuromuscul. Dis. 2014, 4, 85. [Google Scholar] [CrossRef] [Green Version]
- Elwishahy, A.; Antia, K.; Bhusari, S.; Ilechukwu, N.C.; Horstick, O.; Winkler, V. Porphyromonas Gingivalis as a Risk Factor to Alzheimer’s Disease: A Systematic Review. J. Alzheimers Dis. Rep. 2021, 5, 721. [Google Scholar] [CrossRef]
- Mei, F.; Xie, M.; Huang, X.; Long, Y.; Lu, X.; Wang, X.; Chen, L. Porphyromonas gingivalis and Its Systemic Impact: Current Status. Pathogens 2020, 9, 944. [Google Scholar] [CrossRef]
- Li, X.; Kolltveit, K.M.; Tronstad, L.; Olsen, I. Systemic Diseases Caused by Oral Infection. Clin. Microbiol. Rev. 2000, 13, 547. [Google Scholar] [CrossRef]
- O’Brien, M.A.; Costin, B.N.; Miles, M.F. Using Genome-Wide Expression Profiling to Define Gene Networks Relevant to the Study of Complex Traits: From RNA Integrity to Network Topology. Int. Rev. Neurobiol. 2012, 104, 91. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.; Zakharkin, S.O.; Allison, D.B. Expectations, Validity, and Reality in Gene Expression Profiling. J. Clin. Epidemiol. 2010, 63, 950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olsen, I.; Singhrao, S.K. Interaction between Genetic Factors, Porphyromonas Gingivalis and Microglia to Promote Alzheimer’s Disease. J. Oral Microbiol. 2020, 12, 1820834. [Google Scholar] [CrossRef] [PubMed]
- What Causes Alzheimer’s Disease?|National Institute on Aging. Available online: https://www.nia.nih.gov/health/what-causes-alzheimers-disease (accessed on 6 January 2023).
- Kumari, A.; Rahaman, A.; Zeng, X.-A.; Farooq, M.A.; Huang, Y.; Yao, R.; Ali, M.; Ishrat, R.; Ali, R. Temporal Cortex Microarray Analysis Revealed Impaired Ribosomal Biogenesis and Hyperactivity of the Glutamatergic System: An Early Signature of Asymptomatic Alzheimer’s Disease. Front. Neurosci. 2022, 16, 966877. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.C.; Gray, J.D.; Kogan, J.F.; Davidson, R.L.; Rubin, T.G.; Okamoto, M.; Morrison, J.H.; McEwen, B.S. Age and Alzheimer’s Disease Gene Expression Profiles Reversed by the Glutamate Modulator Riluzole. Mol. Psychiatry 2017, 22, 296. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Wu, Q.; Su, J.; Wang, W.; Zhao, X.; Cao, P. Circulating MiRNAs as Epigenetic Mediators of Periodontitis and Preeclampsia Association. Dis. Markers 2022, 2022, 2771492. [Google Scholar] [CrossRef]
- Gonzalez, O.A.; Kirakodu, S.; Nguyen, L.M.; Orraca, L.; Novak, M.J.; Gonzalez-Martinez, J.; Ebersole, J.L. Comparative Analysis of Gene Expression Patterns for Oral Epithelial Cell Functions in Periodontitis. Front. Oral Health 2022, 3, 863231. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, P.; Ren, L.; Hu, C.; Bi, J. Protective Effect of Melatonin on Soluble Aβ1-42-Induced Memory Impairment, Astrogliosis, and Synaptic Dysfunction via the Musashi1/Notch1/Hes1 Signaling Pathway in the Rat Hippocampus. Alzheimer’s Res. Ther. 2016, 8, 40. [Google Scholar] [CrossRef] [Green Version]
- Woo, B.H.; Kim, D.J.; Choi, J.I.; Kim, S.J.; Park, B.S.; Song, J.M.; Lee, J.H.; Park, H.R. Oral Cancer Cells Sustainedly Infected with Porphyromonas Gingivalis Exhibit Resistance to Taxol and Have Higher Metastatic Potential. Oncotarget 2017, 8, 46981. [Google Scholar] [CrossRef] [Green Version]
