
Pathophysiological Aspects of Muscle Atrophy and Osteopenia Induced by Chronic Constriction Injury (CCI) of the Sciatic Nerve in Rats
 , , , , ,
, , , , ,  , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Results
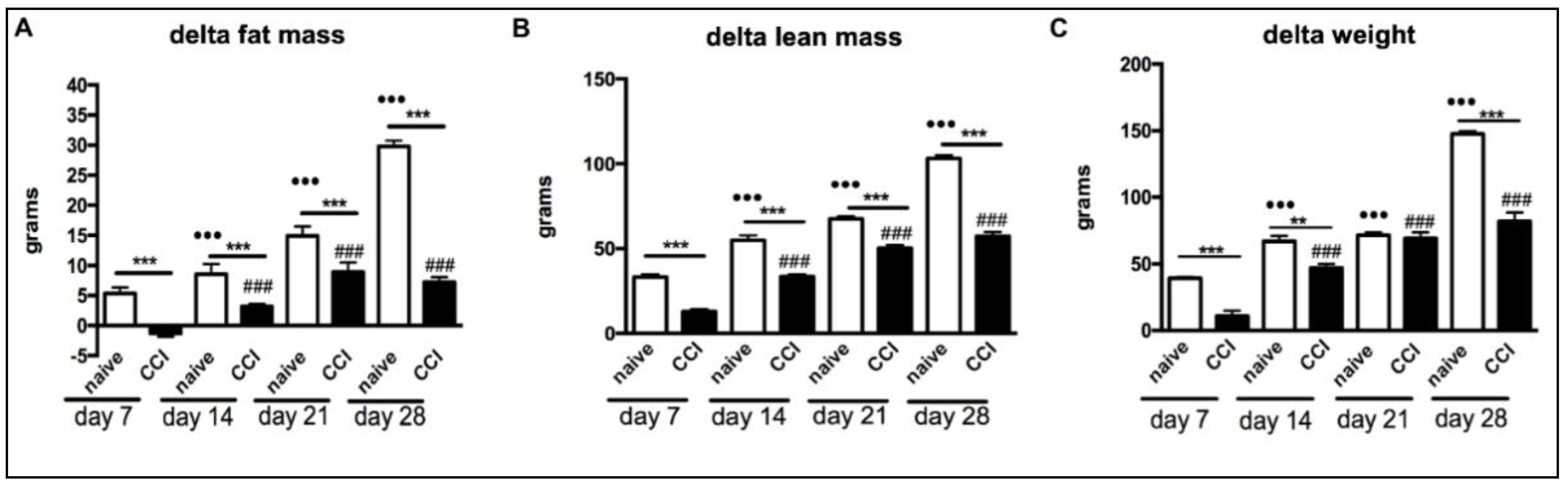
2.1. Body Weight and Body Composition of Rats That Underwent Chronic Constriction Injury
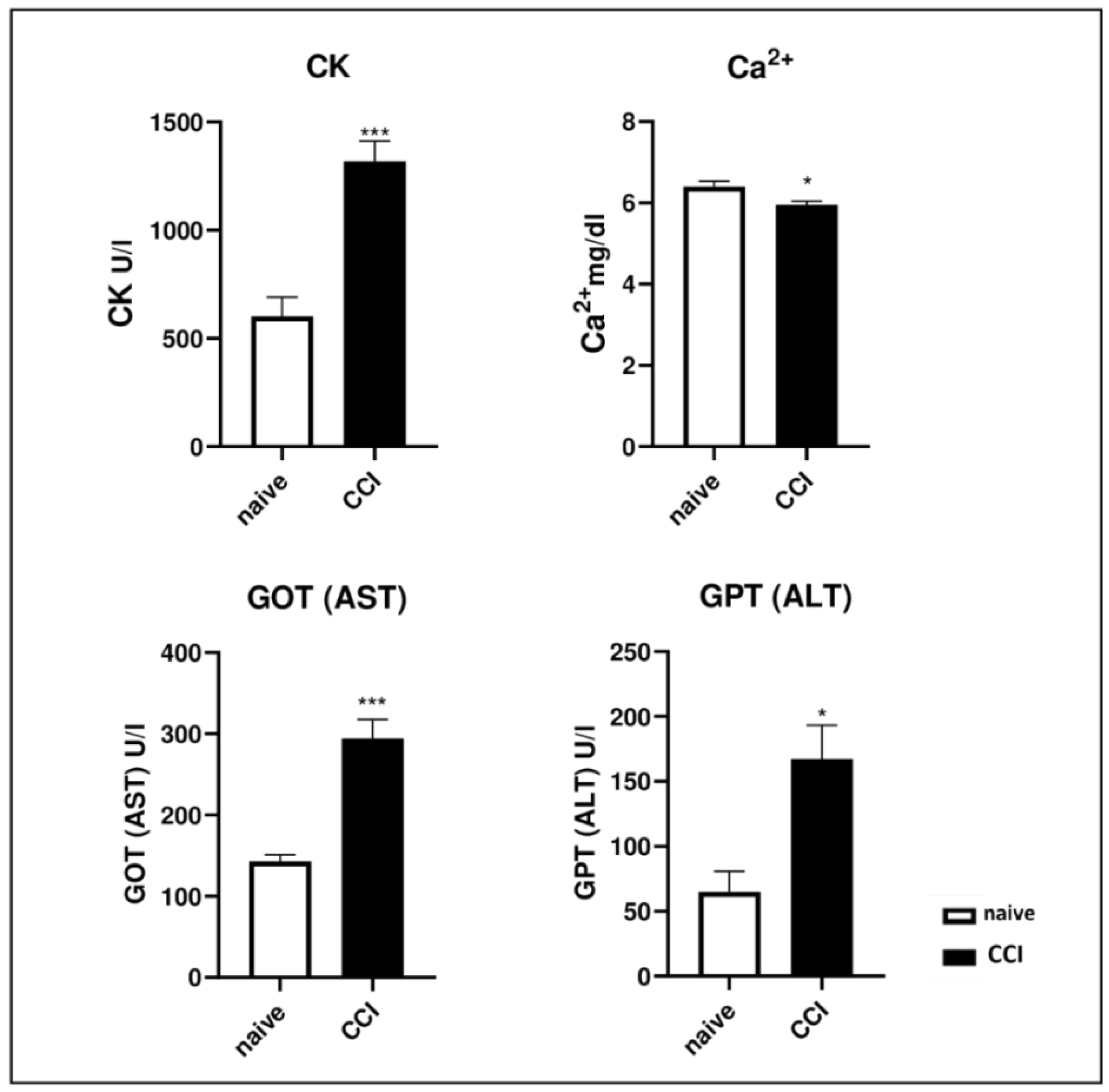
2.2. Biochemical Analysis after Chronic Constriction Injury
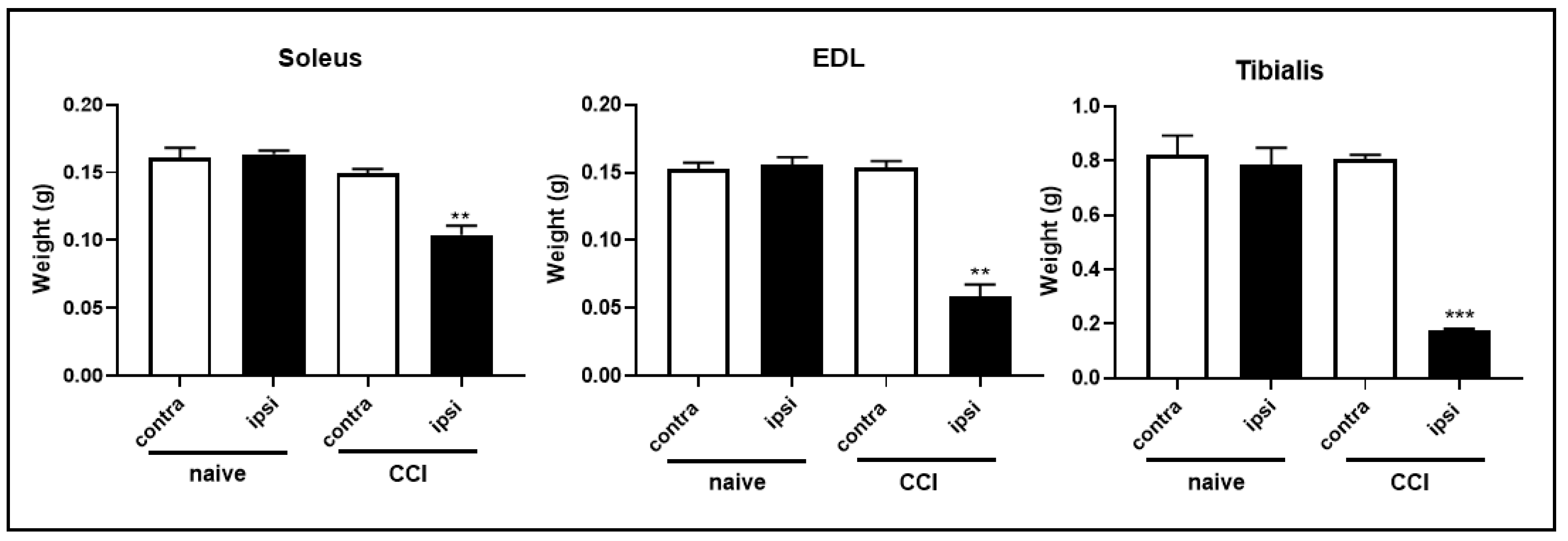
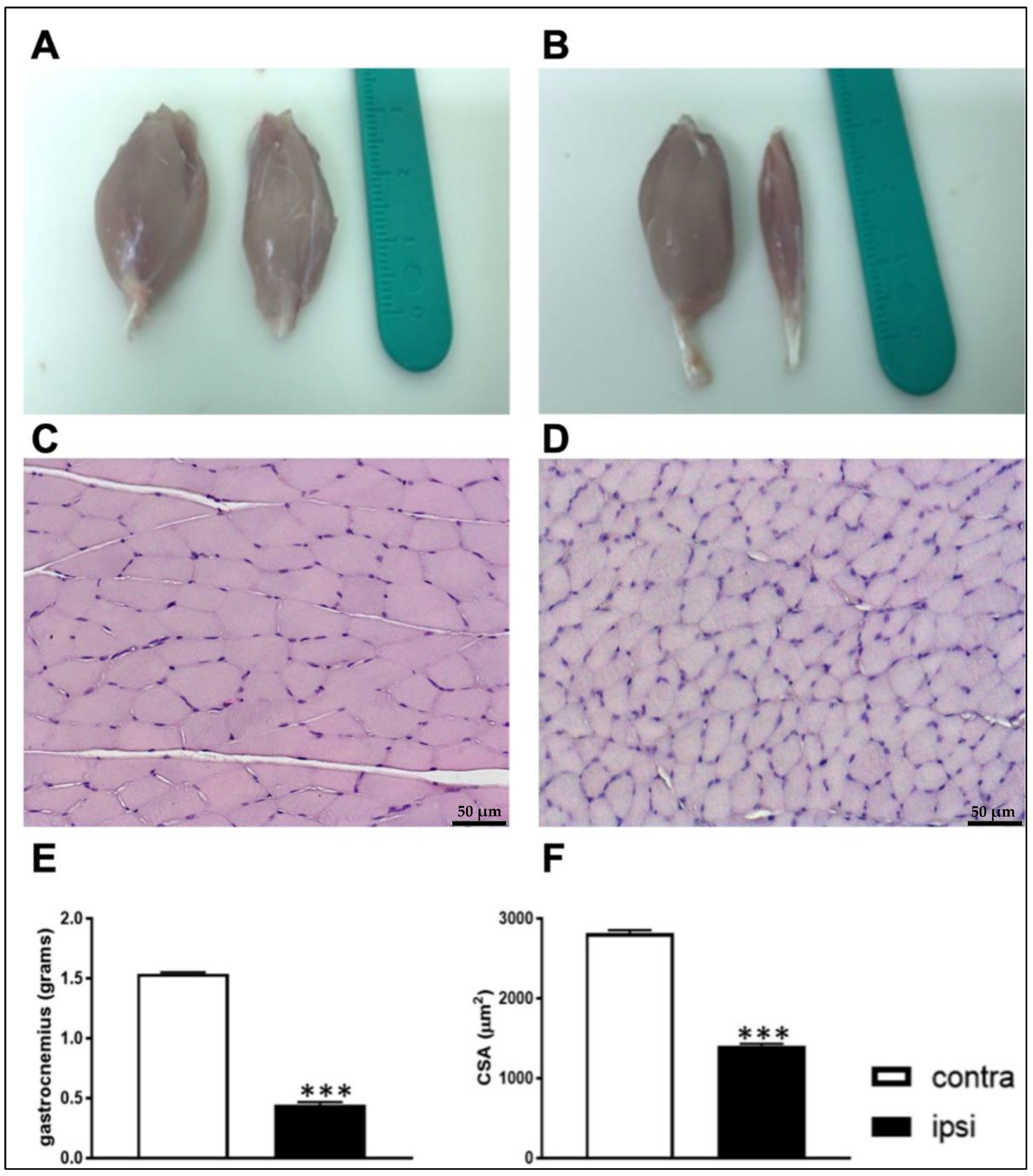
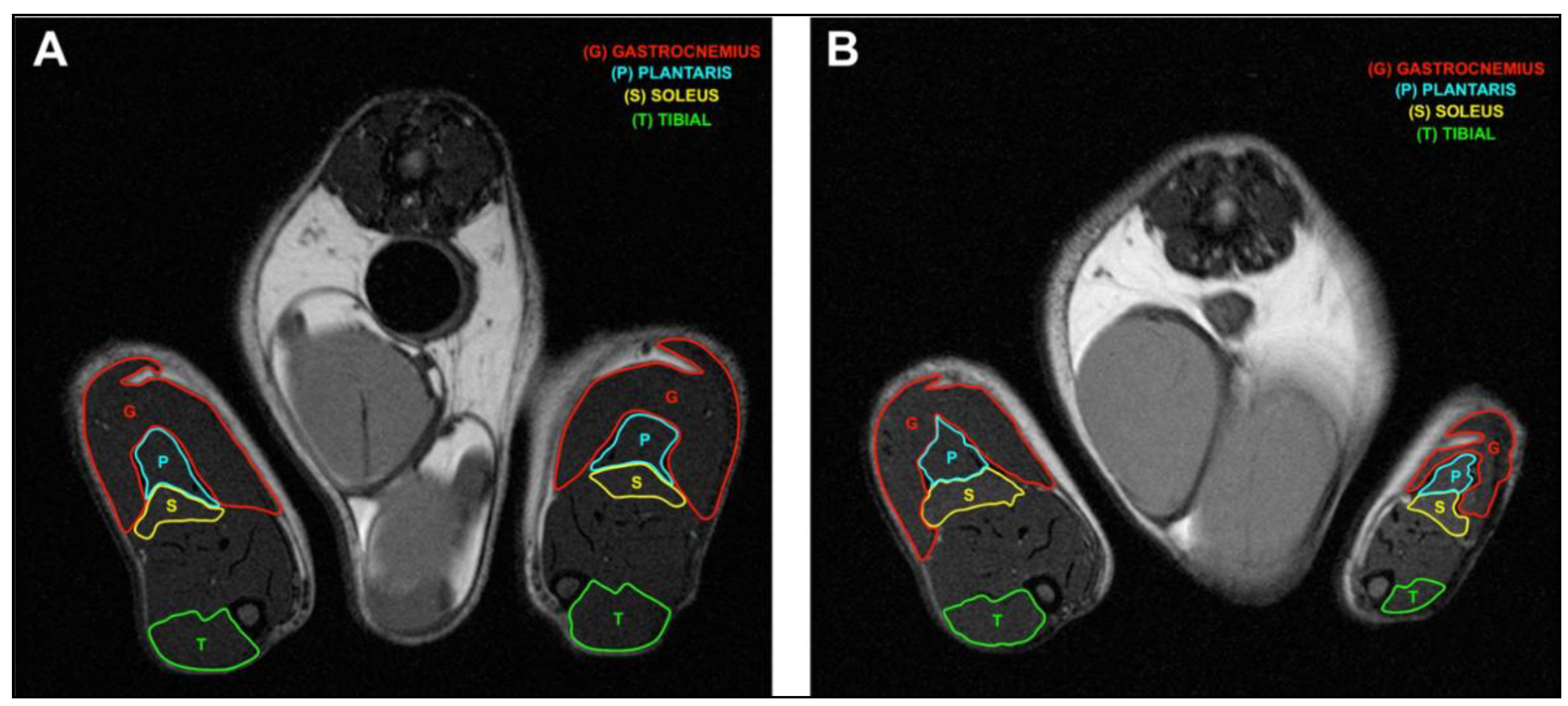
2.3. Constriction Injury of the Sciatic Nerve Induced Muscle Wasting in Rats
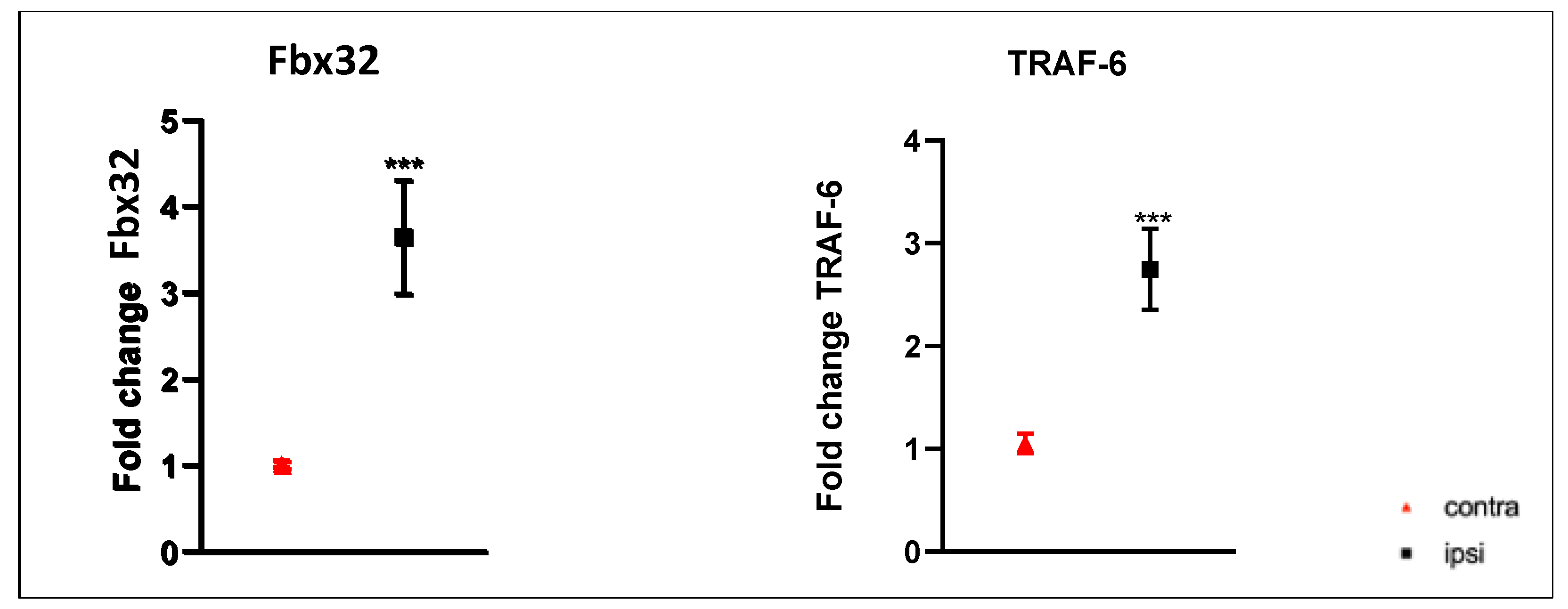
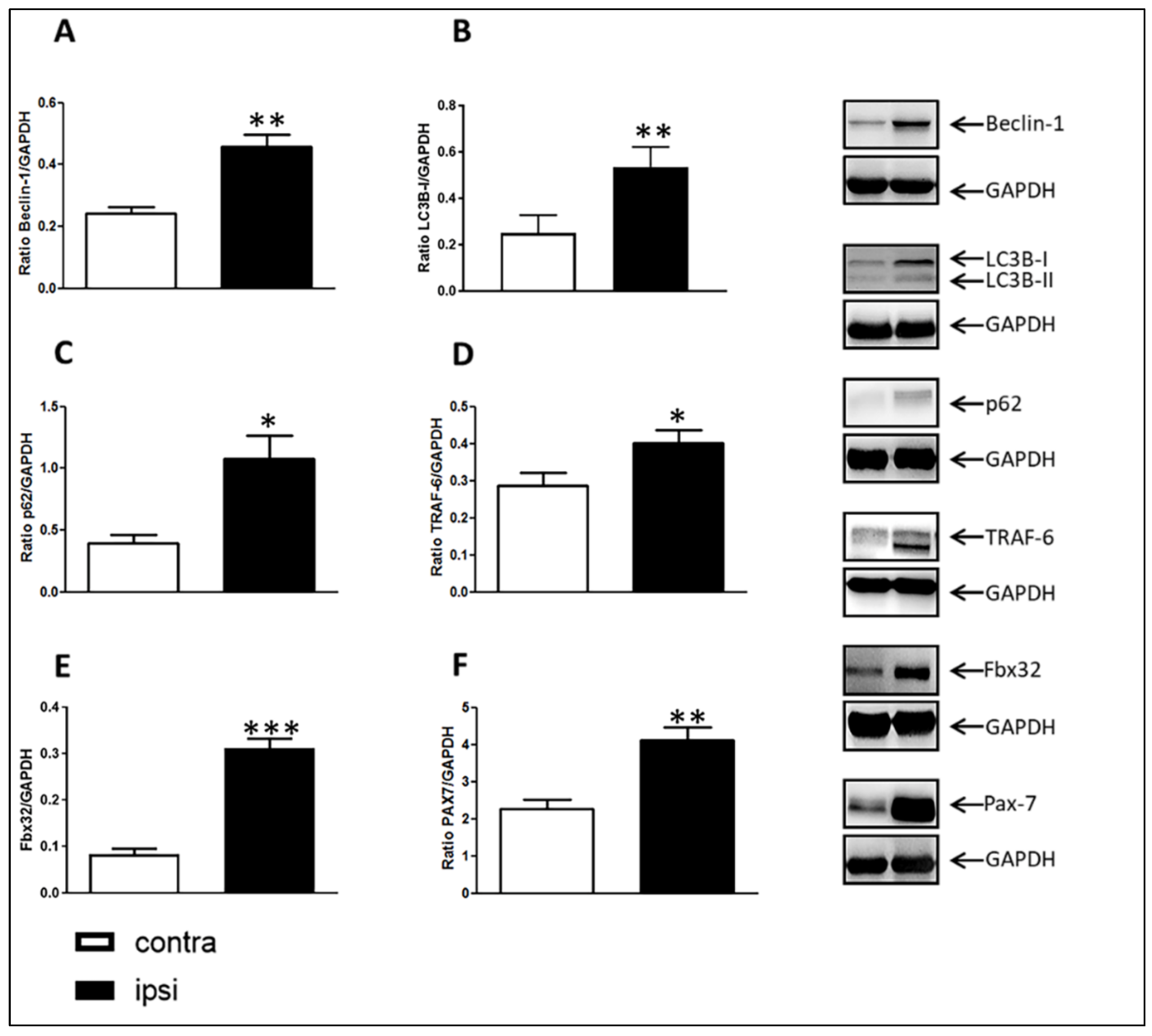
2.4. E3 Ubiquitin Ligase Gene Expression Was Up-Regulated in Gastrocnemius Muscle after CCI of the Sciatic Nerve
2.5. Catabolic Pathways in Rat Gastrocnemius Muscle after Chronic Sciatic Nerve Constriction Injury
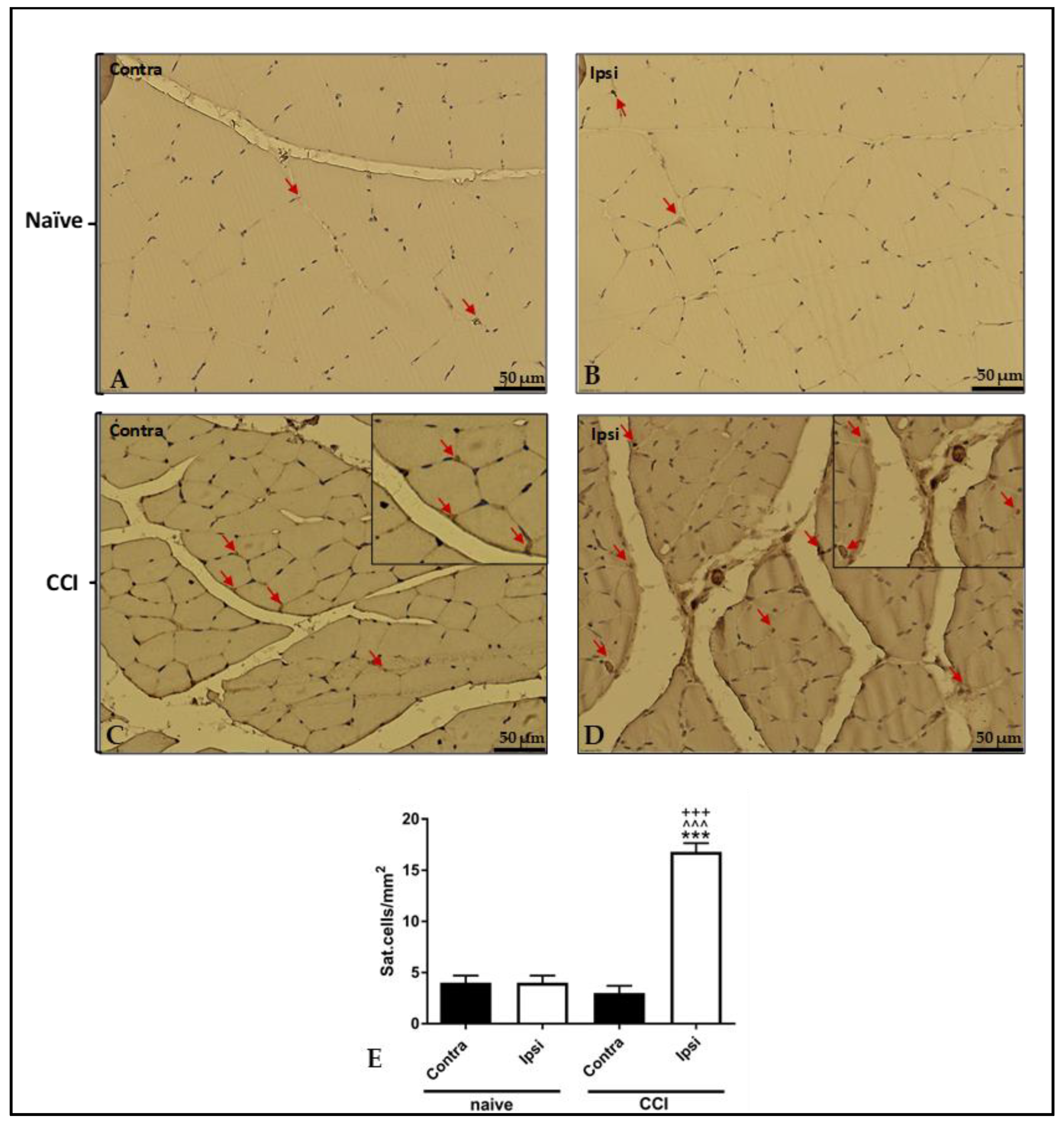
2.6. Increased Stem Cell Presence in Ipsilateral Gastrocnemius Evaluated with Pax-7 Immunostaining
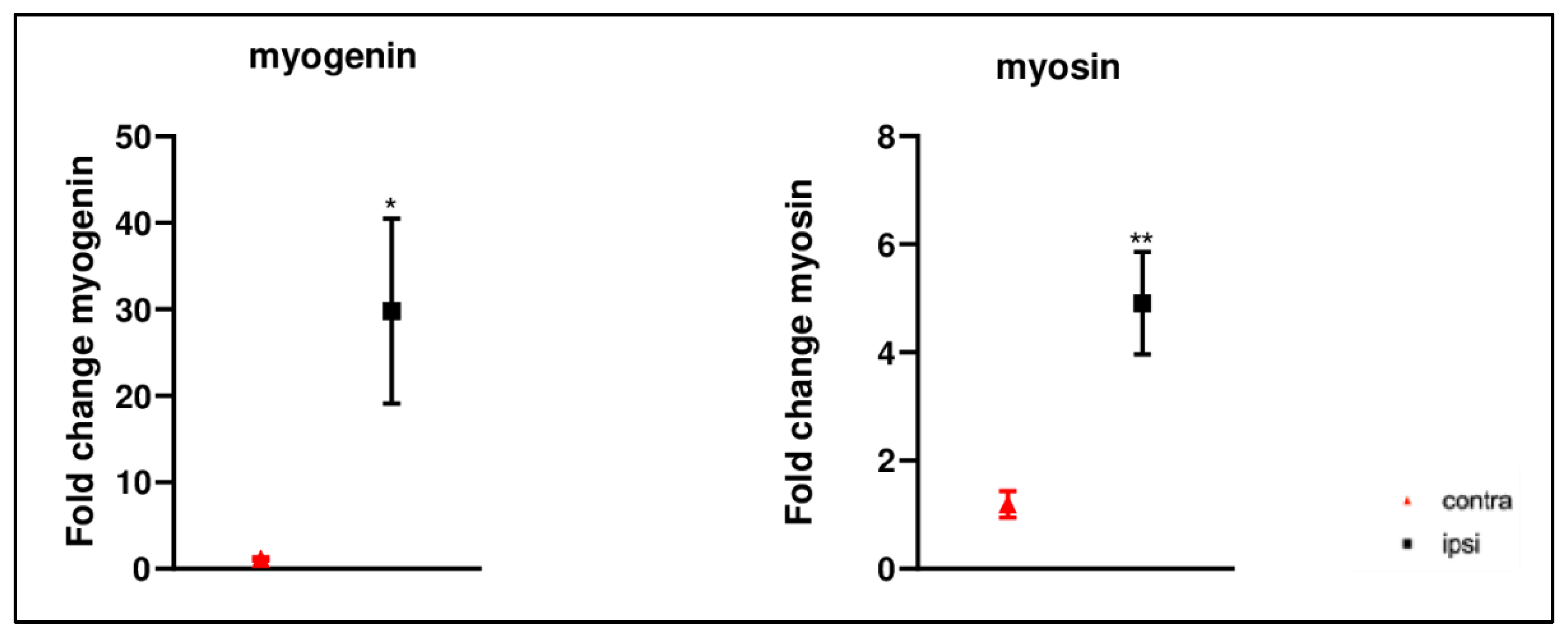
2.7. Increased Myogenic Markers Gene Expression in Gastrocnemius Muscle after CCI of Sciatic Nerve
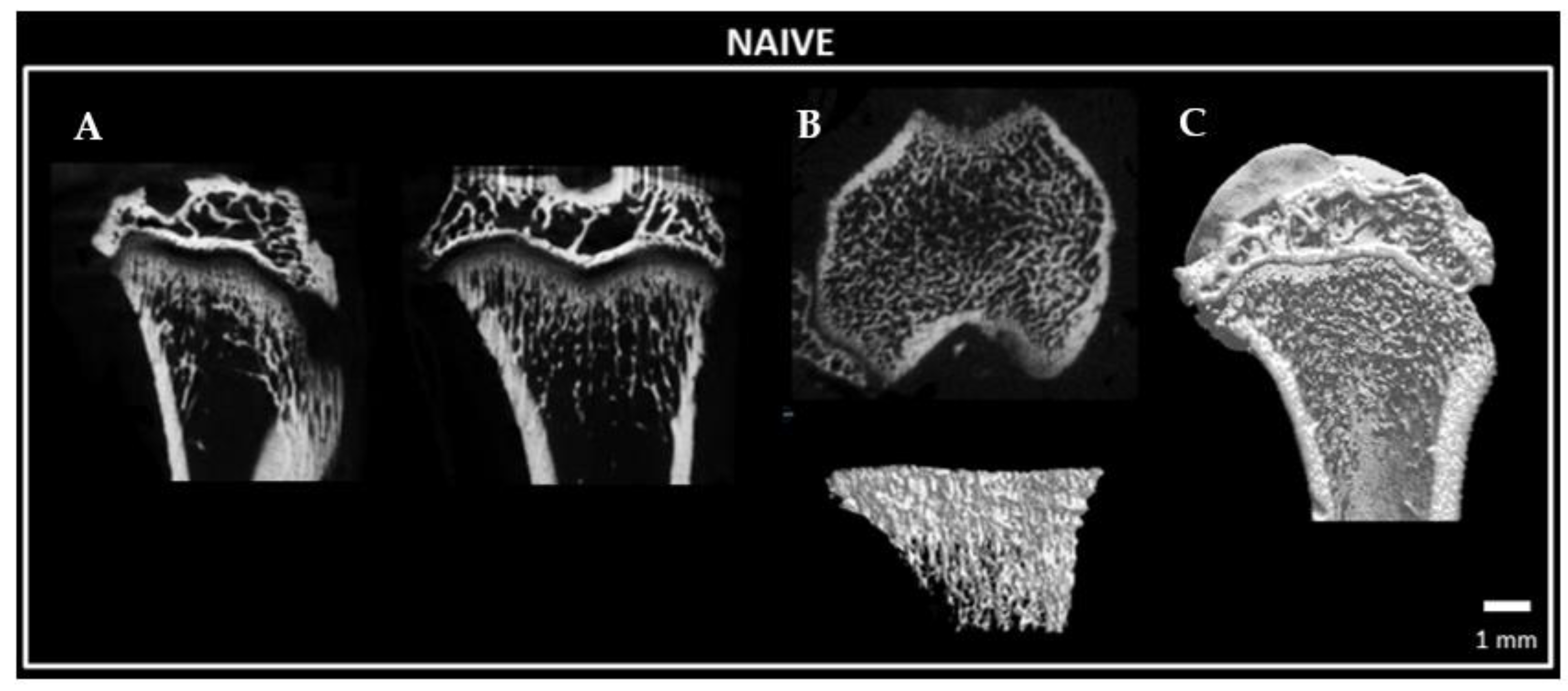
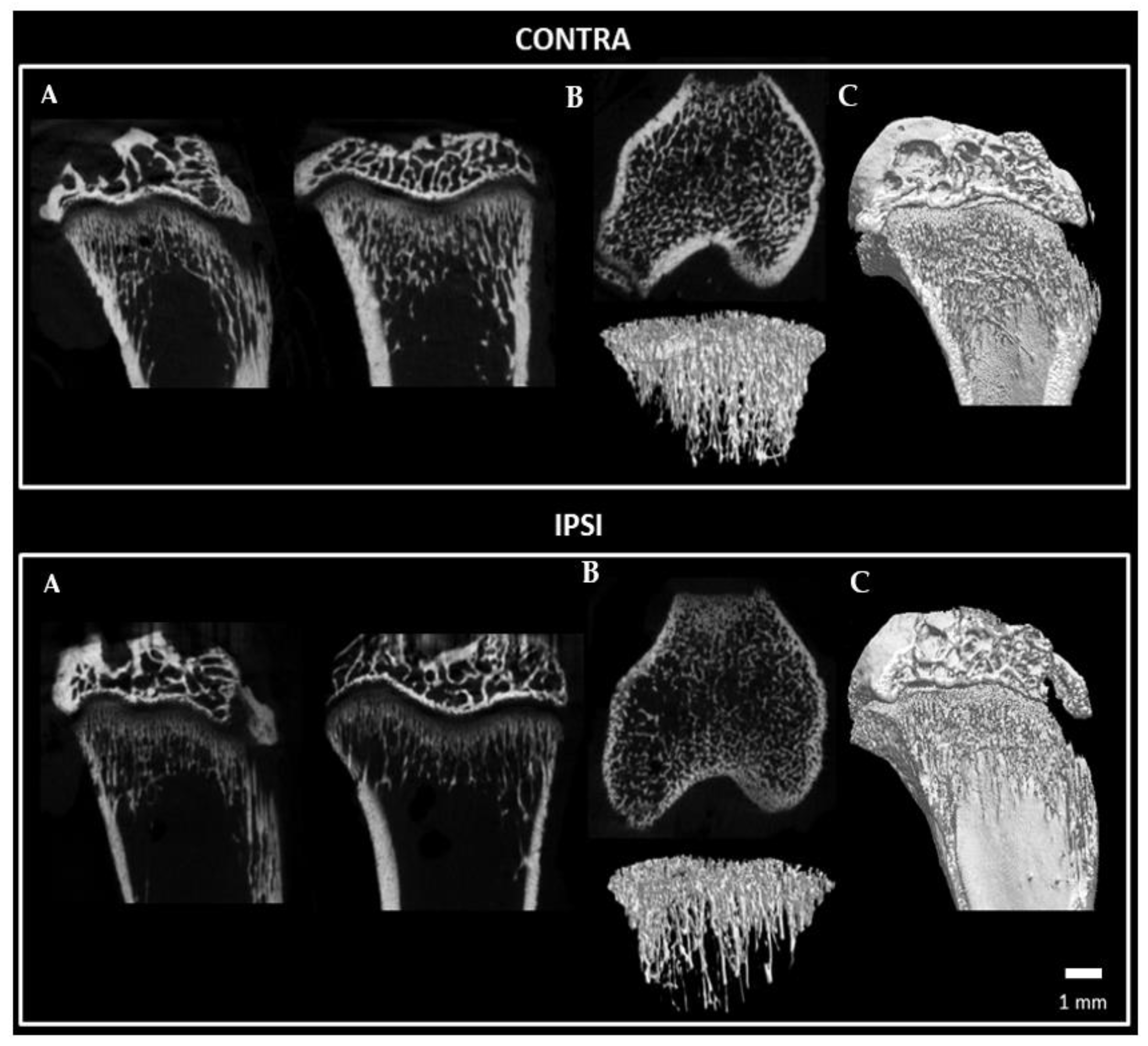
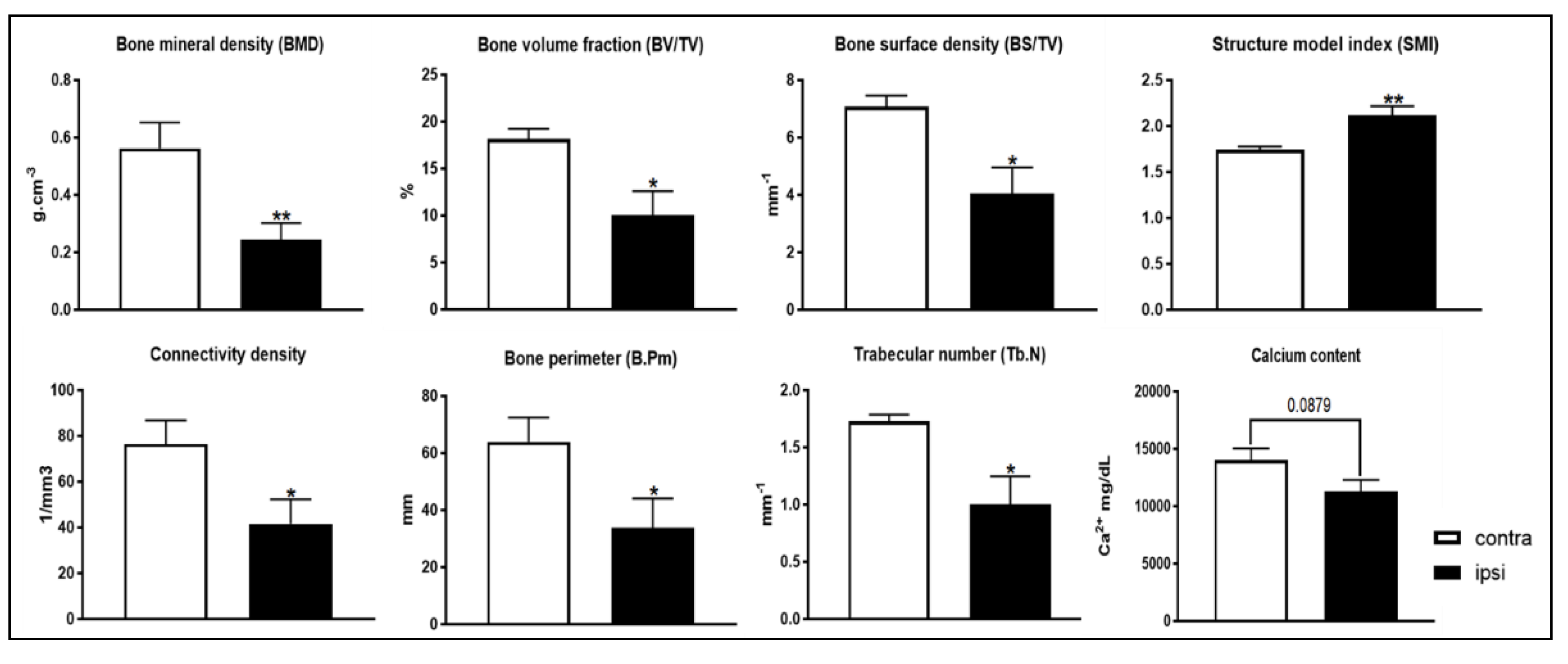
2.8. Bone Microstructure Observation of Tibia Bone following Constriction Injury of Sciatic Nerve
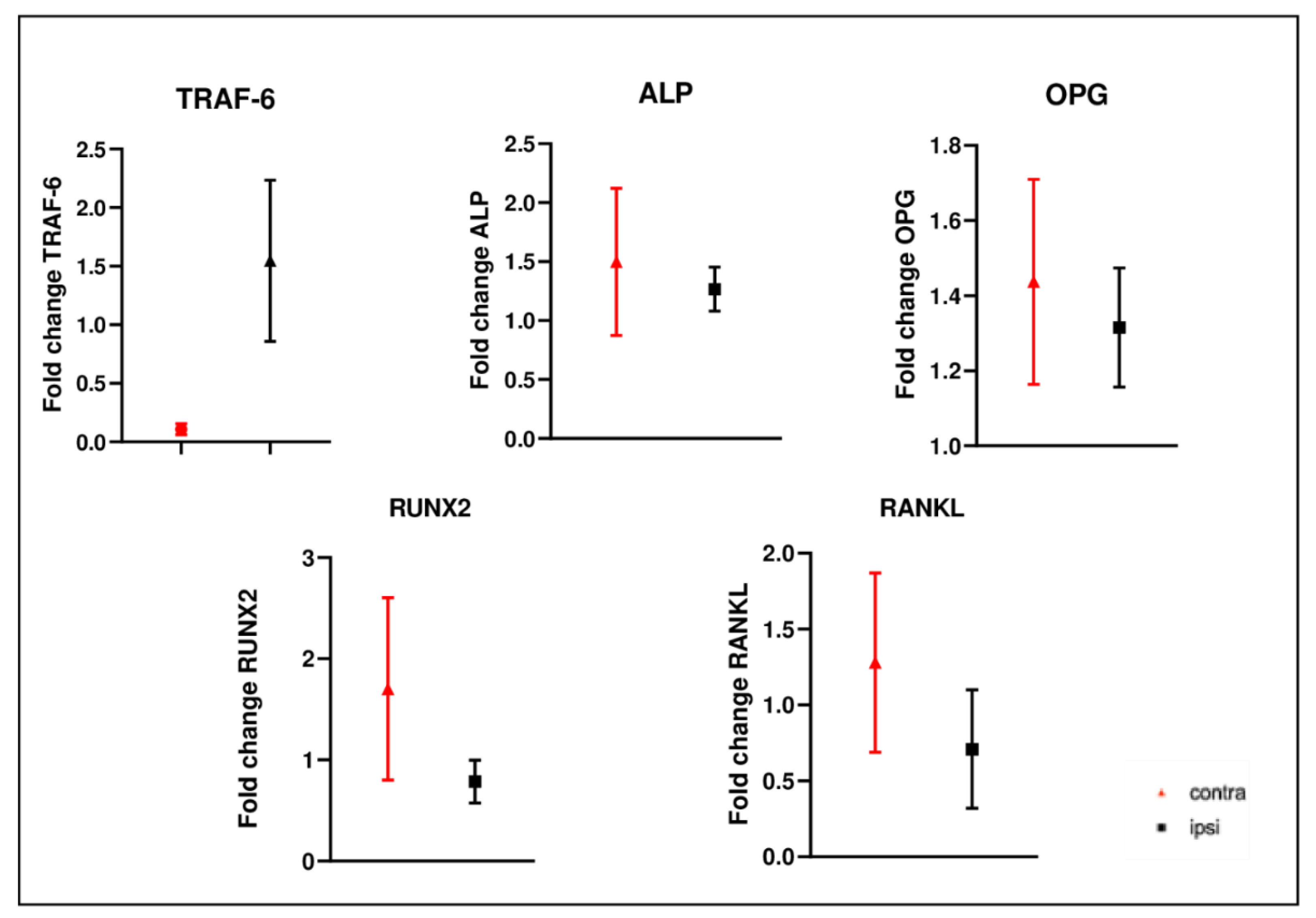
2.9. Gene Expression Analysis in Femurs after CCI of Sciatic Nerve
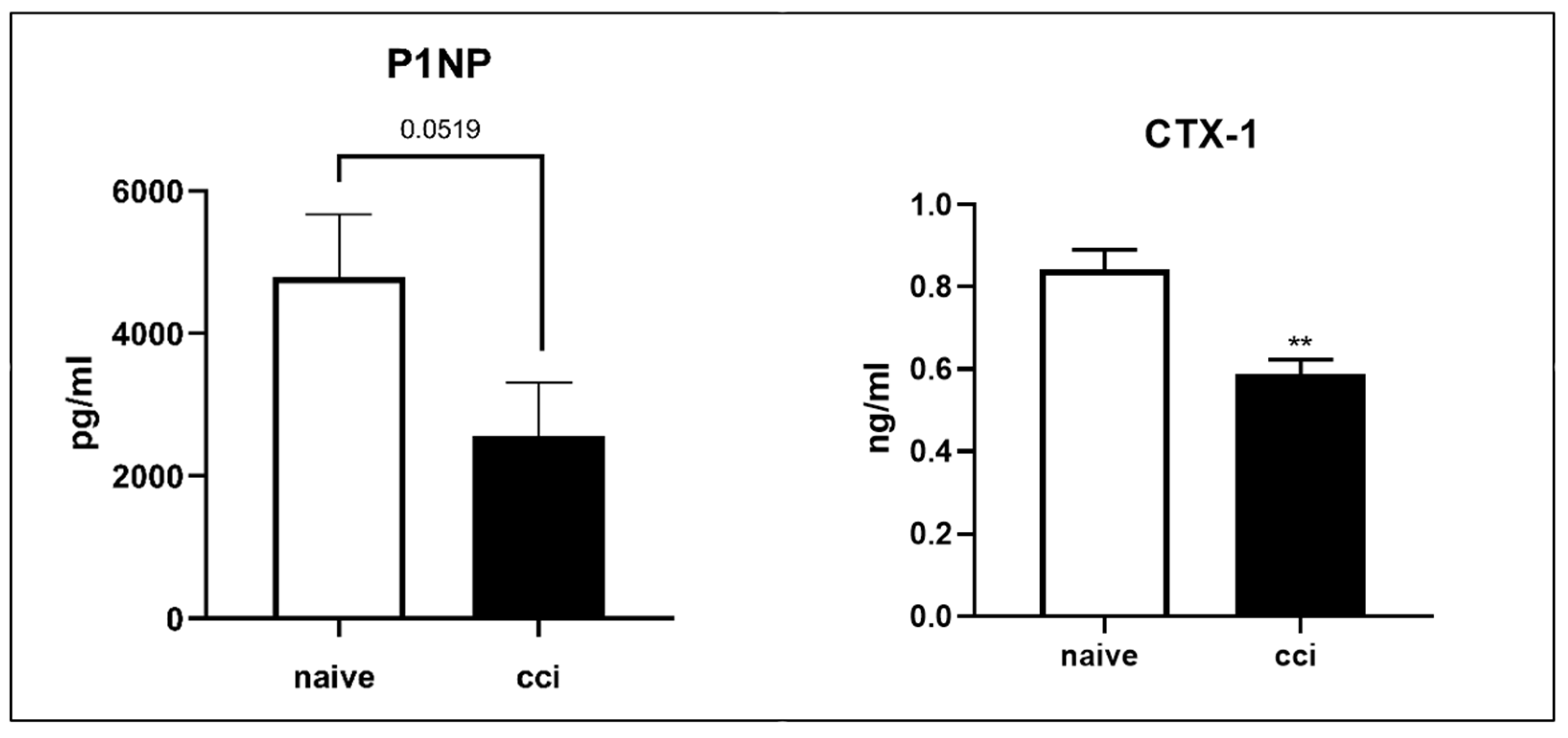
2.10. Serum Markers Procollagen Type 1 Amino-Terminal Propeptide (P1NP) and (C-Terminal Collagen Cross-Linkage 1) CTX-1 Analysis after CCI of the Sciatic Nerve
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Chronic Constriction Injury
4.3. Analysis of Blood-Based Endpoints
4.4. Body Composition Analysis
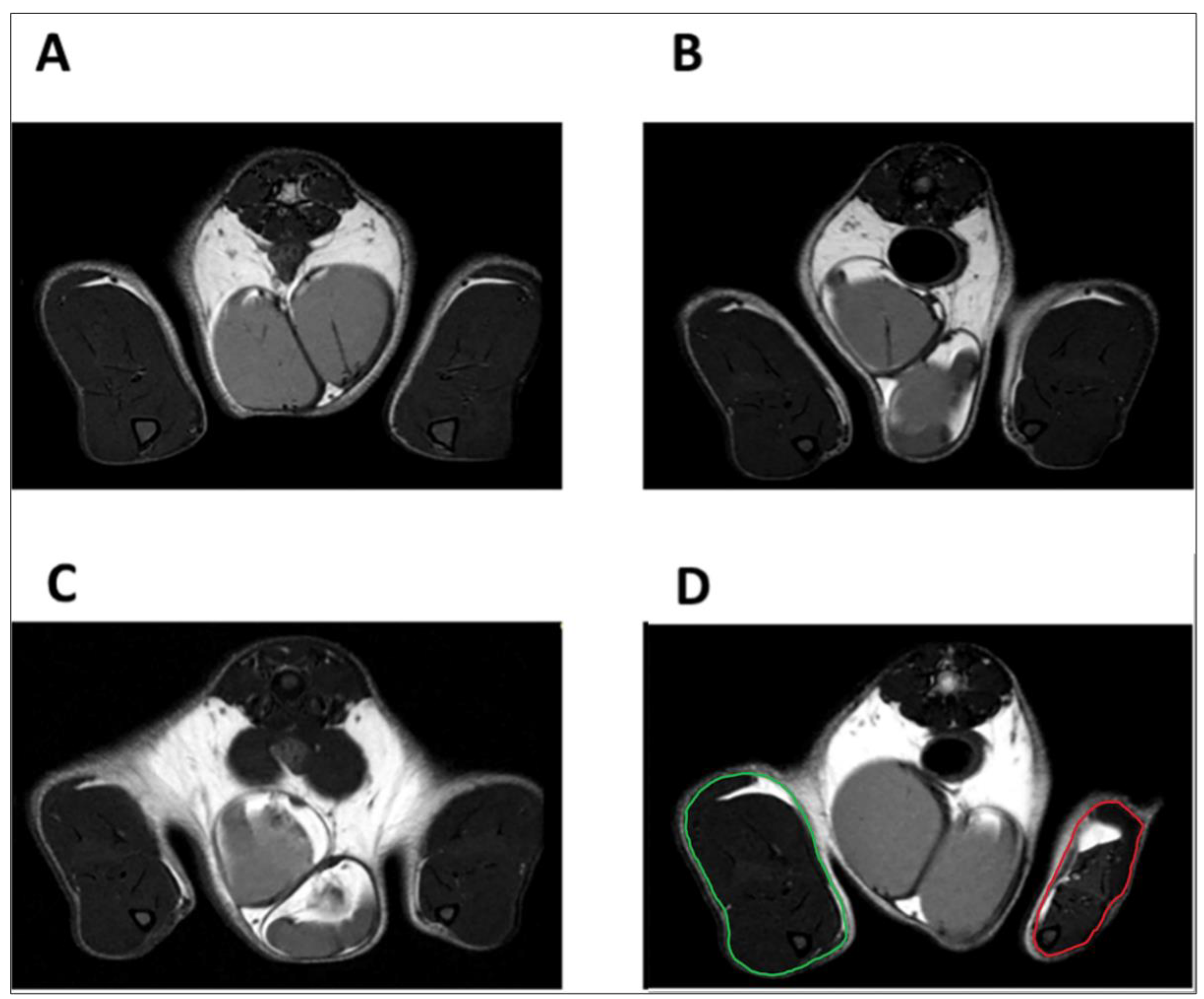
4.5. Magnetic Resonance Imaging
4.6. Muscle Segmentation
4.7. Protein Extraction and Western Blot
4.8. Histological Analysis
4.9. Immunohistochemistry
4.10. Micro-Computed Tomography
4.11. Bone Calcium Content
4.12. Reverse Transcription and Quantitative Real-Time PCR (qRT-PCR) Performed on GC Muscle Tissue
4.13. Reverse Transcription and Quantitative Real-Time PCR (qRT-PCR) Performed on Femur Bones
4.14. Real-Time PCR Data Analysis
4.15. ELISA Analysis
4.16. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Argilés, J.M.; Campos, N.; Lopez-Pedrosa, J.M.; Rueda, R.; Rodriguez-Mañas, L. Skeletal Muscle Regulates Metabolism via Interorgan Crosstalk: Roles in Health and Disease. J. Am. Med. Dir. Assoc. 2016, 17, 789–796. [Google Scholar] [CrossRef]
- Chargi, N.; Bril, S.I.; Swartz, J.E.; Wegner, I.; Willems, S.M.; de Bree, R. Skeletal muscle mass is an imaging biomarker for decreased survival in patients with oropharyngeal squamous cell carcinoma. Oral Oncol. 2020, 101, 104519. [Google Scholar] [CrossRef]
- von Haehling, S.; Garfias Macedo, T.; Valentova, M.; Anker, M.S.; Ebner, N.; Bekfani, T.; Haarmann, H.; Schefold, J.C.; Lainscak, M.; Cleland, J.G.F.; et al. Muscle wasting as an independent predictor of survival in patients with chronic heart failure. J. Cachexia Sarcopenia Muscle 2020, 11, 1242–1249. [Google Scholar] [CrossRef] [PubMed]
- Tieland, M.; Trouwborst, I.; Clark, B.C. Skeletal muscle performance and ageing. J. Cachexia Sarcopenia Muscle 2018, 9, 3–19. [Google Scholar] [CrossRef] [PubMed]
