
Mutations in Complex I of the Mitochondrial Electron-Transport Chain Sensitize the Fruit Fly (Drosophila melanogaster) to Ether and Non-Ether Volatile Anesthetics
 ,
,  ,
,
Abstract
:1. Introduction
2. Results
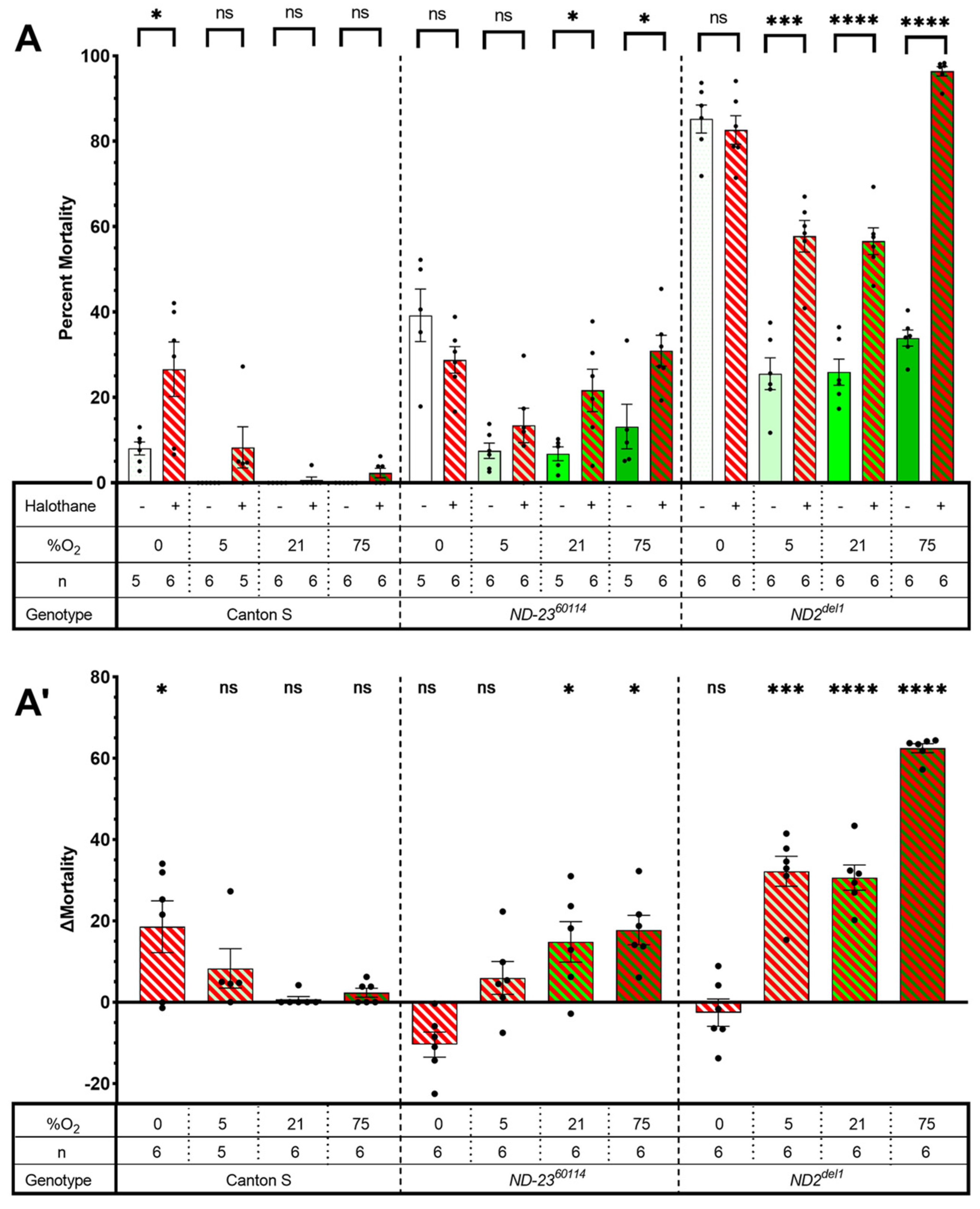
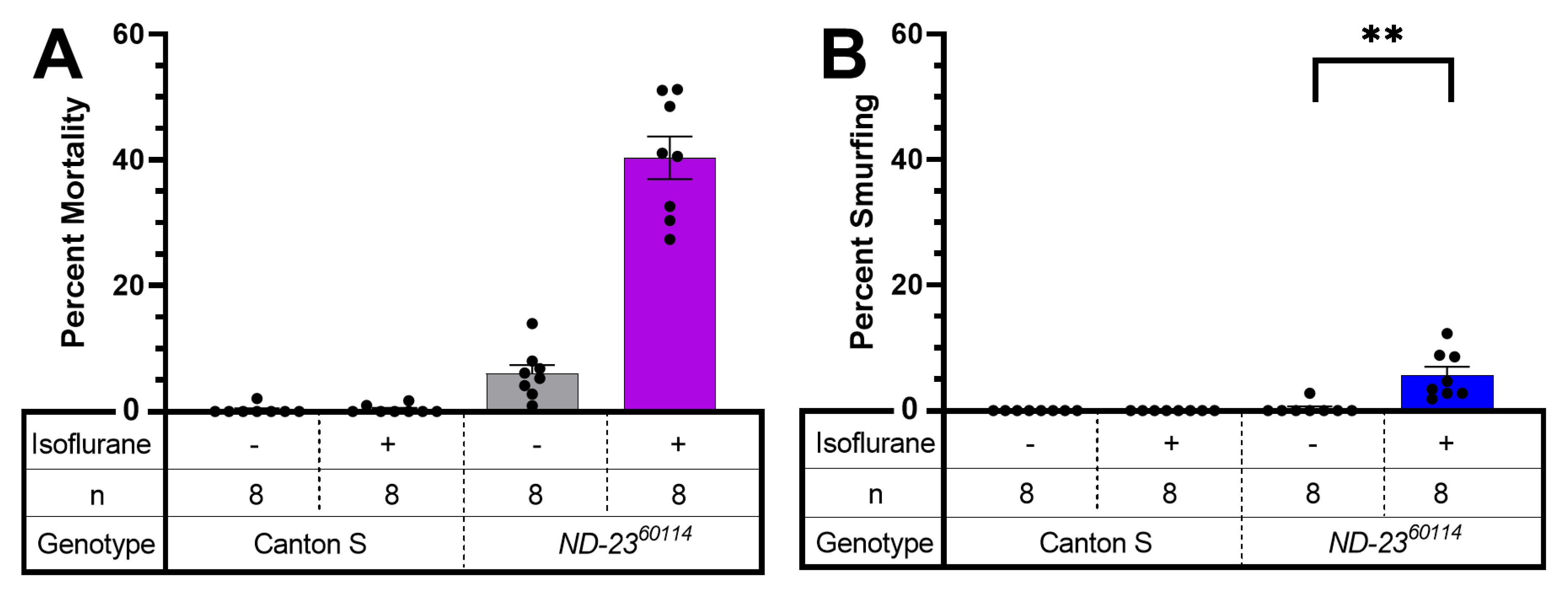
2.1. Mutants of Complex I Subunits ND-23 and Mt:ND2 Are Sensitive to Halothane Toxicity
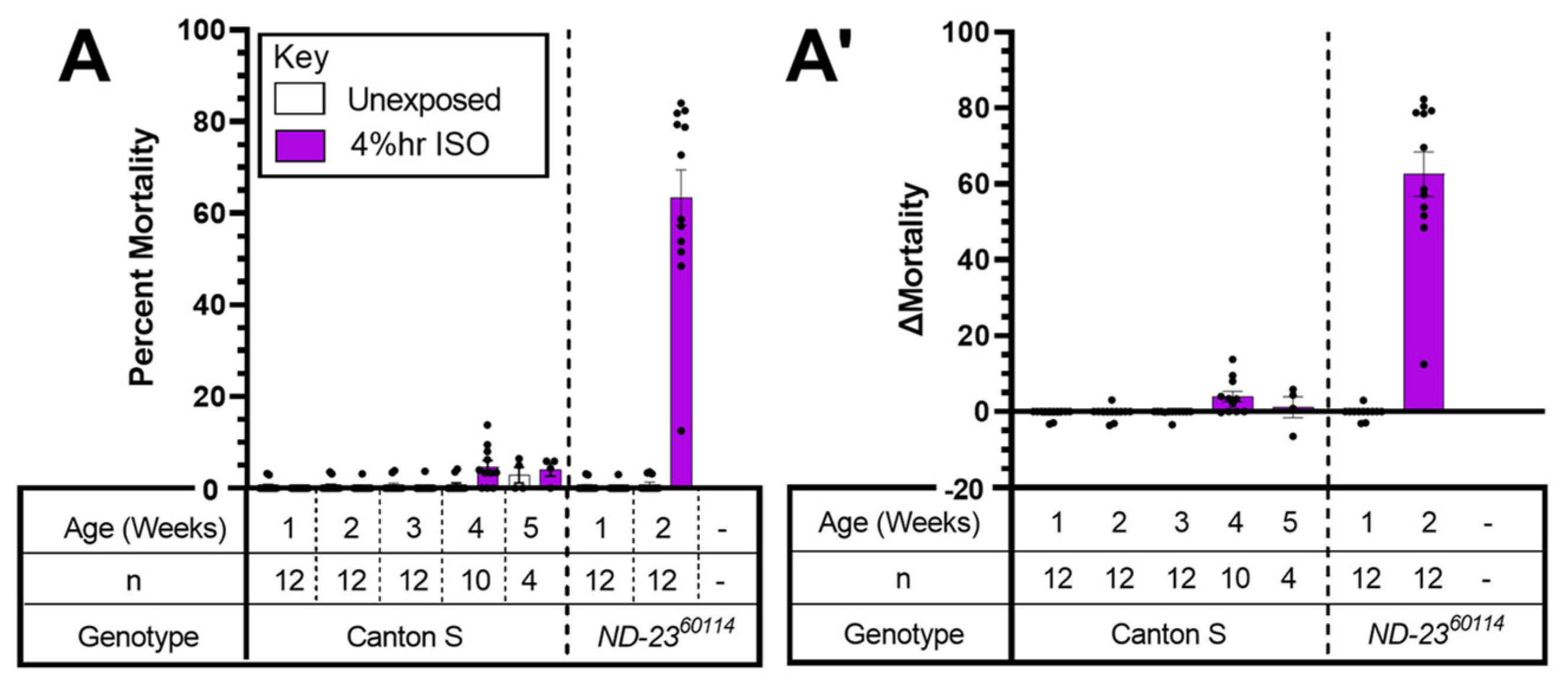
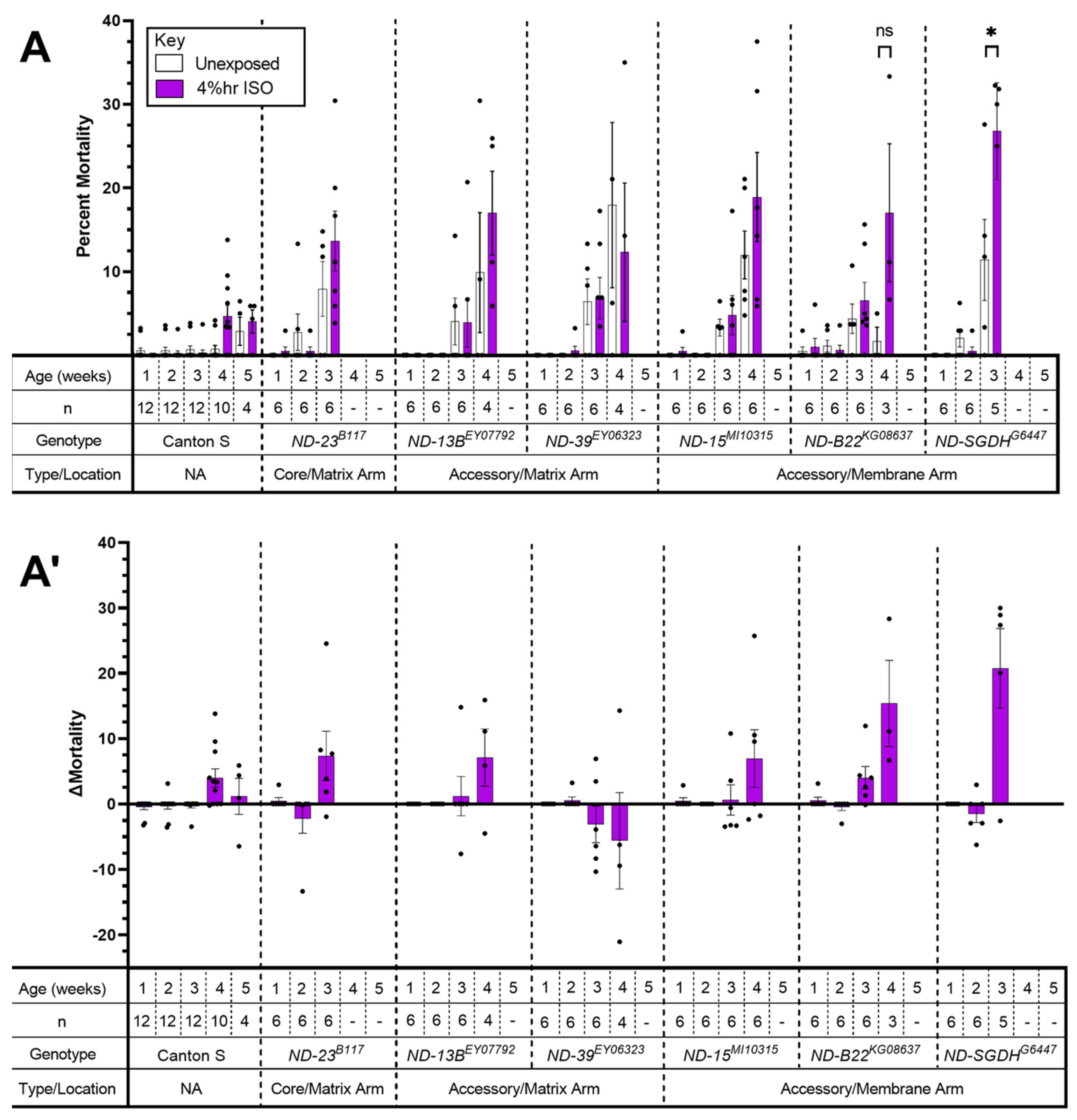
2.2. Complex I: A Mutant Allele of ND-SGDH Results in AiN
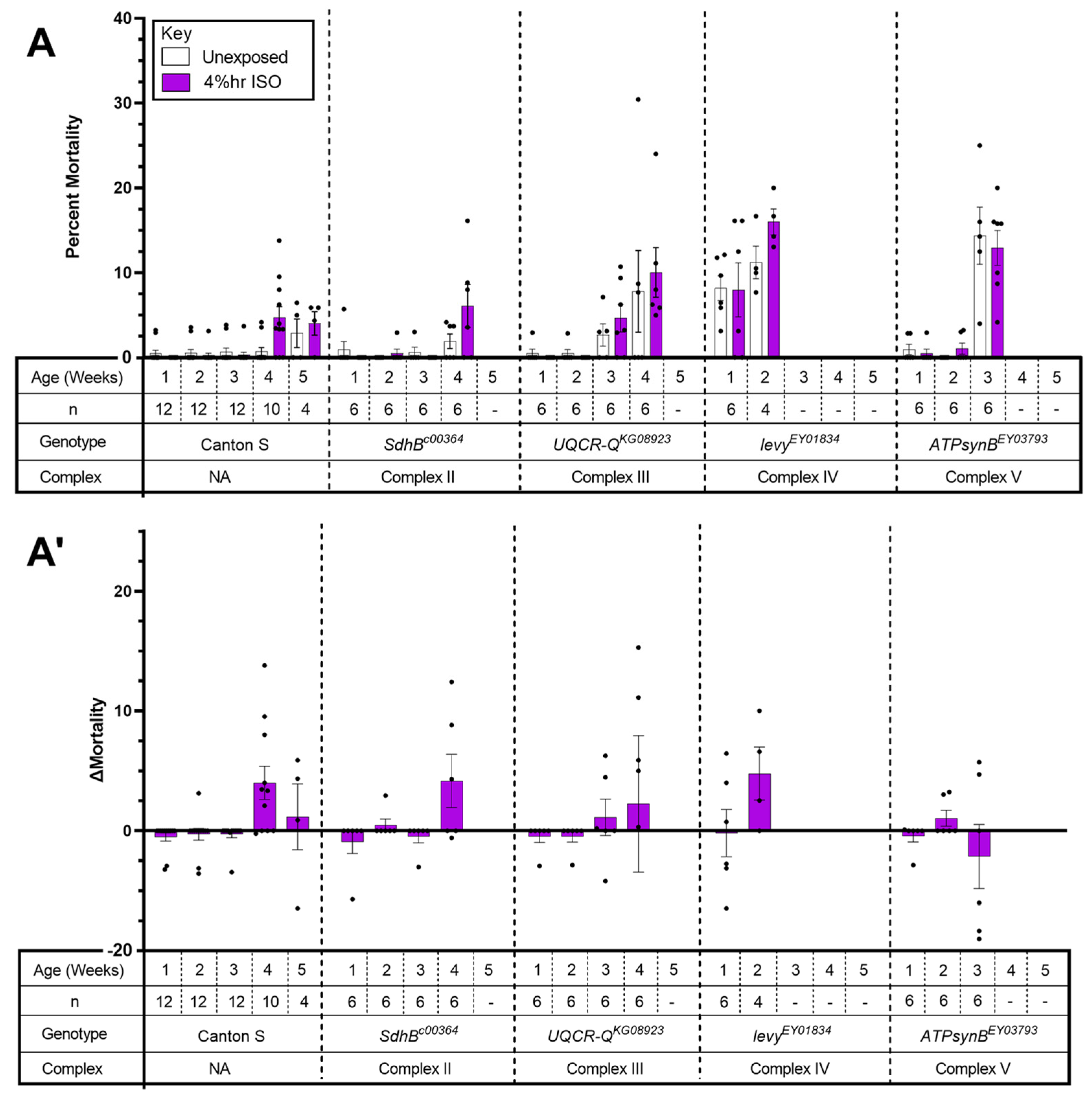
2.3. The Tested Complex II-V Mutants Did Not Show AiN
2.4. AiN Is weakly Associated with Increased Intestinal Permeability
3. Discussion
3.1. HAL Causes AiN in Mitochondrial Mutants
3.2. Mutations in Accessory Subunits Sensitize to AiN
3.3. Mutations in Complexes II–V Do Not Result in AiN
3.4. AiN Differs from TBI in Its Systemic Impact
3.5. Limitations
4. Materials and Methods
4.1. Fly Lines and Culturing
4.2. Anesthesia and O2 Exposure
4.3. Smurfing
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Perouansky, M. General anesthetics and long-term neurotoxicity. Handb. Exp. Pharmacol. 2008, 143–157. [Google Scholar] [CrossRef]
- Kayser, E.B.; Morgan, P.G.; Sedensky, M.M. GAS-1: A mitochondrial protein controls sensitivity to volatile anesthetics in the nematode Caenorhabditis elegans. Anesthesiology 1999, 90, 545–554. [Google Scholar] [CrossRef] [PubMed]