- Ellmeier, W.; Aguzzi, A.; Kleiner, E.; Kurzbauer, R.; Weith, A. Mutually Exclusive Expression of a Helix-Loop-Helix Gene and N-Myc in Human Neuroblastomas and in Normal Development. EMBO J. 1992, 11, 2563–2571. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Roberts, J.A.S.; Atanasova, K.R.; Chowdhury, N.; Han, K.; Yilmaz, Ö. Human Primary Epithelial Cells Acquire an Epithelial-Mesenchymal-Transition Phenotype during Long-Term Infection by the Oral Opportunistic Pathogen, Porphyromonas Gingivalis. Front. Cell. Infect. Microbiol. 2017, 7, 493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebersole, J.L.; Al-Sabbagh, M.; Dawson, D.R. Heterogeneity of Human Serum Antibody Responses to P. Gingivalis in Periodontitis: Effects of Age, Race/Ethnicity, and Sex. Immunol. Lett. 2020, 218, 11. [Google Scholar] [CrossRef]
- Martínez-Drudis, L.; Sheta, R.; Pellegrinato, R.; Loukili, S.; Tremblay, C.; Calon, F.; Rivest, S.; Oueslati, A. Inhibition of PLK2 Activity Affects APP and Tau Pathology and Improves Synaptic Content in a Sex-Dependent Manner in a 3xTg Mouse Model of Alzheimer’s Disease. Neurobiol. Dis. 2022, 172, 105833. [Google Scholar] [CrossRef]
- de Cárcer, G. The Mitotic Cancer Target Polo-Like Kinase 1: Oncogene or Tumor Suppressor? Genes 2019, 10, 208. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.Y.; Chi, Y.H.; Mutalif, R.A.; Starost, M.F.; Myers, T.G.; Anderson, S.A.; Stewart, C.L.; Jeang, K.T. Accumulation of the Inner Nuclear Envelope Protein Sun1 Is Pathogenic in Progeric and Dystrophic Laminopathies. Cell 2012, 149, 565–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visanji, N.P.; Sarvestani, I.K.; Creed, M.C.; Shoaei, Z.S.; Nobrega, J.N.; Hamani, C.; Hazrati, L.N. Deep Brain Stimulation of the Subthalamic Nucleus Preferentially Alters the Translational Profile of Striatopallidal Neurons in an Animal Model of Parkinson’s Disease. Front. Cell. Neurosci. 2015, 9, 221. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Hou, J.; Wu, Y.; Liu, Y.; Li, R.; Xu, T.; Liu, J.; Pan, Y. Distinct Gene Expression Characteristics in Epithelial Cell-Porphyromonas Gingivalis Interactions by Integrating Transcriptome Analyses. Int. J. Med. Sci. 2019, 16, 1320. [Google Scholar] [CrossRef] [Green Version]
- Ottoboni, L.; Radassi, K.; De Jager, P.; Comi, G.; Farina, C.; Martino, G. Characterization of ZFP36L1 in the Context of Multiple Sclerosis and Functional Immunological Consequences Associated with the Susceptibility to the Disease. J. Neuroimmunol. 2014, 275, 52. [Google Scholar] [CrossRef]
- Park, P.H.; McMullen, M.R.; Huang, H.; Thakur, V.; Nagy, L.E. Short-Term Treatment of RAW264.7 Macrophages with Adiponectin Increases Tumor Necrosis Factor-α (TNF-α) Expression via ERK1/2 Activation and Egr-1 Expression: Role of TNF-α in adiponectin-stimulated interleukin-10 production. J. Biol. Chem. 2007, 282, 21695–21703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shrestha, A.; Pun, N.T.; Park, P.H. ZFP36L1 and AUF1 Induction Contribute to the Suppression of Inflammatory Mediators Expression by Globular Adiponectin via Autophagy Induction in Macrophages. Biomol. Ther. 2018, 26, 446. [Google Scholar] [CrossRef] [PubMed]
- Doran, J.F.; Jackson, P.; Kynoch, P.A.M.; Thompson, R.J. Isolation of PGP 9.5, a New Human Neurone-Specific Protein Detected by High-Resolution Two-Dimensional Electrophoresis. J. Neurochem. 1983, 40, 1542–1547. [Google Scholar] [CrossRef] [PubMed]