- Constantin-Teodosiu, D.; Constantin, D. Molecular Mechanisms of Muscle Fatigue. Int. J. Mol. Sci. 2021, 22, 11587. [Google Scholar] [CrossRef] [PubMed]
- Ebert, S.M.; Al-Zougbi, A.; Bodine, S.C.; Adams, C.M. Skeletal Muscle Atrophy: Discovery of Mechanisms and Potential Therapies. Physiology (Bethesda) 2019, 34, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Bosco, F.; Musolino, V.; Gliozzi, M.; Nucera, S.; Carresi, C.; Zito, M.C.; Scarano, F.; Scicchitano, M.; Reale, F.; Ruga, S.; et al. The muscle to bone axis (and viceversa): An encrypted language affecting tissues and organs and yet to be codified? Pharmacol. Res. 2021, 165, 105427. [Google Scholar] [CrossRef]
- Calvani, R.; Miccheli, A.; Landi, F.; Bossola, M.; Cesari, M.; Leeuwenburgh, C.; Sieber, C.C.; Bernabei, R.; Marzetti, E. Current nutritional recommendations and novel dietary strategies to manage sarcopenia. J. Frailty Aging 2013, 2, 38–53. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Meng, X.; Zhang, Z.; Wang, T. Physical Exercise for Muscle Atrophy. Adv. Exp. Med. Biol. 2018, 1088, 529–545. [Google Scholar] [CrossRef] [PubMed]
- Kirk, B.; Feehan, J.; Lombardi, G.; Duque, G. Muscle, Bone, and Fat Crosstalk: The Biological Role of Myokines, Osteokines, and Adipokines. Curr. Osteoporos. Rep. 2020, 18, 388–400. [Google Scholar] [CrossRef]
- Ramdharry, G. Peripheral nerve disease. Handb. Clin. Neurol. 2018, 159, 403–415. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.E.; Meyer, R.M.; Faillace, W.J.; Landau, M.E.; Smith, J.K.; McKay, P.L.; Nesti, L.J. Combat Injury of the Sciatic Nerve—An Institutional Experience. Mil. Med. 2018, 183, e434–e441. [Google Scholar] [CrossRef] [PubMed]
- Welch, J.A. Peripheral nerve injury. Semin Vet. Med. Surg. Small Anim. 1996, 11, 273–284. [Google Scholar] [CrossRef]
- Bennett, G.J.; Xie, Y.K. A peripheral mononeuropathy in rat that produces disorders of pain sensation like those seen in man. Pain 1988, 33, 87–107. [Google Scholar] [CrossRef] [PubMed]
- Langer, H.T.; Senden, J.M.G.; Gijsen, A.P.; Kempa, S.; van Loon, L.J.C.; Spuler, S. Muscle Atrophy Due to Nerve Damage Is Accompanied by Elevated Myofibrillar Protein Synthesis Rates. Front. Physiol. 2018, 9, 1220. [Google Scholar] [CrossRef] [PubMed]
- Langer, H.T.; Afzal, S.; Kempa, S.; Spuler, S. Nerve damage induced skeletal muscle atrophy is associated with increased accumulation of intramuscular glucose and polyol pathway intermediates. Sci. Rep. 2020, 10, 1908. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Bhatnagar, S.; Paul, P.K. TWEAK and TRAF6 regulate skeletal muscle atrophy. Curr. Opin. Clin. Nutr. Metab. Care 2012, 15, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Musolino, V.; Palus, S.; Latouche, C.; Gliozzi, M.; Bosco, F.; Scarano, F.; Nucera, S.; Carresi, C.; Scicchitano, M.; von Haehling, S.; et al. Cardiac expression of neutrophil gelatinase-associated lipocalin in a model of cancer cachexia-induced cardiomyopathy. ESC Heart Fail. 2019, 6, 89–97. [Google Scholar] [CrossRef]
- Vainshtein, A.; Sandri, M. Signaling Pathways That Control Muscle Mass. Int. J. Mol. Sci. 2020, 21, 4759. [Google Scholar] [CrossRef] [PubMed]
- Guadagnin, E.; Mázala, D.; Chen, Y.W. STAT3 in Skeletal Muscle Function and Disorders. Int. J. Mol. Sci. 2018, 19, 2265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anthony, T.G. Mechanisms of protein balance in skeletal muscle. Domest. Anim. Endocrinol. 2016, 56, S23–S32. [Google Scholar] [CrossRef]