- Kayser, E.B.; Morgan, P.G.; Sedensky, M.M. Mitochondrial complex I function affects halothane sensitivity in Caenorhabditis elegans. Anesthesiology 2004, 101, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Morgan, P.G.; Hoppel, C.L.; Sedensky, M.M. Mitochondrial defects and anesthetic sensitivity. Anesthesiology 2002, 96, 1268–1270. [Google Scholar] [CrossRef]
- Niezgoda, J.; Morgan, P.G. Anesthetic considerations in patients with mitochondrial defects. Paediatr. Anaesth. 2013, 23, 785–793. [Google Scholar] [CrossRef] [Green Version]
- Finsterer, J. Leigh and Leigh-like syndrome in children and adults. Pediatr. Neurol. 2008, 39, 223–235. [Google Scholar] [CrossRef]
- Quintana, A.; Kruse, S.E.; Kapur, R.P.; Sanz, E.; Palmiter, R.D. Complex I deficiency due to loss of Ndufs4 in the brain results in progressive encephalopathy resembling Leigh syndrome. Proc. Natl. Acad. Sci. USA 2010, 107, 10996–11001. [Google Scholar] [CrossRef] [Green Version]
- Loewen, C.A.; Ganetzky, B. Mito-Nuclear Interactions Affecting Lifespan and Neurodegeneration in a Drosophila Model of Leigh Syndrome. Genetics 2018, 208, 1535–1552. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; DeLuca, S.Z.; O’Farrell, P.H. Manipulating the metazoan mitochondrial genome with targeted restriction enzymes. Science 2008, 321, 575–577. [Google Scholar] [CrossRef] [Green Version]