- Das, C.; Hoang, Q.Q.; Kreinbring, C.A.; Luchansky, S.J.; Meray, R.K.; Ray, S.S.; Lansbury, P.T.; Ringe, D.; Petsko, G.A. Structural Basis for Conformational Plasticity of the Parkinson’s Disease-Associated Ubiquitin Hydrolase UCH-L1. Proc. Natl. Acad. Sci. USA 2006, 103, 4675–4680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Z.B.; Wu, L.; Xiong, R.; Wang, L.L.; Zhang, B.; Wang, C.; Li, H.; Liang, L.; Chen, S.D. MicroRNA-922 Promotes Tau Phosphorylation by Downregulating Ubiquitin Carboxy-Terminal Hydrolase L1 (UCHL1) Expression in the Pathogenesis of Alzheimer’s Disease. Neuroscience 2014, 275, 232–237. [Google Scholar] [CrossRef]
- Oddo, S. The Ubiquitin-Proteasome System in Alzheimer’s Disease. J. Cell. Mol. Med. 2008, 12, 363. [Google Scholar] [CrossRef] [Green Version]
- Lin, L.; Li, S.; Hu, S.; Yu, W.; Jiang, B.; Mao, C.; Li, G.; Yang, R.; Miao, X.; Jin, M.; et al. UCHL1 Impairs Periodontal Ligament Stem Cell Osteogenesis in Periodontitis. J. Dent. Res. 2022, 102, 00220345221116031. [Google Scholar] [CrossRef]
- Cobb, C.M.; Singla, O.; Feil, P.H.; Theiesen, F.C.; Schultz, R.E. Comparison of NK-Cell (Leu-7+ and Leu-11b+) Populations in Clinically Healthy Gingiva, Chronic Gingivitis and Chronic Adult Periodontitis. J. Periodontal Res. 1989, 24, 1–7. [Google Scholar] [CrossRef]
- Jiang, Z.; Shi, Y.; Zhao, W.; Zhou, L.; Zhang, B.; Xie, Y.; Zhang, Y.; Tan, G.; Wang, Z. Association between Chronic Periodontitis and the Risk of Alzheimer’s Disease: Combination of Text Mining and GEO Dataset. BMC Oral Health 2021, 21, 466. [Google Scholar] [CrossRef]
- Solarski, M.; Wang, H.; Wille, H.; Schmitt-Ulms, G. Somatostatin in Alzheimer’s Disease: A New Role for an Old Player. Prion 2018, 12, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Saido, T.C. Metabolism of Amyloid β Peptide and Pathogenesis of Alzheimer’s Disease. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2013, 89, 321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Memmert, S.; Damanaki, A.; Nokhbehsaim, M.; Nogueira, A.V.B.; Eick, S.; Cirelli, J.A.; Jäger, A.; Deschner, J. Regulation of Somatostatin Receptor 2 by Proinflammatory, Microbial and Obesity-Related Signals in Periodontal Cells and Tissues. Head Face Med. 2019, 15, 2. [Google Scholar] [CrossRef] [PubMed]
- Lechner, T.; Adlassnig, C.; Humpel, C.; Kaufmann, W.A.; Maier, H.; Reinstadler-Kramer, K.; Hinterhölzl, J.; Mahata, S.K.; Jellinger, K.A.; Marksteiner, J. Chromogranin Peptides in Alzheimer’s Disease. Exp. Gerontol. 2004, 39, 101–113. [Google Scholar] [CrossRef]
- Heijtz, R.D.; Alexeyenko, A.; Xavier, F.X. Calcyon MRNA Expression in the Frontal-Striatal Circuitry and Its Relationship to Vesicular Processes and ADHD. Behav. Brain Funct. BBF 2007, 3, 33. [Google Scholar] [CrossRef] [Green Version]
- Muthusamy, N.; Chen, Y.J.; Yin, D.M.; Mei, L.; Bergson, C. Complementary Roles of the Neuron-Enriched Endosomal Proteins NEEP21 and Calcyon in Neuronal Vesicle Trafficking. J. Neurochem. 2015, 132, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Vazdarjanova, A.; Bunting, K.; Muthusamy, N.; Bergson, C. Calcyon Up-Regulation in Adolescence Impairs Response Inhibition and Working Memory in Adulthood. Mol. Psychiatry 2011, 16, 672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levavasseur, F.; Zhu, Q.; Julien, J.P. No Requirement of α-Internexin for Nervous System Development and for Radial Growth of Axons. Mol. Brain Res. 1999, 69, 104–112. [Google Scholar] [CrossRef]
- Lariviere, R.C.; Julien, J.P. Functions of Intermediate Filaments in Neuronal Development and Disease. J. Neurobiol. 2004, 58, 131–148. [Google Scholar] [CrossRef]
