
Tear Film MicroRNAs as Potential Biomarkers: A Review


Abstract
:1. Background
2. Dry Eye Disease
3. Sjögren’s Syndrome
4. Herpes Epithelial Keratitis
5. Vernal Keratoconjunctivitis
6. Primary Open-Angle Glaucoma
7. Diabetic Macular Edema
8. Diabetic Retinopathy
9. Alzheimer’s Disease
10. Breast Cancer
11. Overall Summary
12. Concluding Remarks and Future Directions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hammond, S.M. An overview of microRNAs. Adv. Drug Deliv. Rev. 2015, 87, 3–14. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of microRNA biogenesis, mechanisms of actions, and circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef]
- Conti, I.; Varano, G.; Simioni, C.; Laface, I.; Milani, D.; Rimondi, E.; Neri, L.M. miRNAs as influencers of cell–cell communication in tumor microenvironment. Cells 2020, 9, 220. [Google Scholar] [CrossRef] [PubMed]
- Lawrie, C.H.; Gal, S.; Dunlop, H.M.; Pushkaran, B.; Liggins, A.P.; Pulford, K.; Banham, A.H.; Pezzella, F.; Boultwood, J.; Wainscoat, J.S. Detection of elevated levels of tumour-associated microRNAs in serum of patients with diffuse large B-cell lymphoma. Br. J. Haematol. 2008, 141, 672–675. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, J.; Sen, S. MicroRNA as biomarkers and diagnostics. J. Cell. Physiol. 2016, 231, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Hrašovec, S.; Glavač, D. MicroRNAs as novel biomarkers in colorectal cancer. Front. Genet. 2012, 3, 180. [Google Scholar] [CrossRef]
- Etheridge, A.; Lee, I.; Hood, L.; Galas, D.; Wang, K. Extracellular microRNA: A new source of biomarkers. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2011, 717, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Van den Berg, M.; Krauskopf, J.; Ramaekers, J.; Kleinjans, J.; Prickaerts, J.; Briedé, J. Circulating microRNAs as potential biomarkers for psychiatric and neurodegenerative disorders. Prog. Neurobiol. 2020, 185, 101732. [Google Scholar] [CrossRef]
- Matias-Garcia, P.R.; Wilson, R.; Mussack, V.; Reischl, E.; Waldenberger, M.; Gieger, C.; Anton, G.; Peters, A.; Kuehn-Steven, A. Impact of long-term storage and freeze-thawing on eight circulating microRNAs in plasma samples. PLoS ONE 2020, 15, e0227648. [Google Scholar] [CrossRef] [PubMed]
- Weber, J.A.; Baxter, D.H.; Zhang, S.; Huang, D.Y.; How Huang, K.; Jen Lee, M.; Galas, D.J.; Wang, K. The microRNA spectrum in 12 body fluids. Clin. Chem. 2010, 56, 1733–1741. [Google Scholar] [CrossRef] [Green Version]
- Xi, X.; Li, T.; Huang, Y.; Sun, J.; Zhu, Y.; Yang, Y.; Lu, Z.J. RNA biomarkers: Frontier of precision medicine for cancer. Non-Coding RNA 2017, 3, 9. [Google Scholar] [CrossRef] [PubMed]
- Chan, H.W.; Yang, B.; Wong, W.; Blakeley, P.; Seah, I.; Tan, Q.S.W.; Wang, H.; Bhargava, M.; Lin, H.A.; Chai, C.H. A pilot study on microRNA profile in tear fluid to predict response to anti-VEGF treatments for diabetic macular edema. J. Clin. Med. 2020, 9, 2920. [Google Scholar] [CrossRef] [PubMed]
- Bachhuber, F.; Huss, A.; Senel, M.; Tumani, H. Diagnostic biomarkers in tear fluid: From sampling to preanalytical processing. Sci. Rep. 2021, 11, 10064. [Google Scholar] [CrossRef] [PubMed]
- Quah, J.H.M.; Tong, L.; Barbier, S. Patient acceptability of tear collection in the primary healthcare setting. Optom. Vis. Sci. 2014, 91, 452. [Google Scholar] [CrossRef]
- Posa, A.; Bräuer, L.; Schicht, M.; Garreis, F.; Beileke, S.; Paulsen, F. Schirmer strip vs. capillary tube method: Non-invasive methods of obtaining proteins from tear fluid. Ann. Anat.-Anat. Anz. 2013, 195, 137–142. [Google Scholar] [CrossRef]
- Kenny, A.; Jiménez-Mateos, E.M.; Zea-Sevilla, M.A.; Rábano, A.; Gili-Manzanaro, P.; Prehn, J.H.; Henshall, D.C.; Ávila, J.; Engel, T.; Hernández, F. Proteins and microRNAs are differentially expressed in tear fluid from patients with Alzheimer’s disease. Sci. Rep. 2019, 9, 15437. [Google Scholar] [CrossRef] [PubMed]
- López-López, M.; Regueiro, U.; Bravo, S.B.; del Pilar Chantada-Vázquez, M.; Varela-Fernández, R.; Ávila-Gómez, P.; Hervella, P.; Lema, I. Tear proteomics in keratoconus: A quantitative SWATH-MS analysis. Investig. Ophthalmol. Vis. Sci. 2021, 62, 30. [Google Scholar] [CrossRef]
- Sun, L.; Liu, X.; Zuo, Z. Regulatory role of miRNA-23a in diabetic retinopathy. Exp. Ther. Med. 2021, 22, 1477. [Google Scholar] [CrossRef]
- Kim, Y.J.; Yeon, Y.; Lee, W.J.; Shin, Y.U.; Cho, H.; Sung, Y.-K.; Lim, H.W.; Kang, M.H. Comparison of MicroRNA expression in tears of normal subjects and Sjögren syndrome patients. Investig. Ophthalmol. Vis. Sci. 2019, 60, 4889–4895. [Google Scholar] [CrossRef] [PubMed]
- Raga-Cervera, J.; Bolarin, J.M.; Millan, J.M.; Garcia-Medina, J.J.; Pedrola, L.; Abellán-Abenza, J.; Valero-Vello, M.; Sanz-González, S.M.; O’Connor, J.E.; Galarreta-Mira, D. miRNAs and genes involved in the interplay between ocular hypertension and primary open-angle glaucoma. Oxidative stress, inflammation, and apoptosis networks. J. Clin. Med. 2021, 10, 2227. [Google Scholar] [CrossRef]
