
Expression of RBMS3 in Breast Cancer Progression
 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results
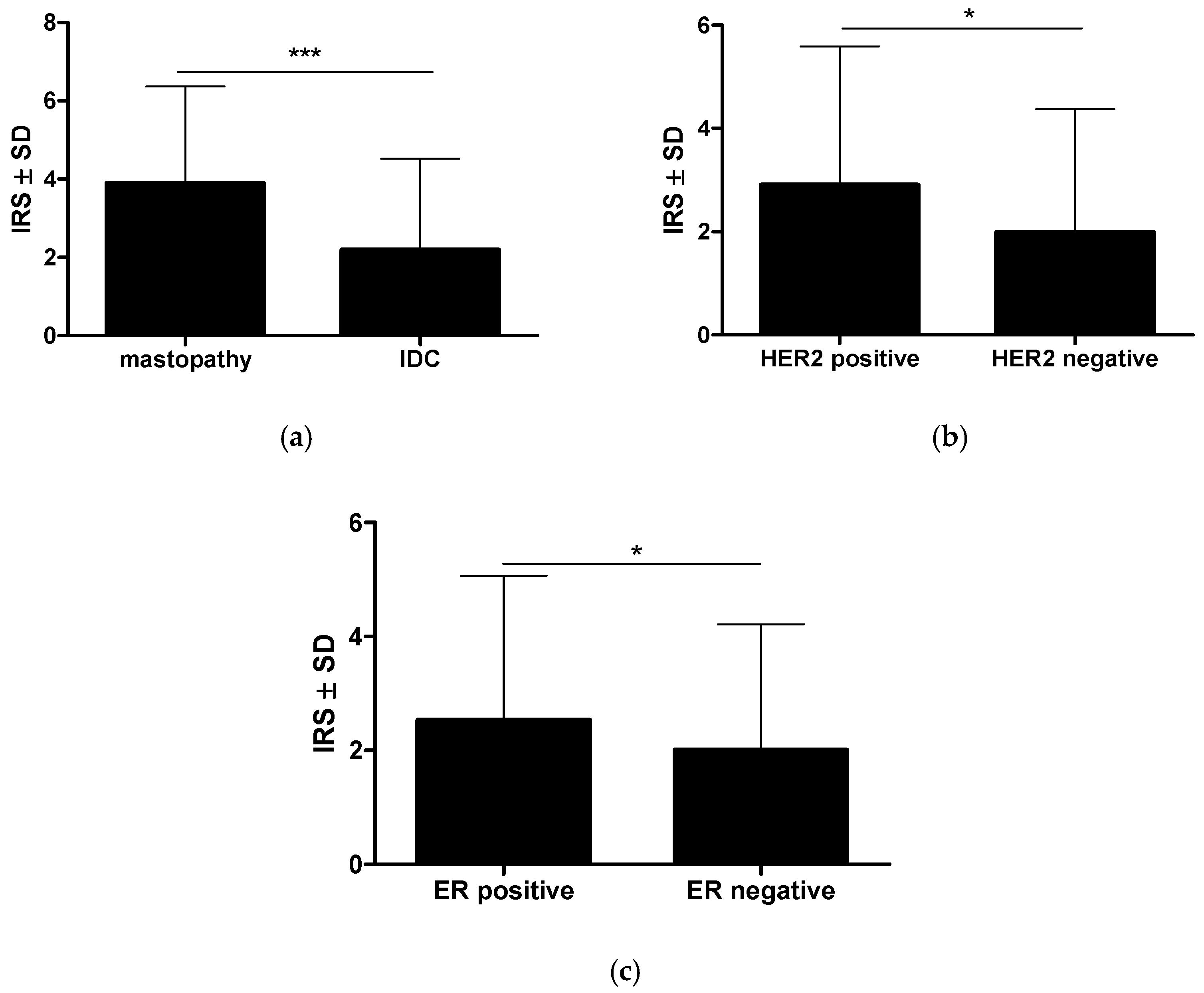
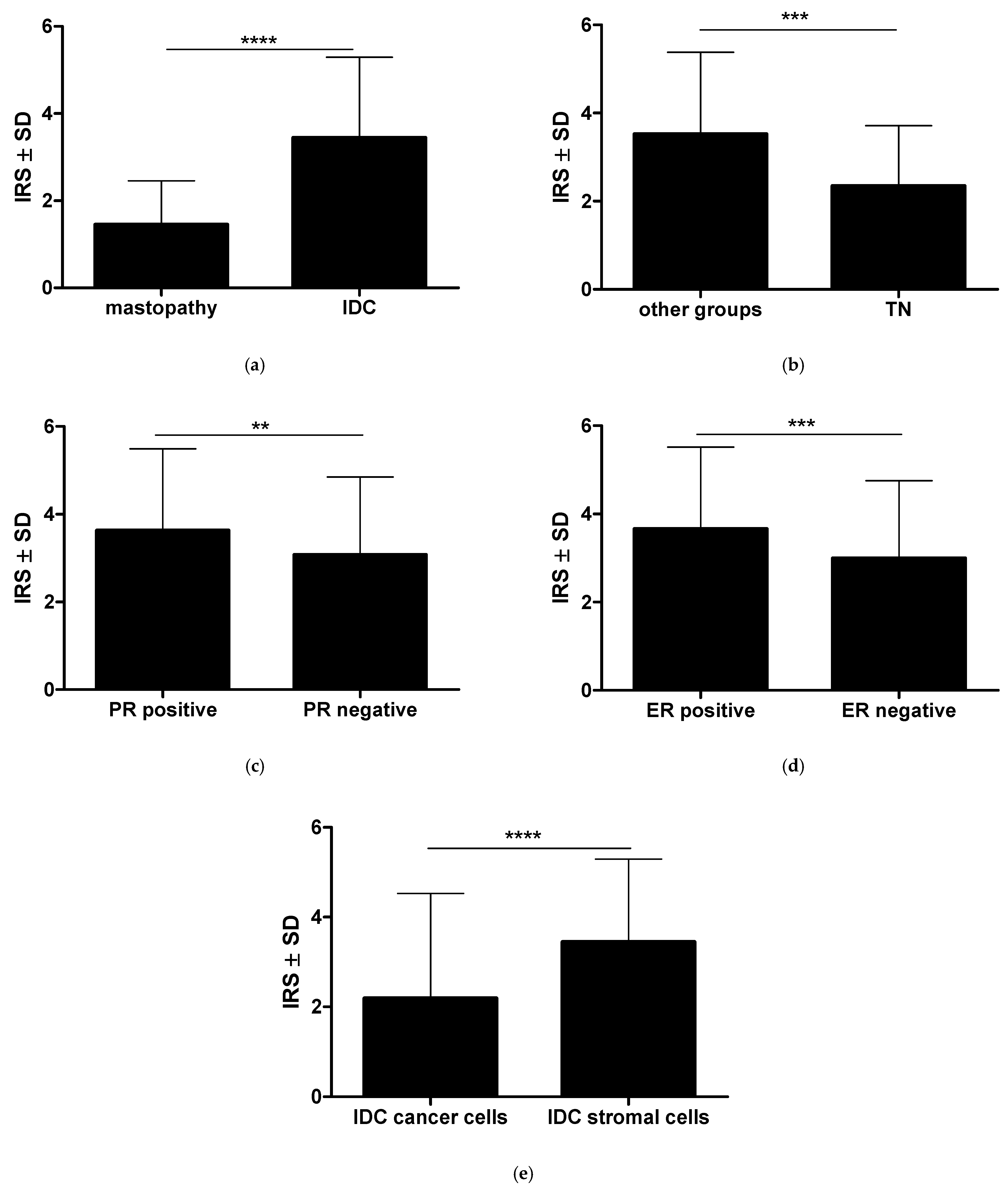
2.1. The Immunohistochemical Intensity of RBMS3′s Expression Varies in Cancer Cells, the Stroma of the Tumor, and the Control Mastopathy Cases, Exhibiting a Dependence on the Expression of Crucial Breast Cancer Receptors
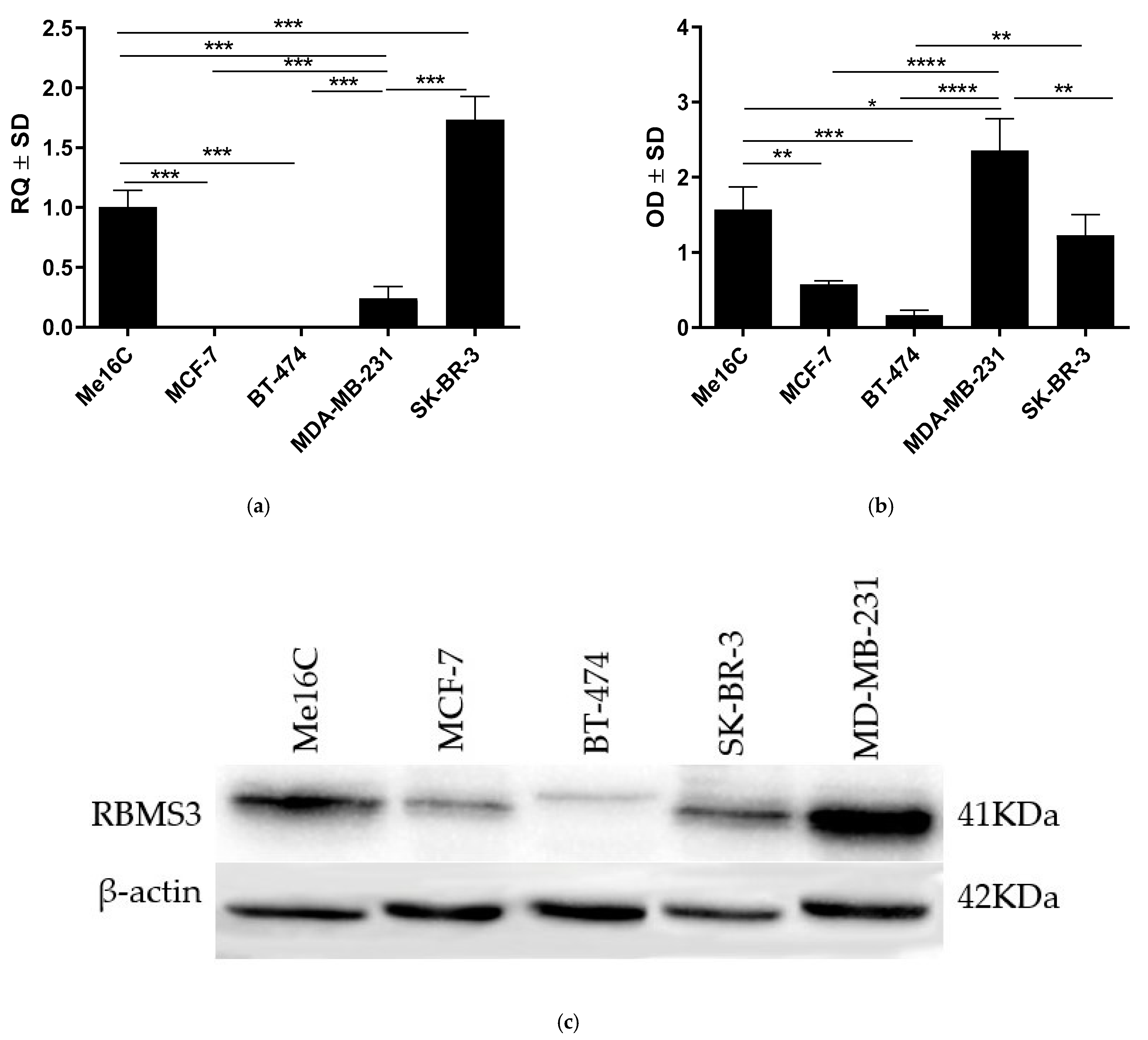
2.2. In Vitro Analysis of RBMS3 Expression Differs from RBMS3 Expression in Clinical Material
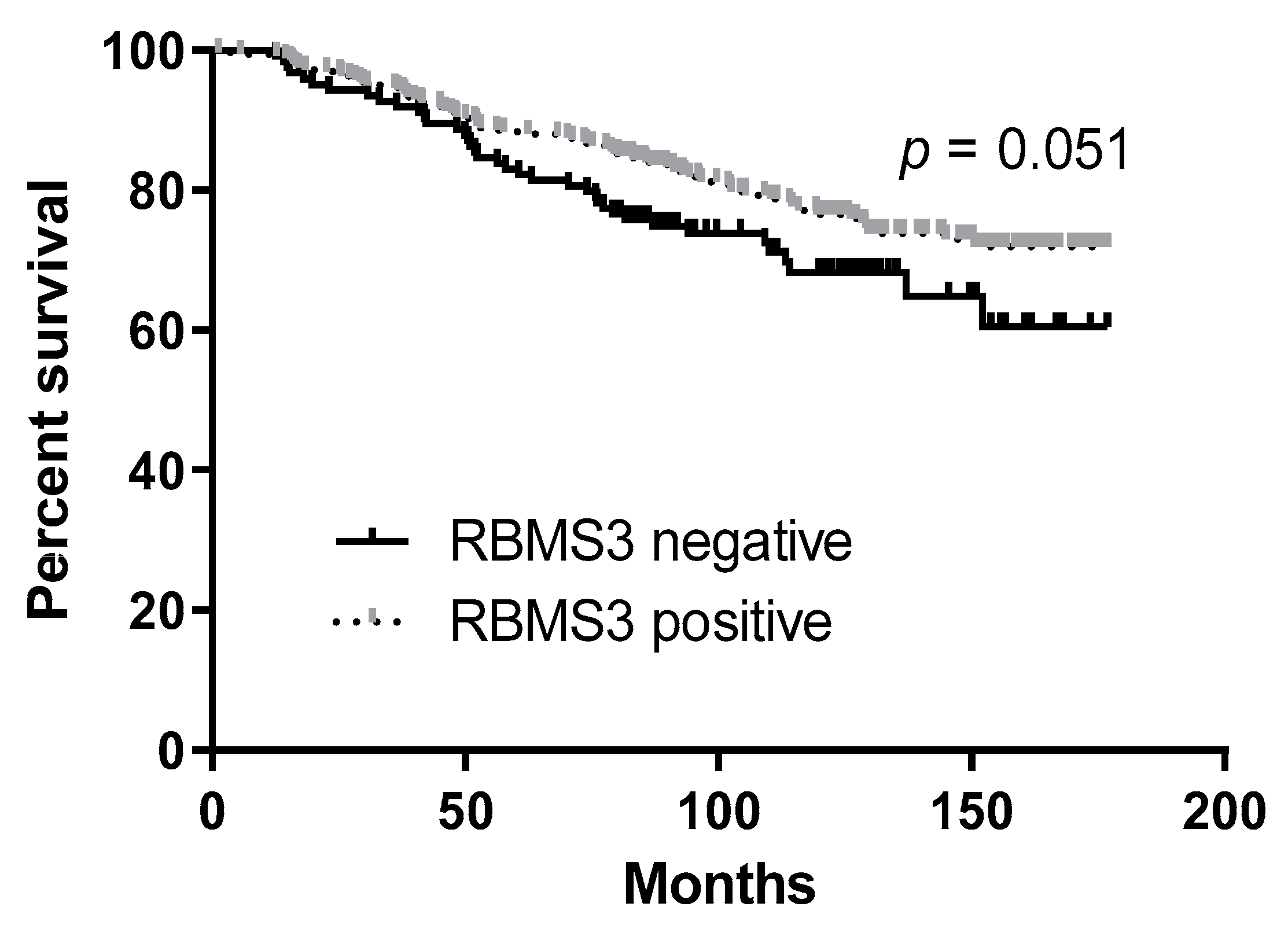
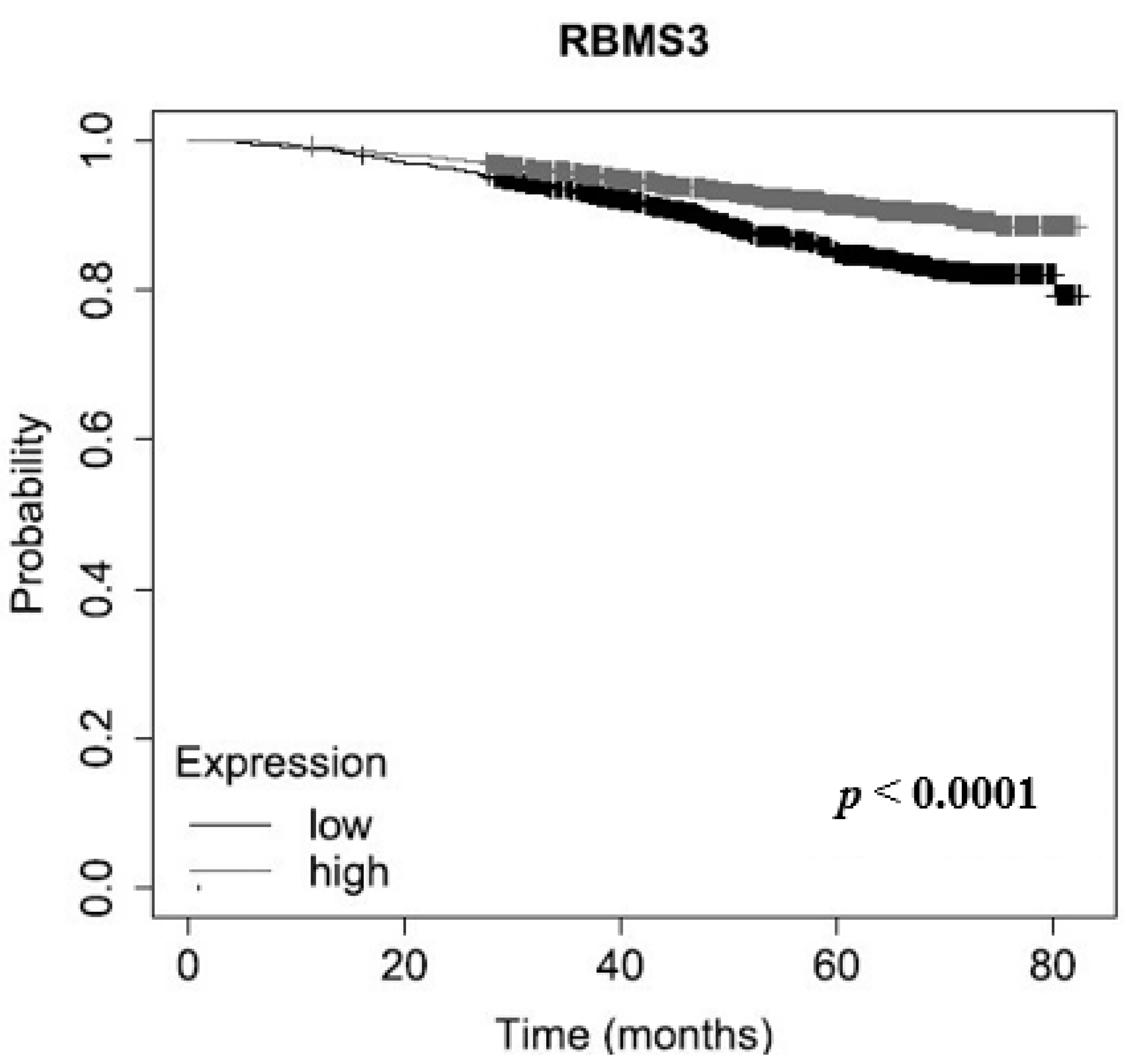
2.3. RBMS3 Expression May Be an Indicator of Longer Overall Survival
3. Discussion
4. Materials and Methods
4.1. Patients’ Characteristics
4.2. Tissue Microarrays (TMAs)
4.3. Immunohistochemistry
4.4. Kaplan–Meier Plotter
4.5. Cell Lines
4.6. Real-Time PCR
4.7. Western Blotting
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Ferlay, J.; Laversanne, M.; Ervik, M.; Lam, F.; Colombet, M.; Mery, L.; Piñeros, M.; Znaor, A.; Soerjomataram, I.; Bray, F. Global Cancer Observatory: Cancer Tomorrow; International Agency for Research on Cancer: Lyon, France, 2018; Available online: https://gco.iarc.fr/tomorrow (accessed on 8 October 2018).
- Łukasiewicz, S.; Czeczelewski, M.; Forma, A.; Baj, J.; Sitarz, R.; Stanisławek, A. Breast Cancer—Epidemiology, Risk Factors, Classification, Prognostic Markers, and Current Treatment Strategies—An Updated Review. Cancers 2021, 13, 4287. [Google Scholar] [CrossRef] [PubMed]
