
Classification of Common Food Lipid Sources Regarding Healthiness Using Advanced Lipidomics: A Four-Arm Crossover Study

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results
2.1. Characteristics of the Sample
2.2. Impact of the Test Diets on Clinical Parameters
2.3. Impact of the Test Diets on Lipidomics
3. Discussion
4. Material and Methods
4.1. Participants
4.2. Inclusion Criteria
4.3. Exclusion Criteria
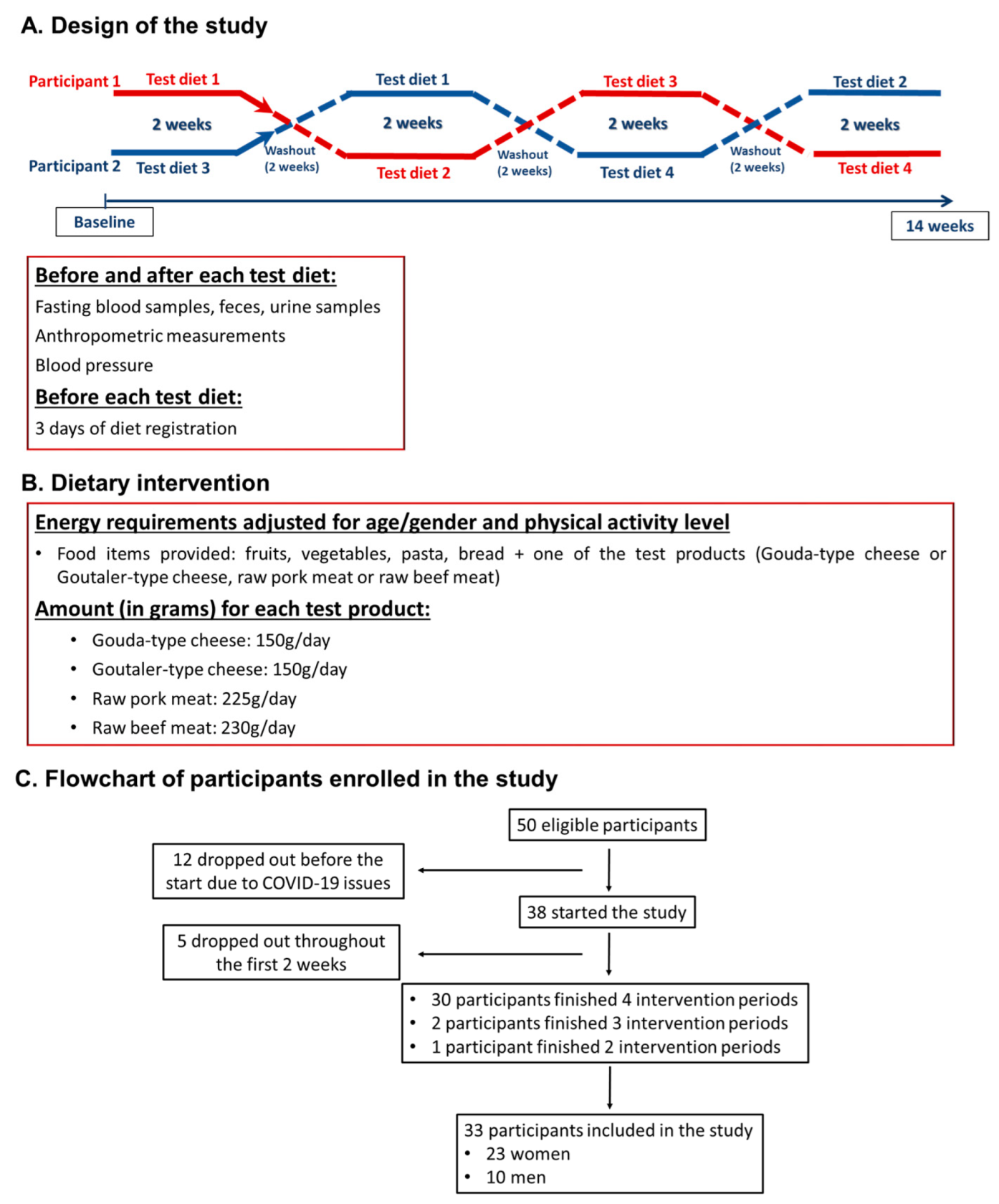
4.4. Design of the Study and Research Protocol
4.5. Test Diets
4.6. Food Composition Analysis
4.7. Diet Assessment
4.8. Health Questionnaire, Risk Analysis, and Physical Activity
4.9. Anthropometric Measurementsand Clinical Variables
4.10. Blood Sampling
4.11. Analytical Procedures
4.12. Lipidomics and Lipoprotein Profile
4.13. Power Calculation
4.14. Statistical Analyses: Clinical and Biochemical Data
4.15. Lipidomics and NMR Lipoprotein Profile Data
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Acylcarnitines (ACs) |
| Alanine transaminase(ALT) |
| Aspartate transaminase (AST) |
| Bile acids (BAs) |
| Body mass index (BMI) |
| Cardiovascular diseases (CVDs) |
| Ceramides (Cers) |
| Cholesteryl esters (ChoEs) |
| Diglycerides (DGs) |
| Fatty acids (FAs) |
| Free sphingoid bases (FSBs) |
| Homeostatic Model Assessment for insulin resistance (HOMA-IR) |
| Interleukin 1-beta (IL-1β) |
| Interleukin-6 (IL-6) |
| International Physical Activity Questionnaire (IPAQ) |
| Liquid chromatography–mass spectrometry (LC-MS) |
| Lyso-PE (LPE) |
| Lyso-PI (LPI) |
| Lyso-PC (LPC) |
| Monohexosylceramides (CMHs) |
| Monounsaturated FAs (MUFAs) |
| N-acyl ethanolamines (NAEs) |
| Non-communicable chronic diseases (NCDs) |
| Nuclear magnetic resonance (NMR) |
| Oxidized FAs (oxFAs) |
| Plasmanyl-PC or ether-linked PC (O-PC) |
| Plasmenyl-PC or vinyl-ether-linked PC (P-PC) |
| Plasmanyl-LPC or ether-linked PC (O-LPC) |
| Plasmenyl-LPC or vinyl-ether-linked LPC (P-LPC) |
| Plasmanyl-PE or ether-linked PE (O-PE) |
| Plasmenyl-PE or vinyl-ether-linked PE (P-PE) |
| Plasmanyl-LPC or ether-linked PE (O-LPE) |
| Plasmenyl-LPE or vinyl-ether-linked LPE (P-LPE) |
| Phosphatidylcholines (PCs) |
| Phosphatidylethanolamines (PEs) |
| Phosphatidylinositols (PIs) |
| Polyunsaturated FAs (PUFAs) |
| Saturated FAs (SFAs) |
| Sphingomyelins (SMs) |
| Steroids (STs) |
| Total energy intake (TEI) |
| Triglycerides (TGs) |
References
- GBD 2017 Diet Collaborators. Health effects of dietary risks in 195 countries, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2019, 393, 1958–1972. [Google Scholar] [CrossRef] [Green Version]
- GBD 2013 Risk Factors Collaborators. Global, regional, and national comparative risk assessment of 79 behavioural, environmental and occupational, and metabolic risks or clusters of risks in 188 countries, 1990–2013: A systematic analysis for the Global Burden of Disease Study 2013. Lancet 2015, 386, 2287–3233. [Google Scholar] [CrossRef] [Green Version]
