

Molecular Dynamics Assessment of Mechanical Properties of the Thin Filaments in Cardiac Muscle


Abstract
:1. Introduction
2. Results
2.1. Structural Model of the Thin Filament in Ca-Free State, Actin Helix Parameters
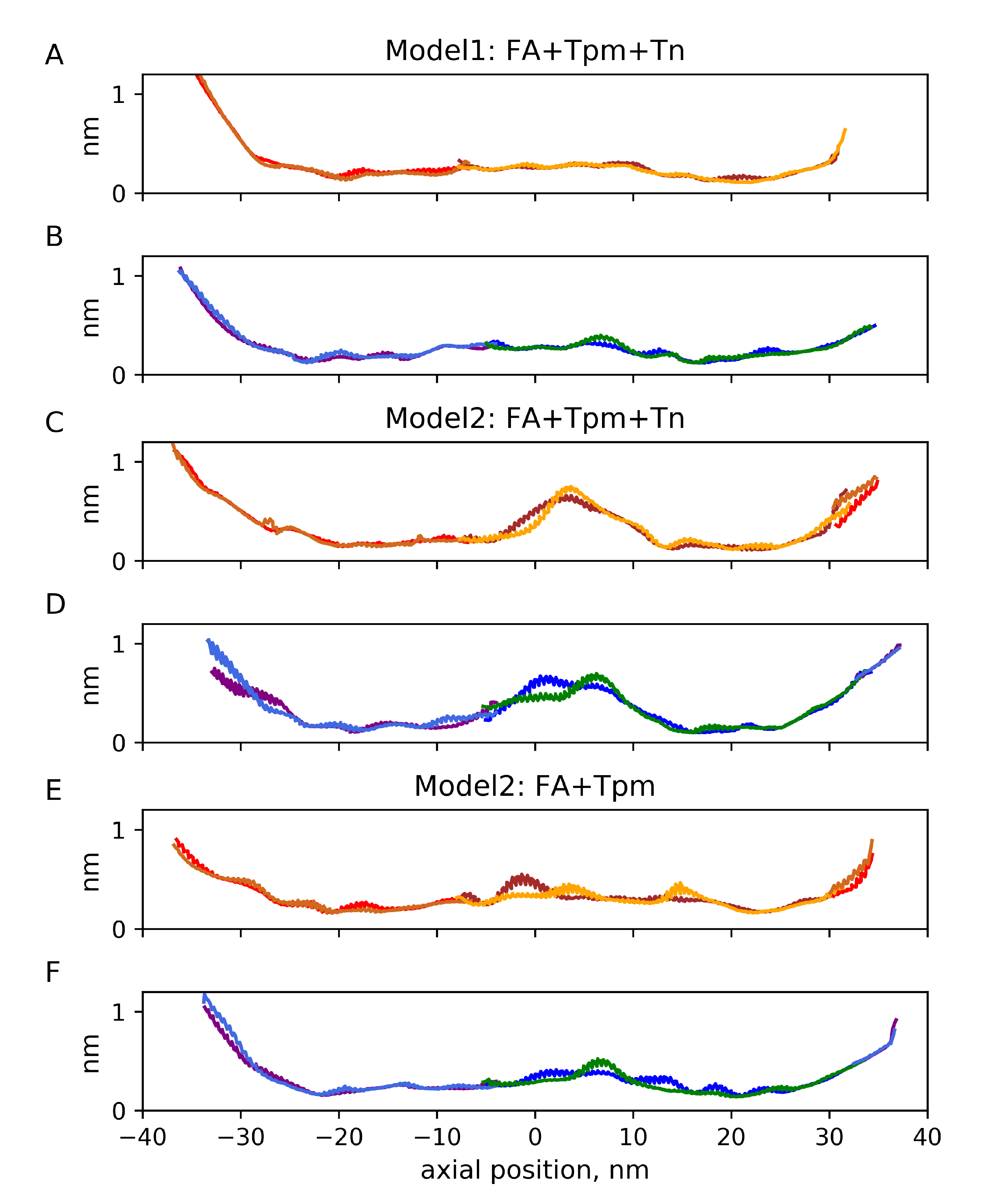
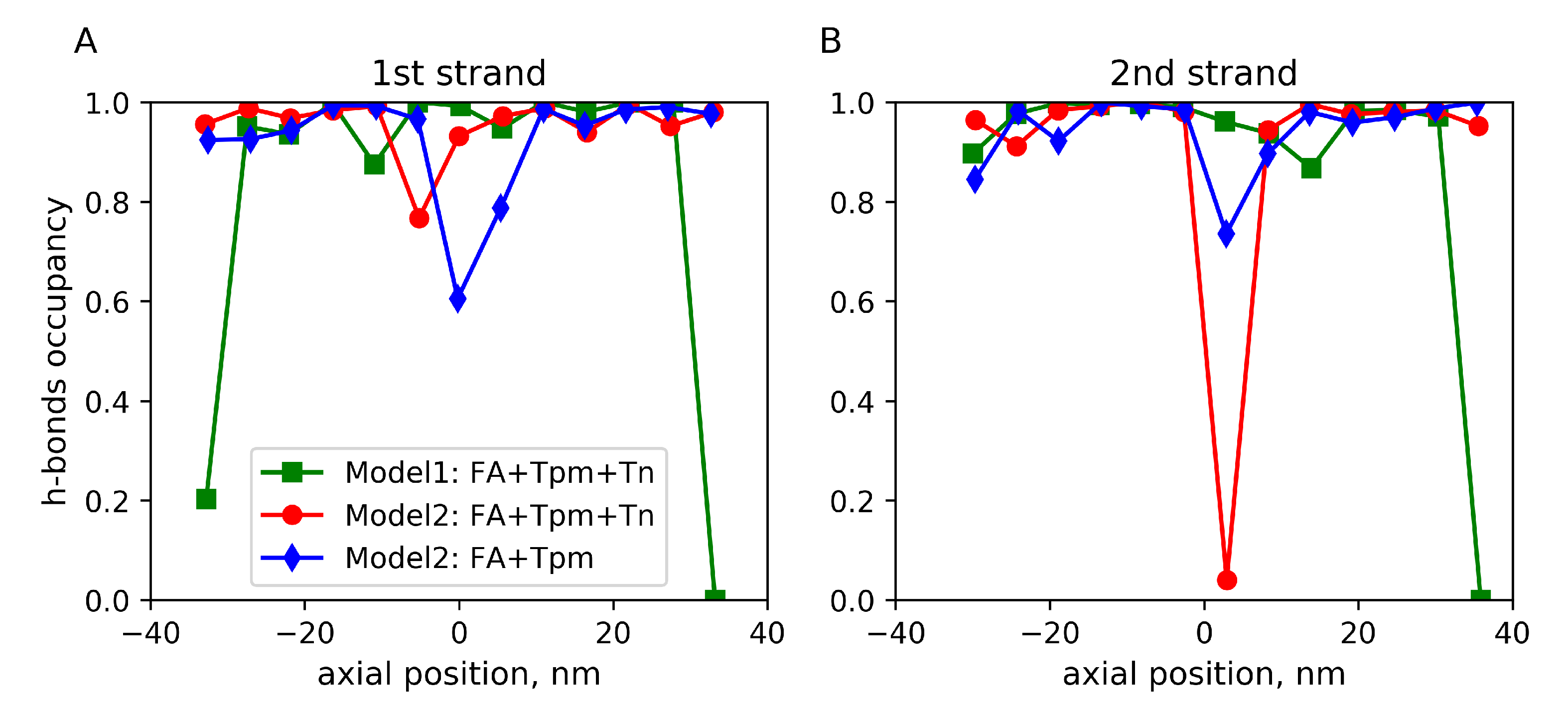
2.2. Mechanical Properties of the Filaments
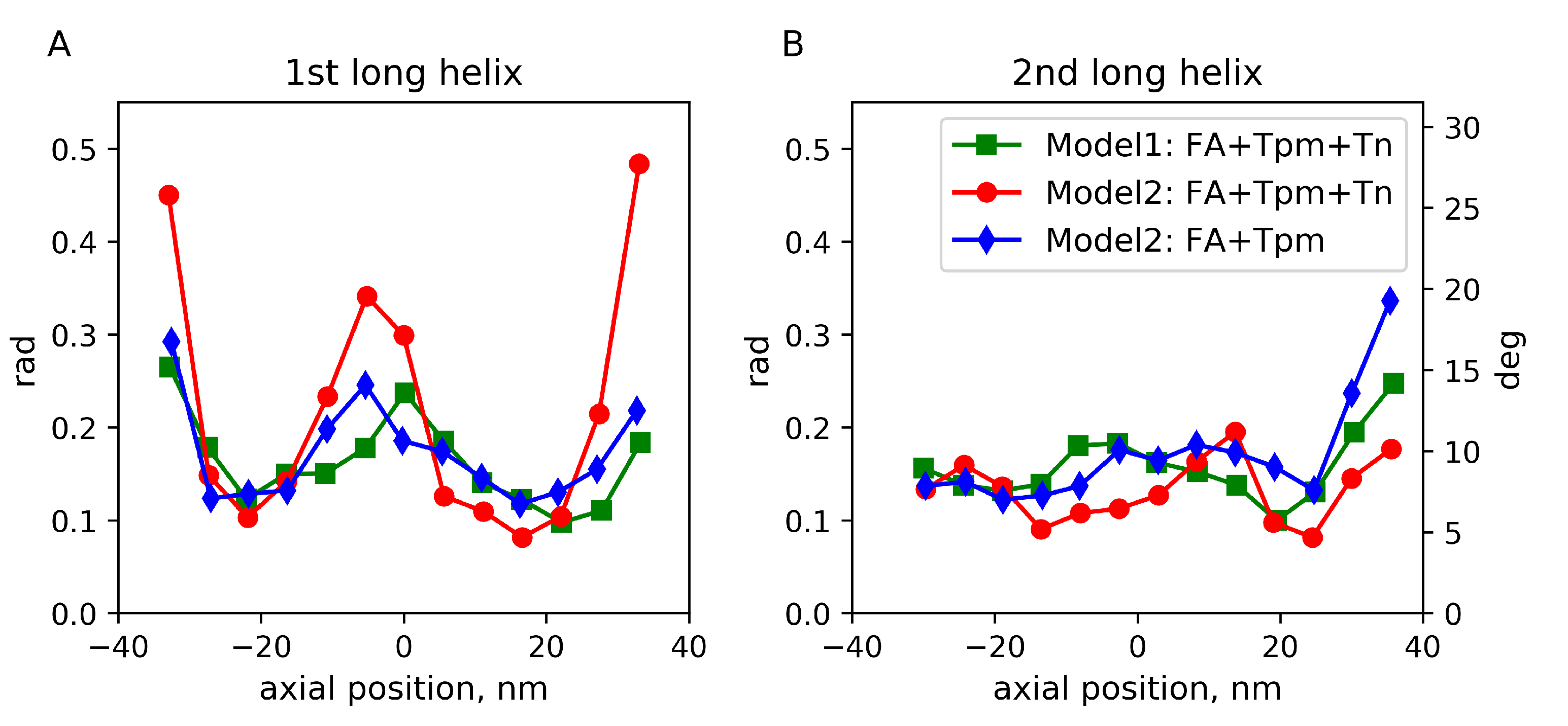
2.3. Fluctuations of the Angular and Radial Position of Tpm on F-Actin
3. Discussion
4. Materials and Methods
4.1. MD Simulation
4.2. Analysis of MD Trajectories
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| F-actin | Filamentous actin |
| Tn | Troponin |
| Tpm | Tropomyosin |
| M | Mean |
| SD | Standard deviation |
References
- Nevzorov, I.A.; Levitsky, D.I. Tropomyosin: Double helix from the protein world. Biochem. Biokhimiia 2011, 76, 1507–1527. [Google Scholar] [CrossRef] [PubMed]
- Lehman, W.; Rynkiewicz, M.J.; Moore, J.R. A new twist on tropomyosin binding to actin filaments: Perspectives on thin filament function, assembly and biomechanics. J. Muscle Res. Cell Motil. 2020, 41, 23–38. [Google Scholar] [CrossRef]
- Tobacman, L.S. Troponin revealed: Uncovering the structure of the thin filament on-off switch in striated muscle. Biophys. J. 2021, 120, 1–9. [Google Scholar] [CrossRef]
- Haselgrove, J.C. X-ray evidence for a conformational change in actin-containing filaments of vertebrate striated muscle. Cold Spring Harb. Symp. Quant. Biol. 1972, 37, 341–352. [Google Scholar] [CrossRef] [Green Version]
- Huxley, H.E. Structural changes in actin- and myosin-containing filaments during contraction. Cold Spring Harb. Symp. Quant. Biol. 1972, 37, 361–376. [Google Scholar] [CrossRef]
- Spudich, J.A.; Huxley, H.E.; Finch, J.T. Regulation of skeletal muscle contraction. II. Structural studies of the interaction of the tropomyosin-troponin complex with actin. J. Mol. Biol. 1972, 72, 619–632. [Google Scholar] [CrossRef] [PubMed]
- Parry, D.A.; Squire, J.M. Structural role of tropomyosin in muscle regulation: Analysis of the X-ray diffraction patterns from relaxed and contracting muscles. J. Mol. Biol. 1973, 75, 33–55. [Google Scholar] [CrossRef]
