

Culturing the Chicken Intestinal Microbiota and Potential Application as Probiotics Development
 ,
,
Abstract
:1. Introduction
2. Results
2.1. Microbial Diversity of Chicken Intestine
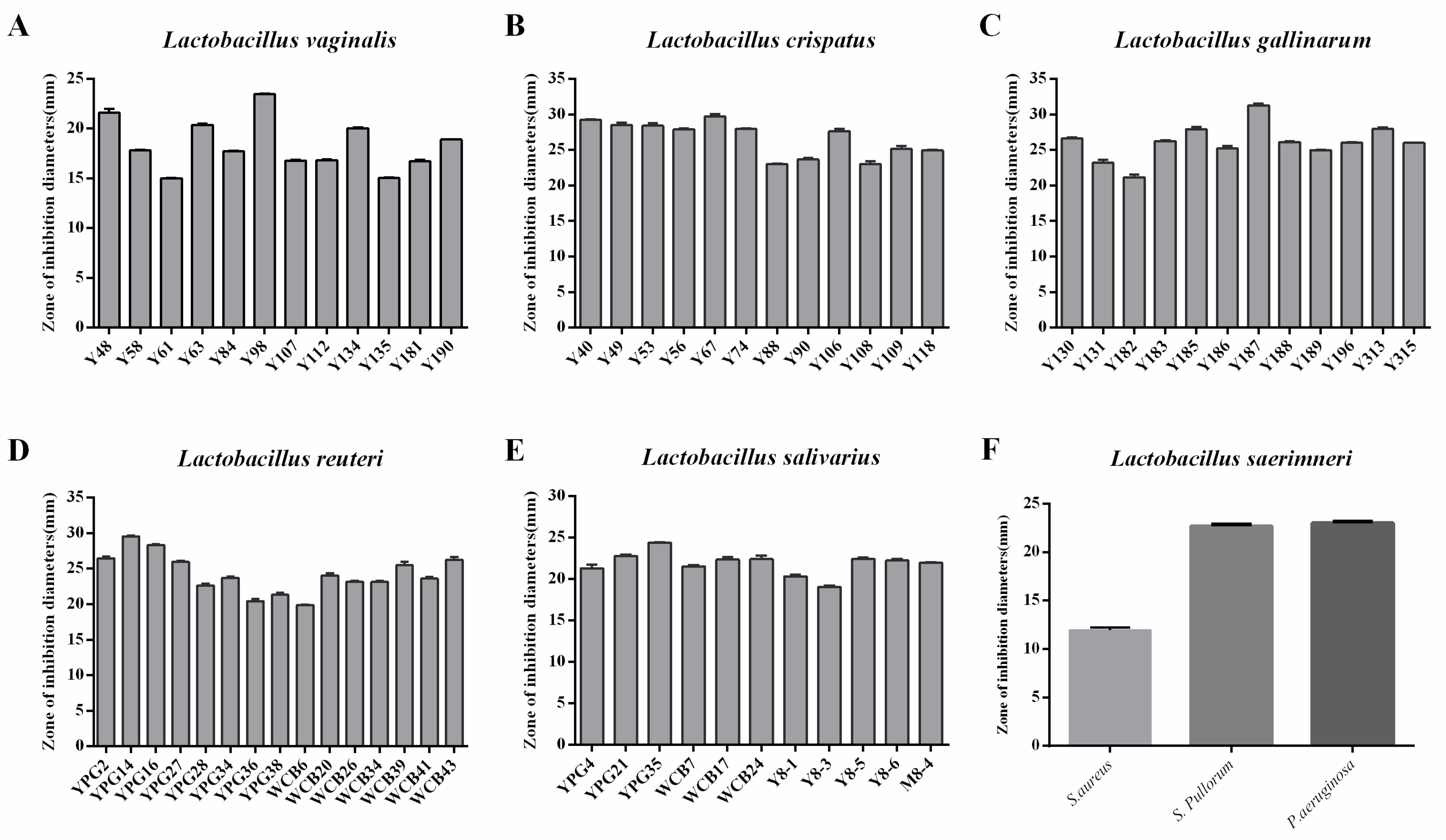
2.2. Selection of Lactobacillus Strains with Antimicrobial Activity against Salmonella Pullorum
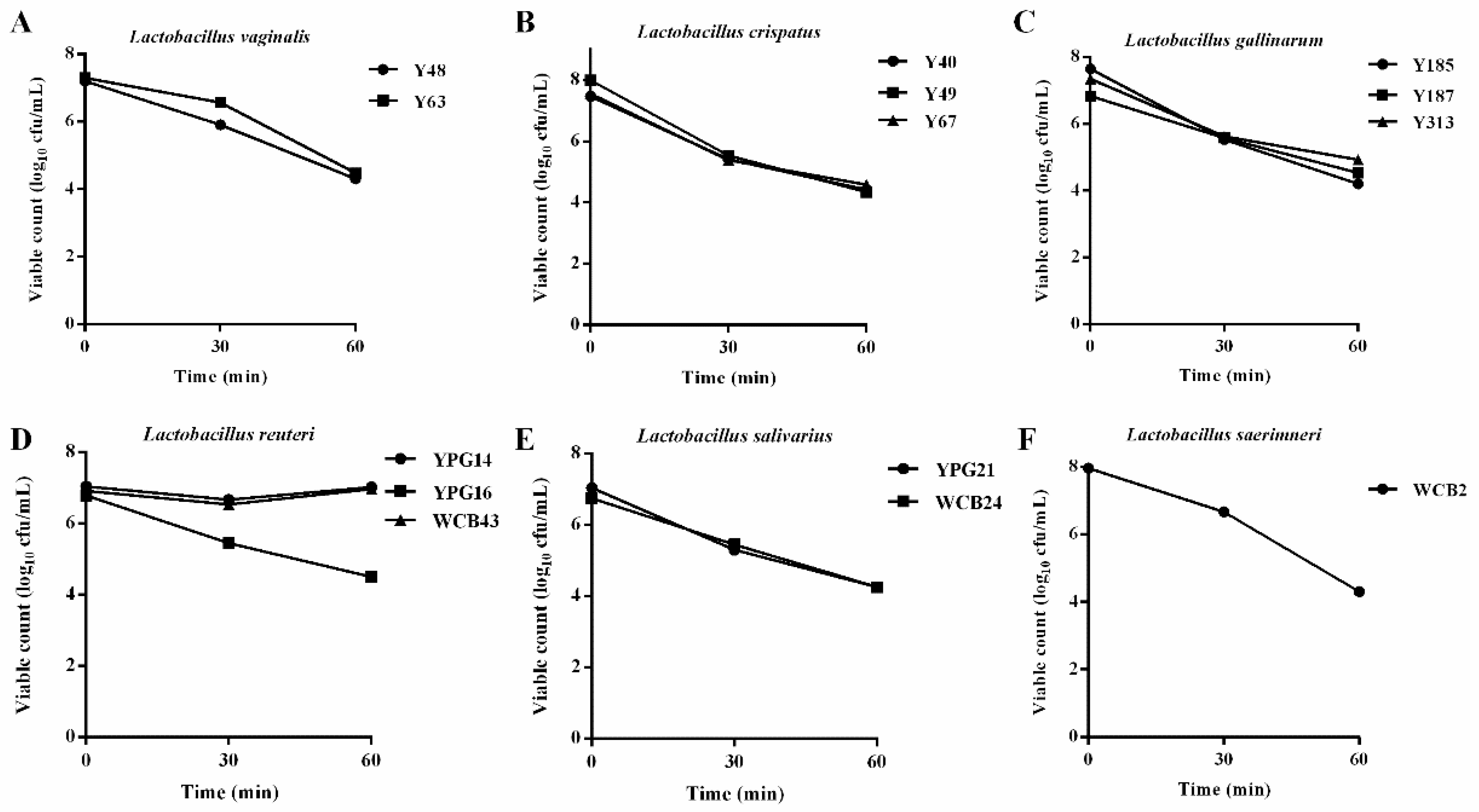
2.3. Acid Tolerance Properties of the Selected Lactobacillus Strains
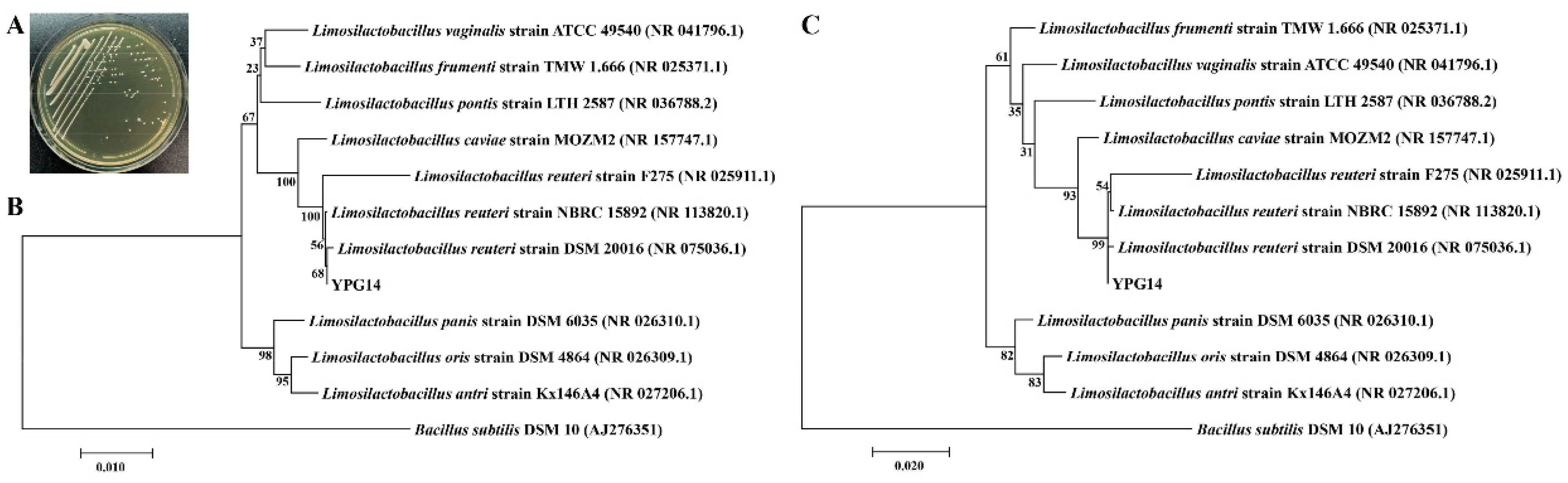
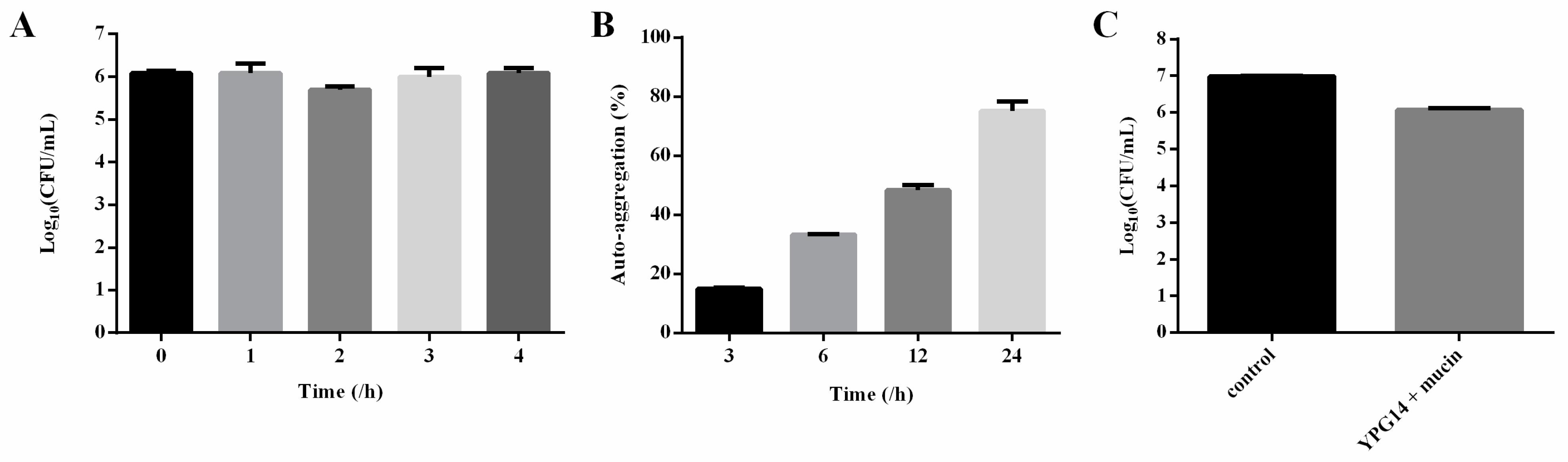
2.4. Identification of the Strain YPG14 and Growth Time Curve
2.5. Simulated Intestinal Fluid Tolerance of the Strain YPG14
2.6. Auto-Aggregation Test of the Strain YPG14
2.7. Mucin Binding Assay of the Strain YPG14

2.8. Antibiotic Sensitivity Testing
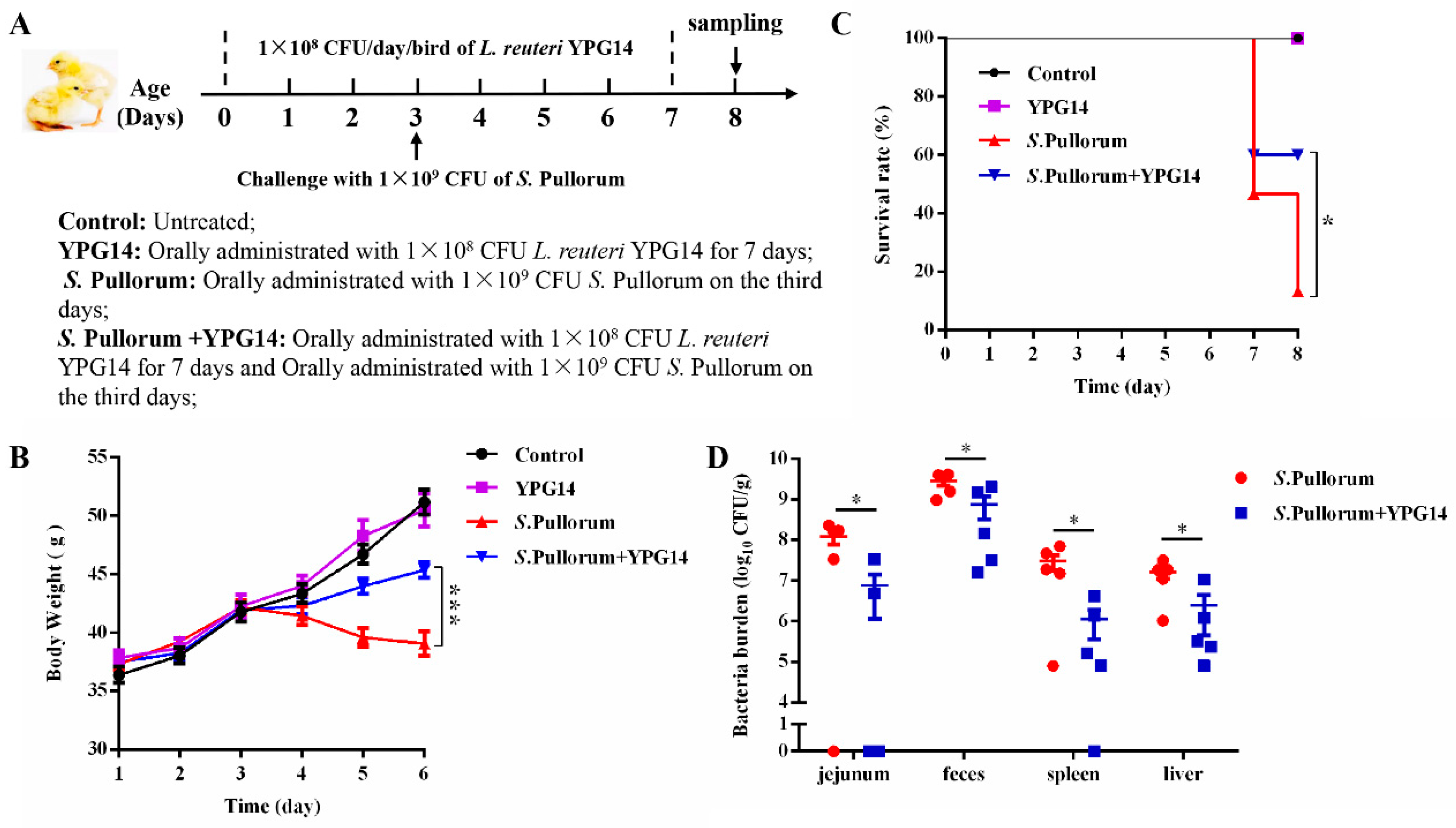
2.9. Survival Rate and Growth Status of Chicks and Enumeration of Salmonella Pullorum
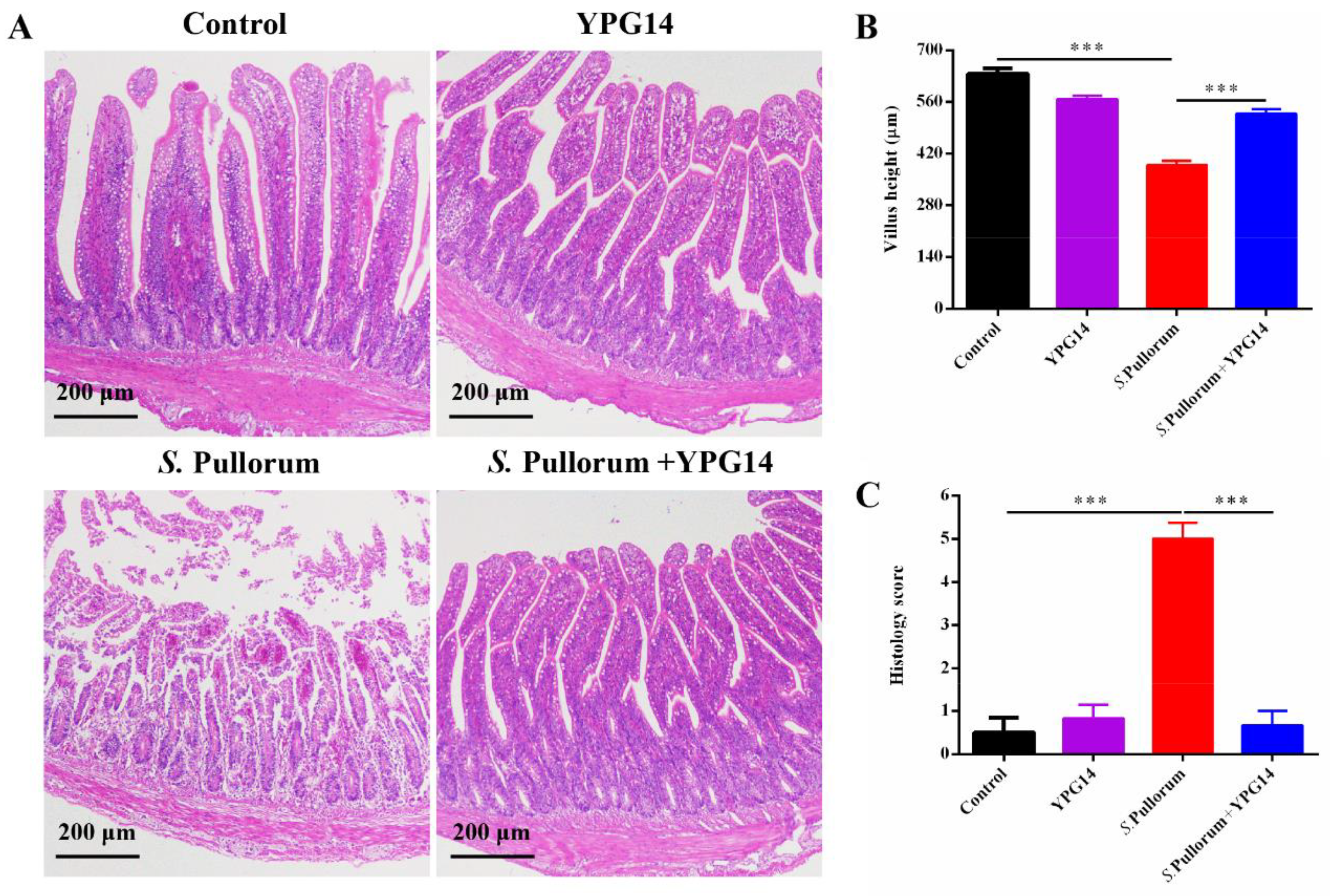
2.10. Histopathology Analysis
3. Discussion
4. Materials and Methods
4.1. Sample Collection and Processing
4.2. Culture Media
4.3. 16S rRNA Gene Sequencing and Phylogenetic Analysis
4.4. Selection of Lactobacillus Strains with Antimicrobial Activity against Salmonella Pullorum
4.5. Acid Tolerance Properties of the Selected Lactobacillus Strains
4.6. Determination of Growth Performance of the Strains YPG14
4.7. Simulated Intestinal Fluid Tolerance of the Strain YPG14
4.8. Auto-Aggregation Test of the Strain YPG14