- Wang, Q.; Xie, X.; Li, H.; Hao, S. Discovery of microRNA expression profiles involved in regulating TGF-β 2 expression in the tears of dry eye patients. Ann. Clin. Biochem. 2020, 57, 420–428. [Google Scholar] [CrossRef]
- Kim, Y.J.; Yeon, Y.; Lee, W.J.; Shin, Y.U.; Cho, H.; Lim, H.W.; Kang, M.H. Analysis of MicroRNA Expression in Tears of Patients with Herpes Epithelial Keratitis: A Preliminary Study. Investig. Ophthalmol. Vis. Sci. 2022, 63, 21. [Google Scholar] [CrossRef] [PubMed]
- Dartt, D.; Willcox, M. Complexity of the tear film: Importance in homeostasis and dysfunction during disease. Exp. Eye Res. 2013, 117, 1. [Google Scholar] [CrossRef]
- Hagan, S.; Martin, E.; Enríquez-de-Salamanca, A.J.E.J. Tear fluid biomarkers in ocular and systemic disease: Potential use for predictive, preventive and personalised medicine. EPMA J. 2016, 7, 15. [Google Scholar] [CrossRef]
- Kalmodia, S.; Son, K.-N.; Cao, D.; Lee, B.-S.; Surenkhuu, B.; Shah, D.; Ali, M.; Balasubramaniam, A.; Jain, S.; Aakalu, V.K. Presence of histatin-1 in human tears and association with aqueous deficient dry eye diagnosis: A preliminary study. Sci. Rep. 2019, 9, 10304. [Google Scholar] [CrossRef] [PubMed]
- Pinazo-Durán, M.; Zanón-Moreno, V.; Lleó-Perez, A.; García-Medina, J.; Galbis-Estrada, C.; Roig-Revert, M.; Marco-Ramírez, C.; López-Gálvez, M.; Dolz-Marco, R.; Duarte, L. Genetic systems for a new approach to risk of progression of diabetic retinopathy. Arch. De La Soc. Española De Oftalmol. Engl. Ed. 2016, 91, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Zhang, T.; Ma, H.; Pan, Y.; Wang, S.; Liu, X.; Dai, X.; Zheng, Y.; Lee, L.P.; Liu, F. Discovering the Secret of Diseases by Incorporated Tear Exosomes Analysis via Rapid-Isolation System: iTEARS. ACS Nano 2022, 16, 11720–11732. [Google Scholar] [CrossRef] [PubMed]
- Tamkovich, S.; Grigor’eva, A.; Eremina, A.; Tupikin, A.; Kabilov, M.; Chernykh, V.; Vlassov, V.; Ryabchikova, E. What information can be obtained from the tears of a patient with primary open angle glaucoma? Clin. Chim. Acta 2019, 495, 529–537. [Google Scholar] [CrossRef] [PubMed]
- Pucker, A.D.; Ngo, W.; Postnikoff, C.K.; Fortinberry, H.; Nichols, J.J. Tear Film miRNAs and Their Association With Human Dry Eye Disease. Curr. Eye Res. 2022, 47, 1479–1487. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Feng, J.; Lin, Y.; Reinach, P.S.; Liu, Y.; Xia, X.; Ma, X.; Chen, W.; Zheng, Q. MiR-223 inhibits hyperosmolarity-induced inflammation through downregulating NLRP3 activation in human corneal epithelial cells and dry eye patients. Exp. Eye Res. 2022, 220, 109096. [Google Scholar] [CrossRef] [PubMed]
- Syed, N.H.; Shahidan, W.N.S.; Shatriah, I.; Zunaina, E. MicroRNA Profiling of the Tears of Children with Vernal Keratoconjunctivitis. Front. Genet. 2022, 13, 847168. [Google Scholar] [CrossRef]
- Inubushi, S.; Kawaguchi, H.; Mizumoto, S.; Kunihisa, T.; Baba, M.; Kitayama, Y.; Takeuchi, T.; Hoffman, R.M.; Sasaki, R. Oncogenic miRNAs identified in tear exosomes from metastatic breast cancer patients. Anticancer. Res. 2020, 40, 3091–3096. [Google Scholar] [CrossRef] [PubMed]
- Ravishankar, P.; Daily, A. Tears as the Next Diagnostic Biofluid: A Comparative Study between Ocular Fluid and Blood. Appl. Sci. 2022, 12, 2884. [Google Scholar] [CrossRef]
- Messmer, E.M. The pathophysiology, diagnosis, and treatment of dry eye disease. Dtsch. Ärzteblatt Int. 2015, 112, 71. [Google Scholar] [CrossRef]
- Craig, J.P.; Nichols, K.K.; Akpek, E.K.; Caffery, B.; Dua, H.S.; Joo, C.-K.; Liu, Z.; Nelson, J.D.; Nichols, J.J.; Tsubota, K. TFOS DEWS II definition and classification report. Ocul. Surf. 2017, 15, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Wolffsohn, J.S.; Arita, R.; Chalmers, R.; Djalilian, A.; Dogru, M.; Dumbleton, K.; Gupta, P.K.; Karpecki, P.; Lazreg, S.; Pult, H. TFOS DEWS II diagnostic methodology report. Ocul. Surf. 2017, 15, 539–574. [Google Scholar] [CrossRef]
- Gopalan, V.; Pillai, S.; Ebrahimi, F.; Salajegheh, A.; Lam, T.C.; Le, T.K.; Langsford, N.; Ho, Y.H.; Smith, R.A.; Lam, A.K.Y. Regulation of microRNA-1288 in colorectal cancer: Altered expression and its clinicopathological significance. Mol. Carcinog. 2014, 53, E36–E44. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Zhang, J.-G.; Shi, Y.; Qin, Q.; Liu, Y.; Wang, B.; Tian, K.; Deng, S.-C.; Li, X.; Zhu, S. MiR-130b is a prognostic marker and inhibits cell proliferation and invasion in pancreatic cancer through targeting STAT3. PLoS ONE 2013, 8, e73803. [Google Scholar]
