
Pomegranate Seed Oil as a Source of Conjugated Linolenic Acid (CLnA) Has No Effect on Atherosclerosis Development but Improves Lipid Profile and Affects the Expression of Lipid Metabolism Genes in apoE/LDLR−/− Mice
 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Effect of CLnA on Body Weight
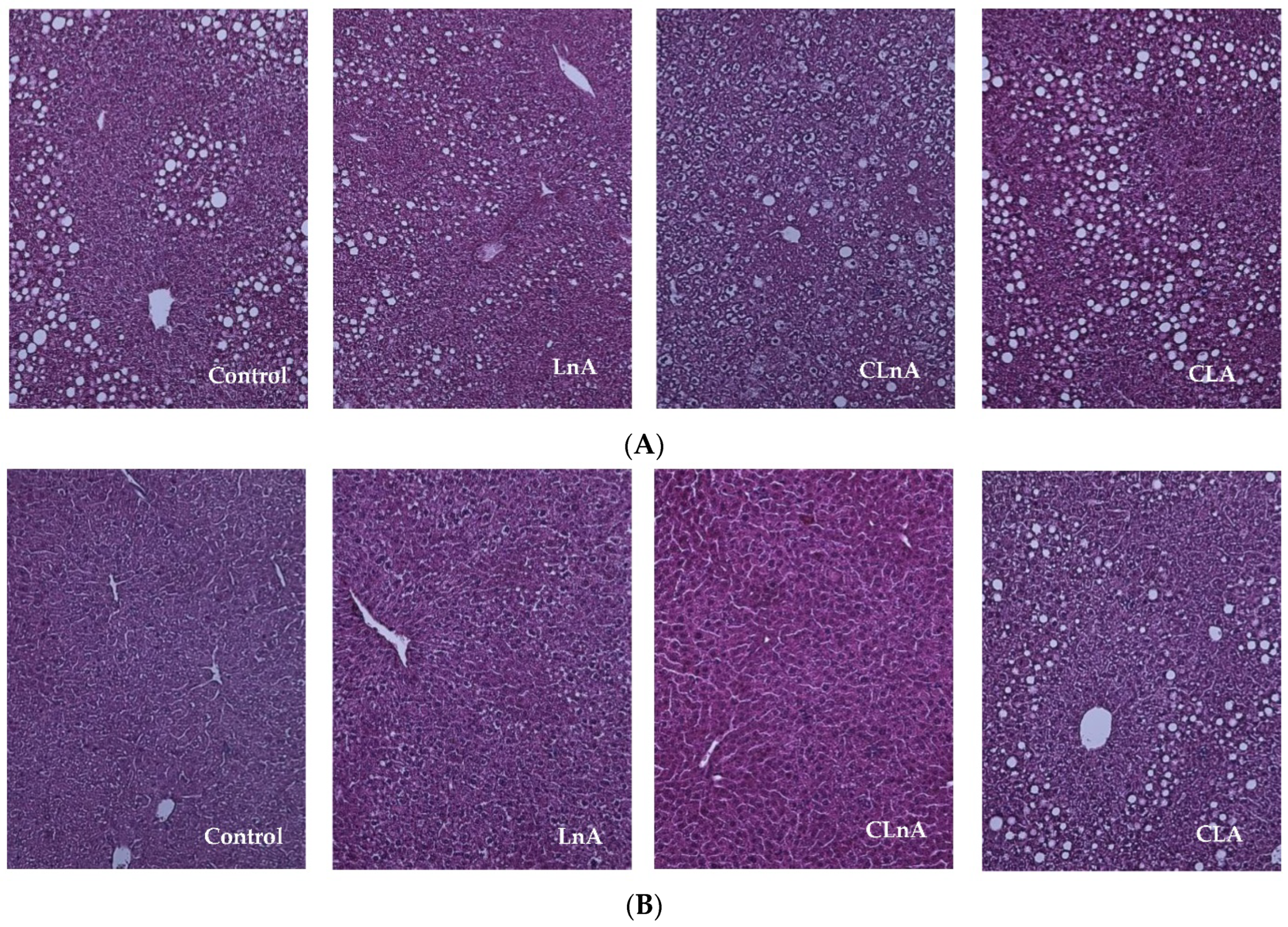
2.2. Effect of CLnA on Liver Weight and Histology
2.3. Effect of CLnA on Glucose Concentration
2.4. Effect of CLnA on Plasma Lipid Profile and Expression of PPARα and SREBP-1 and Their Target Genes (ACO and FAS )
2.5. Effect of CLnA on Fatty Acid Profile in Tissues
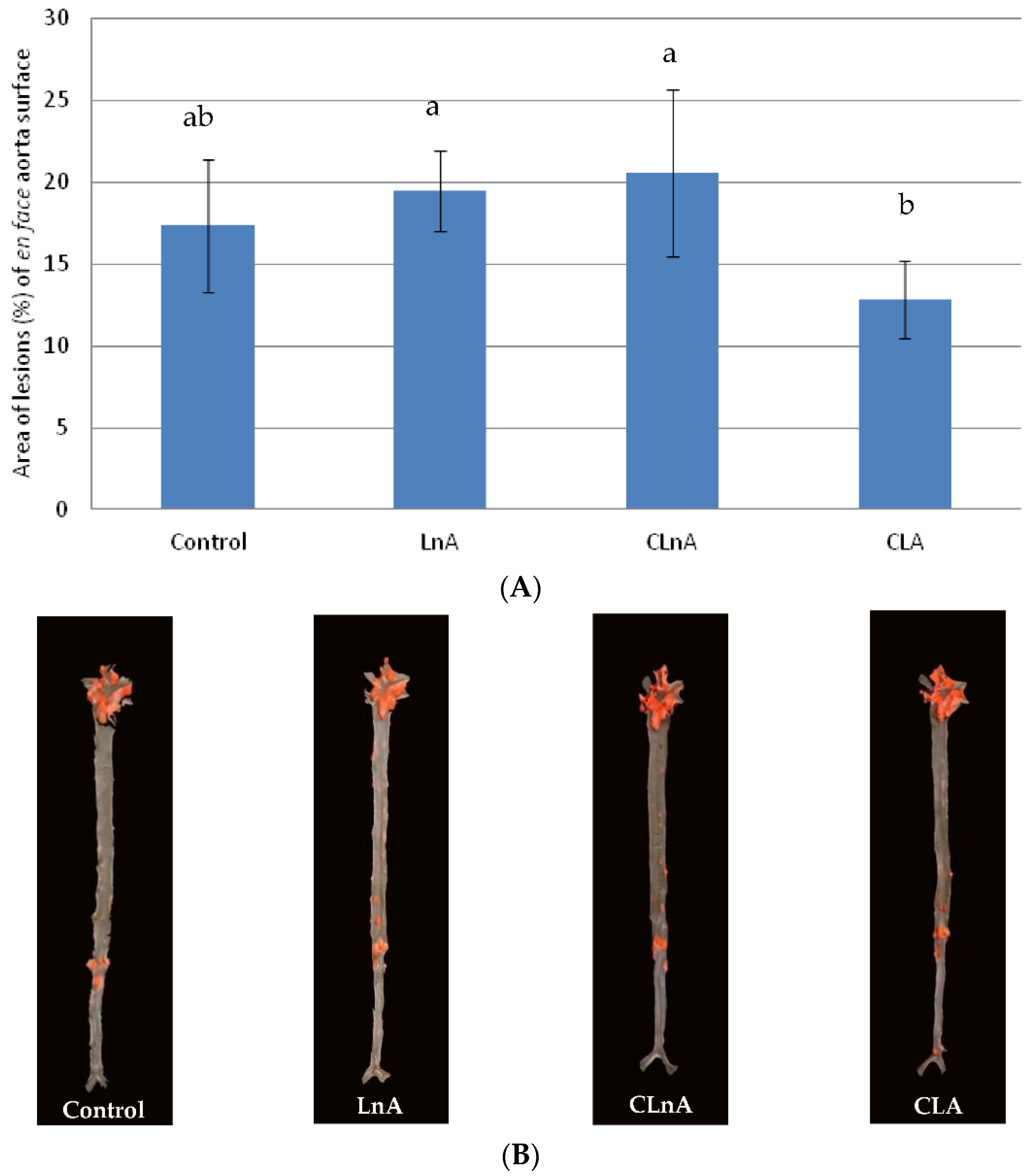
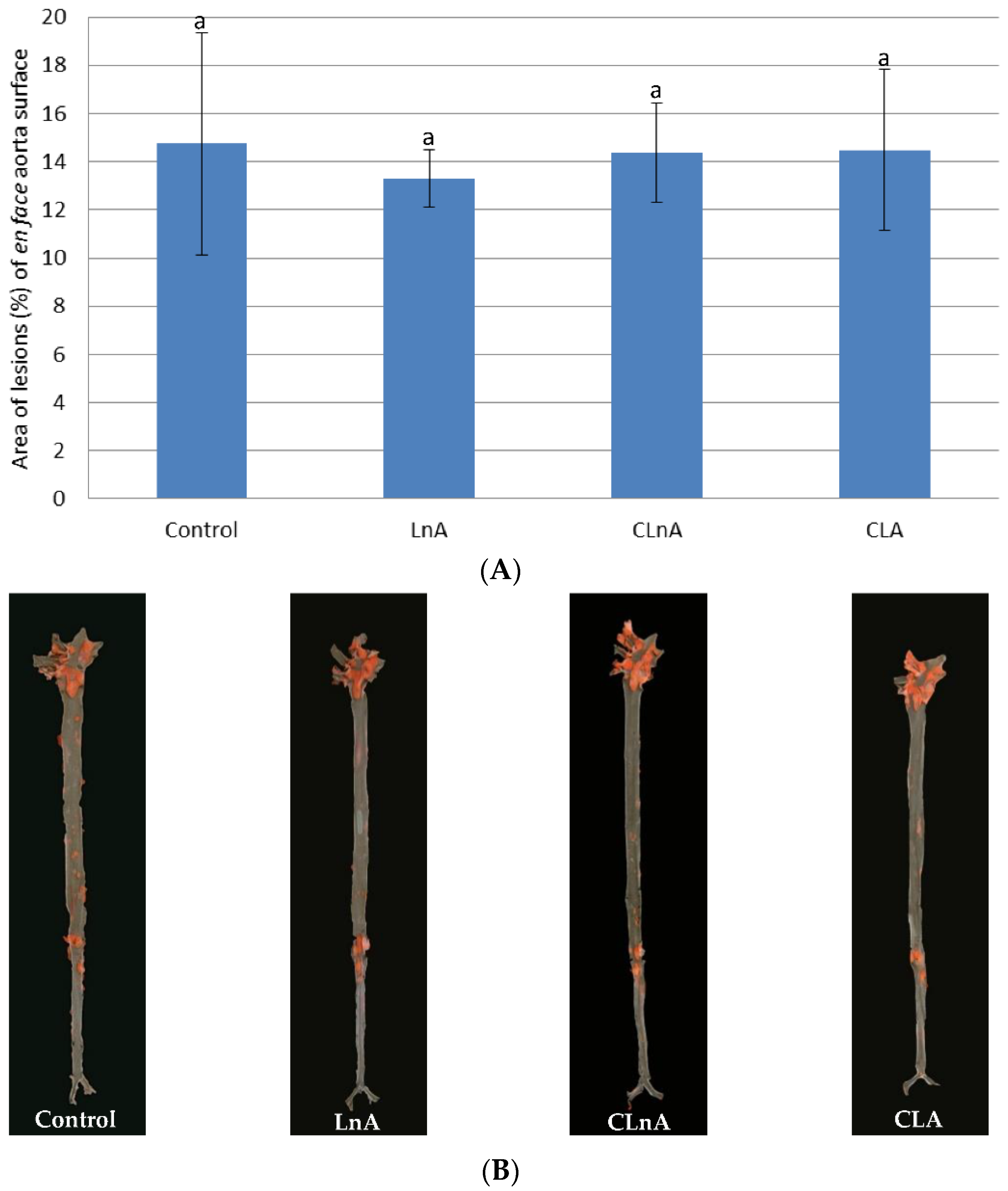
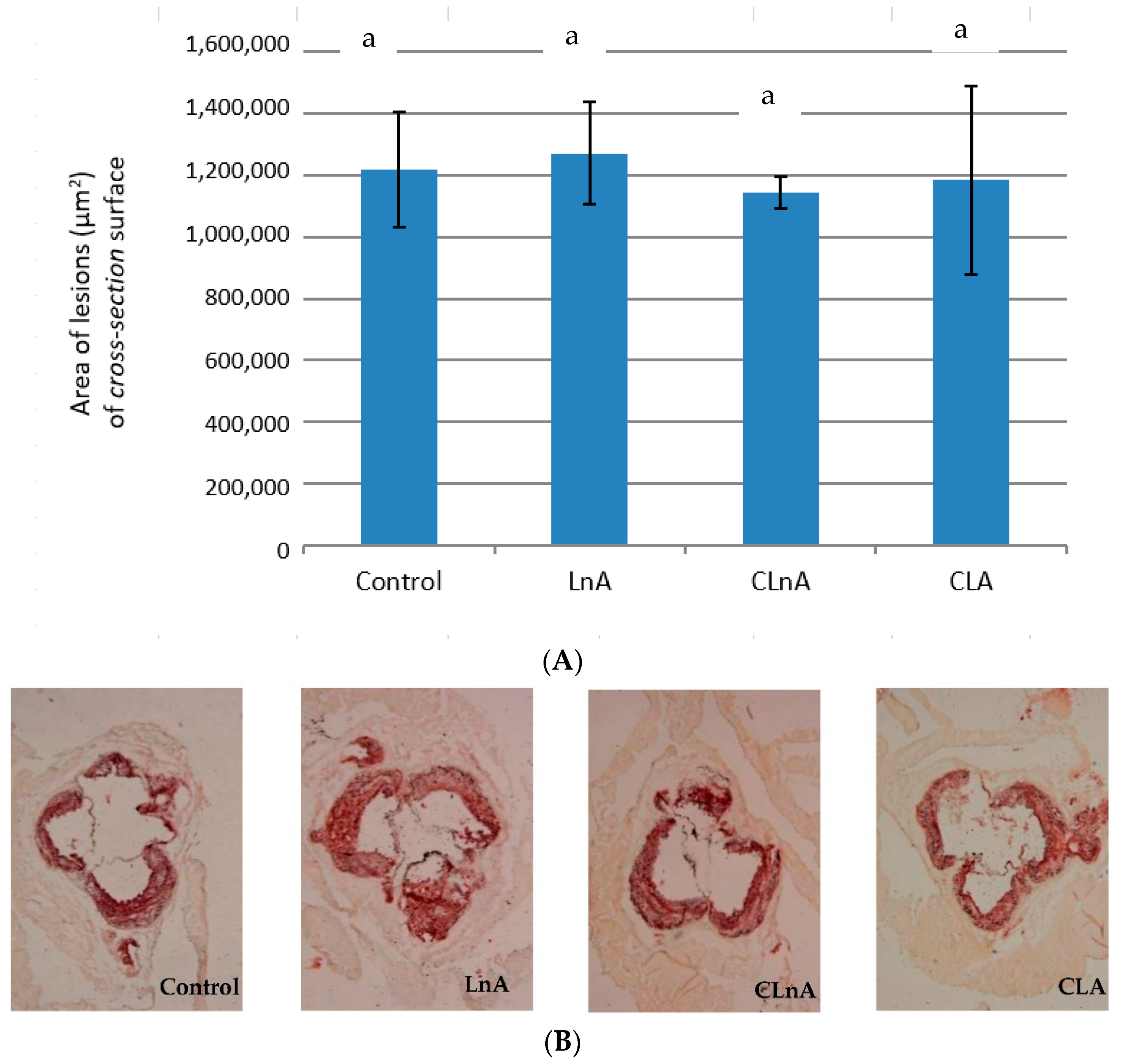
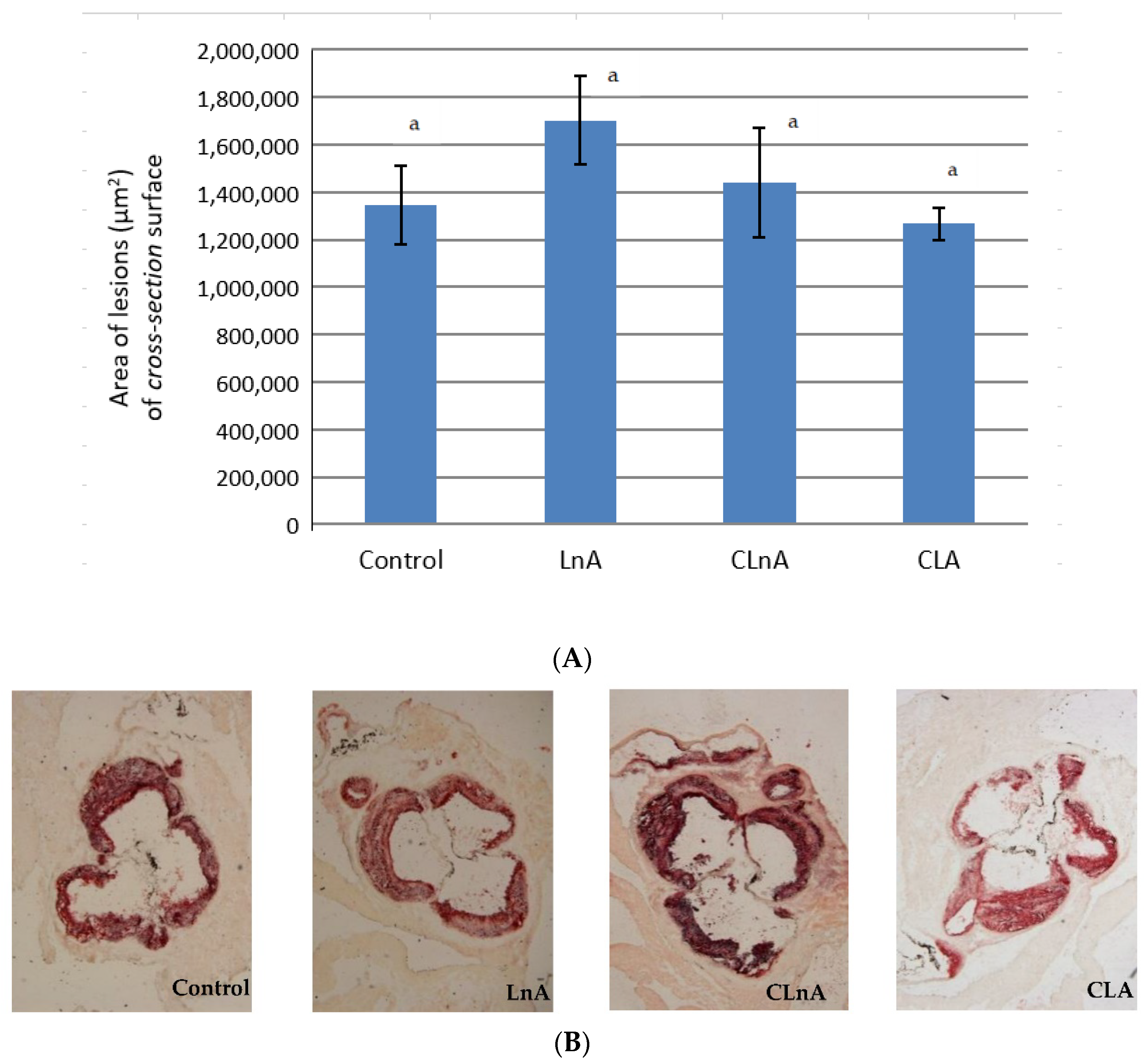
2.6. Effect of CLnA on the Development of Atherosclerosis
3. Discussion
3.1. Effect of CLnA on Body Weight
3.2. Effect of CLA Isomers on Liver Weight and Histology
3.3. Effect of CLA Isomers on Glucose Concentration
3.4. Effect of CLA Isomers on Plasma Lipid Profile and Expression of PPARα and SREBP-1 and Their Target Genes (ACO and FAS )
3.5. Effect of CLA on Fatty Acid Profile in Tissues
3.6. Effect of CLnA on the Development of Atherosclerosis
4. Materials and Methods
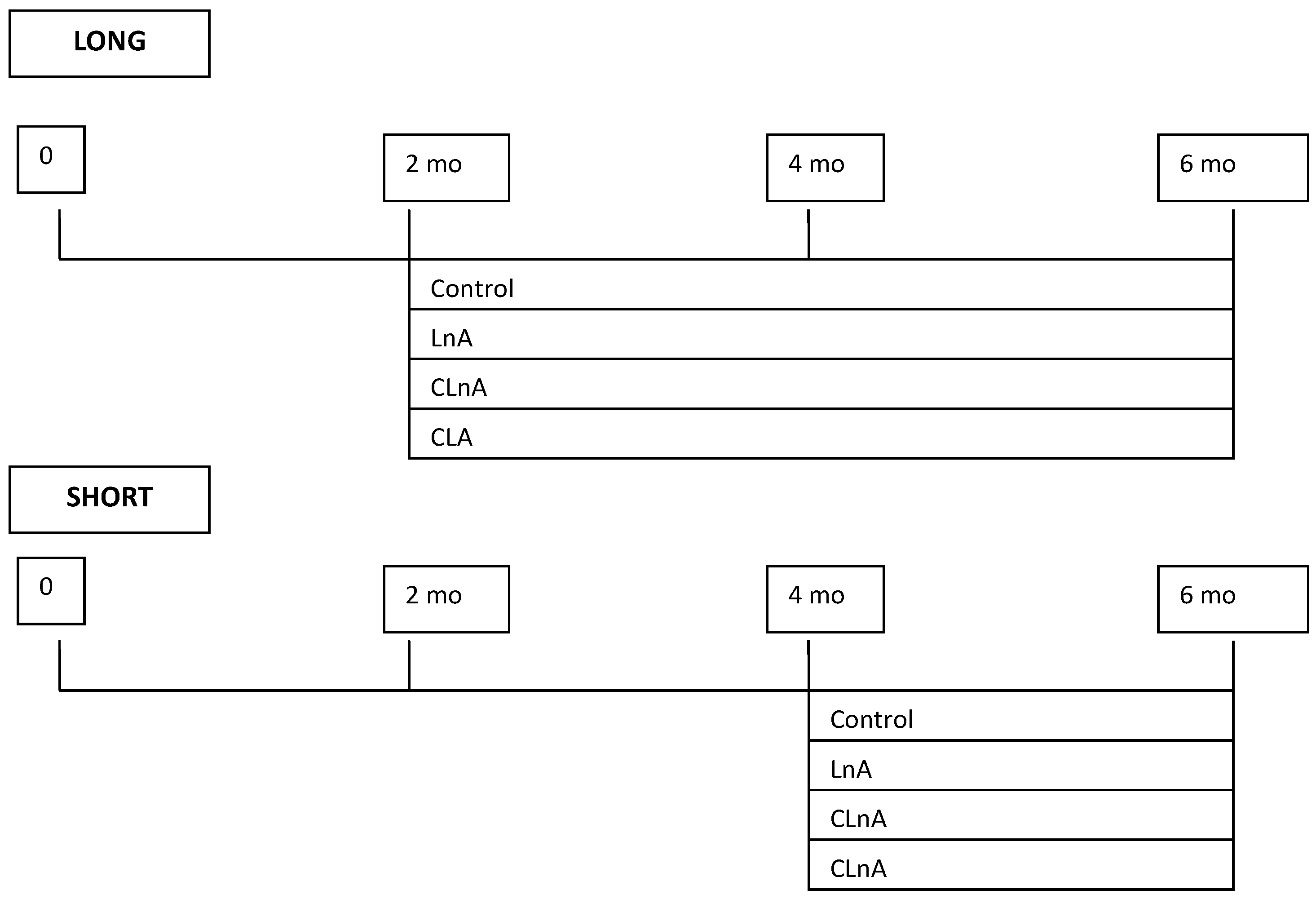
4.1. Animals and Housing
4.2. Diets and Feeding
4.3. Blood Sampling, Plasma Lipid Profile and Glucose Analyses
4.4. Fatty Acid Profile of Tissues
4.5. Quantification of Atherosclerosis in Descending Aortas (en Face Analysis)