- Burman, J.L.; Itsara, L.S.; Kayser, E.B.; Suthammarak, W.; Wang, A.M.; Kaeberlein, M.; Sedensky, M.M.; Morgan, P.G.; Pallanck, L.J. A Drosophila model of mitochondrial disease caused by a complex I mutation that uncouples proton pumping from electron transfer. Dis. Model. Mech. 2014, 7, 1165–1174. [Google Scholar] [CrossRef]
- Olufs, Z.P.G.; Loewen, C.A.; Ganetzky, B.; Wassarman, D.A.; Perouansky, M. Genetic variability affects absolute and relative potencies and kinetics of the anesthetics isoflurane and sevoflurane in Drosophila melanogaster. Sci. Rep. 2018, 8, 2348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olufs, Z.P.G.; Ganetzky, B.; Wassarman, D.A.; Perouansky, M. Mitochondrial Complex I Mutations Predispose Drosophila to Isoflurane Neurotoxicity. Anesthesiology 2020, 133, 839–851. [Google Scholar] [CrossRef] [PubMed]
- Rhooms, S.K.; Murari, A.; Goparaju, N.S.V.; Vilanueva, M.; Owusu-Ansah, E. Insights from Drosophila on mitochondrial complex I. Cell. Mol. Life Sci. 2020, 77, 607–618. [Google Scholar] [CrossRef] [PubMed]
- Sazanov, L.A. A giant molecular proton pump: Structure and mechanism of respiratory complex I. Nat. Rev. Mol. Cell Biol. 2015, 16, 375–388. [Google Scholar] [CrossRef]