- Tsang, J.Y.S.; Tse, G.M. Molecular Classification of Breast Cancer. Adv. Anat. Pathol. 2020, 27, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Vuong, D.; Simpson, P.; Green, B.; Cummings, M.; Lakhani, S.R. Molecular classification of breast cancer. Virchows Arch. 2014, 465, 1–14. [Google Scholar] [CrossRef]
- Penkov, D.; Ni, R.; Else, C.; Piñol-Roma, S.; Ramirez, F.; Tanaka, S. Cloning of a human gene closely related to the genes coding for the c-myc single-strand binding proteins. Gene 2000, 243, 27–36. [Google Scholar] [CrossRef]
- Niki, T.; Izumi, S.; Saëgusa, Y.; Taira, T.; Takai, T.; Iguchi-Ariga, S.M.M.; Ariga, H. MSSP promotes ras/myc cooperative cell transforming activity by binding to c-Myc. Genes Cells 2000, 5, 127–141. [Google Scholar] [CrossRef]
- Lu, C.-K.; Lai, Y.-C.; Chen, H.-R.; Chiang, M.-K. Rbms3, an RNA-Binding Protein, Mediates the Expression of Ptf1a by Binding to Its 3′UTR During Mouse Pancreas Development. DNA Cell Biol. 2012, 31, 1245–1251. [Google Scholar] [CrossRef]
- Fritz, D.; Stefanovic, B. RNA-binding Protein RBMS3 Is Expressed in Activated Hepatic Stellate Cells and Liver Fibrosis and Increases Expression of Transcription Factor Prx1. J. Mol. Biol. 2007, 371, 585–595. [Google Scholar] [CrossRef]