- Milanski, M.; Degasperi, G.; Coope, A.; Morari, J.; Denis, R.; Cintra, D.E.; Tsukumu, D.M.L.; Anhe, G.; Amaral, M.E.; Takashi, H.K.; et al. Saturated fatty acids produce an inflammatory response predominantly through the activation of TLR4 signaling in hypothalamus: Implications for the pathogenesis of obesity. J. Neurosci. 2009, 29, 359–370. [Google Scholar] [CrossRef] [Green Version]
- Rocha, D.M.; Caldas, A.P.; Oliveira, L.L.; Bressan, J.; Hermsdorff, H.H. Saturated fatty acids trigger TLR4-mediated inflammatory response. Atherosclerosis 2016, 244, 211–215. [Google Scholar] [CrossRef] [PubMed]
- Hunter, J.E.; Zhang, J.; Kris-Etherton, P.M. Cardiovascular disease risk of dietary stearic acid compared with trans, other saturated, and unsaturated fatty acids: A systematic review. Am. J. Clin. Nutr. 2010, 91, 46–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, H.; Kokoeva, M.V.; Inouye, K.; Tzameli, I.; Yin, H.; Flier, J.S. TLR4 links innate immunity and fatty acid-induced insulin resistance. J. Clin. Investig. 2006, 116, 3015–3025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.A.; Vijayakumar, A.; Kahn, B.B. Metabolites as regulators of insulin sensitivity and metabolism. Nat. Rev. Mol. Cell Biol. 2018, 19, 654–672. [Google Scholar] [CrossRef] [PubMed]
- de Souza, R.J.; Mente, A.; Maroleanu, A.; Cozma, A.I.; Ha, V.; Kishibe, T.; Uleryk, E.; Budylowski, P.; Schünemann, H.; Beyene, J.; et al. Intake of saturated and trans unsaturated fatty acids and risk of all cause mortality, cardiovascular disease, and type 2 diabetes: Systematic review and meta-analysis of observational studies. BMJ 2015, 351, h3978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siri-Tarino, P.W.; Sun, Q.; Hu, F.B.; Krauss, R.M. Meta-analysis of prospective cohort studies evaluating the association of saturated fat with cardiovascular disease. Am. J. Clin. Nutr. 2010, 91, 535–546. [Google Scholar] [CrossRef] [Green Version]
- de Oliveira Otto, M.C.; Mozaffarian, D.; Kromhout, D.; Bertoni, A.G.; Sibley, C.T.; Jacobs, D.R., Jr.; Nettleton, J.A. Dietary intake of saturated fat by food source and incident cardiovascular disease: The Multi-Ethnic Study of Atherosclerosis. Am. J. Clin. Nutr. 2012, 96, 397–404. [Google Scholar] [CrossRef] [Green Version]
- Astrup, A.; Dyerberg, J.; Elwood, P.; Hermansen, K.; Hu, F.B.; Jakobsen, M.U.; Kok, F.J.; Krauss, R.M.; Lecerf, J.M.; LeGrand, P.; et al. The role of reducing intakes of saturated fat in the prevention of cardiovascular disease: Where does the evidence stand in 2010? Am. J. Clin. Nutr. 2011, 93, 684–688. [Google Scholar] [CrossRef] [Green Version]
- US Department of Agriculture and US Department of Health and Human Services. Dietary Guidelines for Americans, 2020–2025, 9th ed.; US Department of Agriculture and US Department of Health and Human Services: Washington, DC, USA, 2020.
- Nordic Council of Ministers. Nordic Nutrition Recommendations 2012: Integrating Nutrition and Physical Activity; Nordic Council of Ministers: Copenhagen, Denmark, 2014; p. 629.