- Kirby, D.M.; Crawford, M.; Cleary, M.A.; Dahl, H.H.; Dennett, X.; Thorburn, D.R. Respiratory chain complex I deficiency: An underdiagnosed energy generation disorder. Neurology 1999, 52, 1255–1264. [Google Scholar] [CrossRef]
- Weaver, J.L. The brain-gut axis: A prime therapeutic target in traumatic brain injury. Brain Res. 2021, 1753, 147225. [Google Scholar] [CrossRef]
- Katzenberger, R.J.; Chtarbanova, S.; Rimkus, S.A.; Fischer, J.A.; Kaur, G.; Seppala, J.M.; Swanson, L.C.; Zajac, J.E.; Ganetzky, B.; Wassarman, D.A. Death following traumatic brain injury in Drosophila is associated with intestinal barrier dysfunction. eLife 2015, 4, e04790. [Google Scholar] [CrossRef]
- Rera, M.; Bahadorani, S.; Cho, J.; Koehler, C.L.; Ulgherait, M.; Hur, J.H.; Ansari, W.S.; Lo, T., Jr.; Jones, D.L.; Walker, D.W. Modulation of longevity and tissue homeostasis by the Drosophila PGC-1 homolog. Cell Metab. 2011, 14, 623–634. [Google Scholar] [CrossRef] [Green Version]
- Rera, M.; Clark, R.I.; Walker, D.W. Intestinal barrier dysfunction links metabolic and inflammatory markers of aging to death in Drosophila. Proc. Natl. Acad. Sci. USA 2012, 109, 21528–21533. [Google Scholar] [CrossRef] [Green Version]
- Njoku, D.; Laster, M.J.; Gong, D.H.; Eger, E.I., 2nd; Reed, G.F.; Martin, J.L. Biotransformation of halothane, enflurane, isoflurane, and desflurane to trifluoroacetylated liver proteins: Association between protein acylation and hepatic injury. Anesth. Analg. 1997, 84, 173–178. [Google Scholar] [CrossRef]