- Wu, G.; Cao, L.; Zhu, J.; Tan, Z.; Tang, M.; Li, Z.; Hu, Y.; Yu, R.; Zhang, S.; Song, L.; et al. Loss of RBMS3 Confers Platinum Resistance in Epithelial Ovarian Cancer via Activation of miR-126-5p/β-catenin/CBP signaling. Clin. Cancer Res. 2019, 25, 1022–1035. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, Y.; Feng, L.; Li, F.; Sun, Z.; Wu, T.; Shi, X.; Li, J.; Li, X. Comprehensive characterization of lncRNA-mRNA related ceRNA network across 12 major cancers. Oncotarget 2016, 7, 64148–64167. [Google Scholar] [CrossRef] [Green Version]
- Górnicki, T.; Lambrinow, J.; Mrozowska, M.; Podhorska-Okołów, M.; Dzięgiel, P.; Grzegrzółka, J. Role of RBMS3 Novel Potential Regulator of the EMT Phenomenon in Physiological and Pathological Processes. Int. J. Mol. Sci. 2022, 23, 10875. [Google Scholar] [CrossRef]
- Yang, Y.; Quan, L.; Ling, Y. RBMS3 Inhibits the Proliferation and Metastasis of Breast Cancer Cells. Oncol. Res. Featur. Preclin. Clin. Cancer Ther. 2018, 26, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wu, Y.; Liu, Y.; Pan, F.; Zeng, H.; Li, X.; Yu, L. Tumor Suppressor Effect of RBMS3 in Breast Cancer. Technol. Cancer Res. Treat. 2021, 20. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Ma, M.; Li, M.; Guo, Y.; Zuo, X.; Gu, X.; Zhang, M.; Shi, Y. LncRNA MEG3 regulates breast cancer proliferation and apoptosis through miR-141-3p/RBMS3 axis. Genomics 2021, 113, 1689–1704. [Google Scholar] [CrossRef] [PubMed]
- Uddin, N.; Wang, X. Identification of key tumor stroma-associated transcriptional signatures correlated with survival prognosis and tumor progression in breast cancer. Breast Cancer 2022, 29, 541–561. [Google Scholar] [CrossRef] [PubMed]
