
Oligo—Not Only for Silencing: Overlooked Potential for Multidirectional Action in Plants

Abstract
:1. Introduction
2. Oligos Treatment—Alternative Method for Screening of Gene Functions
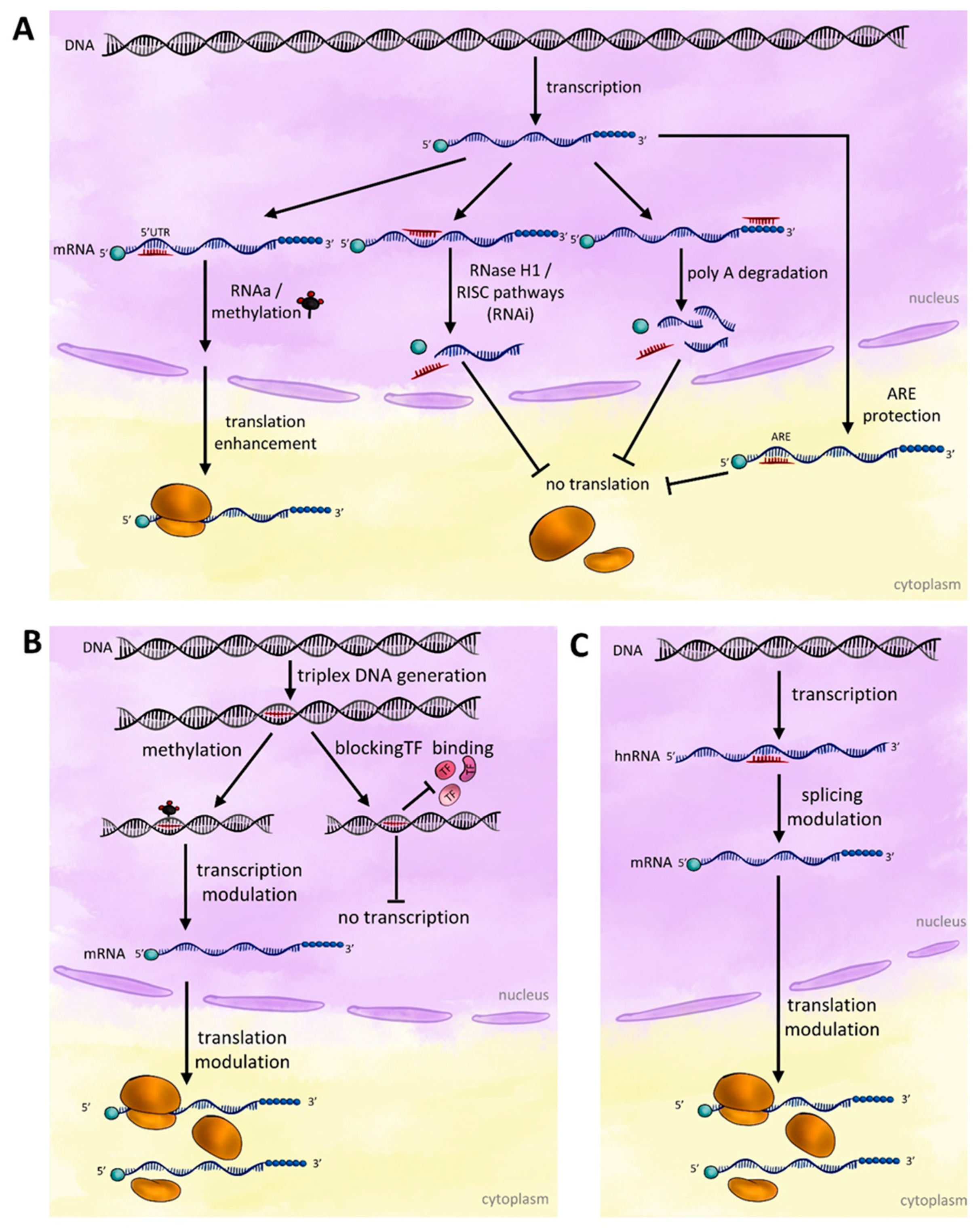
3. Mechanisms of Action
4. Oligo Design Based on the Relationship between Sequence and Effect on Gene Expression
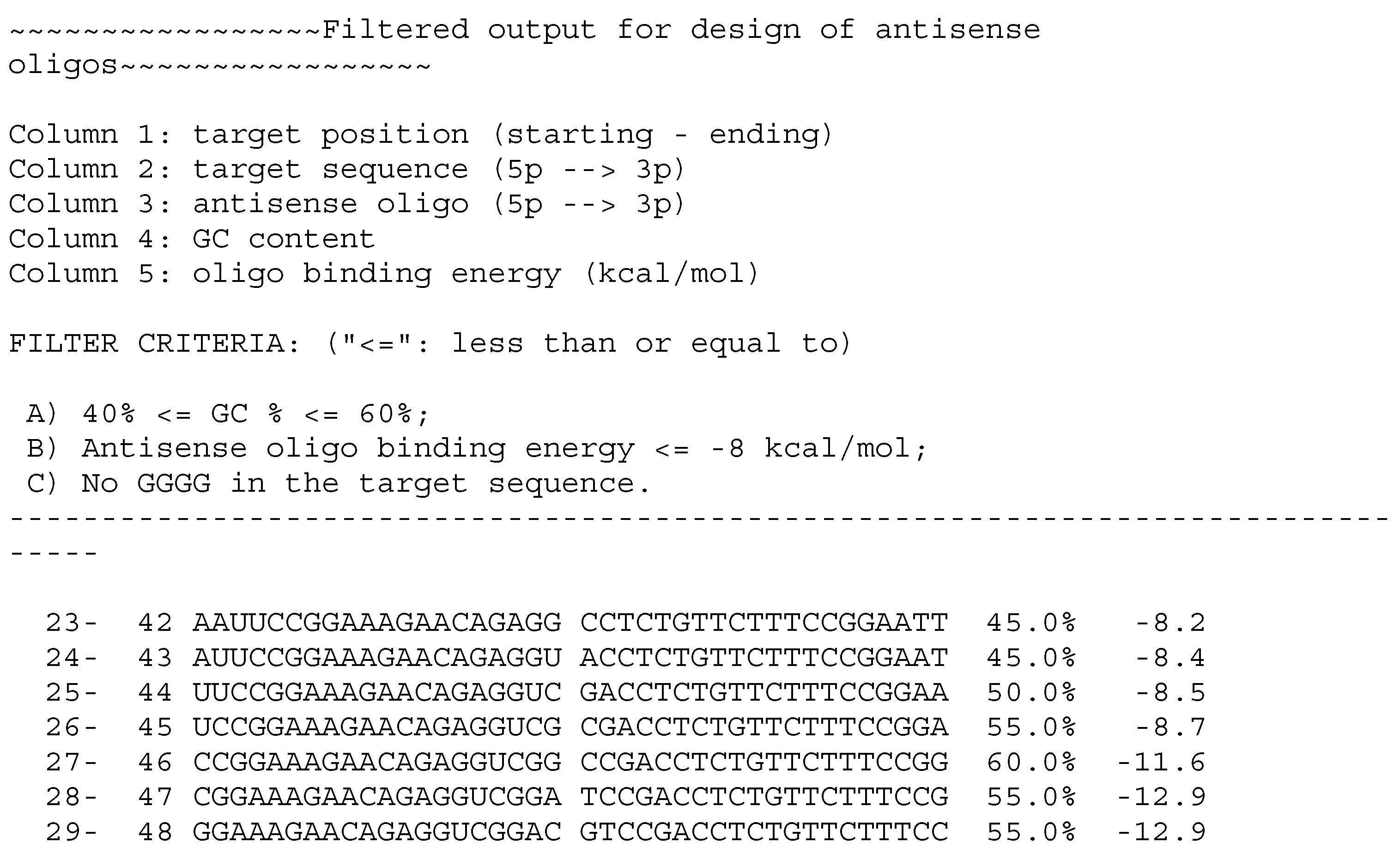
4.1. Basic Rules of Design
4.2. IT Tools
4.3. Oligo Action Based on Target Sequence
5. Methods of Introducing OLIGOs into the Plant Cell
6. Analysis Strategy for Oligo Actions
7. Summary
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Axtell, M.J.; Bartel, D.P. Antiquity of microRNAs and their targets in land plants. Plant Cell 2005, 17, 1658–1673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, K.; Rajewsky, N. The evolution of gene regulation by transcription factors and microRNAs. Nat. Rev. Genet. 2007, 8, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Shabalina, S.A.; Koonin, E.V. Origins and evolution of eukaryotic RNA interference. Trends Ecol. Evol. 2008, 23, 578–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pashkovskiy, P.P.; Ryazansky, S.S. Biogenesis, evolution, and functions of plant microRNAs. Biochemistry 2013, 78, 627–637. [Google Scholar] [CrossRef]
- Graham, I.R.; Dickson, G. Gene repair and mutagenesis mediated by chimeric RNA–DNA oligonucleotides: Chimeraplasty for gene therapy and conversion of single nucleotide polymorphisms (SNPs). Biochim. Biophys. Acta 2002, 1587, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Beetham, P.R.; Kipp, P.B.; Sawycky, X.L.; Arntzen, C.J.; May, G.D. A tool for functional plant genomics: Chimeric RNA/DNA oligonucleotides cause in vivo gene-specific mutations. Proc. Natl. Acad. Sci. USA 1999, 96, 8774–8778. [Google Scholar] [CrossRef] [Green Version]
- Kochevenko, A.; Willmitzer, L. Chimeric RNA/DNA oligonucleotide-based site-specific modification of the tobacco acetolactate syntase gene. Plant Physiol. 2003, 132, 174–184. [Google Scholar] [CrossRef] [Green Version]
- Sauer, N.J.; Mozoruk, J.; Miller, R.B.; Warburg, Z.J.; Walker, K.A.; Beetham, P.R.; Schöpke, C.R.; Gocal, G.F. Oligonucleotide-directed mutagenesis for precision gene editing. Plant Biotechnol. J. 2016, 14, 496–502. [Google Scholar] [CrossRef] [Green Version]