- Gordon, A.M.; Homsher, E.; Regnier, M. Regulation of contraction in striated muscle. Physiol. Rev. 2000, 80, 853–924. [Google Scholar] [CrossRef]
- McKillop, D.F.; Geeves, M.A. Regulation of the interaction between actin and myosin subfragment 1: Evidence for three states of the thin filament. Biophys. J. 1993, 65, 693–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehman, W. Thin filament structure and the steric blocking model. Compr. Physiol. 2016, 6, 1043–1069. [Google Scholar] [CrossRef]
- Parry, D.A.D. 50 Years of the steric-blocking mechanism in vertebrate skeletal muscle: A retrospective. J. Muscle Res. Cell Motil. 2022. [Google Scholar] [CrossRef] [PubMed]
- Holmes, K.; Popp, D.; Gebhard, W.; Kabsch, W. Atomic model of the actin filament. Nature 1990, 347, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Poole, K.J.V.; Lorenz, M.; Evans, G.; Rosenbaum, G.; Pirani, A.; Craig, R.; Tobacman, L.S.; Lehman, W.; Holmes, K.C. A comparison of muscle thin filament models obtained from electron microscopy reconstructions and low-angle X-ray fibre diagrams from non-overlap muscle. J. Struct. Biol. 2006, 155, 273–284. [Google Scholar] [CrossRef]
- Huxley, H.; Stewart, A.; Sosa, H.; Irving, T. X-ray diffraction measurements of the extensibility of actin and myosin filaments in contracting muscle. Biophys. J. 1994, 67, 2411–2421. [Google Scholar] [CrossRef] [Green Version]
- Wakabayashi, K.; Sugimoto, Y.; Tanaka, H.; Ueno, Y.; Takezawa, Y.; Amemiya, Y. X-ray diffraction measurements for the extensibility of actin and myosin filaments during muscle contraction. Biophys. J. 1994, 67, 2422–2435. [Google Scholar] [CrossRef] [Green Version]
- Tsaturyan, A.K.; Koubassova, N.; Ferenczi, M.A.; Narayanan, T.; Roessle, M.; Bershitsky, S.Y. Strong binding of myosin heads stretches and twists the actin helix. Biophys. J. 2005, 88, 1902–1910. [Google Scholar] [CrossRef] [Green Version]
- Bordas, J.; Svensson, A.; Rothery, M.; Lowy, J.; Diakun, G.; Boesecke, P. Extensibility and symmetry of actin filaments in contracting muscles. Biophys. J. 1999, 77, 3197–3207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinogradova, M.; Stone, D.; Malanina, G.; Karatzaferi, C.; Cooke, R. Ca2+-regulated structural changes in troponin. Proc. Natl. Acad. Sci. USA 2005, 102, 