4.9. Mucin Binding Assay of the Strain YPG14
4.10. Antibiotic Sensitivity Testing
4.11. Chicks’ Management and Experimental Design
4.12. Enumeration of Salmonella Pullorum
4.13. Histopathology Analysis and Scoring
4.14. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Clavijo, V.; Florez, M.J.V. The gastrointestinal microbiome and its association with the control of pathogens in broiler chicken production: A review. Poult. Sci. 2018, 97, 1006–1021. [Google Scholar] [PubMed]
- Zhai, Q.; Feng, S.; Arjan, N.; Chen, W. A next generation probiotic, Akkermansia muciniphila. Crit. Rev. Food Sci. Nutr. 2019, 59, 3227–3236. [Google Scholar] [CrossRef] [PubMed]
- Shine, E.E.; Crawford, J.M. Molecules from the Microbiome. Annu. Rev. Biochem. 2021, 90, 789–815. [Google Scholar] [PubMed]
- Ma, K.; Chen, W.; Yan, S.Q.; Liu, Z.Z.; Lin, X.Q.; Zhang, J.B.; Gao, Y.; Wang, T.; Zhang, J.G.; Yang, Y.J. Purification, Characterization, Mode of Action, and Application of Jileicin, a Novel Antimicrobial from Paenibacillus jilinensis YPG26. J. Agric. Food Chem. 2022, 70, 5570–5578. [Google Scholar] [PubMed]
- Lagier, J.C.; Khelaifia, S.; Alou, M.T.; Ndongo, S.; Dione, N.; Hugon, P.; Caputo, A.; Cadoret, F.; Traore, S.I.; Seck, E.H.; et al. Culture of previously uncultured members of the human gut microbiota by culturomics. Nat. Microbiol. 2016, 1, 16203. [Google Scholar] [CrossRef] [PubMed]
- Lewis, W.H.; Tahon, G.; Geesink, P.; Sousa, D.Z.; Ettema, T.J.G. Innovations to culturing the uncultured microbial majority. Nat. Rev. Microbiol. 2021, 19, 225–240. [Google Scholar]