- Block, C.J.; Mitchell, A.V.; Wu, L.; Glassbrook, J.; Craig, D.; Chen, W.; Dyson, G.; DeGracia, D.; Polin, L.; Ratnam, M.; et al. RNA binding protein RBMS3 is a common EMT effector that modulates triple-negative breast cancer progression via stabilizing PRRX1 mRNA. Oncogene 2021, 40, 6430–6442. [Google Scholar] [CrossRef]
- Zhou, Y.; Liang, Z.; Xia, Y.; Li, S.; Liang, J.; Hu, Z.; Tang, C.; Zhao, Q.; Gong, Q.; Ouyang, Y. Disruption of RBMS3 suppresses PD-L1 and enhances antitumor immune activities and therapeutic effects of auranofin against triple-negative breast cancer. Chem. Biol. Interact. 2023, 369, 110260. [Google Scholar] [CrossRef]
- Zhu, L.; Xi, P.-W.; Li, X.-X.; Sun, X.; Zhou, W.-B.; Xia, T.-S.; Shi, L.; Hu, Y.; Ding, Q.; Wei, J.-F. The RNA binding protein RBMS3 inhibits the metastasis of breast cancer by regulating Twist1 expression. J. Exp. Clin. Cancer Res. 2019, 38, 105, Erratum in J. Exp. Clin. Cancer Res. 2020, 39, 21. [Google Scholar] [CrossRef]
- Grzegrzolka, J.; Biala, M.; Wojtyra, P.; Kobierzycki, C.; Olbromski, M.; Gomulkiewicz, A.; Piotrowska, A.; Rys, J.; Podhorska-Okolow, M.; Dziegiel, P. Expression of EMT Markers SLUG and TWIST in Breast Cancer. Anticancer. Res. 2015, 35, 3961–3968. [Google Scholar]