- Krishnan, K.; Steptoe, A.L.; Martin, H.C.; Pattabiraman, D.R.; Nones, K.; Waddell, N.; Mariasegaram, M.; Simpson, P.T.; Lakhani, S.R.; Vlassov, A. miR-139-5p is a regulator of metastatic pathways in breast cancer. RNA 2013, 19, 1767–1780. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Takeshita, F.; Hino, Y.; Fukunaga, S.; Kudo, Y.; Tamaki, A.; Matsunaga, J.; Takahashi, R.-U.; Takata, T.; Shimamoto, A. miR-22 represses cancer progression by inducing cellular senescence. J. Cell Biol. 2011, 193, 409–424. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wang, X.; Liang, H.; Wang, T.; Yan, X.; Cao, M.; Wang, N.; Zhang, S.; Zen, K.; Zhang, C. miR-203 inhibits cell proliferation and migration of lung cancer cells by targeting PKCα. PLoS ONE 2013, 8, e73985. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Jiang, Z.; Guan, X.; Chen, Y.; Tang, Q.; Wang, G.; Wang, X. miR-450b-5p suppresses stemness and the development of chemoresistance by targeting SOX2 in colorectal cancer. DNA Cell Biol. 2016, 35, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Ayaz, L.; Dinç, E. Evaluation of microRNA responses in ARPE-19 cells against the oxidative stress. Cutan. Ocul. Toxicol. 2018, 37, 121–126. [Google Scholar] [CrossRef]
- Seen, S.; Tong, L. Dry eye disease and oxidative stress. Acta Ophthalmol. 2018, 96, e412–e420. [Google Scholar] [CrossRef] [PubMed]
- Ying, H.; Kang, Y.; Zhang, H.; Zhao, D.; Xia, J.; Lu, Z.; Wang, H.; Xu, F.; Shi, L. MiR-127 modulates macrophage polarization and promotes lung inflammation and injury by activating the JNK pathway. J. Immunol. 2015, 194, 1239–1251. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, A.; Nakajima, T.; Azuma, M. Tear miRNA expression analysis reveals miR-203 as a potential regulator of corneal epithelial cells. BMC Ophthalmol. 2021, 21, 377. [Google Scholar] [CrossRef] [PubMed]
- Manicone, A.M.; McGuire, J.K. Matrix metalloproteinases as modulators of inflammation. In Seminars in Cell & Developmental Biology; Academic Press: Cambridge, MA, USA, 2008; pp. 34–41. [Google Scholar]
- Yao, X.; Huang, J.; Zhong, H.; Shen, N.; Faggioni, R.; Fung, M.; Yao, Y. Targeting interleukin-6 in inflammatory autoimmune diseases and cancers. Pharmacol. Ther. 2014, 141, 125–139. [Google Scholar] [CrossRef]
- Hirano, T. IL-6 in inflammation, autoimmunity and cancer. Int. Immunol. 2021, 33, 127–148. [Google Scholar] [CrossRef] [PubMed]
- Shalom-Feuerstein, R.; Serror, L.; De La Forest Divonne, S.; Petit, I.; Aberdam, E.; Camargo, L.; Damour, O.; Vigouroux, C.; Solomon, A.; Gaggioli, C.; et al. Pluripotent stem cell model reveals essential roles for miR-450b-5p and miR-184 in embryonic corneal lineage specification. Stem Cells 2012, 30, 898–909. [Google Scholar] [CrossRef]
- Ramaesh, T.; Collinson, J.M.; Ramaesh, K.; Kaufman, M.H.; West, J.D.; Dhillon, B. Corneal abnormalities in Pax6+/− small eye mice mimic human aniridia-related keratopathy. Investig. Ophthalmol. Vis. Sci. 2003, 44, 1871–1878. [Google Scholar] [CrossRef]
- Li, W.; Chen, Y.-T.; Hayashida, Y.; Blanco, G.; Kheirkah, A.; He, H.; Chen, S.-Y.; Liu, C.-Y.; Tseng, S.C.G. Down-regulation of Pax6 is associated with abnormal differentiation of corneal epithelial cells in severe ocular surface diseases. J. Pathol. 2008, 214, 114–122. [Google Scholar] [CrossRef] [Green Version]
- Patel, R.; Shahane, A. The epidemiology of Sjögren’s syndrome. Clin. Epidemiol. 2014, 6, 247. [Google Scholar] [PubMed]
- Cha, S.; Mona, M.; Lee, K.E.; Kim, D.H.; Han, K. MicroRNAs in autoimmune Sjögren’s syndrome. Genom. Inform. 2018, 16, e19. [Google Scholar] [CrossRef] [PubMed]
- Alevizos, I.; Alexander, S.; Turner, R.J.; Illei, G.G. MicroRNA expression profiles as biomarkers of minor salivary gland inflammation and dysfunction in Sjögren’s syndrome. Arthritis Rheum. 2011, 63, 535–544. [Google Scholar] [CrossRef]
- Chen, C.-Z.; Li, L.; Lodish, H.F.; Bartel, D.P. MicroRNAs modulate hematopoietic lineage differentiation. Science 2004, 303, 83–86. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Lin, Y.; Huang, L.; Hu, J. MiR-223-3p Regulates Autophagy and Inflammation by Targeting ATG16L1 in Fusarium solani–Induced Keratitis. Investig. Ophthalmol. Vis. Sci. 2022, 63, 41. [Google Scholar] [CrossRef] [PubMed]
- Cortes-Troncoso, J.; Jang, S.-I.; Perez, P.; Hidalgo, J.; Ikeuchi, T.; Greenwell-Wild, T.; Warner, B.M.; Moutsopoulos, N.M.; Alevizos, I. T cell exosome–derived miR-142-3p impairs glandular cell function in Sjögren’s syndrome. JCI Insight 2020, 5, e133497. [Google Scholar] [CrossRef]
- Taheri, F.; Ebrahimi, S.O.; Shareef, S.; Reiisi, S. Regulatory and immunomodulatory role of miR-34a in T cell immunity. Life Sci. 2020, 262, 118209. [Google Scholar] [CrossRef]
- Hart, M.; Walch-Rückheim, B.; Friedmann, K.S.; Rheinheimer, S.; Tänzer, T.; Glombitza, B.; Sester, M.; Lenhof, H.-P.; Hoth, M.; Schwarz, E.C. miR-34a: A new player in the regulation of T cell function by modulation of NF-κB signaling. Cell Death Dis. 2019, 10, 46. [Google Scholar] [CrossRef]
- Youinou, P.; Pennec, Y.; Blaschek, M.; Gentric, A.; Jouquan, J.; Lamour, A.; Angelidis, P. Activation of peripheral blood lymphocytes in patients with primary Sjögren’s syndrome. Rheumatol. Int. 1988, 8, 125–130. [Google Scholar] [CrossRef]