- Gordon, B.S.; Kelleher, A.R.; Kimball, S.R. Regulation of muscle protein synthesis and the effects of catabolic states. Int. J. Biochem. Cell Biol. 2013, 45, 2147–2157. [Google Scholar] [CrossRef]
- Musolino, V.; Palus, S.; Tschirner, A.; Drescher, C.; Gliozzi, M.; Carresi, C.; Vitale, C.; Muscoli, C.; Doehner, W.; von Haehling, S.; et al. Megestrol acetate improves cardiac function in a model of cancer cachexia-induced cardiomyopathy by autophagic modulation. J. Cachexia Sarcopenia Muscle 2016, 7, 555–566. [Google Scholar] [CrossRef] [PubMed]
- Paul, P.K.; Bhatnagar, S.; Mishra, V.; Srivastava, S.; Darnay, B.G.; Choi, Y.; Kumar, A. The E3 ubiquitin ligase TRAF6 intercedes in starvation-induced skeletal muscle atrophy through multiple mechanisms. Mol. Cell Biol. 2012, 32, 1248–1259. [Google Scholar] [CrossRef]
- Paul, P.K.; Kumar, A. TRAF6 coordinates the activation of autophagy and ubiquitin-proteasome systems in atrophying skeletal muscle. Autophagy 2011, 7, 555–556. [Google Scholar] [CrossRef]
- Schiaffino, S.; Dyar, K.A.; Ciciliot, S.; Blaauw, B.; Sandri, M. Mechanisms regulating skeletal muscle growth and atrophy. FEBS J. 2013, 280, 4294–4314. [Google Scholar] [CrossRef]
- Song, J.Y.; Pineault, K.M.; Wellik, D.M. Development, repair, and regeneration of the limb musculoskeletal system. Curr. Top. Dev. Biol. 2019, 132, 451–486. [Google Scholar] [CrossRef]
- Mañas-García, L.; Guitart, M.; Duran, X.; Barreiro, E. Satellite Cells and Markers of Muscle Regeneration during Unloading and Reloading: Effects of Treatment with Resveratrol and Curcumin. Nutrients 2020, 12, 1870. [Google Scholar] [CrossRef] [PubMed]
- McKenna, C.F.; Fry, C.S. Altered satellite cell dynamics accompany skeletal muscle atrophy during chronic illness, disuse, and aging. Curr. Opin. Clin. Nutr. Metab. Care 2017, 20, 447–452. [Google Scholar] [CrossRef]
- Bettis, T.; Kim, B.J.; Hamrick, M.W. Impact of muscle atrophy on bone metabolism and bone strength: Implications for muscle-bone crosstalk with aging and disuse. Osteoporos. Int. 2018, 29, 1713–1720. [Google Scholar] [CrossRef] [PubMed]
- Bateman, T.A.; Dunstan, C.R.; Lacey, D.L.; Ferguson, V.L.; Ayers, R.A.; Simske, S.J. Osteoprotegerin ameliorates sciatic nerve crush induced bone loss. J. Orthop. Res. 2001, 19, 518–523. [Google Scholar] [CrossRef] [PubMed]
- Suyama, H.; Moriwaki, K.; Niida, S.; Maehara, Y.; Kawamoto, M.; Yuge, O. Osteoporosis following chronic constriction injury of sciatic nerve in rats. J. Bone Miner. Metab. 2002, 20, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Strotmeyer, E.S.; Cauley, J.A.; Schwartz, A.V.; de Rekeneire, N.; Resnick, H.E.; Zmuda, J.M.; Shorr, R.I.; Tylavsky, F.A.; Vinik, A.I.; Harris, T.B.; et al. Reduced peripheral nerve function is related to lower hip BMD and calcaneal QUS in older white and black adults: The Health, Aging, and Body Composition Study. J. Bone Miner. Res. 2006, 21, 1803–1810. [Google Scholar] [CrossRef] [PubMed]
- Fonseca-Rodrigues, D.; Amorim, D.; Almeida, A.; Pinto-Ribeiro, F. Emotional and cognitive impairments in the peripheral nerve chronic constriction injury model (CCI) of neuropathic pain: A systematic review. Behav. Brain Res. 2021, 399, 113008. [Google Scholar] [CrossRef]
- Matias Júnior, I.; Medeiros, P.; de Freita, R.L.; Vicente-César, H.; Ferreira Junior, J.R.; Machado, H.R.; Menezes-Reis, R. Effective Parameters for Gait Analysis in Experimental Models for Evaluating Peripheral Nerve Injuries in Rats. Neurospine 2019, 16, 305–316. [Google Scholar] [CrossRef] [PubMed]