- Sawyer, D.C.; Eger, E.I., 2nd; Bahlman, S.H.; Cullen, B.F.; Impelman, D. Concentration dependence of hepatic halothane metabolism. Anesthesiology 1971, 34, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.N.; Hunter, F.E., Jr. The effect of halothane on electron transport, oxidative phosphorylation, and swelling in rat liver mitochondria. Mol. Pharmacol. 1970, 6, 67–77. [Google Scholar] [PubMed]
- Ross, W.T., Jr.; Daggy, B.P.; Cardell, R.R., Jr. Hepatic necrosis caused by halothane and hypoxia in phenobarbital-treated rats. Anesthesiology 1979, 51, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Quintana, A.; Morgan, P.G.; Kruse, S.E.; Palmiter, R.D.; Sedensky, M.M. Altered anesthetic sensitivity of mice lacking Ndufs4, a subunit of mitochondrial complex I. PLoS ONE 2012, 7, e42904. [Google Scholar] [CrossRef] [Green Version]
- Falk, M.J.; Kayser, E.B.; Morgan, P.G.; Sedensky, M.M. Mitochondrial complex I function modulates volatile anesthetic sensitivity in C. elegans. Curr. Biol. CB 2006, 16, 1641–1645. [Google Scholar] [CrossRef] [Green Version]
- Kirkman, M.A.; Yu-Wai-Man, P.; Korsten, A.; Leonhardt, M.; Dimitriadis, K.; De Coo, I.F.; Klopstock, T.; Chinnery, P.F. Gene-environment interactions in Leber hereditary optic neuropathy. Brain A J. Neurol. 2009, 132, 2317–2326. [Google Scholar] [CrossRef]