- Zhang, Z.; Lv, J.; Pan, L.; Zhang, Y. Roles and applications of probiotic Lactobacillus strains. Appl. Microbiol. Biotechnol. 2018, 102, 8135–8143. [Google Scholar]
- Neveling, D.P.; Dicks, L.M.T. Probiotics: An Antibiotic Replacement Strategy for Healthy Broilers and Productive Rearing. Probiot. Antimicrob. Proteins 2021, 13, 1–11. [Google Scholar] [CrossRef]
- Lutful Kabir, S.M. The role of probiotics in the poultry industry. Int. J. Mol. Sci. 2009, 10, 3531–3546. [Google Scholar]
- Stanley, D.; Hughes, R.J.; Moore, R.J. Microbiota of the chicken gastrointestinal tract: Influence on health, productivity and disease. Appl. Microbiol. Biotechnol. 2014, 98, 4301–4310. [Google Scholar] [CrossRef]
- Kim, M.; Oh, H.S.; Park, S.C.; Chun, J. Towards a taxonomic coherence between average nucleotide identity and 16S rRNA gene sequence similarity for species demarcation of prokaryotes. Int. J. Syst. Evol. Microbiol. 2014, 64, 346–351. [Google Scholar] [PubMed]
- Ma, K.; Chen, W.; Yan, S.Q.; Lin, X.Q.; Liu, Z.Z.; Zhang, J.B.; Gao, Y.; Yang, Y.J. Genome mining reveals polysaccharide-degrading potential and new antimicrobial gene clusters of novel intestinal bacterium Paenibacillus jilinensis sp. nov. BMC Genom. 2022, 23, 380. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Xiang, Y.; Zhou, W.; Chen, J.; Li, K.; Yang, H. Microbial community mapping in intestinal tract of broiler chicken. Poult. Sci. 2017, 96, 1387–1393. [Google Scholar] [CrossRef] [PubMed]