5038–5043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doran, M.H.; Pavadai, E.; Rynkiewicz, M.J.; Walklate, J.; Bullitt, E.; Moore, J.R.; Regnier, M.; Geeves, M.A.; Lehman, W. Cryo-EM and molecular docking shows myosin loop 4 contacts actin and tropomyosin on thin filaments. Biophys. J. 2020, 119, 821–830. [Google Scholar] [CrossRef]
- Yamada, Y.; Namba, K.; Fujii, T. Cardiac muscle thin filament structures reveal calcium regulatory mechanism. Nat. Commun. 2020, 11, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Risi, C.; Schäfer, L.U.; Belknap, B.; Pepper, I.; White, H.D.; Schröder, G.F.; Galkin, V.E. High-resolution cryo-EM structure of the cardiac actomyosin complex. Structure 2021, 29, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Risi, C.M.; Pepper, I.; Belknap, B.; Landim-Vieira, M.; White, H.D.; Dryden, K.; Pinto, J.R.; Chase, P.B.; Galkin, V.E. The structure of the native cardiac thin filament at systolic Ca2+ levels. Proc. Natl. Acad. Sci. USA 2021, 118, e2024288118. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Grange, M.; Wagner, T.; Kho, A.L.; Gautel, M.; Raunser, S. The molecular basis for sarcomere organization in vertebrate skeletal muscle. Cell 2021, 184, 2135–2150. [Google Scholar] [CrossRef]
- Wang, Z.; Grange, M.; Pospich, S.; Wagner, T.; Kho, A.L.; Gautel, M.; Raunser, S. Structures from intact myofibrils reveal mechanism of thin filament regulation through nebulin. Science 2022, 375, eabn1934. [Google Scholar] [CrossRef] [PubMed]
- Pavadai, E.; Rynkiewicz, M.J.; Ghosh, A.; Lehman, W. Docking Troponin T onto the tropomyosin overlapping domain of thin filaments. Biophys. J. 2020, 118, 325–336. [Google Scholar] [CrossRef]
- Rynkiewicz, M.J.; Pavadai, E.; Lehman, W. Modeling human cardiac thin filament structures. Front. Physiol. 2022, 13, 932333. [Google Scholar] [CrossRef]
- Deranek, A.E.; Baldo, A.P.; Lynn, M.L.; Schwartz, S.D.; Tardiff, J.C. Structure and dynamics of the flexible cardiac troponin T linker domain in a fully reconstituted thin filament. Biochemistry 2022, 61, 1229–1242. [Google Scholar] [CrossRef]
- Tobacman, L.S.; Cammarato, A. Cardiomyopathic troponin mutations predominantly occur at its interface with actin and tropomyosin. J. Gen. Physiol. 2021, 153, e202012815. [Google Scholar] [CrossRef]