4.6. Quantitation of Atherosclerosis in Aortic Roots (Cross-Section Analysis)
4.7. Liver Histological Examination
4.8. Real-Time qRT-PCR
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yamasaki, M.; Kitagawa, T.; Koyanagi; Chujo, N.H.; Maeda, H.; Kohno-Murase, J.; Imamura, J.; Tachibana, H.; Yamada, K. Dietary effect of pomegranate seed oil on immune function and lipid metabolism in mice. Nutrition 2006, 22, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Koba, K.; Belury, M.A.; Sugano, M. Potential health benefits of conjugated trienoic acids. Lipid Technol. 2007, 19, 200–220. [Google Scholar] [CrossRef]
- Bassaganya-Riera, J.; DiGuardo, M.; Climent, M.; Vives, C.; Carbo, A.; Jouni, Z.E.; Einerhand, A.W.C.; O’Shea, M.; Hontecillas, R. Activation of PPARγ and δ by dietary punicic acid ameliorates intestinal inflammation in mice. Br. J. Nutr. 2011, 106, 878–886. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, R.; Noguchi, R.; Ota, T.; Abe, M.; Miyashita, K.; Kawada, T. Cytotoxic Effect of Conjugated Trienoic Fatty Acids on Mouse Tumor and Human Monocytic Leukemia Cells. Lipids 2001, 36, 477–482. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.D.; Mehta, R.; Yu, W.; Neeman, I.; Livney, T.; Amichay, A.; Poirier, D.; Nicholls, P.; Kirby, A.; Jiang, W.; et al. Chemopreventive and adjuvant therapeutic potential of pomegranate (Punica granatum) for human breast cancer. Breast Can. Res. Treat. 2002, 71, 203–217. [Google Scholar] [CrossRef]