- Xiao, S.S.; Mi, J.D.; Mei, L.; Liang, J.; Feng, K.X.; Wu, Y.B.; Liao, X.D.; Wang, Y. Microbial Diversity and Community Variation in the Intestines of Layer Chickens. Animals 2021, 11, 840. [Google Scholar] [CrossRef]
- Ferrario, C.; Alessandri, G.; Mancabelli, L.; Gering, E.; Mangifesta, M.; Milani, C.; Lugli, G.A.; Viappiani, A.; Duranti, S.; Turroni, F.; et al. Untangling the cecal microbiota of feral chickens by culturomic and metagenomic analyses. Environ. Microbiol. 2017, 19, 4771–4783. [Google Scholar]
- Gilroy, R. Spotlight on the avian gut microbiome: Fresh opportunities in discovery. Avian. Pathol. 2021, 50, 291–294. [Google Scholar]
- Pourabedin, M.; Zhao, X. Prebiotics and gut microbiota in chickens. FEMS Microbiol. Lett. 2015, 362, fnv122. [Google Scholar] [CrossRef]
- Dicks, L.M.; Botes, M. Probiotic lactic acid bacteria in the gastro-intestinal tract: Health benefits, safety and mode of action. Benef. Microbes 2010, 1, 11–29. [Google Scholar] [CrossRef]
- Singh, T.P.; Kaur, G.; Malik, R.K.; Schillinger, U.; Guigas, C.; Kapila, S. Characterization of Intestinal Lactobacillus reuteri Strains as Potential Probiotics. Probiotics Antimicrob Proteins 2012, 4, 47–58. [Google Scholar]
- Collado, M.C.; Meriluoto, J.; Salminen, S. Adhesion and aggregation properties of probiotic and pathogen strains. Eur. Food Res. Technol. 2007, 226, 1065–1073. [Google Scholar] [CrossRef]