- Mason, A.B.; Lynn, M.L.; Baldo, A.P.; Deranek, A.E.; Tardiff, J.C.; Schwartz, S.D. Computational and biophysical determination of pathogenicity of variants of unknown significance in cardiac thin filament. JCI Insight 2021, 6, e154350. [Google Scholar] [CrossRef]
- Chalovich, J.M.; Zhu, L.; Johnson, D. Hypertrophic Cardiomyopathy Mutations of Troponin Reveal Details of Striated Muscle Regulation. Front. Physiol. 2022, 13, 902079. [Google Scholar] [CrossRef]
- Gerull, B.; Klaassen, S.; Brodehl, A. The Genetic Landscape of Cardiomyopathies. In Genetic Causes of Cardiac Disease; Springer International Publishing: Cham, Switzerlnad, 2019; pp. 45–91. [Google Scholar] [CrossRef]
- Matsushita, S.; Adachi, T.; Inoue, Y.; Hojo, M.; Sokabe, M. Evaluation of extensional and torsional stiffness of single actin filaments by molecular dynamics analysis. J. Biomech. 2010, 43, 3162–3167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaendtner, J.; Branduardi, D.; Parrinello, M.; Pollard, T.D.; Voth, G.A. Nucleotide-dependent conformational states of actin. Proc. Natl. Acad. Sci. USA 2009, 106, 12723–12728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaendtner, J.; Lyman, E.; Pollard, T.D.; Voth, G.A. Structure and dynamics of the actin filament. J. Mol. Biol. 2010, 396, 252–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deriu, M.A.; Bidone, T.C.; Mastrangelo, F.; Di Benedetto, G.; Soncini, M.; Montevecchi, F.M.; Morbiducci, U. Biomechanics of actin filaments: A computational multi-level study. J. Biomech. 2011, 44, 630–636. [Google Scholar] [CrossRef] [Green Version]
- Fan, J.; Saunders, M.G.; Voth, G.A. Coarse-graining provides insights on the essential nature of heterogeneity in actin filaments. Biophys. J. 2012, 103, 1334–1342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koubassova, N.A.; Tsaturyan, A.K. Molecular dynamics assessment of the mechanical properties of fibrillar actin. Comput. Res. Model. 2022, 14, 1081–1092. [Google Scholar] [CrossRef]
- Manning, E.P.; Tardiff, J.C.; Schwartz, S.D. A model of calcium activation of the cardiac thin filament. Biochemistry 2011, 50, 7405–7413. [Google Scholar] [CrossRef] [Green Version]