- Zhou, H.; Yang, J.; Tian, J.; Wang, S. CD8+ T lymphocytes: Crucial players in sjögren’s syndrome. Front. Immunol. 2021, 11, 602823. [Google Scholar] [CrossRef] [PubMed]
- Mun, J.; Tam, C.; Chan, G.; Kim, J.H.; Evans, D.; Fleiszig, S. MicroRNA-762 is upregulated in human corneal epithelial cells in response to tear fluid and Pseudomonas aeruginosa antigens and negatively regulates the expression of host defense genes encoding RNase7 and ST2. PLoS ONE 2013, 8, e57850. [Google Scholar] [CrossRef] [PubMed]
- Yan, T.; Shen, J.; Chen, J.; Zhao, M.; Guo, H.; Wang, Y. Differential expression of miR-17-92 cluster among varying histological stages of minor salivary gland in patients with primary Sjögren’s syndrome. Clin. Exp. Rheumatol. 2019, 37, 49–54. [Google Scholar]
- Xiao, C.; Srinivasan, L.; Calado, D.P.; Patterson, H.C.; Zhang, B.; Wang, J.; Henderson, J.M.; Kutok, J.L.; Rajewsky, K. Lymphoproliferative disease and autoimmunity in mice with increased miR-17-92 expression in lymphocytes. Nat. Immunol. 2008, 9, 405–414. [Google Scholar] [CrossRef] [PubMed]
- Wang-Renault, S.-F.; Boudaoud, S.; Nocturne, G.; Roche, E.; Sigrist, N.; Daviaud, C.; Tinggaard, A.B.; Renault, V.; Deleuze, J.-F.; Mariette, X. Deregulation of microRNA expression in purified T and B lymphocytes from patients with primary Sjögren’s syndrome. Ann. Rheum. Dis. 2018, 77, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Mackay, F.; Woodcock, S.A.; Lawton, P.; Ambrose, C.; Baetscher, M.; Schneider, P.; Tschopp, J.; Browning, J.L. Mice transgenic for BAFF develop lymphocytic disorders along with autoimmune manifestations. J. Exp. Med. 1999, 190, 1697–1710. [Google Scholar] [CrossRef] [PubMed]
- Kaye, S.; Choudhary, A. Herpes simplex keratitis. Prog. Retin. Eye Res. 2006, 25, 355–380. [Google Scholar] [CrossRef] [PubMed]
- Azher, T.N.; Yin, X.-T.; Tajfirouz, D.; Huang, A.J.; Stuart, P.M. Herpes simplex keratitis: Challenges in diagnosis and clinical management. Clin. Ophthalmol. 2017, 11, 185. [Google Scholar] [CrossRef] [PubMed]
- Bhela, S.; Mulik, S.; Gimenez, F.; Reddy, P.B.; Richardson, R.L.; Varanasi, S.K.; Jaggi, U.; Xu, J.; Lu, P.Y.; Rouse, B.T. Role of miR-155 in the pathogenesis of herpetic stromal keratitis. Am. J. Pathol. 2015, 185, 1073–1084. [Google Scholar] [CrossRef] [PubMed]
- Anand, S.; Majeti, B.K.; Acevedo, L.M.; Murphy, E.A.; Mukthavaram, R.; Scheppke, L.; Huang, M.; Shields, D.J.; Lindquist, J.N.; Lapinski, P.E. MicroRNA-132–mediated loss of p120RasGAP activates the endothelium to facilitate pathological angiogenesis. Nat. Med. 2010, 16, 909–914. [Google Scholar] [CrossRef]
- Suryawanshi, A.; Mulik, S.; Sharma, S.; Reddy, P.B.; Sehrawat, S.; Rouse, B.T. Ocular neovascularization caused by herpes simplex virus type 1 infection results from breakdown of binding betweenendothelial growth factor A and its soluble receptor. J. Immunol. 2011, 186, 3653–3665. [Google Scholar] [CrossRef] [PubMed]
- Yun, H.; Yee, M.B.; Lathrop, K.L.; Kinchington, P.R.; Hendricks, R.L.; Leger, A.J.S. Production of the cytokine VEGF-A by CD4+ T and myeloid cells disrupts the corneal nerve landscape and promotes herpes stromal keratitis. Immunity 2020, 53, 1050–1062.e5. [Google Scholar] [CrossRef] [PubMed]
- Mulik, S.; Xu, J.; Reddy, P.B.; Rajasagi, N.K.; Gimenez, F.; Sharma, S.; Lu, P.Y.; Rouse, B.T. Role of miR-132 in angiogenesis after ocular infection with herpes simplex virus. Am. J. Pathol. 2012, 181, 525–534. [Google Scholar] [CrossRef]
- Muraleedharan, C.K.; McClellan, S.A.; Barrett, R.P.; Li, C.; Montenegro, D.; Carion, T.; Berger, E.; Hazlett, L.D.; Xu, S. Inactivation of the miR-183/96/182 cluster decreases the severity of Pseudomonas aeruginosa-induced keratitis. Investig. Ophthalmol. Vis. Sci. 2016, 57, 1506–1517. [Google Scholar] [CrossRef] [PubMed]
- Coku, A.; McClellan, S.A.; Van Buren, E.; Back, J.B.; Hazlett, L.D.; Xu, S. The miR-183/96/182 cluster regulates the functions of corneal resident macrophages. ImmunoHorizons 2020, 4, 729–744. [Google Scholar] [CrossRef]
- Muraleedharan, C.K.; McClellan, S.A.; Ekanayaka, S.A.; Francis, R.; Zmejkoski, A.; Hazlett, L.D.; Xu, S. The miR-183/96/182 cluster regulates macrophage functions in response to Pseudomonas aeruginosa. J. Innate Immun. 2019, 11, 347–358. [Google Scholar] [CrossRef]
- Boomiraj, H.; Mohankumar, V.; Lalitha, P.; Devarajan, B. Human corneal microRNA expression profile in fungal keratitis. Investig. Ophthalmol. Vis. Sci. 2015, 56, 7939–7946. [Google Scholar] [CrossRef] [PubMed]
- Arenas-Padilla, M.; Mata-Haro, V. Regulation of TLR signaling pathways by microRNAs: Implications in inflammatory diseases. Cent. Eur. J. Immunol. 2018, 43, 482–489. [Google Scholar] [CrossRef] [PubMed]
- Cimmino, A.; Calin, G.A.; Fabbri, M.; Iorio, M.V.; Ferracin, M.; Shimizu, M.; Wojcik, S.E.; Aqeilan, R.I.; Zupo, S.; Dono, M. miR-15 and miR-16 induce apoptosis by targeting BCL2. Proc. Natl. Acad. Sci. USA 2005, 102, 13944–13949. [Google Scholar] [CrossRef]