- Győrffy, B. Survival analysis across the entire transcriptome identifies biomarkers with the highest prognostic power in breast cancer. Comput. Struct. Biotechnol. J. 2021, 19, 4101–4109. [Google Scholar] [CrossRef]
- Wu, Y.; Meng, D.; You, Y.; Sun, R.; Yan, Q.; Bao, J.; Sun, Y.; Yun, D.; Li, Y.; Sun, D. Increased expression of RBMS3 predicts a favorable prognosis in human gallbladder carcinoma. Oncol. Rep. 2020, 44, 55–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, T.; Wu, Y.; Fang, Z.; Yan, Q.; Zhang, S.; Sun, R.; Khaliq, J.; Li, Y. Low expression of RBMS3 and SFRP1 are associated with poor prognosis in patients with gastric cancer. Am. J. Cancer Res. 2016, 6, 2679–2689. [Google Scholar] [PubMed]
- Li, Y.; Chen, L.; Nie, C.-J.; Zeng, T.-T.; Liu, H.; Mao, X.; Qin, Y.; Zhu, Y.-H.; Fu, L.; Guan, X.-Y. Downregulation of RBMS3 Is Associated with Poor Prognosis in Esophageal Squamous Cell Carcinoma. Cancer Res 2011, 71, 6106–6115. [Google Scholar] [CrossRef] [PubMed]
- Weigelt, B.; Geyer, F.C.; Reis-Filho, J.S. Histological types of breast cancer: How special are they? Mol. Oncol. 2010, 4, 192–208. [Google Scholar] [CrossRef]
- Anderson, N.M.; Simon, M.C. The tumor microenvironment. Curr. Biol. 2020, 30, R921–R925. [Google Scholar] [CrossRef]
- Arneth, B. Tumor Microenvironment. Medicina 2019, 56, 15. [Google Scholar] [CrossRef]
- Ratajczak-Wielgomas, K.; Grzegrzolka, J.; Piotrowska, A.; Gomulkiewicz, A.; Witkiewicz, W.; Dziegiel, P. Periostin expression in cancer-associated fibroblasts of invasive ductal breast carcinoma. Oncol. Rep. 2016, 36, 2745–2754. [Google Scholar] [CrossRef]