- Sinha, R.; Cross, A.J.; Graubard, B.; Leitzmann, M.F.; Schatzkin, A. Meat intake and mortality: A prospective study of over half a million people. Arch. Intern. Med. 2009, 169, 562–571. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, R.; Dehghan, M.; Mente, A.; Rangarajan, S.; Wielgosz, A.; Avezum, A.; Seron, P.; AlHabib, K.F.; Lopez-Jaramillo, P.; Swaminathan, S.; et al. Associations of unprocessed and processed meat intake with mortality and cardiovascular disease in 21 countries [Prospective Urban Rural Epidemiology (PURE) Study]: A prospective cohort study. Am. J. Clin. Nutr. 2021, 114, 1049–1058. [Google Scholar] [CrossRef]
- Hjerpsted, J.; Tholstrup, T. Cheese and Cardiovascular Disease Risk: A Review of the Evidence and Discussion of Possible Mechanisms. Crit. Rev. Food Sci. Nutr. 2016, 56, 1389–1403. [Google Scholar] [CrossRef] [PubMed]
- Sonestedt, E.; Wirfalt, E.; Wallstrom, P.; Gullberg, B.; Orho-Melander, M.; Hedblab, B. Dairy products and its association with incidence of cardiovascular disease: The Malmo diet and cancer cohort. Eur. J. Epidemiol. 2011, 26, 609–618. [Google Scholar] [CrossRef]
- O’Connor, L.E.; Kim, J.E.; Campbell, W.W. Total red meat intake of >/=0.5 servings/d does not negatively influence cardiovascular disease risk factors: A systemically searched meta-analysis of randomized controlled trials. Am. J. Clin. Nutr. 2017, 105, 57–69. [Google Scholar] [CrossRef] [Green Version]
- Dehghan, M.; Mente, A.; Rangarajan, S.; Sheridan, P.; Mohan, V.; Iqbal, R.; Gupta, R.; Lear, S.; Wentzel-Viljoen, E.; Avezum, A.; et al. Association of dairy intake with cardiovascular disease and mortality in 21 countries from five continents (PURE): A prospective cohort study. Lancet 2018, 392, 2288–2297. [Google Scholar] [CrossRef]
- Hammad, S.; Pu, S.; Jones, P.J. Current Evidence Supporting the Link Between Dietary Fatty Acids and Cardiovascular Disease. Lipids 2016, 51, 507–517. [Google Scholar] [CrossRef]
- Thorning, T.K.; Bertram, H.C.; Bonjour, J.P.; de Groot, L.; Dupon, D.; Feeney, E.; Ipsen, R.; Lecerf, J.M.; Mackie, A.; McKinley, M.C.; et al. Whole dairy matrix or single nutrients in assessment of health effects: Current evidence and knowledge gaps. Am. J. Clin. Nutr. 2017, 105, 1033–1045. [Google Scholar] [CrossRef] [Green Version]
- Hyotylainen, T.; Bondia-Pons, I.; Oresic, M. Lipidomics in nutrition and food research. Mol. Nutr. Food Res. 2013, 57, 1306–1318. [Google Scholar] [CrossRef]
- Havulinna, A.S.; Sysi-Aho, M.; Hilvo, M.; Kauhanen, D.; Hurme, R.; Ekroos, K.; Salomaa, V.; Laaksonen, R. Circulating Ceramides Predict Cardiovascular Outcomes in the Population-Based FINRISK 2002 Cohort. Arterioscler. Thromb. Vasc. Biol. 2016, 36, 2424–2430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seah, J.Y.H.; Chew, W.S.; Torta, F.; Khoo, C.M.; Wenk, M.R.; Herr, D.R.; Tai, E.S.; van Dam, R.M. Dietary Fat and Protein Intake in Relation to Plasma Sphingolipids as Determined by a Large-Scale Lipidomic Analysis. Metabolites 2021, 11, 93. [Google Scholar] [CrossRef]
- Paul, S.; Lancaster, G.I.; Meikle, P.J. Plasmalogens: A potential therapeutic target for neurodegenerative and cardiometabolic disease. Prog. Lipid. Res. 2019, 74, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Lankinen, M.; Schwab, U.; Kolehmainen, M.; Paananen, J.; Nygren, H.; Seppanen-Laakso, T.; Poutanen, K.; Hyötyläinen, T.; Riserus, U. A Healthy Nordic Diet Alters the Plasma Lipidomic Profile in Adults with Features of Metabolic Syndrome in a Multicenter Randomized Dietary Intervention. J. Nutr. 2015, 146, 662–672. [Google Scholar] [CrossRef] [Green Version]
- Wade, A.T.; Davis, C.R.; Dyer, K.A.; Hodgson, J.M.; Woodman, R.J.; Murphy, K.J. Effects of Mediterranean diet supplemented with lean pork on blood pressure and markers of cardiovascular risk: Findings from the MedPork trial. Br. J. Nutr. 2019, 122, 873–883. [Google Scholar] [CrossRef]