- Austin, P.J.; Wu, A.; Moalem-Taylor, G. Chronic constriction of the sciatic nerve and pain hypersensitivity testing in rats. J. Vis. Exp. 2012, 61, e3393. [Google Scholar] [CrossRef]
- Nathwani, R.A.; Pais, S.; Reynolds, T.B.; Kaplowitz, N. Serum alanine aminotransferase in skeletal muscle diseases. Hepatology 2005, 41, 380–382. [Google Scholar] [CrossRef]
- Green, R.M.; Flamm, S. AGA technical review on the evaluation of liver chemistry tests. Gastroenterology 2002, 123, 1367–1384. [Google Scholar] [CrossRef]
- Baird, M.F.; Graham, S.M.; Baker, J.S.; Bickerstaff, G.F. Creatine-kinase- and exercise-related muscle damage implications for muscle performance and recovery. J. Nutr. Metab. 2012, 2012, 960363. [Google Scholar] [CrossRef]
- Choe, M.A.; Kim, K.H.; An, G.J.; Lee, K.S.; Heitkemper, M. Hindlimb muscle atrophy occurs from peripheral nerve damage in a rat neuropathic pain model. Biol. Res. Nurs. 2011, 13, 44–54. [Google Scholar] [CrossRef]
- Richner, M.; Bjerrum, O.J.; Nykjaer, A.; Vaegter, C.B. The spared nerve injury (SNI) model of induced mechanical allodynia in mice. J. Vis. Exp. 2011, 54, e3092. [Google Scholar] [CrossRef]
- Beehler, B.C.; Sleph, P.G.; Benmassaoud, L.; Grover, G.J. Reduction of skeletal muscle atrophy by a proteasome inhibitor in a rat model of denervation. Exp. Biol. Med. 2006, 231, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Daemen, M.A.; Kurvers, H.A.; Bullens, P.H.; Slaaf, D.W.; Freling, G.; Kitslaar, P.J.; van den Wildenberg, F.A. Motor denervation induces altered muscle fibre type densities and atrophy in a rat model of neuropathic pain. NeuroSci. Lett. 1998, 247, 204–208. [Google Scholar] [CrossRef] [PubMed]
- Arnold, W.D.; Kassar, D.; Kissel, J.T. Spinal muscular atrophy: Diagnosis and management in a new therapeutic era. Muscle Nerve 2015, 51, 157–167. [Google Scholar] [CrossRef]
- Wu, H.; Li, L.; Su, X. Vagus nerve through α7 nAChR modulates lung infection and inflammation: Models, cells, and signals. Biomed Res. Int. 2014, 2014, 283525. [Google Scholar] [CrossRef]
- Zhao, Y.X.; He, W.; Jing, X.H.; Liu, J.L.; Rong, P.J.; Ben, H.; Liu, K.; Zhu, B. Transcutaneous auricular vagus nerve stimulation protects endotoxemic rat from lipopolysaccharide-induced inflammation. Evid. Based Complement. Alternat. Med. 2012, 2012, 627023. [Google Scholar] [CrossRef] [PubMed]
- Leterme, D.; Tyc, F. Re-innervation and recovery of rat soleus muscle and motor unit function after nerve crush. Exp. Physiol. 2004, 89, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Shimkus, K.L.; Jefferson, L.S.; Gordon, B.S.; Kimball, S.R. Repressors of mTORC1 act to blunt the anabolic response to feeding in the soleus muscle of a cast-immobilized mouse hindlimb. Physiol. Rep. 2018, 6, e13891. [Google Scholar] [CrossRef]
- Jaspers, S.R.; Tischler, M.E. Atrophy and growth failure of rat hindlimb muscles in tail-cast suspension. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1984, 57, 1472–1479. [Google Scholar] [CrossRef] [PubMed]
- Bederman, I.R.; Lai, N.; Shuster, J.; Henderson, L.; Ewart, S.; Cabrera, M.E. Chronic hindlimb suspension unloading markedly decreases turnover rates of skeletal and cardiac muscle proteins and adipose tissue triglycerides. J. Appl. Physiol. (1985) 2015, 119, 16–26. [Google Scholar] [CrossRef] [Green Version]