- Denton, A.E.; Roberts, E.W.; Fearon, D.T. Stromal Cells in the Tumor Microenvironment. Adv. Exp. Med. Biol. 2018, 1060, 99–114. [Google Scholar] [CrossRef]
- Liang, Y.-N.; Liu, Y.; Meng, Q.; Li, X.; Wang, F.; Yao, G.; Wang, L.; Fu, S.; Tong, D. RBMS3 is a tumor suppressor gene that acts as a favorable prognostic marker in lung squamous cell carcinoma. Med. Oncol. 2015, 32, 30. [Google Scholar] [CrossRef]
- Remmele, W.; Stegner, H.E. Vorschlag zur einheitlichen Definition eines Immunreaktiven Score (IRS) für den immunhistochem-ischen Ostrogenrezeptor-Nachweis (ER-ICA) im Mammakarzinomgewebe [Recommendation for uniform definition of an immunoreactive score (IRS) for immunohistochemical estrogen receptor detection (ER-ICA) in breast cancer tissue]. Pathologe 1987, 8, 138–140. (In German) [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecular Subtype of Breast Cancer | Receptor Status | ||
|---|---|---|---|
| Estrogen Receptor (ER) | Progesterone Receptor (PR) | Human Epidermal Growth Factor Receptor 2 (HER2) | |
| Luminal A | + | ≥20% | - |
| Luminal B | + | <20% | +/- |
| HER2-enriched | - | - | + |
| Triple-negative breast cancer (TNBC) | - | - | - |
| Univariate Cox Analysis of Survival | Multivariate Cox Analysis of Survival | |||||||
|---|---|---|---|---|---|---|---|---|
| Characteristics | p-Value | Hazard Ratio | HR 95% CI Lower | HR 95% CI Upper | p-Value | Hazard Ratio | HR 95% CI Lower | HR 95% CI Upper |