- Wang, P.; Wang, S.M.; Hsieh, C.J.; Chien, C.L. Neural Expression of α-Internexin Promoter In Vitro and In Vivo. J. Cell. Biochem. 2006, 97, 275–287. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.; Domrachev, M.; Lash, A.E. Gene Expression Omnibus: NCBI Gene Expression and Hybridization Array Data Repository. Nucleic Acids Res. 2002, 30, 207–210. [Google Scholar] [CrossRef] [Green Version]
- Readhead, B.; Haure-Mirande, J.V.; Funk, C.C.; Richards, M.A.; Shannon, P.; Haroutunian, V.; Sano, M.; Liang, W.S.; Beckmann, N.D.; Price, N.D.; et al. Multiscale Analysis of Independent Alzheimer’s Cohorts Finds Disruption of Molecular, Genetic, and Clinical Networks by Human Herpesvirus. Neuron 2018, 99, 64–82.e7. [Google Scholar] [CrossRef] [Green Version]
- Liang, W.S.; Dunckley, T.; Beach, T.G.; Grover, A.; Mastroeni, D.; Walker, D.G.; Caselli, R.J.; Kukull, W.A.; McKeel, D.; Morris, J.C.; et al. Gene Expression Profiles in Anatomically and Functionally Distinct Regions of the Normal Aged Human Brain. Physiol. Genom. 2007, 28, 311–322. [Google Scholar] [CrossRef]
- Handfield, M.; Mans, J.J.; Zheng, G.; Lopez, M.C.; Mao, S.; Progulske-Fox, A.; Narasimhan, G.; Baker, H.V.; Lamont, R.J. Distinct Transcriptional Profiles Characterize Oral Epithelium-Microbiota Interactions. Cell. Microbiol. 2005, 7, 811–823. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and Integrative Analysis of Large Gene Lists Using DAVID Bioinformatics Resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Blake, J.A.; Christie, K.R.; Dolan, M.E.; Drabkin, H.J.; Hill, D.P.; Ni, L.; Sitnikov, D.; Burgess, S.; Buza, T.; Gresham, C.; et al. Gene Ontology Consortium: Going Forward. Nucleic Acids Res. 2015, 43, D1049–D1056. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING Database in 2021: Customizable Protein–Protein Networks, and Functional Characterization of User-Uploaded Gene/Measurement Sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Chemical Structure | IUPAC Name |
|---|---|---|
| 24971422 | C21H17N3O3 | 3-(1,3-benzodioxol-5-yl)-N-[(1S)-1-phenylethyl]-[1,2] oxazolo [5,4-c] pyridin-5-amine |
| 11364421 | C28H39N7O3 | 4-[[(7R)-8-cyclopentyl-7-ethyl-5-methyl-6-oxo-7H-pteridin-2-yl] amino]-3-methoxy-N-(1-methylpiperidin-4-yl) benzamide |
| 49792852 | C24H27F3N8O3 | 1-(2-hydroxyethyl)-8-[5-(4-methylpiperazin-1-yl)-2-(trifluoromethoxy) anilino]-4,5-dihydropyrazolo[4,3-h] quinazoline-3-carboxamide |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamarsha, A.; Balachandran, K.; Sailan, A.T.; Nasruddin, N.S. Predicting Key Genes and Therapeutic Molecular Modelling to Explain the Association between Porphyromonas gingivalis (P. gingivalis) and Alzheimer’s Disease (AD). Int. J. Mol. Sci. 2023, 24, 5432. https://doi.org/10.3390/ijms24065432
Hamarsha A, Balachandran K, Sailan AT, Nasruddin NS. Predicting Key Genes and Therapeutic Molecular Modelling to Explain the Association between Porphyromonas gingivalis (P. gingivalis) and Alzheimer’s Disease (AD). International Journal of Molecular Sciences. 2023; 24(6):5432. https://doi.org/10.3390/ijms24065432
Chicago/Turabian StyleHamarsha, Ahmed, Kumarendran Balachandran, Ahmad Tarmidi Sailan, and Nurrul Shaqinah Nasruddin. 2023. "Predicting Key Genes and Therapeutic Molecular Modelling to Explain the Association between Porphyromonas gingivalis (P. gingivalis) and Alzheimer’s Disease (AD)" International Journal of Molecular Sciences 24, no. 6: 5432. https://doi.org/10.3390/ijms24065432