- Ahire, J.J.; Jakkamsetty, C.; Kashikar, M.S.; Lakshmi, S.G.; Madempudi, R.S. In Vitro Evaluation of Probiotic Properties of Lactobacillus plantarum UBLP40 Isolated from Traditional Indigenous Fermented Food. Probiot. Antimicrob. Proteins 2021, 13, 1413–1424. [Google Scholar]
- El-Saadony, M.T.; Salem, H.M.; El-Tahan, A.M.; Abd El-Mageed, T.A.; Soliman, S.M.; Khafaga, A.F.; Swelum, A.A.; Ahmed, A.E.; Alshammari, F.A.; Abd El-Hack, M.E. The control of poultry salmonellosis using organic agents: An updated overview. Poult. Sci. 2022, 101, 101716. [Google Scholar] [PubMed]
- Zhang, D.; Li, R.; Li, J. Lactobacillus reuteri ATCC 55730 and L22 display probiotic potential in vitro and protect against Salmonella-induced pullorum disease in a chick model of infection. Res. Vet. Sci. 2012, 93, 366–373. [Google Scholar] [PubMed]
- Shanmugasundaram, R.; Mortada, M.; Cosby, D.E.; Singh, M.; Applegate, T.J.; Syed, B.; Pender, C.M.; Curry, S.; Murugesan, G.R.; Selvaraj, R.K. Synbiotic supplementation to decrease Salmonella colonization in the intestine and carcass contamination in broiler birds. PLoS ONE 2019, 14, e0223577. [Google Scholar] [CrossRef] [PubMed]
- Lagkouvardos, I.; Pukall, R.; Abt, B.; Foesel, B.U.; Meier-Kolthoff, J.P.; Kumar, N.; Bresciani, A.; Martinez, I.; Just, S.; Ziegler, C.; et al. The Mouse Intestinal Bacterial Collection (miBC) provides host-specific insight into cultured diversity and functional potential of the gut microbiota. Nat. Microbiol. 2016, 1, 16131. [Google Scholar] [CrossRef]