- Kohno, H.; Suzuki, R.; Yasui, Y.; Hosokawa, M.; Miyashita, K.; Tanakaet, T. Pomegranate seed oil rich in conjugated linolenic acid suppresses chemically induced colon carcinogenesis in rats. Cancer Sci. 2004, 95, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Arao, K.; Wang, Y.M.; Inoue, N.; Hirata, J.; Cha, J.-Y.; Nagao, K.; Yanagitaet, T. Dietary effect of pomegranate seed oil rich in 9cis, 11trans, 13cis conjugated linolenic acid on lipid metabolism in obese, hyperlipidemic OLETF rats. Lipids Health Dis. 2004, 9, 24–30. [Google Scholar] [CrossRef] [Green Version]
- Grossmann, M.E.; Mizuno, N.K.; Schuster, T.; Cleary, M.P. Punicic acid is an ω-5 fatty acid capable of inhibiting breast cancer proliferation. Int. J. Oncol. 2010, 36, 421–426. [Google Scholar]
- Tsuzuki, T.; Tokuyama, Y.; Igarashi, M.; Nakagawa, K.; Ohsaki, Y.; Komai, M.; Miyazawat, T. α-Eleostearic Acid (9Z11E13E-18:3) Is Quickly Converted to Conjugated Linoleic Acid (9Z11E-18:2) in Rats. J. Nutr. 2004, 134, 2634–2639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuzuki, T.; Kawakami, Y.; Abe, R.; Nakagawa, K.; Koba, K.; Imamura, J.; Iwata, T.; Ikeda, I.; Miyazawa, T. Conjugated Linolenic Acid Is Slowly Absorbed in Rat Intestine, but Quickly Converted to Conjugated Linoleic Acid. J. Nutr. 2006, 136, 2153–2159. [Google Scholar] [CrossRef] [Green Version]
- Yuan, G.; Sun, H.; Sinclair, A.J.; Li, D. Effects of conjugated linolenic acid and conjugated linoleic acid on lipid metabolism in mice. Eur. J. Lipid Sci. Technol. 2009, 111, 537–545. [Google Scholar] [CrossRef]