- Zheng, J.S.; Sharp, S.J.; Imamura, F.; Koulman, A.; Schulze, M.B.; Ye, Z.; Griffin, J.; Guevara, M.; Huerta, J.M.; Kröger, J.; et al. Association between plasma phospholipid saturated fatty acids and metabolic markers of lipid, hepatic, inflammation and glycaemic pathways in eight European countries: A cross-sectional analysis in the EPIC-InterAct study. BMC Med. 2017, 15, 203. [Google Scholar] [CrossRef] [Green Version]
- Ericson, U.; Hellstrand, S.; Brunkwall, L.; Schulz, C.A.; Sonestedt, E.; Wallstrom, P.; Gullberg, B.; Wirfält, E.; Orho-Melander, M. Food sources of fat may clarify the inconsistent role of dietary fat intake for incidence of type 2 diabetes. Am. J. Clin. Nutr. 2015, 101, 1065–1080. [Google Scholar] [CrossRef] [Green Version]
- Thorning, T.K.; Raziani, F.; Bendsen, N.; Astrup, A.; Tholstrup, T.; Raben, A. Diets with high-fat cheese, high-fat meat, or carbohydrate on cardiovascular risk markers in overweight postmenopausal women: A randomized crossover trial. Am. J. Clin. Nutr. 2015, 102, 573–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergeron, N.; Chiu, S.; Williams, P.T.; King, S.M.; Krauss, R.M. Effects of red meat, white meat, and nonmeat protein sources on atherogenic lipoprotein measures in the context of low compared with high saturated fat intake: A randomized controlled trial. Am. J. Clin. Nutr. 2019, 110, 24–33. [Google Scholar] [CrossRef]
- Chiu, S.; Williams, P.T.; Krauss, R.M. Effects of a very high saturated fat diet on LDL particles in adults with atherogenic dyslipidemia: A randomized controlled trial. PLoS ONE 2017, 12, e0170664. [Google Scholar] [CrossRef] [Green Version]
- Sacks, F.M.; Katan, M. Randomized clinical trials on the effects of dietary fat and carbohydrate on plasma lipoproteins and cardiovascular disease. Am. J. Med. 2002, 113 (Suppl. S9B), 13S–24S. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Xia, M.; Gao, X. Biomarker Discovery in Atherosclerotic Diseases Using Quantitative Nuclear Magnetic Resonance Metabolomics. Front. Cardiovasc. Med. 2021, 8, 681444. [Google Scholar] [CrossRef] [PubMed]
- Lankinen, M.; Kolehmainen, M.; Jaaskelainen, T.; Paananen, J.; Joukamo, L.; Kangas, A.J.; Soininen, P.; Poutanen, K.; Mykkänen, H.; Gylling, H.; et al. Effects of whole grain, fish and bilberries on serum metabolic profile and lipid transfer protein activities: A randomized trial (Sysdimet). PLoS ONE 2014, 9, e90352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erkkila, A.T.; Schwab, U.S.; Lehto, S.; de Mello, V.D.; Kangas, A.J.; Soininen, P.; Ala-Korpela, M.; Uusitupa, M.I.J. Effect of fatty and lean fish intake on lipoprotein subclasses in subjects with coronary heart disease: A controlled trial. J. Clin. Lipidol. 2014, 8, 126–133. [Google Scholar] [CrossRef] [Green Version]
- Manninen, S.M.; Lankinen, M.A.; de Mello, V.D.; Laaksonen, D.E.; Schwab, U.S.; Erkila, A.T. Intake of Fatty Fish Alters the Size and the Concentration of Lipid Components of HDL Particles and Camelina Sativa Oil Decreases IDL Particle Concentration in Subjects with Impaired Glucose Metabolism. Mol. Nutr. Food Res. 2018, 62, e1701042. [Google Scholar] [CrossRef] [Green Version]
- Hilvo, M.; Vasile, V.C.; Donato, L.J.; Hurme, R.; Laaksonen, R. Ceramides and Ceramide Scores: Clinical Applications for Cardiometabolic Risk Stratification. Front. Endocrinol. 2020, 11, 570628. [Google Scholar] [CrossRef]
- Larsen, P.J.; Tennagels, N. On ceramides, other sphingolipids and impaired glucose homeostasis. Mol. Metab. 2014, 3, 252–260. [Google Scholar] [CrossRef]
- Yin, X.; Willinger, C.M.; Keefe, J.; Liu, J.; Fernandez-Ortiz, A.; Ibanez, B.; Peñalvo, J.; Adourian, A.; Chen, G.; Corella, D.; et al. Lipidomic profiling identifies signatures of metabolic risk. EBioMedicine 2020, 51, 102520. [Google Scholar] [CrossRef] [Green Version]
- Murphy, K.J.; Thomson, R.L.; Coates, A.M.; Buckley, J.D.; Rowe, P.R.C. Effects of eating fresh lean pork on cardiometabolic health parameters. Nutrients 2012, 4, 711–723. [Google Scholar] [CrossRef] [Green Version]