- Aw, Y.K.; Ong, K.S.; Lee, L.H.; Cheow, Y.L.; Yule, C.M.; Lee, S.M. Newly Isolated Paenibacillus tyrfis sp. nov., from Malaysian Tropical Peat Swamp Soil with Broad Spectrum Antimicrobial Activity. Front. Microbiol. 2016, 7, 219. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Zhao, S.; Han, J.; Bie, X.; Lu, Z.; Zhang, C.; Lv, F. Purification and Characterization of Plantaricin JLA-9: A Novel Bacteriocin against Bacillus spp. Produced by Lactobacillus plantarum JLA-9 from Suan-Tsai, a Traditional Chinese Fermented Cabbage. J. Agric. Food Chem. 2016, 64, 2754–2764. [Google Scholar] [CrossRef]
- Wang, Y.; Li, A.; Jiang, X.; Zhang, H.; Mehmood, K.; Zhang, L.; Jiang, J.; Waqas, M.; Iqbal, M.; Li, J. Probiotic Potential of Leuconostoc pseudomesenteroides and Lactobacillus Strains Isolated From Yaks. Front. Microbiol. 2018, 9, 2987. [Google Scholar]
- Yang, J.M.; Jiang, H.; Dai, H.L.; Wang, Z.W.; Jia, G.Z.; Meng, X.C. Feeble Antipyretic, Analgesic, and Anti-inflammatory Activities were Found with Regular Dose 4′-O-beta-D-Glucosyl-5-O-Methylvisamminol, One of the Conventional Marker Compounds for Quality Evaluation of Radix Saposhnikoviae. Pharmacogn. Mag. 2017, 13, 168–174. [Google Scholar]
- Monteiro, C.; do Carmo, M.S.; Melo, B.O.; Alves, M.S.; Dos Santos, C.I.; Monteiro, S.G.; Bomfim, M.R.Q.; Fernandes, E.S.; Monteiro-Neto, V. In Vitro Antimicrobial Activity and Probiotic Potential of Bifidobacterium and Lactobacillus against Species of Clostridium. Nutrients 2019, 11, 448. [Google Scholar] [PubMed]