- Schneider, A.C.; Mignolet, E.; Schneider, Y.J.; Larondelle, Y. Uptake of conjugated linolenic acids and conversion to cis-9, trans-11-or trans-9, trans-11-conjugated linoleic acids in Caco-2 cells. Br. J. Nutr. 2013, 109, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Boussetta, T.; Raad, H.; Lettéron, P.; Gougerot-Pocidalo, M.-A.; Marie, J.C.; Driss, F.; El-Benna, J. Punicic Acid a Conjugated Linolenic Acid Inhibits TNFα-Induced Neutrophil Hyperactivation and Protectsfrom Experimental Colon Inflammation in Rats. PLoS ONE 2009, 4, e6458–e6469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhar, P.; Ghosh, S.; Bhattacharyya, D.K. Dietary effects of conjugated octadecatrienoic fatty acid (9 cis, 11 trans, 13 trans) levels on blood lipids and nonenzymatic in vitro lipid peroxidation in rats. Lipids 1999, 34, 109–114. [Google Scholar] [CrossRef]
- Mukherjee, C.; Bhattacharyya, S.; Ghosh, S.; Bhattacharyya, D.K. Dietary effects of punicic acid on the composition and peroxidation of rat plasma lipid. J. Oleo Sci. 2002, 51, 513–522. [Google Scholar] [CrossRef] [Green Version]
- Saha, S.S.; Ghosh, M. Antioxidant effect of vegetable oils containing conjugated linolenic acid isomers against induced tissue lipid peroxidation and inflammation in rat model. Chem. Biol. Interact. 2011, 190, 109–120. [Google Scholar] [CrossRef]