- Wall, B.T.; Snijders, T.; Senden, J.M.; Ottenbros, C.L.; Gijsen, A.P.; Verdijk, L.B.; van Loon, L.J. Disuse impairs the muscle protein synthetic response to protein ingestion in healthy men. J. Clin. Endocrinol. Metab. 2013, 98, 4872–4881. [Google Scholar] [CrossRef] [PubMed]
- Phillips, S.M. A brief review of critical processes in exercise-induced muscular hypertrophy. Sports Med. 2014, 44 (Suppl. S1), S71–S77. [Google Scholar] [CrossRef] [PubMed]
- Talbert, E.E.; Smuder, A.J.; Min, K.; Kwon, O.S.; Szeto, H.H.; Powers, S.K. Immobilization-induced activation of key proteolytic systems in skeletal muscles is prevented by a mitochondria-targeted antioxidant. J. Appl. Physiol. (1985) 2013, 115, 529–538. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.L.; Yeo, D. Mitochondrial dysregulation and muscle disuse atrophy. F1000Res 2019, 8, 1621. [Google Scholar] [CrossRef] [PubMed]
- Hyatt, H.; Deminice, R.; Yoshihara, T.; Powers, S.K. Mitochondrial dysfunction induces muscle atrophy during prolonged inactivity: A review of the causes and effects. Arch. Biochem. Biophys. 2019, 662, 49–60. [Google Scholar] [CrossRef]
- Cohen, S.; Nathan, J.A.; Goldberg, A.L. Muscle wasting in disease: Molecular mechanisms and promising therapies. Nat. Rev. Drug Discov. 2015, 14, 58–74. [Google Scholar] [CrossRef]
- Joshi, S.K.; Kim, H.T.; Feeley, B.T.; Liu, X. Differential ubiquitin-proteasome and autophagy signaling following rotator cuff tears and suprascapular nerve injury. J. Orthop. Res. 2014, 32, 138–144. [Google Scholar] [CrossRef]
- Johansen, T.; Lamark, T. Selective autophagy mediated by autophagic adapter proteins. Autophagy 2011, 7, 279–296. [Google Scholar] [CrossRef]
- Hanai, J.; Cao, P.; Tanksale, P.; Imamura, S.; Koshimizu, E.; Zhao, J.; Kishi, S.; Yamashita, M.; Phillips, P.S.; Sukhatme, V.P.; et al. The muscle-specific ubiquitin ligase atrogin-1/MAFbx mediates statin-induced muscle toxicity. J. Clin. Invest. 2007, 117, 3940–3951. [Google Scholar] [CrossRef]
- Bialek, P.; Morris, C.; Parkington, J.; St Andre, M.; Owens, J.; Yaworsky, P.; Seeherman, H.; Jelinsky, S.A. Distinct protein degradation profiles are induced by different disuse models of skeletal muscle atrophy. Physiol. Genom. 2011, 43, 1075–1086. [Google Scholar] [CrossRef]
- Bodine, S.C.; Latres, E.; Baumhueter, S.; Lai, V.K.; Nunez, L.; Clarke, B.A.; Poueymirou, W.T.; Panaro, F.J.; Na, E.; Dharmarajan, K.; et al. Identification of ubiquitin ligases required for skeletal muscle atrophy. Science 2001, 294, 1704–1708. [Google Scholar] [CrossRef] [PubMed]
- Collins, C.A.; Gnocchi, V.F.; White, R.B.; Boldrin, L.; Perez-Ruiz, A.; Relaix, F.; Morgan, J.E.; Zammit, P.S. Integrated functions of Pax3 and Pax7 in the regulation of proliferation, cell size and myogenic differentiation. PLoS ONE 2009, 4, e4475. [Google Scholar] [CrossRef] [PubMed]
- Zammit, P.S.; Relaix, F.; Nagata, Y.; Ruiz, A.P.; Collins, C.A.; Partridge, T.A.; Beauchamp, J.R. Pax7 and myogenic progression in skeletal muscle satellite cells. J. Cell Sci. 2006, 119, 1824–1832. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Price, F.; Rudnicki, M.A. Satellite cells and the muscle stem cell niche. Physiol. Rev. 2013, 93, 23–67. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Garry, D.J. Muscle stem cells in development, regeneration, and disease. Genes Dev. 2006, 20, 1692–1708. [Google Scholar] [CrossRef] [PubMed]
- Ganassi, M.; Badodi, S.; Wanders, K.; Zammit, P.S.; Hughes, S.M. Myogenin is an essential regulator of adult myofibre growth and muscle stem cell homeostasis. Elife 2020, 9, e60445. [Google Scholar] [CrossRef]