- Zheng, W.; Barua, B.; Hitchcock-DeGregori, S.E. Probing the flexibility of tropomyosin and its binding to filamentous actin using molecular dynamics simulations. Biophys. J. 2013, 105, 1882–1892. [Google Scholar] [CrossRef] [Green Version]
- Zheng, W.; Hitchcock-DeGregori, S.E.; Barua, B. Investigating the effects of tropomyosin mutations on its flexibility and interactions with filamentous actin using molecular dynamics simulation. J. Muscle Res. Cell Motil. 2016, 37, 131–147. [Google Scholar] [CrossRef]
- Viswanathan, M.C.; Schmidt, W.; Franz, P.; Rynkiewicz, M.J.; Newhard, C.S.; Madan, A.; Lehman, W.; Swank, D.M.; Preller, M.; Cammarato, A. A role for actin flexibility in thin filament-mediated contractile regulation and myopathy. Nat. Commun. 2020, 11, 2417. [Google Scholar] [CrossRef]
- Pavadai, E.; Lehman, W.; Rynkiewicz, M.J. Protein-protein docking reveals dynamic interactions of tropomyosin on actin filaments. Biophys. J. 2020, 119, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Kuruba, B.; Kaczmarek, M.; Kęsik-Brodacka, M.; Fojutowska, M.; Śliwinska, M.; Kostyukova, A.S.; Moraczewska, J. Structural effects of disease-related mutations in actin-binding period 3 of tropomyosin. Molecules 2021, 26, 6980. [Google Scholar] [CrossRef]
- Pavadai, E.; Rynkiewicz, M.J.; Yang, Z.; Gould, I.R.; Marston, S.B.; Lehman, W. Modulation of cardiac thin filament structure by phosphorylated troponin-I analyzed by protein-protein docking and molecular dynamics simulation. Arch. Biochem. Biophys. 2022, 725, 109282. [Google Scholar] [CrossRef] [PubMed]
- Kiani, F.A.; Lehman, W.; Fischer, S.; Rynkiewicz, M.J. Spontaneous transitions of actin-bound tropomyosin toward blocked and closed states. J. Gen. Physiol. 2019, 151, 4–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsaturyan, A.K.; Zaklyazminskaya, E.V.; Polyak, M.E.; Kopylova, G.V.; Shchepkin, D.V.; Kochurova, A.M.; Gonchar, A.D.; Kleymenov, S.Y.; Koubasova, N.A.; Bershitsky, S.Y.; et al. De novo Asp219Val mutation in cardiac tropomyosin associated with hypertrophic cardiomyopathy. Int. J. Mol. Sci. 2022, 24, 18. [Google Scholar] [CrossRef] [PubMed]
- Whitby, F.G.; Phillips, G.N., Jr. Crystal structure of tropomyosin at 7 Angstroms resolution. Proteins 2000, 38, 49–59. [Google Scholar] [CrossRef]