- Charteris, W.P.; Kelly, P.M.; Morelli, L.; Collins, J.K. Antibiotic susceptibility of potentially probiotic Lactobacillus species. J. Food Prot. 1998, 61, 1636–1643. [Google Scholar] [CrossRef]
- Xie, S.; Zhang, H.; Matjeke, R.S.; Zhao, J.; Yu, Q. Bacillus coagulans protect against Salmonella enteritidis-induced intestinal mucosal damage in young chickens by inducing the differentiation of goblet cells. Poult. Sci. 2022, 101, 101639. [Google Scholar] [CrossRef]
- Peng, X.; Ed-Dra, A.; Song, Y.; Elbediwi, M.; Nambiar, R.B.; Zhou, X.; Yue, M. Lacticaseibacillus rhamnosus alleviates intestinal inflammation and promotes microbiota-mediated protection against Salmonella fatal infections. Front. Immunol. 2022, 13, 973224. [Google Scholar] [CrossRef] [PubMed]
- De Koning, B.A.; van Dieren, J.M.; Lindenbergh-Kortleve, D.J.; van der Sluis, M.; Matsumoto, T.; Yamaguchi, K.; Einerhand, A.W.; Samsom, J.N.; Pieters, R.; Nieuwenhuis, E.E. Contributions of mucosal immune cells to methotrexate-induced mucositis. Int. Immunol. 2006, 18, 941–949. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Medium | Number of Isolates | Number of Uncultured Isolates | Genus | Species |
|---|---|---|---|---|
| M1 | 19 | 5 | 5 | 9 |
| M2 | 110 | 20 | 6 | 17 |
| M3 | 209 | 63 | 4 | 12 |
| M4 | 27 | 0 | 6 | 10 |
| M5 | 17 | 0 | 2 | 3 |
| M6 | 6 | 0 | 2 | 3 |
| M7 | 24 | 0 | 1 | 1 |