- Changhua, L.; Jindong, Y.; Defa, L.; Lidan, Z.; Shiyan, Q.; Jianjun, X. Conjugated linoleic acid attenuates the production and gene expression of proinflammatory cytokines in weaned pigs challenged with lipopolysaccharide. J. Nutr. 2005, 135, 239–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hontecillas, R.; Diguardo, M.; Duran, E.; Orpi, M.; Bassaganya-Riera, J. Catalpic acid decreases abdominal fat deposition, improves glucose homeostasis and upregulates PPAR alpha expression in adipose tissue. Clin. Nutr. 2008, 27, 764–772. [Google Scholar] [CrossRef]
- Hontecillas, R.; O’Shea, M.; Einerhand, A.; Diguardo, M.; Bassaganya-Riera, J. Activation of PPAR gamma and alpha by punicic acid ameliorates glucose tolerance and suppresses obesity-related inflammation. J. Am. Coll. Nutr. 2009, 28, 184–195. [Google Scholar] [CrossRef]
- McFarlin, B.K.; Strohacker, K.A.; Kueht, M.L. Pomegranate seed oil consumption during a period of high-fat feeding reduces weight gain and reduces type 2 diabetes risk in CD-1 mice. Br. J. Nutr. 2009, 102, 54–59. [Google Scholar] [CrossRef] [Green Version]
- Meerts, I.A.; Verspeek-Rip, C.M.; Buskens, C.A.; Keizer, H.G.; Bassaganya-Riera, J.; Jouni, Z.E.; van Huygevoort, A.H.B.M.; van Otterdijk, F.M.; van de Waartet, E.J. Toxicological evaluation of pomegranate seed oil. Food Chem. Toxicol. 2009, 47, 1085–1092. [Google Scholar] [CrossRef]
- Mirmiran, P.; Fazeli, M.R.; Asghari, G.; Shafiee, A.; Azizi, F. Effect of pomegranate seed oil on hyperlipidaemic subjects: A doubleblind placebo-controlled clinical trial. Br. J. Nutr. 2010, 104, 402–406. [Google Scholar] [CrossRef] [Green Version]