- von Maltzahn, J.; Jones, A.E.; Parks, R.J.; Rudnicki, M.A. Pax7 is critical for the normal function of satellite cells in adult skeletal muscle. Proc. Natl. Acad. Sci. USA 2013, 110, 16474–16479. [Google Scholar] [CrossRef]
- Macpherson, P.C.; Wang, X.; Goldman, D. Myogenin regulates denervation-dependent muscle atrophy in mouse soleus muscle. J. Cell Biochem. 2011, 112, 2149–2159. [Google Scholar] [CrossRef]
- Abmayr, S.M.; Pavlath, G.K. Myoblast fusion: Lessons from flies and mice. Development 2012, 139, 641–656. [Google Scholar] [CrossRef]
- Steinacker, J.M.; Opitz-Gress, A.; Baur, S.; Lormes, W.; Bolkart, K.; Sunder-Plassmann, L.; Liewald, F.; Lehmann, M.; Liu, Y. Expression of myosin heavy chain isoforms in skeletal muscle of patients with peripheral arterial occlusive disease. J. Vasc. Surg. 2000, 31, 443–449. [Google Scholar] [CrossRef]
- Isesele, P.O.; Mazurak, V.C. Regulation of Skeletal Muscle Satellite Cell Differentiation by Omega-3 Polyunsaturated Fatty Acids: A Critical Review. Front. Physiol. 2021, 12, 682091. [Google Scholar] [CrossRef] [PubMed]
- Viguie, C.A.; Lu, D.X.; Huang, S.K.; Rengen, H.; Carlson, B.M. Quantitative study of the effects of long-term denervation on the extensor digitorum longus muscle of the rat. Anat. Rec. 1997, 248, 346–354. [Google Scholar] [CrossRef]
- Reilly, B.D.; Franklin, C.E. Prevention of muscle wasting and osteoporosis: The value of examining novel animal models. J. Exp. Biol. 2016, 219, 2582–2595. [Google Scholar] [CrossRef] [PubMed]
- Abrams, S.A.; Copeland, K.C.; Gunn, S.K.; Gundberg, C.M.; Klein, K.O.; Ellis, K.J. Calcium absorption, bone mass accumulation, and kinetics increase during early pubertal development in girls. J. Clin. Endocrinol. Metab. 2000, 85, 1805–1809. [Google Scholar] [CrossRef]
- Cifuentes, M.; Morano, A.B.; Chowdhury, H.A.; Shapses, S.A. Energy restriction reduces fractional calcium absorption in mature obese and lean rats. J. Nutr. 2002, 132, 2660–2666. [Google Scholar] [CrossRef]
- Larsson, S.C.; Burgess, S.; Michaëlsson, K. Association of Genetic Variants Related to Serum Calcium Levels with Coronary Artery Disease and Myocardial Infarction. JAMA 2017, 318, 371–380. [Google Scholar] [CrossRef]
- Zheng, H.F.; Forgetta, V.; Hsu, Y.H.; Estrada, K.; Rosello-Diez, A.; Leo, P.J.; Dahia, C.L.; Park-Min, K.H.; Tobias, J.H.; Kooperberg, C.; et al. Whole-genome sequencing identifies EN1 as a determinant of bone density and fracture. Nature 2015, 526, 112–117. [Google Scholar] [CrossRef]
- Nakamura, K.; Saito, T.; Yoshihara, A.; Ishikawa, M.; Tsuchiya, Y.; Oshiki, R.; Kobayashi, R.; Maruyama, K.; Hyodo, K.; Nashimoto, M.; et al. Low calcium intake is associated with increased bone resorption in postmenopausal Japanese women: Yokogoshi Study. Public Health Nutr 2009, 12, 2366–2370. [Google Scholar] [CrossRef]
- Chen, R.A.; Goodman, W.G. Role of the calcium-sensing receptor in parathyroid gland physiology. Am. J. Physiol. Renal. Physiol. 2004, 286, F1005–F1011. [Google Scholar] [CrossRef]
- Watanabe, Y. An assessment of osteoporosis in stroke patients on rehabilitation admission. Int. J. Rehabil. Res. 2004, 27, 163–166. [Google Scholar] [CrossRef]
- Suyama, H.; Kawamoto, M.; Gaus, S.; Yuge, O. Effect of etodolac, a COX-2 inhibitor, on neuropathic pain in a rat model. Brain Res. 2004, 1010, 144–150. [Google Scholar] [CrossRef]