- Sauer, N.J.; Narváez-Vásquez, J.; Mozoruk, J.; Miller, R.B.; Warburg, Z.J.; Woodward, M.J.; Mihiret, Y.A.; Lincoln, T.A.; Segami, R.E.; Sanders, S.L.; et al. Oligonucleotide-Mediated Genome Editing Provides Precision and Function to Engineered Nucleases and Antibiotics in Plants. Plant Physiol. 2016, 170, 1917–1928. [Google Scholar] [CrossRef] [Green Version]
- Crooke, S.T.; Wang, S.; Vickers, T.A.; Shen, W.; Liang, X.H. Cellular uptake and trafficking of antisense oligonucleotides. Nat. Biotechnol. 2017, 35, 230–237. [Google Scholar] [CrossRef]
- Dinç, E.; Tóth, S.Z.; Schansker, G.; Ayaydin, F.; Kovács, L.; Dudits, D.; Garab, G.; Bottka, S. Synthetic antisense oligodeoxynucleotides to transiently suppress different nucleus- and chloroplast-encoded proteins of higher plant chloroplasts. Plant Physiol. 2011, 157, 1628–1641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dzialo, M.; Szopa, J.; Czuj, T.; Zuk, M. Oligodeoxynucleotides Can Transiently Up- and Downregulate CHS Gene Expression in Flax by Changing DNA Methylation in a Sequence-Specific Manner. Front. Plant Sci. 2017, 8, 755. [Google Scholar] [CrossRef] [PubMed]
- Dzialo, M.; Szopa, J.; Hnitecka, A.; Zuk, M. Transgenerational Perpetuation of CHS Gene Expression and DNA Methylation Status Induced by Short Oligodeoxynucleotides in Flax (Linum usitatissimum). Int. J. Mol. Sci. 2019, 20, 3983. [Google Scholar] [CrossRef] [Green Version]
- Liang, X.H.; Shen, W.; Sun, H.; Migawa, M.T.; Vickers, T.A.; Crooke, S.T. Translation efficiency of mRNAs is increased by antisense oligonucleotides targeting upstream open reading frames. Nat. Biotechnol. 2016, 34, 875–880. [Google Scholar] [CrossRef]
- Liang, X.H.; Sun, H.; Nichols, J.G.; Crooke, S.T. RNase H1-Dependent Antisense Oligonucleotides Are Robustly Active in Directing RNA Cleavage in Both the Cytoplasm and the Nucleus. Mol. Ther. 2017, 25, 2075–2092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, X.H.; Sun, H.; Shen, W.; Wang, S.; Yao, J.; Migawa, M.T.; Bui, H.H.; Damle, S.S.; Riney, S.; Graham, M.J.; et al. Antisense oligonucleotides targeting translation inhibitory elements in 5’ UTRs can selectively increase protein levels. Nucleic Acids Res. 2017, 45, 9528–9546. [Google Scholar] [CrossRef] [Green Version]