- Li, Y.; Mui, S.; Brown, J.H.; Strand, J.; Reshetnikova, L.; Tobacman, L.S.; Cohen, C. The crystal structure of the C-terminal fragment of striated-muscle alpha-tropomyosin reveals a key troponin T recognition site. Proc. Natl. Acad. Sci. USA 2002, 99, 7378–7383. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.H.; Zhou, Z.; Reshetnikova, L.; Robinson, H.; Yammani, R.D.; Tobacman, L.S.; Cohen, C. Structure of the mid-region of tropomyosin: Bending and binding sites for actin. Proc. Natl. Acad. Sci. USA 2005, 102, 18878–18883. [Google Scholar] [CrossRef] [Green Version]
- Bershitsky, S.Y.; Ferenczi, M.A.; Koubassova, N.A.; Tsaturyan, A.K. Insight into the actin-myosin motor from X-ray diffraction on muscle. Front. Biosci. 2009, 14, 3188–3213. [Google Scholar] [CrossRef] [Green Version]
- Lehman, W.; Pavadai, E.; Rynkiewicz, M.J. C-terminal troponin-I residues trap tropomyosin in the muscle thin filament blocked-state. Biochem. Biophys. Res. Commun. 2021, 551, 27–32. [Google Scholar] [CrossRef]
- Strelkov, S.V.; Burkhard, P. Analysis of alpha-helical coiled coils with the program TWISTER reveals a structural mechanism for stutter compensation. J. Struct. Biol. 2002, 137, 54–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barua, B. Periodicities designed in the tropomyosin sequence and structure define its functions. Bioarchitecture 2013, 3, 51–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmes, K.C.; Lehman, W. Gestalt-binding of tropomyosin to actin filaments. J. Muscle Res. Cell Motil. 2008, 29, 213–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koubassova, N.A.; Bershitsky, S.Y.; Ferenczi, M.A.; Narayanan, T.; Tsaturyan, A.K. Tropomyosin movement is described by a quantitative high-resolution model of X-ray diffraction of contracting muscle. Eur. Biophys. J. 2017, 46, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Isambert, H.; Venier, P.; Maggs, A.C.; Fattoum, A.; Kassab, R.; Pantaloni, D.; Carlier, M.F. Flexibility of actin filaments derived from thermal fluctuations. Effect of bound nucleotide, phalloidin, and muscle regulatory proteins. J. Biol. Chem. 1995, 270, 11437–11444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1–2, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Webb, B.; Sali, A. Comparative Protein Structure Modeling Using MODELLER. Curr. Protoc. Bioinform. 2016, 54, 5.6.1–5.6.37. [Google Scholar] [CrossRef] [Green Version]