| M8 | 15 | 0 | 2 | 5 |
| M9 | 24 | 0 | 1 | 5 |
| M10 | 35 | 0 | 4 | 7 |
| M11 | 21 | 1 | 4 | 7 |
| Antibiotics | Disc Conc (μg) | Diameter (mm) | Sensitivity/Resistance |
|---|---|---|---|
| Penicillin | 10U | 46 | S |
| Ampicillin | 10 | 38 | S |
| Cefalexin | 30 | 43 | S |
| Cefuroxime | 30 | 34 | S |
| Ceftriaxone | 30 | 44 | S |
| Cefoperazone | 75 | 41 | S |
| Chloramphenicol | 30 | 31 | S |
| Gentamicin | 10 | 12 | R |
| Carbenicillin | 100 | 32 | S |
| Vancomycin | 30 | - | R |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, K.; Chen, W.; Lin, X.-Q.; Liu, Z.-Z.; Wang, T.; Zhang, J.-B.; Zhang, J.-G.; Zhou, C.-K.; Gao, Y.; Du, C.-T.; et al. Culturing the Chicken Intestinal Microbiota and Potential Application as Probiotics Development. Int. J. Mol. Sci. 2023, 24, 3045. https://doi.org/10.3390/ijms24033045
Ma K, Chen W, Lin X-Q, Liu Z-Z, Wang T, Zhang J-B, Zhang J-G, Zhou C-K, Gao Y, Du C-T, et al. Culturing the Chicken Intestinal Microbiota and Potential Application as Probiotics Development. International Journal of Molecular Sciences. 2023; 24(3):3045. https://doi.org/10.3390/ijms24033045
Chicago/Turabian StyleMa, Ke, Wei Chen, Xiao-Qi Lin, Zhen-Zhen Liu, Tao Wang, Jia-Bao Zhang, Jian-Gang Zhang, Cheng-Kai Zhou, Yu Gao, Chong-Tao Du, and et al. 2023. "Culturing the Chicken Intestinal Microbiota and Potential Application as Probiotics Development" International Journal of Molecular Sciences 24, no. 3: 3045. https://doi.org/10.3390/ijms24033045