- Wojtasik, W.; Kulma, A.; Boba, A.; Szopa, J. Oligonucleotide treatment causes flax β-glucanase up-regulation via changes in gene-body methylation. BMC Plant Biol. 2014, 14, 261. [Google Scholar] [CrossRef] [Green Version]
- Gleave, M.E.; Monia, B.P. Antisense therapy for cancer. Nat. Rev. Cancer 2005, 5, 468–479. [Google Scholar] [CrossRef]
- Rayburn, E.R.; Zhang, R. Antisense, RNAi, and gene silencing strategies for therapy: Mission possible or impossible? Drug Discov. Today 2008, 13, 513–521. [Google Scholar] [CrossRef] [Green Version]
- Behlke, M.A. Chemical modification of siRNAs for in vivo use. Oligonucleotides 2008, 18, 305–319. [Google Scholar] [CrossRef] [Green Version]
- Sandy, P.; Ventura, A.; Jacks, T. Mammalian RNAi: A practical guide. Biotechniques 2005, 39, 215–224. [Google Scholar] [CrossRef] [Green Version]
- Scherer, L.J.; Rossi, J.J. Approaches for the sequence-specific knockdown of mRNA. Nat. Biotechnol. 2003, 21, 1457–1465. [Google Scholar] [CrossRef]
- Kurreck, J. Antisense technologies. Improvement through novel chemical modifications. Eur. J. Biochem. 2003, 270, 1628–1644. [Google Scholar] [CrossRef]
- Sun, C.; Ghebramedhin, H.; Höglund, A.S.; Jansson, C. Antisense oligodeoxynucleotide inhibition as a potent diagnostic tool for gene function in plant biology. Plant Signal. Behav. 2008, 3, 328–330. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.; Höglund, A.S.; Olsson, H.; Mangelsen, E.; Jansson, C. Antisense oligodeoxynucleotide inhibition as a potent strategy in plant biology: Identification of SUSIBA2 as a transcriptional activator in plant sugar signalling. Plant J. 2005, 44, 128–138. [Google Scholar] [CrossRef] [PubMed]
- Vickers, T.A.; Koo, S.; Bennett, C.F.; Crooke, S.T.; Dean, N.M.; Baker, B.F. Efficient reduction of target RNAs by small interfering RNA and RNase H-dependent antisense agents. A comparative analysis. J. Biol. Chem. 2003, 278, 7108–7118. [Google Scholar] [CrossRef] [Green Version]
- Bennett, C.F.; Cowsert, L.M. Application of antisense oligonucleotides for gene functionalization and target validation. Curr. Opin. Mol. Ther. 1999, 1, 359–371. [Google Scholar] [PubMed]
- Fabian, M.R.; Sonenberg, N.; Filipowicz, W. Regulation of mRNA translation and stability by microRNAs. Annu. Rev. Biochem. 2010, 79, 351–379. [Google Scholar] [CrossRef] [Green Version]
- Hammond, S.M. Dicing and slicing: The core machinery of the RNA interference pathway. FEBS Lett. 2005, 579, 5822–5829. [Google Scholar] [CrossRef] [Green Version]