- Widmer, R.J.; Flammer, A.J.; Lerman, L.O.; Lerman, A. The Mediterranean diet, its components, and cardiovascular disease. Am. J. Med. 2015, 128, 229–238. [Google Scholar] [CrossRef] [Green Version]
- Vessby, B.; Uusitupa, M.; Hermansen, K.; Riccardi, G.; Rivellese, A.A.; Tapsel, L.C.; Nälsén, C.; Berglund, L.; Louheranta, A.; Rasmussen, B.M.; et al. Substituting dietary saturated for monounsaturated fat impairs insulin sensitivity in healthy men and women: The KANWU Study. Diabetologia 2001, 44, 312–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bos, M.B.; de Vries, J.H.M.; Feskens, E.M.J.; van Dijk, S.J.; Hoelen, D.W.M.; Siebelink, E.; Heijligenberg, R.; de Groot, L.C. Effect of a high monounsaturated fatty acids diet and a Mediterranean diet on serum lipids and insulin sensitivity in adults with mild abdominal obesity. Nutr. Metab. Cardiovasc. Dis. 2010, 20, 591–598. [Google Scholar] [CrossRef]
- Michielsen, C.; Hangelbroek, R.W.J.; Feskens, E.J.M.; Afman, L.A. Disentangling the Effects of Monounsaturated Fatty Acids from Other Components of a Mediterranean Diet on Serum Metabolite Profiles: A Randomized Fully Controlled Dietary Intervention in Healthy Subjects at Risk of the Metabolic Syndrome. Mol. Nutr. Food Res. 2019, 63, e1801095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hulander, E.; Barebring, L.; Wadell, A.T.; Gjertsson, I.; Calder, P.C.; Winkvist, A.; Lindqvist, H.M. Diet intervention improves cardiovascular profile in patients with rheumatoid arthritis: Results from the randomized controlled cross-over trial ADIRA. Nutr. J. 2021, 20, 9. [Google Scholar] [CrossRef] [PubMed]
- Razquin, C.; Toledo, E.; Clish, C.B.; Ruiz-Canela, M.; Dennis, C.; Corella, D.; Papandreou, C.; Ros, E.; Estruch, R.; Guasch-Ferré, M.; et al. Plasma Lipidomic Profiling and Risk of Type 2 Diabetes in the PREDIMED Trial. Diabetes Care 2018, 41, 2617–2624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotronen, A.; Velagapudi, V.R.; Yetukuri, L.; Westerbacka, J.; Bergholm, R.; Ekroos, K.; Makkonen, J.; Taskinen, M.R.; Oresic, M.; Yki-Järvinen, H. Serum saturated fatty acids containing triacylglycerols are better markers of insulin resistance than total serum triacylglycerol concentrations. Diabetologia 2009, 52, 684–690. [Google Scholar] [CrossRef] [Green Version]
- Schwab, U.; Teppanen-Laakso, T.; Yetukuri, L.; Agren, J.; Kolehmainen, M.; Laaksonen, D.E.; Ruskeepää, A.L.; Gylling, H.; Uusitupa, M.; Oresic, M.; et al. Triacylglycerol fatty acid composition in diet-induced weight loss in subjects with abnormal glucose metabolism--the GENOBIN study. PLoS ONE 2008, 3, e2630. [Google Scholar] [CrossRef]
- Djekic, D.; Shi, L.; Calais, F.; Carlsson, F.; Landberg, R.; Hyötyläinen, T.; Frøbert, O. Effects of a Lacto-Ovo-Vegetarian Diet on the Plasma Lipidome and Its Association with Atherosclerotic Burden in Patients with Coronary Artery Disease-A Randomized, Open-Label, Cross-over Study. Nutrients 2020, 12, 3586. [Google Scholar] [CrossRef]
- Stegemann, C.; Pechlaner, R.; Willeit, P.; Langley, S.R.; Mangino, M.; Mayr, U.; Menni, C.; Moayyeri, A.; Santer, P.; Rungger, G.; et al. Lipidomics profiling and risk of cardiovascular disease in the prospective population-based Bruneck study. Circulation 2014, 129, 1821–1831. [Google Scholar] [CrossRef] [Green Version]
- Hu, C.; Kong, H.; Qu, F.; Li, Y.; Yu, Z.; Gao, P.; Peng, S.; Xu, G. Application of plasma lipidomics in studying the response of patients with essential hypertension to antihypertensive drug therapy. Mol. Biosyst. 2011, 7, 3271–3279. [Google Scholar] [CrossRef]