- Saha, S.S.; Chakraborty, A.; Ghosh, S.; Ghosh, M. Comparative study of hypocholesterolemic and hypolipidemic effects of conjugated linolenic acid isomers against induced biochemical perturbations and aberration in erythrocyte membrane fluidity. Eur. J. Nutr. 2012, 51, 483–495. [Google Scholar] [CrossRef]
- Miranda, J.; Aguirre, L.; Fernández-Quintela, A.; Macarulla, M.T.; Guadalupe Martínez-Castaño, M.; Ayo, J.; Bilbao, E.; Portillo, M.P. Effects of pomegranate seed oil on glucose and lipid metabolism-related organs in rats fed an obesogenic diet. J. Agric. Food Chem. 2013, 61, 5089–5096. [Google Scholar] [CrossRef] [PubMed]
- Miranda, J.; Fernández-Quintela, A.; Macarulla, M.T.; Churruca, I.; García, C.; Rodríguez, V.M.; Simón, E.; Portillo, M.P. A comparison between CLNA and CLA effects on body fat, serum parameters and liver composition. J. Physiol. Biochem. 2009, 65, 25–32. [Google Scholar] [CrossRef]
- Shinohara, N.; Tsuduki, T.; Ito, J.; Honma, T.; Kijima, R.; Sugawara, S.; Arai, T.; Yamasaki, M.; Ikezaki, A.; Yokoyama, M.; et al. Jacaric acid, a linolenic acid isomer with a conjugated triene system, reduces stearoyl-CoA desaturase expression in liver of mice. J. Oleo Sci. 2012, 61, 433–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vroegrijk, I.O.; van Diepen, J.A.; van den Berg, S.; Westbroek, I.; Keizer, H.; Gambelli, L.; Hontecillas, R.; Bassaganya-Riera, J.; Zondag, G.C.M.; Romijn, J.A.; et al. Pomegranate seed oil, a rich source of punicic acid, prevents diet-induced obesity and insulin resistance in mice. Food Chem. Toxicol. 