- Ahluwalia, J.K.; Khan, S.Z.; Soni, K.; Rawat, P.; Gupta, A.; Hariharan, M.; Scaria, V.; Lalwani, M.; Pillai, B.; Mitra, D. Human cellular microRNA hsa-miR-29a interferes with viral nef protein expression and HIV-1 replication. Retrovirology 2008, 5, 117. [Google Scholar] [CrossRef]
- Lobo, A.-M.; Agelidis, A.M.; Shukla, D. Pathogenesis of herpes simplex keratitis: The host cell response and ocular surface sequelae to infection and inflammation. Ocul. Surf. 2019, 17, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Addis, H.; Jeng, B.H. Vernal keratoconjunctivitis. Clin. Ophthalmol. 2018, 12, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arif, A.S.; Aaqil, B.; Siddiqui, A.; Nazneen, Z.; Farooq, U. Corneal complications and visual impairment in vernal keratoconjunctivitis patients. J. Ayub Med. Coll. Abbottabad 2017, 29, 58–60. [Google Scholar] [PubMed]
- Jing, L.; Li, H.; Zhang, T.; Lu, J.; Zhong, L. MicroRNA-4530 suppresses cell proliferation and induces apoptosis by targeting RASA1 in human umbilical vein endothelial cells. Mol. Med. Rep. 2019, 19, 3393–3402. [Google Scholar] [CrossRef]
- Pagliari, M.; Munari, F.; Toffoletto, M.; Lonardi, S.; Chemello, F.; Codolo, G.; Millino, C.; Della Bella, C.; Pacchioni, B.; Vermi, W. Helicobacter pylori affects the antigen presentation activity of macrophages modulating the expression of the immune receptor CD300E through miR-4270. Front. Immunol. 2017, 8, 1288. [Google Scholar] [CrossRef]
- Singh, N.; Diebold, Y.; Sahu, S.K.; Leonardi, A. Epithelial barrier dysfunction in ocular allergy. Allergy 2022, 77, 1360–1372. [Google Scholar] [CrossRef]
- Mantelli, F.; Mauris, J.; Argüeso, P. The ocular surface epithelial barrier and other mechanisms of mucosal protection: From allergy to infectious diseases. Curr. Opin. Allergy Clin. Immunol. 2013, 13, 563–568. [Google Scholar] [CrossRef]
- Lozano, V.; Martín, C.; Blanco, N.; Alcalde, I.; Fernandez-Vega Cueto, L.; Merayo-Lloves, J.; Quirós, L.M. Exosomes Released by Corneal Stromal Cells Show Molecular Alterations in Keratoconus Patients and Induce Different Cellular Behavior. Biomedicines 2022, 10, 2348. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.H.; Fingert, J.H.; Kuehn, M.H.; Alward, W.L. Primary open-angle glaucoma. New Engl. J. Med. 2009, 360, 1113–1124. [Google Scholar] [CrossRef]
- Drewry, M.D.; Challa, P.; Kuchtey, J.G.; Navarro, I.; Helwa, I.; Hu, Y.; Mu, H.; Stamer, W.D.; Kuchtey, R.W.; Liu, Y. Differentially expressed microRNAs in the aqueous humor of patients with exfoliation glaucoma or primary open-angle glaucoma. Hum. Mol. Genet. 2018, 27, 1263–1275. [Google Scholar] [CrossRef]
- Sabirzhanov, B.; Zhao, Z.; Stoica, B.A.; Loane, D.J.; Wu, J.; Borroto, C.; Dorsey, S.G.; Faden, A.I. Downregulation of miR-23a and miR-27a following experimental traumatic brain injury induces neuronal cell death through activation of proapoptotic Bcl-2 proteins. J. Neurosci. 2014, 34, 10055–10071. [Google Scholar] [CrossRef] [PubMed]
- Jayaram, H.; Cepurna, W.O.; Johnson, E.C.; Morrison, J.C. MicroRNA expression in the glaucomatous retina. Investig. Ophthalmol. Vis. Sci. 2015, 56, 7971–7982. [Google Scholar] [CrossRef] [Green Version]
- Ren, C.; Hu, W.; Wei, Q.; Cai, W.; Jin, H.; Yu, D.; Liu, C.; Shen, T.; Zhu, M.; Liang, X. MicroRNA-27a Promotes Oxidative-Induced RPE Cell Death through Targeting FOXO1. BioMed Res. Int. 2021, 2021, 6666506. [Google Scholar] [CrossRef] [PubMed]
- Tabak, S.; Schreiber-Avissar, S.; Beit-Yannai, E. Crosstalk between MicroRNA and oxidative stress in primary open-angle glaucoma. Int. J. Mol. Sci. 2021, 22, 2421. [Google Scholar] [CrossRef] [PubMed]
- Parisi, C.; Napoli, G.; Amadio, S.; Spalloni, A.; Apolloni, S.; Longone, P.; Volonté, C. MicroRNA-125b regulates microglia activation and motor neuron death in ALS. Cell Death Differ. 2016, 23, 531–541. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Cho, K.-S.; Vu, T.; Shen, C.-H.; Kaur, M.; Chen, G.; Mathew, R.; McHam, M.L.; Fazelat, A.; Lashkari, K. Commensal microflora-induced T cell responses mediate progressive neurodegeneration in glaucoma. Nat. Commun. 2018, 9, 3209. [Google Scholar] [CrossRef] [PubMed]
- Cowan, C.; Muraleedharan, C.K.; O’Donnell, J.J.; Singh, P.K.; Lum, H.; Kumar, A.; Xu, S. MicroRNA-146 inhibits thrombin-induced NF-κB activation and subsequent inflammatory responses in human retinal endothelial cells. Investig. Ophthalmol. Vis. Sci. 2014, 55, 4944–4951. [Google Scholar] [CrossRef]
- Zhuang, P.; Muraleedharan, C.K.; Xu, S. Intraocular delivery of miR-146 inhibits diabetes-induced retinal functional defects in diabetic rat model. Investig. Ophthalmol. Vis. Sci. 2017, 58, 1646–1655. [Google Scholar] [CrossRef]
- Comer, B.S.; Camoretti-Mercado, B.; Kogut, P.C.; Halayko, A.J.; Solway, J.; Gerthoffer, W.T. MicroRNA-146a and microRNA-146b expression and anti-inflammatory function in human airway smooth muscle. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2014, 307, L727–L734. [Google Scholar] [CrossRef]