- Sanders, F.; McNally, B.; Griffin, J.L. Blood triacylglycerols: A lipidomic window on diet and disease. Biochem. Soc. Trans. 2016, 44, 638–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meikle, P.J.; Wong, G.; Barlow, C.K.; Kingwell, B.A. Lipidomics: Potential role in risk prediction and therapeutic monitoring for diabetes and cardiovascular disease. Pharmacol. Ther. 2014, 143, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Gowda, S.G.B.; Gao, Z.J.; Chen, Z.; Abe, T.; Hori, S.; Fukiya, S.; Ishizuka, S.; Yokota, A.; Chiba, H.; Hui, S.P. Untargeted Lipidomic Analysis of Plasma from High-fat Diet-induced Obese Rats Using UHPLC-Linear Trap Quadrupole-Orbitrap MS. Anal. Sci. 2020, 36, 821–828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maeba, R.; Maeda, T.; Kinoshita, M.; Takao, K.; Takenaka, H.; Kusano, J.; Yoshimura, N.; Takeoka, Y.; Yasuda, D.; Okazaki, T.; et al. Plasmalogens in human serum positively correlate with high- density lipoprotein and decrease with aging. J. Atheroscler. Thromb. 2007, 14, 12–18. [Google Scholar] [CrossRef] [Green Version]
- An, P.; Wan, S.; Luo, Y.; Luo, J.; Zhang, X.; Zhou, S.; Xu, T.; He, J.; Mechanick, J.I.; Wu, W.C.; et al. Micronutrient Supplementation to Reduce Cardiovascular Risk. J. Am. Coll. Cardiol. 2022, 80, 2269–2285. [Google Scholar] [CrossRef]
- Celada, P.; Sanchez-Muniz, S.J.; Delgado-Pando, G.; Bastida, S.; Rodilla, M.E.; Jimenez-Colmenero, F.; Olmedilla-Alonso, B. Effects of improved fat meat products consumption on emergent cardiovascular disease markers of male volunteers at cardiovascular risk. J. Physiol. Biochem. 2016, 72, 669–678. [Google Scholar] [CrossRef] [Green Version]
- Astrup, A.; Magkos, F.; Bier, D.M.; Brena, J.T.; de Oliveira-Otto, M.C.; Hill, J.O.; King, J.C.; Mente, A.; Ordovas, J.M.; Volek, J.S.; et al. Saturated Fats and Health: A Reassessment and Proposal for Food-Based Recommendations: JACC State-of-the-Art Review. J. Am. Coll. Cardiol. 2020, 76, 844–857. [Google Scholar] [CrossRef] [PubMed]
- Ebbeling, C.B.; Knapp, A.; Johnson, A.; Wong, J.M.; Greco, K.F.; Ma, C.; Mora, S.; Ludwig, D.S. Effects of a low-carbohydrate diet on insulin-resistant dyslipoproteinemia-a randomized controlled feeding trial. Am. J. Clin. Nutr. 2022, 115, 154–162. [Google Scholar] [CrossRef]
- Nieto, F.J.; Iribarren, C.; Gross, M.D.; Comstock, G.W.; Cutler, R.G. Uric acid and serum antioxidant capacity: A reaction to atherosclerosis? Atherosclerosis 2000, 148, 131–139. [Google Scholar] [CrossRef]
- Dattilo, A.M.; Kris-Etherton, P.M. Effects of weight reduction on blood lipids and lipoproteins: A meta-analysis. Am. J. Clin. Nutr. 1992, 56, 320–328. [Google Scholar] [CrossRef] [Green Version]
- Craig, C.L.; Marshall, A.L.; Sjostrom, M.; Bauman, A.E.; Booth, M.L.; Ainsworth, B.E.; Pratt, M.; Ekelund, U.; Yngve, A.; Sallis, J.F.; et al. International physical activity questionnaire: 12-country reliability and validity. Med. Sci. Sports Exerc. 2003, 35, 1381–1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campos-Giménez, E.; Fontannaz, P.; Trisconi, M.J.; Kilinc, T.; Gimenez, C.; Andrieux, P. Determination of vitamin B12 in food products by liquid chromatography/UV detection with immunoaffinity extraction: Single-laboratory validation. J. AOAC Int. 2008, 91, 786–793. [Google Scholar] [CrossRef] [Green Version]
- Jørgensen, C.E.; Abrahamsen, R.K.; Rukke, E.O.; Johansen, A.G.; Schüller, R.B.; Skeie, S.B. Improving the structure and rheology of high protein, low fat yoghurt with undenatured whey proteins. Int. Dairy, J. 2015, 47, 6–18. [Google Scholar] [CrossRef]
- Norwegian Institute of Public Health. HUBRO-Protocol, Protocol for The Oslo Health Study; Norwegian Institute of Public Health: Oslo, Norway, 2002.
- Barr, J.; Caballería, J.; Martínez-Arranz, I.; Domínguez-Díez, A.; Alonso, C.; Muntané, J.; Pérez-Cormenzana, M.; Gar-cía-Monzón, C.; Mayo, R.; Martín-Duce, A.; et al. Obesity-dependent metabolic signatures associated with nonalcoholic fatty liver disease progression. J. Proteome Res. 2012, 11, 2521–2532. [Google Scholar] [CrossRef]
- Martínez-Arranz, I.; Mayo, R.; Pérez-Cormenzana, M.; Mincholé, I.; Salazar, L.; Alonso, C.; Mato, J.M. Enhancing metabolomics research through data mining. J. Proteom. 2015, 127, 275–288. [Google Scholar] [CrossRef]