2011, 49, 1426–1430. [Google Scholar] [CrossRef]
- Koba, K.; Akahoshi, A.; Yamasaki, M.; Tanaka, K.; Yamada, K.; Iwata, T.; Kamegai, T.; Tsutsumi, K.; Sugano, M. Dietary conjugated linolenic acid in relation to CLA differently modifies body fat mass and serum and liver lipid levels in rats. Lipids 2002, 37, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, R.; Yasui, Y.; Suzuki, R.; Hosokawa, M.; Fukunaga, K.; Miyashita, K. Dietary Effects of Bitter Gourd Oil on Blood and Liver Lipids of Rats. Arch. Biochem. Biophys. 2001, 396, 207–212. [Google Scholar] [CrossRef]
- Yang, L.; Leung, K.A.; Cao, Y.; Huang, Y.; Ratnayake, W.M.N.; Chen, Z.-Y. α-Linolenic acid but not conjugated linolenic acid is hypocholesterolaemic in hamsters. Br. J. Nutr. 2005, 93, 433–438. [Google Scholar] [CrossRef]
- Ashwell, M.S.; Ceddia, R.P.; House, R.L.; Cassady, J.P.; Eisen, E.J.; Eling, T.E.; Collins, J.B.; Grissom, S.F.; Odle, J. Trans-10, cis-12-conjugated linoleic acid alters hepatic gene expression in a polygenic obese line of mice displaying hepatic lipidosis. J. Nutr. Biochem. 2010, 21, 848–855. [Google Scholar] [CrossRef] [PubMed]