- Boldin, M.P.; Taganov, K.D.; Rao, D.S.; Yang, L.; Zhao, J.L.; Kalwani, M.; Garcia-Flores, Y.; Luong, M.; Devrekanli, A.; Xu, J. miR-146a is a significant brake on autoimmunity, myeloproliferation, and cancer in mice. J. Exp. Med. 2011, 208, 1189–1201. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Dong, L.; Tang, Y.; Li, M.; Zhang, M. MiR-146b protects against the inflammation injury in pediatric pneumonia through MyD88/NF-κB signaling pathway. Infect. Dis. 2020, 52, 23–32. [Google Scholar] [CrossRef]
- Kuchtey, J.; Rezaei, K.A.; Jaru-Ampornpan, P.; Sternberg, P.; Kuchtey, R.W. Multiplex cytokine analysis reveals elevated concentration of interleukin-8 in glaucomatous aqueous humor. Investig. Ophthalmol. Vis. Sci. 2010, 51, 6441–6447. [Google Scholar] [CrossRef] [Green Version]
- Zanon-Moreno, V.; Marco-Ventura, P.; Lleo-Perez, A.; Pons-Vazquez, S.; Garcia-Medina, J.J.; Vinuesa-Silva, I.; Moreno-Nadal, M.A.; Pinazo-Duran, M.D. Oxidative stress in primary open-angle glaucoma. J. Glaucoma 2008, 17, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Baleriola, J.; García-Feijoo, J.; Martínez-de-la-Casa, J.M.; Fernández-Cruz, A.; Enrique, J.; Fernández-Durango, R. Apoptosis in the trabecular meshwork of glaucomatous patients. Mol. Vis. 2008, 14, 1513. [Google Scholar]
- Wang, L.; Wang, X.; Li, Z.; Kou, D.; Zhang, D.; Xu, Z. MiR-126 facilitates apoptosis of retinal ganglion cells in glaucoma rats via VEGF-Notch signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 8635–8641. [Google Scholar]
- Ye, P.; Liu, J.; He, F.; Xu, W.; Yao, K. Hypoxia-induced deregulation of miR-126 and its regulative effect on VEGF and MMP-9 expression. Int. J. Med. Sci. 2014, 11, 17. [Google Scholar] [CrossRef]
- Lang, G.E. Diabetic macular edema. Ophthalmologica 2012, 227, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Fei, X.; Lu, Y.; Xu, B.; Ma, Y.; Wan, H. miRNA-214 suppresses oxidative stress in diabetic nephropathy via the ROS/Akt/mTOR signaling pathway and uncoupling protein 2. Exp. Ther. Med. 2019, 17, 3530–3538. [Google Scholar] [CrossRef] [PubMed]
- Pang, B.; Qiao, L.; Wang, S.; Guo, X.; Xie, Y.; Han, L. MiR-214-3p plays a protective role in diabetic neuropathic rats by regulating Nav1. 3 and TLR4. Cell Biol. Int. 2021, 45, 2294–2303. [Google Scholar] [CrossRef] [PubMed]
- Yao, T.; Zha, D.; Gao, P.; Shui, H.; Wu, X. MiR-874 alleviates renal injury and inflammatory response in diabetic nephropathy through targeting toll-like receptor-4. J. Cell. Physiol. 2019, 234, 871–879. [Google Scholar] [CrossRef] [PubMed]
- Feng, B.; Chakrabarti, S. miR-320 regulates glucose-induced gene expression in diabetes. Int. Sch. Res. Not. 2012, 2012, 549875. [Google Scholar] [CrossRef] [PubMed]
- Aihara, M.; Kubota, N.; Minami, T.; Shirakawa, R.; Sakurai, Y.; Hayashi, T.; Iwamoto, M.; Takamoto, I.; Kubota, T.; Suzuki, R. Association between tear and blood glucose concentrations: Random intercept model adjusted with confounders in tear samples negative for occult blood. J. Diabetes Investig. 2021, 12, 266–276. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Tan, H.-Y.; Feng, Y.-G.; Zhang, C.; Chen, F.; Feng, Y. microRNA-23a in human cancer: Its roles, mechanisms and therapeutic relevance. Cancers 2018, 11, 7. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Mansoor, S.; Sharma, A.; Sapkal, A.; Sheth, J.; Falatoonzadeh, P.; Kuppermann, B.; Kenney, M. Diabetic retinopathy and VEGF. Open Ophthalmol. J. 2013, 7, 4. [Google Scholar] [CrossRef] [Green Version]
- Nättinen, J.; Aapola, U.; Jylhä, A.; Vaajanen, A.; Uusitalo, H. Comparison of capillary and Schirmer strip tear fluid sampling methods using SWATH-MS proteomics approach. Transl. Vis. Sci. Technol. 2020, 9, 16. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Cui, C.; Xu, H. Downregulation of miR-145-5p elevates retinal ganglion cell survival to delay diabetic retinopathy progress by targeting FGF5. Biosci. Biotechnol. Biochem. 2019, 83, 1655–1662. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Li, S.; Yang, Q.; Zhou, Z.; Fu, M.; Yang, X.; Hao, K.; Liu, Y.; Ji, H. MicroRNA-145-5p targeting of TRIM2 mediates the apoptosis of retinal ganglion cells via the PI3K/AKT signaling pathway in glaucoma. J. Gene Med. 2021, 23, e3378. [Google Scholar] [CrossRef]
- Hui, Y.; Yin, Y. MicroRNA-145 attenuates high glucose-induced oxidative stress and inflammation in retinal endothelial cells through regulating TLR4/NF-κB signaling. Life Sci. 2018, 207, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Bresciani, E.; Saletti, C.; Squillace, N.; Rizzi, L.; Molteni, L.; Meanti, R.; Omeljaniuk, R.J.; Biagini, G.; Gori, A.; Locatelli, V. miRNA-218 targets lipin-1 and glucose transporter type 4 genes in 3T3-L1 cells treated with lopinavir/ritonavir. Front. Pharmacol. 2019, 10, 461. [Google Scholar] [CrossRef] [PubMed]