- Mallol, R.; Amigó, N.; Rodríguez, M.A.; Heras, M.; Vinaixa, M.; Plana, N. Liposcale: A novel advanced lipoprotein test based on 2D diffusion-ordered 1H NMR spectroscopy. J. Lipid Res. 2015, 56, 737–746. [Google Scholar] [CrossRef] [Green Version]
- McKinney, W. Pandas: A python Data Analysis Library. Available online: http://pandas.pydata.org (accessed on 20 November 2022).
- Virtanen, P.; Gommers, R.; Oliphant, T.E.; Haberland, M.; Reddy, T.; Cournapeau, D.; Burovski, E.; Peterson, P.; Weckesser, W.; Bright, J.; et al. SciPy 1.0 Contributors. SciPy 1.0: Fundamental algorithms for scientific computing in Python. Nat. Methods 2020, 17, 261–272. [Google Scholar] [CrossRef] [Green Version]
- Vallat, R. Pingouin: Statistics in Python. J. Open Source Softw. 2018, 3, 1026. [Google Scholar] [CrossRef]
- Waskom, M.; Botvinnik, O.; Ostblom, J.; Gelbart, M.; Lukauskas, S.; Hobson, P.; Gemperline, D.C.; Augspurger, T.; Halchenko, Y.; Cole, J.B.; et al. Mwaskom/Seaborn: v0.10.1. 2020. Available online: https://zenodo.org/record/3767070#.ZAHkHHbMKUk (accessed on 20 November 2022).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Men | Women | p-Value | Total Sample | |
|---|---|---|---|---|
| Clinical Data | ||||
| Number (n; %) | 10 (30%) | 23 (70%) | . | 33 |
| Age (years) | 24.6 ± 2.4 | 23.1 ± 2.7 | 0.15 | 23.6 ± 2.7 |
| Height (m) | 1.8 ± 0.1 | 1.7 ± 0.1 | <0.01 | 1.7 ± 0.1 |
| Weight (kg) | 87.7 ± 14.5 | 68.1 ± 10.8 | <0.01 | 74.0 ± 14.9 |
| BMI (kg/m2) | 27.0 ± 3.7 | 24.3 ± 3.1 | 0.04 | 25.1 ± 3.5 |
| Waist circumference (cm) | 87.7 ± 8.1 | 76.9 ± 7.1 | <0.01 | 79.8 ± 8.7 |
| Systolic blood pressure (mmHg) | 132.4 ± 8.5 | 119.7 ± 10.4 | <0.01 | 123.6 ± 11.4 |
| Diastolic Blood pressure (mmHg) & | 79.7 ± 9.5 | 79.1 ± 8.5 | 0.74 | 79.3 ± 8.7 |
| Glucose (mmol/L) | 4.9 ± 0.3 | 4.6 ± 0.3 | 0.04 | 4.7 ± 0.3 |
| Insulin (µUI/mL) | 7.1 ± 3.2 | 8.3 ± 4.1 | 0.41 | 7.9 ± 3.9 |
| HOMA-IR | 1.5 ± 0.7 | 1.7 ± 0.9 | 0.57 | 1.7 ± 0.9 |
| C-peptide (pmol/L) | 333.6 ± 92.1 | 356.3 ± 87.5 | 0.51 | 349.4 ± 88.1 |
| AST (U/L) & | 22.3 ± 9.2 | 16.5 ± 6.9 | 0.01 | 18.2 ± 8.0 |
| ALT (U/L) & | 28.1 ± 11.3 | 18.9 ± 9.3 | <0.01 | 21.7 ± 10.7 |
| Total cholesterol (mmol/L) | 4.7 ± 1.1 | 4.6 ± 0.9 | 0.89 | 4.6 ± 0.9 |
| LDL cholesterol (mmol/L) | 3.0 ± 1.1 | 2.8 ± 0.8 | 0.64 | 2.9 ± 0.9 |
| HDL cholesterol (mmol/L) | 1.4 ± 0.3 | 1.6 ± 0.3 | 0.21 | 1.5 ± 0.3 |
| Triglycerides (mmol/L) & | 1.0 ± 0.6 | 1.0 ± 0.3 | 0.71 | 1.0 ± 0.4 |
| Apolipoprotein A (g/L) | 1.3 ± 0.2 | 1.4 ± 0.2 | 0.25 | 1.3 ± 0.2 |
| Apolipoprotein B (g/L) | 0.8 ± 0.2 | 0.8 ± 0.2 | 0.77 | 0.8 ± 0.2 |
| ApoB/A ratio & | 0.6 ± 0.2 | 0.6 ± 0.1 | 0.38 | 0.6 ± 0.1 |
| C-reactive protein (mg/L) & | 1.2 ± 1.9 | 1.1 ± 1.2 | 0.55 | 1.1 ± 1.4 |
| Uric acid | 391.9 ± 70.3 | 277.5 ± 40.9 | <0.01 | 312.2 ± 73.4 |
| Calcium (mmol/L) | 2.4 ± 0.1 | 2.3 ± 0.1 | 0.01 | 2.4 ±0.1 |
| Ferritin (µg/L) & | 144.5 ± 64 | 50.9 ± 28.1 | <0.01 | 79.2 ± 60.0 |
| Iron (µmol/L) | 21.2 ± 6 | 23.6 ± 10.1 | 0.42 | 22.8 ± 9.0 |
| Vitamin D (nmol/L) | 48.2 ± 18.3 | 51.0 ± 19.0 | 0.70 | 50.1 ± 18.5 |
| Vitamin B12 (pmol/L) & | 344.2 ± 140.1 | 275.6 ± 110.6 | 0.14 | 296.4 ± 122.3 |
| Women F2 (2300 kcal, n = 15) | Men M2 (3000 kcal, n = 7) | |||||||
|---|---|---|---|---|---|---|---|---|
| Gouda Cheese | Goutaler Cheese | Pork | Beef | Gouda Cheese | Goutaler Cheese | Pork | Beef | |
| Energy (kcal) | 2289 | 2289 | 2254 | 2230 | 3029 | 3029 | 2994 | 2970 |
| Carbohydrates (gram) | 283.2 | 283.2 | 283.2 | 283.2 | 382.9 | 382.9 | 382.9 | 382.9 |
| Carbohydrates (% of TEI) | 46.0 | 46.0 | 47.5 | 47.5 | 47.0 | 47.0 | 47.0 | 47.0 |
| Protein (gram) | 93.2 | 94.4 | 89.3 | 92.9 | 117.1 | 118.1 | 113.0 | 116.8 |
| Protein (% of TEI) | 16.0 | 16.0 | 16.0 | 16.0 | 15.0 | 15.5 | 15.5 | 16 |
| Fat (gram) | 96.5 | 96.2 | 94.2 | 90.2 | 128.1 | 127.5 | 125.8 | 121.9 |
| Fat (% of TEI) | 38.0 | 38.0 | 37.5 | 36.5 | 38.0 | 37.5 | 37.5 | 37.0 |
| Saturated fatty acids (gram) | 35.8 | 35.7 | 22.3 | 25.4 | 40.7 | 40.5 | 27.2 | 30.3 |