- Acharya, N.K.; Goldwaser, E.L.; Forsberg, M.M.; Godsey, G.A.; Johnson, C.A.; Sarkar, A.; DeMarshall, C.; Kosciuk, M.C.; Dash, J.M.; Hale, C.P.; et al. Sevoflurane and Isoflurane induce structural changes in brain vascular endothelial cells and increase blood-brain barrier permeability: Possible link to postoperative delirium and cognitive decline. Brain Res. 2015, 1620, 29–41. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drosophila Gene and Allele | Mammalian Name | BDSC Stock Number | Insertion Location/Description of Mutation | Type/Location of Subunit |
|---|---|---|---|---|
| mt:ND2del1 | ND2 | NA | 9 bp deletion destroys the BglII restriction enzyme site | Core/Membrane Arm |
| ND-2360114 | NDUFS8 | NA | G199A Amino Acid Substitution | Core/Matrix Arm |
| ND-23B117 | NDUFS8 | 16143 | ~50 bp upstream of 5′ UTR | Core/Matrix Arm |
| ND-13BEY07792 | NDUFA5 | 16860 | 5′ UTR | Accessory/Matrix Arm |
| ND-39EY06323 | NDUFA9 | 15821 | Intron/~50 bp upstream of 5′ UTR | Accessory/Matrix Arm |
| ND-15MI10315 | NDUFS5 | 53837 | 5′ UTR/Intron | Accessory/Membrane Arm |
| ND-B22KG08637 | NDUFB9 | 15134 | 5′ UTR | Accessory/Membrane Arm |
| ND-SGDHG6447 | NDUFB5 | 27208 | 5′ UTR | Accessory/Membrane Arm |
| Drosophila Gene | Mammalian Name | BDSC Stock Number | Insertion Location/Description of Mutation | Complex |