- Kadono, Y.; Okada, F.; Perchonock, C.; Jang, H.D.; Lee, S.Y.; Kim, N.; Choi, Y. Strength of TRAF6 signalling determines osteoclastogenesis. EMBO Rep. 2005, 6, 171–176. [Google Scholar] [CrossRef]
- Oya, A.; Katsuyama, E.; Morita, M.; Sato, Y.; Kobayashi, T.; Miyamoto, K.; Nishiwaki, T.; Funayama, A.; Fujita, Y.; Matsumoto, M.; et al. Tumor necrosis factor receptor-associated factor 6 is required to inhibit foreign body giant cell formation and activate osteoclasts under inflammatory and infectious conditions. J. Bone Miner. Metab. 2018, 36, 679–690. [Google Scholar] [CrossRef] [PubMed]
- Vasikaran, S.; Cooper, C.; Eastell, R.; Griesmacher, A.; Morris, H.A.; Trenti, T.; Kanis, J.A. International Osteoporosis Foundation and International Federation of Clinical Chemistry and Laboratory Medicine position on bone marker standards in osteoporosis. Clin. Chem. Lab. Med. 2011, 49, 1271–1274. [Google Scholar] [CrossRef] [PubMed]
- García-Castellano, J.M.; Díaz-Herrera, P.; Morcuende, J.A. Is bone a target-tissue for the nervous system? New advances on the understanding of their interactions. Iowa Orthop. J. 2000, 20, 49–58. [Google Scholar]
- Ellis, A.; Bennett, D.L. Neuroinflammation and the generation of neuropathic pain. Br. J. Anaesth. 2013, 111, 26–37. [Google Scholar] [CrossRef] [PubMed]
- Amarasekara, D.S.; Yun, H.; Kim, S.; Lee, N.; Kim, H.; Rho, J. Regulation of Osteoclast Differentiation by Cytokine Networks. Immune Netw. 2018, 18, e8. [Google Scholar] [CrossRef]
- Vasikaran, S.; Eastell, R.; Bruyère, O.; Foldes, A.J.; Garnero, P.; Griesmacher, A.; McClung, M.; Morris, H.A.; Silverman, S.; Trenti, T.; et al. Markers of bone turnover for the prediction of fracture risk and monitoring of osteoporosis treatment: A need for international reference standards. Osteoporos. Int. 2011, 22, 391–420. [Google Scholar] [CrossRef]
- Takada, J.; Dinavahi, R.; Miyauchi, A.; Hamaya, E.; Hirama, T.; Libanati, C.; Nakamura, Y.; Milmont, C.E.; Grauer, A. Relationship between P1NP, a biochemical marker of bone turnover, and bone mineral density in patients transitioned from alendronate to romosozumab or teriparatide: A post hoc analysis of the STRUCTURE trial. J. Bone Miner. Metab. 2020, 38, 310–315. [Google Scholar] [CrossRef]
- Zaidi, M.; Pazianas, M.; Shankar, V.S.; Bax, B.E.; Bax, C.M.; Bevis, P.J.; Stevens, C.; Huang, C.L.; Blake, D.R.; Moonga, B.S. Osteoclast function and its control. Exp. Physiol. 1993, 78, 721–739. [Google Scholar] [CrossRef]
- Kawana, K.; Takahashi, M.; Hoshino, H.; Kushida, K. Comparison of serum and urinary C-terminal telopeptide of type I collagen in aging, menopause and osteoporosis. Clin. Chim. Acta 2002, 316, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Jorde, R.; Stunes, A.K.; Kubiak, J.; Grimnes, G.; Thorsby, P.M.; Syversen, U. Smoking and other determinants of bone turnover. PLoS ONE 2019, 14, e0225539. [Google Scholar] [CrossRef] [PubMed]
- Lafage-Proust, M.H.; Roche, B.; Langer, M.; Cleret, D.; Vanden Bossche, A.; Olivier, T.; Vico, L. Assessment of bone vascularization and its role in bone remodeling. Bonekey Rep. 2015, 4, 662. [Google Scholar] [CrossRef]
- Ding, W.G.; Yan, W.H.; Wei, Z.X.; Liu, J.B. Difference in intraosseous blood vessel volume and number in osteoporotic model mice induced by spinal cord injury and sciatic nerve resection. J. Bone Miner. Metab. 2012, 30, 400–407. [Google Scholar] [CrossRef] [PubMed]
- Alagiakrishnan, K.; Juby, A.; Hanley, D.; Tymchak, W.; Sclater, A. Role of vascular factors in osteoporosis. J. Gerontol. A Biol. Sci. Med. Sci. 2003, 58, 362–366. [Google Scholar] [CrossRef]