| Saturated fatty acids (% TEI) | 14.0 | 14.0 | 9.0 | 10.0 | 12.0 | 12.0 | 8.0 | 9.0 |
| Trans FA (g) | 0.5 | 0.5 | 0 | 0.7 | 0.5 | 0.5 | 0 | 0.7 |
| Trans FA (% TEI) | 0.2 | 0.2 | 0 | 0.3 | 0.1 | 0.1 | 0 | 0.2 |
| MUFA (gram) | 33.4 | 33.1 | 44.6 | 40.0 | 48.9 | 48.6 | 60.2 | 55.5 |
| MUFA (%TEI) | 13.0 | 13.0 | 18.0 | 16.0 | 15.0 | 14.0 | 18.0 | 17.0 |
| PUFA (gram) | 18.0 | 18.0 | 24.1 | 18.0 | 26.6 | 26.6 | 32.8 | 26.8 |
| PUFA (%TEI) | 7.0 | 7.0 | 10.0 | 7.0 | 8.0 | 8.0 | 10.0 | 8.0 |
| Omega-3 (gram) & | 3.3 | 3.3 | 3.5 | 3.3 | 5.1 | 5.1 | 5.3 | 5.2 |
| Omega-6 (gram) & | 13.1 | 13.1 | 13.1 | 13.2 | 19.6 | 19.6 | 19.6 | 19.6 |
| Cholesterol (mg) & | 135 | 135 | 128 | 165 | 162 | 162 | 155 | 192 |
| Starch (gram) & | 156.2 | 156.2 | 156.2 | 156.2 | 212.8 | 212.8 | 212.8 | 212.8 |
| Mono- and dissaccharides (gram) & | 91.9 | 91.9 | 91.9 | 91.9 | 117 | 117 | 117 | 117 |
| Sugar, added (gram) & | 24.1 | 24.1 | 24.1 | 24.1 | 24.7 | 24.7 | 24.7 | 24.7 |
| Fiber (gram) & | 42 | 42 | 42 | 42 | 60 | 60 | 60 | 60 |
| Salt (gram) & | 10.6 | 10.5 | 9.1 | 9.3 | 11.5 | 11.4 | 9.9 | 10.2 |
| Alcohol (gram)& | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Beta-carotene (µg) & | 8736 | 8729 | 8490 | 8619 | 9039 | 9032 | 8792 | 8922 |
| Calcium (mg) | 1312 | 1443 | 329 | 334 | 1429 | 1560 | 446 | 451 |
| Copper (mg) & | 1.9 | 1.9 | 2 | 1.9 | 2.8 | 2.8 | 2.8 | 2.7 |
| Folate (µg) & | 361 | 352 | 301 | 305 | 509 | 500 | 446 | 453 |
| Iodine (µg) € | 198 | 189 | 166 | 168 | 212 | 203 | 179 | 182 |
| Iron (mg) | 13 | 13 | 14.1 | 17.4 | 18.3 | 18.3 | 19.3 | 22.7 |
| Magnesium (mg) | 453 | 459 | 456 | 454 | 614 | 620 | 618 | 615 |
| Niacin (mg) & | 20.2 | 20.9 | 31.1 | 28 | 29.8 | 30.5 | 40.4 | 37.6 |
| Phosphorus (mg) & | 2078 | 2134 | 1731 | 1696 | 2759 | 2815 | 2413 | 2377 |
| Potassium (mg) | 3751 | 3742 | 4283 | 4205 | 5495 | 5486 | 6028 | 5949 |
| Retinol (µg) & | 595 | 596 | 243 | 267 | 693 | 694 | 340 | 365 |
| Riboflavin (mg) & | 1.1 | 1.1 | 0.9 | 0.9 | 1.4 | 1.4 | 1.1 | 1.2 |
| Selenium (µg) | 63 | 63 | 80 | 60 | 86 | 86 | 103 | 83 |
| Sodium (mg) | 3271 | 3385 | 2880 | 2872 | 3567 | 3681 | 3176 | 3168 |
| Thiamin (mg) & | 1.7 | 1.8 | 2.7 | 1.7 | 2.3 | 2.4 | 3.3 | 2.3 |
| Vitamin A (RAE) & | 1324 | 1324 | 952 | 984 | 1447 | 1447 | 1075 | 1107 |
| Vitamin B12 (µg) | 3.1 | 3 | 2.1 | 4.6 | 4.2 | 4.1 | 3.2 | 5.7 |
| Vitamin B6 (mg) & | 1.6 | 1.6 | 2.5 | 2.2 | 2.5 | 2. 5 | 3.4 | 3.0 |
| Vitamin C (mg) & | 159 | 159 | 159 | 159 | 191 | 191 | 191 | 191 |
| Vitamin D (µg) &,£ | 20.9 | 20.9 | 21.3 | 21.4 | 25.2 | 25.2 | 25.7 | 25.7 |
| Vitamin E (alfa-TE) & | 18 | 18.3 | 18.7 | 18.1 | 25.7 | 26 | 26.3 | 25.8 |
| Zinc (mg) & | 16.2 | 15.6 | 12.8 | 18.5 | 20 | 19.4 | 16.6 | 22.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monfort-Pires, M.; Lamichhane, S.; Alonso, C.; Egelandsdal, B.; Orešič, M.; Jordahl, V.O.; Skjølsvold, O.; Pérez-Ruiz, I.; Blanco, M.E.; Skeie, S.; et al. Classification of Common Food Lipid Sources Regarding Healthiness Using Advanced Lipidomics: A Four-Arm Crossover Study. Int. J. Mol. Sci. 2023, 24, 4941. https://doi.org/10.3390/ijms24054941
Monfort-Pires M, Lamichhane S, Alonso C, Egelandsdal B, Orešič M, Jordahl VO, Skjølsvold O, Pérez-Ruiz I, Blanco ME, Skeie S, et al. Classification of Common Food Lipid Sources Regarding Healthiness Using Advanced Lipidomics: A Four-Arm Crossover Study. International Journal of Molecular Sciences. 2023; 24(5):4941. https://doi.org/10.3390/ijms24054941
Chicago/Turabian StyleMonfort-Pires, Milena, Santosh Lamichhane, Cristina Alonso, Bjørg Egelandsdal, Matej Orešič, Vilde Overrein Jordahl, Oda Skjølsvold, Irantzu Pérez-Ruiz, María Encarnación Blanco, Siv Skeie, and et al. 2023. "Classification of Common Food Lipid Sources Regarding Healthiness Using Advanced Lipidomics: A Four-Arm Crossover Study" International Journal of Molecular Sciences 24, no. 5: 4941. https://doi.org/10.3390/ijms24054941