|---|---|---|---|---|
| SdhBc00364 | SdhB | 10039 | 5′ UTR | Complex II |
| UQCR-QKG08923 | UQCRQ | 16470 | Intron | Complex III |
| LevyEY01834 | COX6A | 15080 | 5′ UTR | Complex IV |
| ATPsynBEY03793 | ATP5PB | 16575 | 5′ UTR | Complex V |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borchardt, L.A.; Scharenbrock, A.R.; Olufs, Z.P.G.; Wassarman, D.A.; Perouansky, M. Mutations in Complex I of the Mitochondrial Electron-Transport Chain Sensitize the Fruit Fly (Drosophila melanogaster) to Ether and Non-Ether Volatile Anesthetics. Int. J. Mol. Sci. 2023, 24, 1843. https://doi.org/10.3390/ijms24031843
Borchardt LA, Scharenbrock AR, Olufs ZPG, Wassarman DA, Perouansky M. Mutations in Complex I of the Mitochondrial Electron-Transport Chain Sensitize the Fruit Fly (Drosophila melanogaster) to Ether and Non-Ether Volatile Anesthetics. International Journal of Molecular Sciences. 2023; 24(3):1843. https://doi.org/10.3390/ijms24031843
Chicago/Turabian StyleBorchardt, Luke A., Amanda R. Scharenbrock, Zachariah P. G. Olufs, David A. Wassarman, and Misha Perouansky. 2023. "Mutations in Complex I of the Mitochondrial Electron-Transport Chain Sensitize the Fruit Fly (Drosophila melanogaster) to Ether and Non-Ether Volatile Anesthetics" International Journal of Molecular Sciences 24, no. 3: 1843. https://doi.org/10.3390/ijms24031843