| G1 vs. G2-G3 | <0.0100 | 3.0873 | 1.5179 | 6.2792 | <0.0100 | 2.5309 | 1.2509 | 5.1208 |
| pT1 vs. pT2-pT4 | <0.0001 | 2.4469 | 1.7123 | 3.4966 | <0.0010 | 2.0371 | 1.4201 | 2.9221 |
| pN0 vs. pN1-pN3 | <0.0001 | 2.6544 | 1.8541 | 3.8001 | <0.0001 | 2.1583 | 1.4997 | 3.1062 |
| ER negative vs. ER positive | 0.2260 | 0.7987 | 0.5550 | 1.1493 | ||||
| PR negative vs. PR positive | 0.1416 | 0.7626 | 0.5313 | 1.0946 | ||||
| HER2 0-HER2 2 vs. HER2 3 | 0.4485 | 1.3206 | 0.64338 | 2.7105 | ||||
| Triple-negative vs. other groups | 0.3742 | 1.3843 | 0.67566 | 2.8361 | ||||
| RBMS3 IRS stromal: 0 vs. 1–12 | 0.3196 | 0.6548 | 0.2844 | 1.5075 | ||||
| RBMS3 IRS cancer: 0 vs. 1–12 | <0.0500 | 0.6470 | 0.4470 | 0.9365 | 0.1429 | 0.7576 | 0.5226 | 1.0983 |
| Parameters | Patients | |
|---|---|---|
| IHC n = 524 | % | |
| Age | ||
| ≤60 | 165 | 31.49 |
| >60 | 359 | 68.51 |
| Tumor grade | ||
| G1 | 87 | 16.60 |
| G2 | 342 | 65.27 |
| G3 | 92 | 17.56 |
| No data | 3 | 0.57 |
| Tumor size | ||
| pT1 | 325 | 62.02 |
| pT2 | 168 | 32.06 |
| pT3 | 3 | 0.57 |
| pT4 | 9 | 1.72 |
| No data | 19 | 3.63 |
| Lymph nodes | ||
| pN0 | 314 | 59.92 |
| pN1-pN3 | 180 | 34.35 |
| pNx | 30 | 5.73 |
| Stage | ||
| I | 224 | 42.75 |
| II | 257 | 49.05 |
| III | 18 | 3.44 |
| IV | 0 | 0.00 |
| ER | ||
| Neg. | 177 | 33.78 |
| Pos. | 344 | 65.65 |
| No data | 3 | 0.57 |
| PR | ||
| Neg. | 183 | 34.92 |
| Pos. | 338 | 64.50 |