- Plaisance, V.; Abderrahmani, A.; Perret-Menoud, V.; Jacquemin, P.; Lemaigre, F.; Regazzi, R. MicroRNA-9 controls the expression of Granuphilin/Slp4 and the secretory response of insulin-producing cells. J. Biol. Chem. 2006, 281, 26932–26942. [Google Scholar] [CrossRef]
- Al-Muhtaresh, H.A.; Al-Kafaji, G. Evaluation of two-diabetes related microRNAs suitability as earlier blood biomarkers for detecting prediabetes and type 2 diabetes mellitus. J. Clin. Med. 2018, 7, 12. [Google Scholar] [CrossRef]
- Shen, J.; Yang, X.; Xie, B.; Chen, Y.; Swaim, M.; Hackett, S.F.; Campochiaro, P.A. MicroRNAs regulate ocular neovascularization. Mol. Ther. 2008, 16, 1208–1216. [Google Scholar] [CrossRef]
- Wang, S.; Li, K. MicroRNA-96 regulates RGC-5 cell growth through caspase-dependent apoptosis. Int. J. Clin. Exp. Med. 2014, 7, 3694. [Google Scholar]
- Kumar, A.; Sidhu, J.; Goyal, A.; Tsao, J.W.; Svercauski, J. Alzheimer disease (nursing). Health Aff. 2021, 33, 650–657. [Google Scholar]
- Hébert, S.S.; Papadopoulou, A.S.; Smith, P.; Galas, M.-C.; Planel, E.; Silahtaroglu, A.N.; Sergeant, N.; Buée, L.; De Strooper, B. Genetic ablation of Dicer in adult forebrain neurons results in abnormal tau hyperphosphorylation and neurodegeneration. Hum. Mol. Genet. 2010, 19, 3959–3969. [Google Scholar] [CrossRef] [PubMed]
- Kalló, G.; Emri, M.; Varga, Z.; Ujhelyi, B.; Tőzsér, J.; Csutak, A.; Csősz, É. Changes in the chemical barrier composition of tears in Alzheimer’s disease reveal potential tear diagnostic biomarkers. PLoS ONE 2016, 11, e0158000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higaki, S.; Muramatsu, M.; Matsuda, A.; Matsumoto, K.; Satoh, J.-I.; Michikawa, M.; Niida, S. Defensive effect of microRNA-200b/c against amyloid-beta peptide-induced toxicity in Alzheimer’s disease models. PLoS ONE 2018, 13, e0196929. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.-G.; Wang, J.-L.; Li, L.; Wang, P.-C. MicroRNA-384 regulates both amyloid precursor protein and β-secretase expression and is a potential biomarker for Alzheimer’s disease. Int. J. Mol. Med. 2014, 34, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Scully, O.J.; Bay, B.-H.; Yip, G.; Yu, Y. Breast cancer metastasis. Cancer Genom. Proteom. 2012, 9, 311–320. [Google Scholar]
- Lim, B.; Hortobagyi, G.N. Current challenges of metastatic breast cancer. Cancer Metastasis Rev. 2016, 35, 495–514. [Google Scholar] [CrossRef] [PubMed]
- Loh, H.-Y.; Norman, B.P.; Lai, K.-S.; Rahman, N.M.A.N.A.; Alitheen, N.B.M.; Osman, M.A. The regulatory role of microRNAs in breast cancer. Int. J. Mol. Sci. 2019, 20, 4940. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Zhang, W.; Li, B.; Stringer-Reasor, E.; Chu, C.; Sun, L.; Bae, S.; Chen, D.; Wei, S.; Jiao, K. MicroRNA-200c and microRNA-141 are regulated by a FOXP3-KAT2B axis and associated with tumor metastasis in breast cancer. Breast Cancer Res. 2017, 19, 73. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.-X.; Huang, X.-F.; Shao, Q.; Huang, M.-Y.; Deng, L.; Wu, Q.-L.; Zeng, Y.-X.; Shao, J.-Y. MicroRNA miR-21 overexpression in human breast cancer is associated with advanced clinical stage, lymph node metastasis and patient poor prognosis. RNA 2008, 14, 2348–2360. [Google Scholar] [CrossRef] [PubMed]
- Volinia, S.; Calin, G.A.; Liu, C.-G.; Ambs, S.; Cimmino, A.; Petrocca, F.; Visone, R.; Iorio, M.; Roldo, C.; Ferracin, M. A microRNA expression signature of human solid tumors defines cancer gene targets. Proc. Natl. Acad. Sci. USA 2006, 103, 2257–2261. [Google Scholar] [CrossRef]
- Feng, Y.-H.; Tsao, C.-J. Emerging role of microRNA-21 in cancer. Biomed. Rep. 2016, 5, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.A.; Krichevsky, A.M.; Kosik, K.S. MicroRNA-21 is an antiapoptotic factor in human glioblastoma cells. Cancer Res. 2005, 65, 6029–6033. [Google Scholar] [CrossRef]
- Löffler, D.; Brocke-Heidrich, K.; Pfeifer, G.; Stocsits, C.; Hackermüller, J.; Kretzschmar, A.K.; Burger, R.; Gramatzki, M.; Blumert, C.; Bauer, K. Interleukin-6–dependent survival of multiple myeloma cells involves the Stat3-mediated induction of microRNA-21 through a highly conserved enhancer. Blood J. Am. Soc. Hematol. 2007, 110, 1330–1333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, S.; Wu, H.; Wu, F.; Nie, D.; Sheng, S.; Mo, Y.-Y. MicroRNA-21 targets tumor suppressor genes in invasion and metastasis. Cell Res. 2008, 18, 350–359. [Google Scholar] [CrossRef]
- Toiyama, Y.; Hur, K.; Tanaka, K.; Inoue, Y.; Kusunoki, M.; Boland, C.R.; Goel, A. Serum miR-200c is a novel prognostic and metastasis-predictive biomarker in patients with colorectal cancer. Ann. Surg. 2014, 259, 735. [Google Scholar] [CrossRef]
- Jurmeister, S.; Baumann, M.; Balwierz, A.; Keklikoglou, I.; Ward, A.; Uhlmann, S.; Zhang, J.D.; Wiemann, S.; Sahin, Ö. MicroRNA-200c represses migration and invasion of breast cancer cells by targeting actin-regulatory proteins FHOD1 and PPM1F. Mol. Cell. Biol. 2012, 32, 633–651. [Google Scholar] [CrossRef] [Green Version]