- Elkon, R.; Zlotorynski, E.; Zeller, K.I.; Agami, R. Major role for mRNA stability in shaping the kinetics of gene induction. BMC Genomics 2010, 11, 259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, N.N.; Luo, D.; Singh, R.N. Pre-mRNA Splicing Modulation by Antisense Oligonucleotides. Methods Mol. Biol. 2018, 1828, 415–437. [Google Scholar] [CrossRef]
- Castanotto, D.; Lin, M.; Kowolik, C.; Wang, L.; Ren, X.Q.; Soifer, H.S.; Koch, T.; Hansen, B.R.; Oerum, H.; Armstrong, B.; et al. A cytoplasmic pathway for gapmer antisense oligonucleotide-mediated gene silencing in mammalian cells. Nucleic Acids Res. 2015, 43, 9350–9361. [Google Scholar] [CrossRef]
- Gheibi-Hayat, S.M.; Jamialahmadi, K. Antisense Oligonucleotide (AS-ODN) Technology: Principle, Mechanism and Challenges. Biotechnol. Appl. Biochem. 2021, 68, 1086–1094. [Google Scholar] [CrossRef] [PubMed]
- Wdowikowska, A.; Janicka, M. Antisense oligonucleotide technology as a research tool in plant biology. Funct. Plant Biol. 2021, 49, 1–12. [Google Scholar] [CrossRef]
- Escudé, C.; François, J.C.; Sun, J.S.; Ott, G.; Sprinzl, M.; Garestier, T.; Hélène, C. Stability of triple helices containing RNA and DNA strands: Experimental and molecular modeling studies. Nucleic Acids Res. 1993, 21, 5547–5553. [Google Scholar] [CrossRef] [PubMed]
- Shibuya, K.; Fukushima, S.; Takatsuji, H. RNA-directed DNA methylation induces transcriptional activation in plants. Proc. Natl. Acad. Sci. USA 2009, 106, 1660–1665. [Google Scholar] [CrossRef] [Green Version]
- Liang, Z.; Shen, L.; Cui, X.; Bao, S.; Geng, Y.; Yu, G.; Liang, F.; Xie, S.; Lu, T.; Gu, X.; et al. DNA N(6)-Adenine Methylation in Arabidopsis thaliana. Dev. Cell 2018, 45, 406–416.e403. [Google Scholar] [CrossRef] [Green Version]
- López, A.; Ramírez, V.; García-Andrade, J.; Flors, V.; Vera, P. The RNA silencing enzyme RNA polymerase v is required for plant immunity. PLoS Genet. 2011, 7, e1002434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Q.; Liu, S.; Yu, L.; Xiao, Y.; Zhang, S.; Wang, X.; Xu, Y.; Yu, H.; Li, Y.; Yang, J.; et al. RNA demethylation increases the yield and biomass of rice and potato plants in field trials. Nat. Biotechnol. 2021, 39, 1581–1588. [Google Scholar] [CrossRef]