| No data | 3 | 0.57 |
| HER2 | ||
| Neg. | 272 | 51.91 |
| Pos. | 36 | 6.87 |
| No data | 216 | 41.22 |
| Molecular tumor types | ||
| Triple-negative | 34 | 6.49 |
| Other types | 487 | 92.94 |
| No data | 3 | 0.57 |
| Points | Percentage of Positive Cancer Cells (A) | Intensity of Color Reaction (B) |
|---|---|---|
| 0 | 0% | No color reaction |
| 1 | <10% | Mild reaction |
| 2 | 10–50% | Moderate reaction |
| 3 | 51–80% | Strong reaction |
| 4 | 81–100% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Górnicki, T.; Lambrinow, J.; Mrozowska, M.; Romanowicz, H.; Smolarz, B.; Piotrowska, A.; Gomułkiewicz, A.; Podhorska-Okołów, M.; Dzięgiel, P.; Grzegrzółka, J. Expression of RBMS3 in Breast Cancer Progression. Int. J. Mol. Sci. 2023, 24, 2866. https://doi.org/10.3390/ijms24032866
Górnicki T, Lambrinow J, Mrozowska M, Romanowicz H, Smolarz B, Piotrowska A, Gomułkiewicz A, Podhorska-Okołów M, Dzięgiel P, Grzegrzółka J. Expression of RBMS3 in Breast Cancer Progression. International Journal of Molecular Sciences. 2023; 24(3):2866. https://doi.org/10.3390/ijms24032866
Chicago/Turabian StyleGórnicki, Tomasz, Jakub Lambrinow, Monika Mrozowska, Hanna Romanowicz, Beata Smolarz, Aleksandra Piotrowska, Agnieszka Gomułkiewicz, Marzena Podhorska-Okołów, Piotr Dzięgiel, and Jędrzej Grzegrzółka. 2023. "Expression of RBMS3 in Breast Cancer Progression" International Journal of Molecular Sciences 24, no. 3: 2866. https://doi.org/10.3390/ijms24032866