- Best, R.B.; Zhu, X.; Shim, J.; Lopes, P.E.M.; Mittal, J.; Feig, M.; Mackerell, A.D. Optimization of the additive CHARMM all-atom protein force field targeting improved sampling of the backbone ϕ, ψ and side-chain χ1 and χ2 dihedral angles. J. Chem. Theory Comput. 2012, 8, 3257–3273. [Google Scholar] [CrossRef] [Green Version]
- Berendsen, H.J.C.; Postma, J.P.M.; van Gunsteren, W.F.; DiNola, A.; Haak, J.R. Molecular dynamics with coupling to an external bath. J. Chem. Phys. 1984, 81, 3684–3690. [Google Scholar] [CrossRef] [Green Version]
- Flory, P.J. Statistical Mechanics of Chain Molecules; Interscience Publishers: New York, NY, USA, 1969. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model Number and Description | Trajectory Duration, ns | d, nm | P, nm | |
|---|---|---|---|---|
| 1: FA+Tpm+Tn | 104.2 | 2.75 (2.75) 1 | 166.2 (166.1) | 35.8 (35.6) |
| 2: FA+Tpm+Tn | 43.2 | 2.72 | 166.0 | 35.0 |
| 2: FA+Tpm | 204.8 | 2.71 (2.71) | 166.5 (166.2) | 36.1 (35.5) |
| Model Number and Description | , m | , 10 Nm | K, 10 N | G, 10 N |
|---|---|---|---|---|
| 1: FA+Tpm+Tn | 6.580 | 2.7 | 48.3 (51.6) 1 | 3.96 (4.45) |
| 2: FA+Tpm+Tn | 8.370 | 3.5 | 42.1 | 2.88 |
| 2: FA+Tpm | 8.061 | 3.3 | 41.0 (35.9) | 5.87 (7.05) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koubassova, N.A.; Tsaturyan, A.K. Molecular Dynamics Assessment of Mechanical Properties of the Thin Filaments in Cardiac Muscle. Int. J. Mol. Sci. 2023, 24, 4792. https://doi.org/10.3390/ijms24054792
Koubassova NA, Tsaturyan AK. Molecular Dynamics Assessment of Mechanical Properties of the Thin Filaments in Cardiac Muscle. International Journal of Molecular Sciences. 2023; 24(5):4792. https://doi.org/10.3390/ijms24054792
Chicago/Turabian StyleKoubassova, Natalia A., and Andrey K. Tsaturyan. 2023. "Molecular Dynamics Assessment of Mechanical Properties of the Thin Filaments in Cardiac Muscle" International Journal of Molecular Sciences 24, no. 5: 4792. https://doi.org/10.3390/ijms24054792