- Zuk, M.; Szperlik, J.; Szopa, J. Linseed Silesia, Diverse Crops for Diverse Diets. New Solutions to Increase Dietary Lipids in Crop Species. Foods 2021, 10, 2675. [Google Scholar] [CrossRef]
- Seffer, I.; Nemeth, Z.; Hoffmann, G.; Matics, R.; Seffer, A.G.; Koller, A. Unexplored potentials of epigenetic mechanisms of plants and animals-theoretical considerations. Genet. Epigenet. 2013, 5, 23–41. [Google Scholar] [CrossRef]
- Kashihara, N.; Maeshima, Y.; Makino, H. Antisense oligonucleotides. Exp. Nephrol. 1998, 6, 84–88. [Google Scholar] [CrossRef]
- Xie, Z.; Sundström, J.F.; Jin, Y.; Liu, C.; Jansson, C.; Sun, C. A selection strategy in plant transformation based on antisense oligodeoxynucleotide inhibition. Plant J. 2014, 77, 954–961. [Google Scholar] [CrossRef] [Green Version]
- Chan, J.H.; Lim, S.; Wong, W.S. Antisense oligonucleotides: From design to therapeutic application. Clin. Exp. Pharmacol. Physiol. 2006, 33, 533–540. [Google Scholar] [CrossRef] [PubMed]
- Sohail, M.; Southern, E.M. Selecting optimal antisense reagents. Adv. Drug Deliv. Rev. 2000, 44, 23–34. [Google Scholar] [CrossRef]
- Sohail, M.; Southern, E.M. Hybridization of antisense reagents to RNA. Curr. Opin. Mol. Ther. 2000, 2, 264–271. [Google Scholar]
- Matveeva, O.V.; Tsodikov, A.D.; Giddings, M.; Freier, S.M.; Wyatt, J.R.; Spiridonov, A.N.; Shabalina, S.A.; Gesteland, R.F.; Atkins, J.F. Identification of sequence motifs in oligonucleotides whose presence is correlated with antisense activity. Nucleic Acids Res. 2000, 28, 2862–2865. [Google Scholar] [CrossRef] [Green Version]
- Ho, S.P.; Britton, D.H.; Stone, B.A.; Behrens, D.L.; Leffet, L.M.; Hobbs, F.W.; Miller, J.A.; Trainor, G.L. Potent antisense oligonucleotides to the human multidrug resistance-1 mRNA are rationally selected by mapping RNA-accessible sites with oligonucleotide libraries. Nucleic Acids Res. 1996, 24, 1901–1907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markham, N.R.; Zuker, M. UNAFold: Software for nucleic acid folding and hybridization. Methods Mol. Biol. 2008, 453, 3–31. [Google Scholar] [CrossRef] [PubMed]
- Gardner, P.P.; Giegerich, R. A comprehensive comparison of comparative RNA structure prediction approaches. BMC Bioinform. 2004, 5, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, L.; Andersen, K.B.; Hovgaard, L.; Jaroszewski, J.W. Rational selection of antisense oligonucleotide sequences. Eur. J. Pharm. Sci. 2000, 11, 191–198. [Google Scholar] [CrossRef]