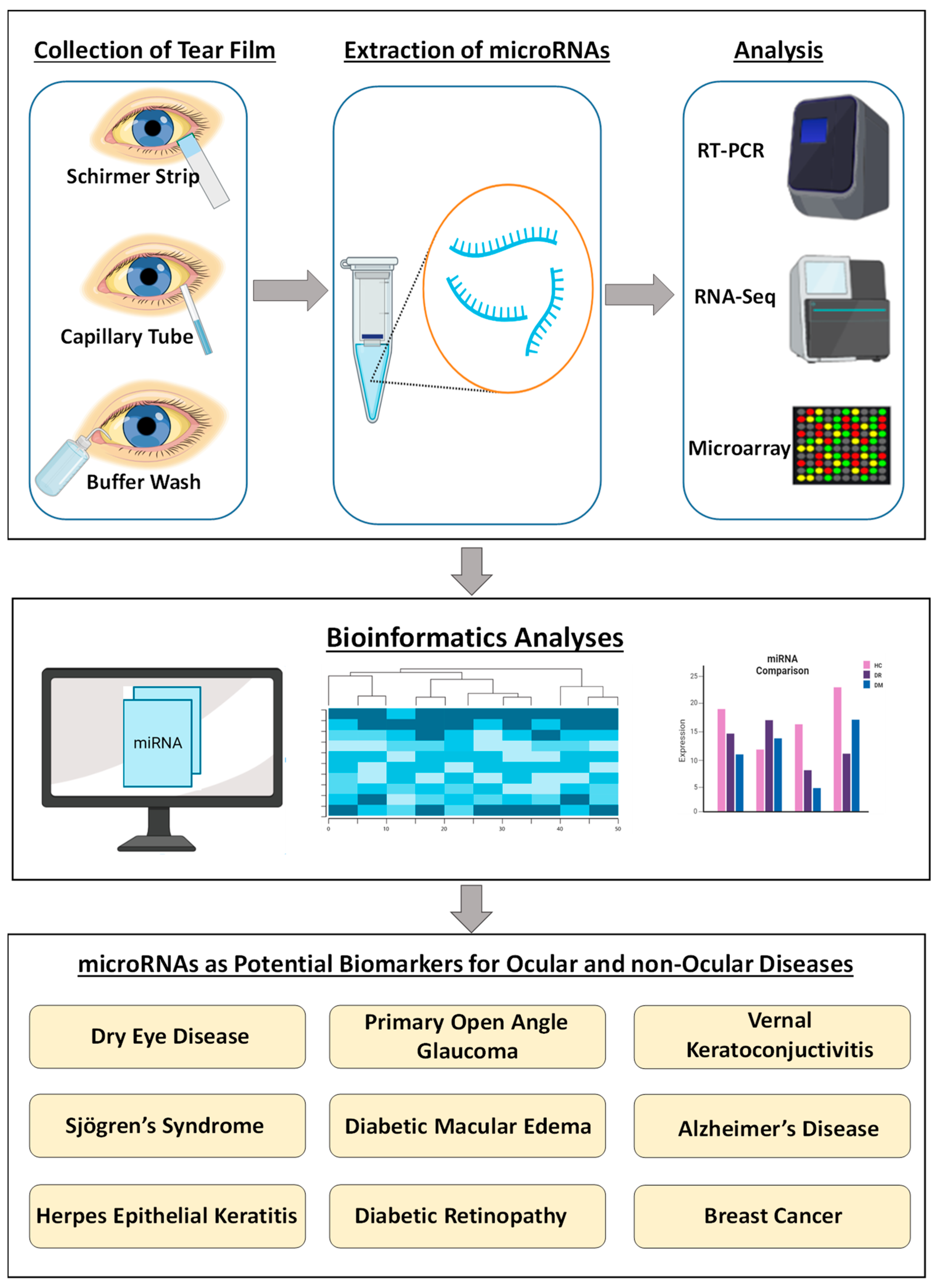{kind=link}
| Study | Disease | Number of Participants | Tear Collection Method | Detection Method |
|---|---|---|---|---|
| Wang, Q. et al. (2020) [21] | Dry eye Disease (DED) | 138 HC; 138 DED | Capillary tube | miRNA microarray |
| Pucker, A.D. et al. (2022) [29] | Dry Eye Disease (DED) | 5 HC; 5 DED | Phosphate buffer saline (PBS) wash | RNA-Seq |
| Kim, Y.J. et al. (2019) [19] | Sjögren Syndrome (SS) | 8 HC; 18 SS | Micropipettes | RT-PCR |
| Kim, Y.J. et al. (2022) [22] | Herpes Epithelial Keratitis (HEK) | 7 HC; 8 HEK | Micropipettes | RT-PCR |
| Syed, N.H. et al. (2022) [31] | Vernal Keratoconjunctivitis (VKC) | 4 HC; 4 VKC | Schirmer strip | miRNA microarray |
| Tamkovich, S. et al. (2019) [28] | Primary Open Angle Glaucoma (POAG) | 29 HC; 33 POAG | Capillary tube | RT-PCR |
| Raga-Cervera, J. et al. (2021) [20] | Primary Open Angle Glaucoma (POAG) | 22 OHT; 20 POAG | Capillary tube | RNA-Seq |
| Chan, H.W. et al. (2020) [12] | Diabetic Macular Edema (DME) | 24 DME | Schirmer strip | RT-PCR |
| Sun, L. et al. (2021) [18] | Diabetic Retinopathy (DR) | 28 HC; 33 DR | Capillary tube | RT-PCR |
| Hu, L. et al. (2022) [27] | Diabetic Retinopathy (DR) | 11 HC; 10 DM; 9 DR | Schirmer strip | RNA-Seq |
| Kenny, A. et al. (2019) [16] | Alzheimer’s Disease (AD) | 15 HC; 8 MCI; 9 AD | Schirmer strip | RT-PCR |
| Inubushi, S. et al. (2020) [32] | Metastatic Breast Cancer (MBC) | 8 HC; 5 MBC | Schirmer strip | RT-PCR |
| Condition | Tear miRNAs in Disease State vs. Controls | |
|---|---|---|
| Upregulated | Downregulated | |
| Dry Eye Disease | miR-127-5p, miR-1273h-3p, miR-1288-5p, miR-130b-5p, miR-139-3p, miR-1910-5p, miR-203b-5p, miR-22-5p, miR-4632-3p, miR-450b-5p, miR-1283, miR-5700, miR-3671 | miR-4673, miR-890, miR-576-5p, miR-337-3p, miR-607, miR-1827, miR-485-5p, miR-6884-5p, miR-335-3p, miR-647, miR-4433b-5p, miR-570-3p, miR-1276, miR-2054, miR-802, miR-539-5p, miR-198, miR-3646, miR-4427, miR-4705, miR-606, miR-20b-3p, miR-4475, miR-4478, miR-4797-5p, miR-142-5p, miR-153-5p, miR-5583-5p |
| Sjögren’s Syndrome | miR-16-5p, miR-34a-5p, miR-142-3p, miR-223-3p | miR-30b-5p, miR-30c-5p, miR-30d-5p, miR-92a-3p, miR-134-5p, miR-137, miR-302d-5p, miR-365b-3p, miR-374c-5p, miR-487b-3p |
| Herpes Epithelial Keratitis | miR-15b-5p, miR-16-5p, miR-20b-5p, miR-21-5p, miR-23b-3p, miR-25-3p, miR-29a-3p, miR-30a-3p, miR-30d-5p, miR-92a-3p, miR-124-3p, miR-127-3p, miR-132-3p, miR-142-3p, miR-145-5p, miR-146a-5p, miR-146b-5p, miR-155-5p, miR-182-5p, miR-183-5p, miR-221-3p, miR-223-3p, miR-338-5p | |
| Vernal Keratoconjunctivitis | miR-1229-5p, miR-6821-5p, miR-6800-5p, miR-4466, miR-3665, miR-4530, miR-7110-5p, miR-1207-5p, miR-6875-5p, miR-762, miR-4741, miR-6740-5p, miR-4298, miR-7107-5p, miR-2861, miR-3663-3p, miR-6891-5p, miR-4672, miR-6785-5p, miR-6510-5p, miR-6803-5p, miR-718, miR-642b-3p, miR-6124, miR-4687-3p, miR-4721, miR-4459, miR-8072, miR-5703, miR-3195, miR-7847-3p, miR-4665-3p, miR-6869-5p, miR-638, miR-6087, miR-4516, miR-3960, miR-6089, miR-4443, miR-5787, miR-642a-3p, miR-3679-5p, miR-6088, miR-8069, miR-4281, miR-1273g-3p, miR-7150, miR-940 | miR-7975, miR-7977, miR-1260a |
| Primary Open-Angle Glaucoma | miR-26b-5p, miR-27a-3p, miR-152-3p, miR-30e-5p, miR-125b-2-5p, miR-224-5p | miR-151a-3p, miR-1307-3p, miR-146b, miR-16, miR-126 |
| Response to Anti-VEGF Treatments for Diabetic Macular Edema | Correlated with good response: miR-214-3p, miR-320d, miR-874-3p, Correlated with poor response: miR-98-5p, miR-196b-5p, miR-454-3p | |
| Diabetic Retinopathy | miR-9-5p, miR-143-3p, miR-411-5p, miR-218-5p, miR-139-5p, miR-10b-5p, miR-129-5p, miR-214-3p, miR-381-3p, miR-130b-4p, miR-382-5p, miR-145-5p, miR-379-5p, miR-127-3p, miR-196a-5p, miR-206, miR-1-3p, miR-199a-5p, miR-642a-3p, miR-642b-5p | miR-23a, miR-361-3p, miR-29c-5p, miR-532-5p, miR-3615, miR-429, miR-224-5p, miR-146a-5p, miR-149-5p, miR-339-3p, miR-203a-3p, miR-24-1-5p, miR-203b-5p, miR-31-5p, miR-147b-3p, miR-205-5p, miR-186-5p, miR-7706, miR-191-5p, miR-135b-5p, miR-96-5p |
| Alzheimer’s Disease | miR-200b-5p | |
| Metastatic Breast Cancer | miR-21, miR-200c | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Altman, J.; Jones, G.; Ahmed, S.; Sharma, S.; Sharma, A. Tear Film MicroRNAs as Potential Biomarkers: A Review. Int. J. Mol. Sci. 2023, 24, 3694. https://doi.org/10.3390/ijms24043694
Altman J, Jones G, Ahmed S, Sharma S, Sharma A. Tear Film MicroRNAs as Potential Biomarkers: A Review. International Journal of Molecular Sciences. 2023; 24(4):3694. https://doi.org/10.3390/ijms24043694
Chicago/Turabian StyleAltman, Jeremy, Garrett Jones, Saleh Ahmed, Shruti Sharma, and Ashok Sharma. 2023. "Tear Film MicroRNAs as Potential Biomarkers: A Review" International Journal of Molecular Sciences 24, no. 4: 3694. https://doi.org/10.3390/ijms24043694