- Vogt, M.T.; Nevitt, M.C.; Cauley, J.A. Back problems and atherosclerosis. The Study of Osteoporotic Fractures. Spine (Phila Pa 1976) 1997, 22, 2741–2747. [Google Scholar] [CrossRef] [PubMed]
- Gerber, H.P.; Ferrara, N. Angiogenesis and bone growth. Trends Cardiovasc. Med. 2000, 10, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Maes, C.; Kobayashi, T.; Selig, M.K.; Torrekens, S.; Roth, S.I.; Mackem, S.; Carmeliet, G.; Kronenberg, H.M. Osteoblast precursors, but not mature osteoblasts, move into developing and fractured bones along with invading blood vessels. Dev. Cell 2010, 19, 329–344. [Google Scholar] [CrossRef]
- Greene, E.C. Anatomy of the Rat; Hafner Publishing Company: New York, NY, USA, 1963. [Google Scholar]
- Zhang, J.; Zhang, G.; Morrison, B.; Mori, S.; Sheikh, K.A. Magnetic resonance imaging of mouse skeletal muscle to measure denervation atrophy. Exp. Neurol. 2008, 212, 448–457. [Google Scholar] [CrossRef]
- Cizkova, K.; Foltynkova, T.; Gachechiladze, M.; Tauber, Z. Comparative Analysis of Immunohistochemical Staining Intensity Determined by Light Microscopy, ImageJ and QuPath in Placental Hofbauer Cells. Acta Histochem. Cytochem. 2021, 54, 21–29. [Google Scholar] [CrossRef]
- Maurotti, S.; Russo, C.; Musolino, V.; Nucera, S.; Gliozzi, M.; Scicchitano, M.; Bosco, F.; Morittu, V.M.; Ragusa, M.; Mazza, E.; et al. Effects of C-Peptide Replacement Therapy on Bone Microarchitecture Parameters in Streptozotocin-Diabetic Rats. Calcif. Tissue Int. 2020, 107, 266–280. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, J.; Pingoud, A. Real-time polymerase chain reaction. Chembiochem 2003, 4, 1120–1128. [Google Scholar] [CrossRef] [PubMed]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer | Ref. Gene |
|---|---|---|---|
| RANKL | ACTTTCGAGCGCAGATGGAT | GCCTGAAGCAAATGTTGGCG | NM_057149.1 |
| OPG | CCCAACGTTCAACAACCCAA | GGGCGCATAGTCAGTAGACA | NM_012870.2 |
| RUNX2 | CACAAGTGCGGTGCAAACTT | TGAAACTCTTGCCTCGTCCG | NM_001278483.1 |
| ALP | TTGCTAGTGGAAGGAGGCAG | CATTGTGGGCTCTTGTGGGA | NM_013059.2 |
| TRAF-6 | ACTTGATCTCGGAGTGGTGC | CGTGACAGCCAAACACACTG | NM_001107754.2 |
| Β-ACTIN | CCGCGAGTACAACCTTCTTG | CGTCATCCATGGCGAACTGG | NM_031144.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bosco, F.; Guarnieri, L.; Nucera, S.; Scicchitano, M.; Ruga, S.; Cardamone, A.; Maurotti, S.; Russo, C.; Coppoletta, A.R.; Macrì, R.; et al. Pathophysiological Aspects of Muscle Atrophy and Osteopenia Induced by Chronic Constriction Injury (CCI) of the Sciatic Nerve in Rats. Int. J. Mol. Sci. 2023, 24, 3765. https://doi.org/10.3390/ijms24043765
Bosco F, Guarnieri L, Nucera S, Scicchitano M, Ruga S, Cardamone A, Maurotti S, Russo C, Coppoletta AR, Macrì R, et al. Pathophysiological Aspects of Muscle Atrophy and Osteopenia Induced by Chronic Constriction Injury (CCI) of the Sciatic Nerve in Rats. International Journal of Molecular Sciences. 2023; 24(4):3765. https://doi.org/10.3390/ijms24043765
Chicago/Turabian StyleBosco, Francesca, Lorenza Guarnieri, Saverio Nucera, Miriam Scicchitano, Stefano Ruga, Antonio Cardamone, Samantha Maurotti, Cristina Russo, Anna Rita Coppoletta, Roberta Macrì, and et al. 2023. "Pathophysiological Aspects of Muscle Atrophy and Osteopenia Induced by Chronic Constriction Injury (CCI) of the Sciatic Nerve in Rats" International Journal of Molecular Sciences 24, no. 4: 3765. https://doi.org/10.3390/ijms24043765