- Baker, B.F.; Lot, S.S.; Condon, T.P.; Cheng-Flournoy, S.; Lesnik, E.A.; Sasmor, H.M.; Bennett, C.F. 2’-O-(2-Methoxy)ethyl-modified anti-intercellular adhesion molecule 1 (ICAM-1) oligonucleotides selectively increase the ICAM-1 mRNA level and inhibit formation of the ICAM-1 translation initiation complex in human umbilical vein endothelial cells. J. Biol. Chem. 1997, 272, 11994–12000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, M.; Zhang, N.; Gao, T.; Jin, J.; Jing, T.; Wang, J.; Wu, Y.; Wan, X.; Schwab, W.; Song, C. Sesquiterpene glucosylation mediated by glucosyltransferase UGT91Q2 is involved in the modulation of cold stress tolerance in tea plants. N. Phytol. 2020, 226, 362–372. [Google Scholar] [CrossRef]
- Zhang, H.; Demirer, G.S.; Zhang, H.; Ye, T.; Goh, N.S.; Aditham, A.J.; Cunningham, F.J.; Fan, C.; Landry, M.P. DNA nanostructures coordinate gene silencing in mature plants. Proc. Natl. Acad. Sci. USA 2019, 116, 7543–7548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmes-Hampton, G.P.; Crooks, D.R.; Haller, R.G.; Guo, S.; Freier, S.M.; Monia, B.P.; Rouault, T.A. Use of antisense oligonucleotides to correct the splicing error in ISCU myopathy patient cell lines. Hum. Mol. Genet. 2016, 25, 5178–5187. [Google Scholar] [CrossRef]
- Crooke, S.T.; Witztum, J.L.; Bennett, C.F.; Baker, B.F. RNA-Targeted Therapeutics. Cell Metab. 2018, 27, 714–739. [Google Scholar] [CrossRef] [Green Version]
- Dhillon, S. Viltolarsen: First Approval. Drugs 2020, 80, 1027–1031. [Google Scholar] [CrossRef]
- Keam, S.J. Inotersen: First Global Approval. Drugs 2018, 78, 1371–1376. [Google Scholar] [CrossRef]
- Paik, J.; Duggan, S. Volanesorsen: First Global Approval. Drugs 2019, 79, 1349–1354. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Hu, C.; Moufawad El Achkar, C.; Black, L.E.; Douville, J.; Larson, A.; Pendergast, M.K.; Goldkind, S.F.; Lee, E.A.; Kuniholm, A.; et al. Patient-Customized Oligonucleotide Therapy for a Rare Genetic Disease. N. Engl. J. Med. 2019, 381, 1644–1652. [Google Scholar] [CrossRef]
- Roberts, M.R. Fast-track applications: The potential for direct delivery of proteins and nucleic acids to plant cells for the discovery of gene function. Plant Methods 2005, 1, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Method | Tissue Type/Developmental Stage | Advantages | Disadvantages |
|---|---|---|---|
| infiltration under reduced pressure [11,12] | almost any type of tissue and any stage of development: whole plant, leaves, roots | quick and easy to perform method, many plants can be infiltrated at the same time, spiking is possible to a certain extent (easy to seedlings) | considerable stress, a large volume of oligonucleotide solution is needed at once and tissue residues contaminate the tissue, it is difficult to transfer the method to adult plants |
| infiltration through the stomata [53] | leaves only (the larger the better), best results with mature leaves, possible study in vivo | small volumes of the oligo solution required at once, no problem with impurities, no equipment required | more variable results, possibility of tissue damage, more difficult to do manually, especially troublesome with some small-leaved species such as flax |
| spraying the cells/tissue [43] | different developmental stages and organs, possible life examination for above-ground part of the plant only | simple scale-up, hardly any equipment required | more variable results, more difficult to control quantitative application of oligo |
| uptake in a sugar solution [17,24,25,43] | whole plant, leaves, roots, best results with seedlings, possible study in vivo | quick and easy to perform method, hardly any equipment re-quired | large volume of oligonucleotide solution is needed at once, it is difficult to transfer the method to adult plants |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krasnodębski, C.; Sawuła, A.; Kaźmierczak, U.; Żuk, M. Oligo—Not Only for Silencing: Overlooked Potential for Multidirectional Action in Plants. Int. J. Mol. Sci. 2023, 24, 4466. https://doi.org/10.3390/ijms24054466
Krasnodębski C, Sawuła A, Kaźmierczak U, Żuk M. Oligo—Not Only for Silencing: Overlooked Potential for Multidirectional Action in Plants. International Journal of Molecular Sciences. 2023; 24(5):4466. https://doi.org/10.3390/ijms24054466
Chicago/Turabian StyleKrasnodębski, Cezary, Agnieszka Sawuła, Urszula Kaźmierczak, and Magdalena Żuk. 2023. "Oligo—Not Only for Silencing: Overlooked Potential for Multidirectional Action in Plants" International Journal of Molecular Sciences 24, no. 5: 4466. https://doi.org/10.3390/ijms24054466