- Raffaele, M.; Licari, M.; Amin, S.; Alex, R.; Shen, H.; Singh, S.P.; Vanella, L.; Rezzani, R.; Bonomini, F.; Peterson, S.J.; et al. Cold Press Pomegranate Seed Oil Attenuates Dietary-Obesity Induced Hepatic Steatosis and Fibrosis through Antioxidant and Mitochondrial Pathways in Obese Mice. Int. J. Mol. Sci. 2020, 21, 5469. [Google Scholar] [CrossRef]
- Nekooeian, A.A.; Eftekhari, M.H.; Adibi, S.; Rajaeifard, A. Effects of pomegranate seed oil on insulin release in rats with type 2 diabetes. Iran J. Med. Sci. 2014, 39, 130–135. [Google Scholar] [PubMed]
- Saha, S.S.; Ghosh, M. Antioxidant and anti-inflammatory effect of conjugated linolenic acid isomers against streptozotocin-induced diabetes. Br. J. Nutr. 2012, 108, 974–983. [Google Scholar] [CrossRef] [Green Version]
- Dhar, P.; Bhattacharyya, D.; Bhattacharyya, D.K.; Ghosh, S. Dietary comparison of conjugated linolenic acid (9cis, 11trans, 13trans) and α-tocopherol effects on blood lipids and lipid peroxidation in alloxan-induced diabetes mellitus rats. Lipids 2006, 41, 49–54. [Google Scholar] [CrossRef]
- Lam, C.K.; Chen, J.; Cao, Y.; Yang, L.; Wong, Y.M.; Yeung, S.Y.V.; Yao, X.; Huang, Y.; Chen, Z.Y. Conjugated and non-conjugated octadecaenoic acids affect differently intestinal acyl coenzyme A: Cholesterol acyltransferase activity. Atherosclerosis 2008, 198, 85–93. [Google Scholar] [CrossRef]
- Koba, K.; Imamura, J.; Akashoshi, A.; Kohno-Murase, J.; Nishizono, S.; Iwabuchi, M.; Tanaka, K.; Sugano, M. Genetically Modified Rapeseed Oil Containing cis-9, trans-11, cis-13-Octadecatrienoic Acid Affects Body Fat Mass and Lipid Metabolism in Mice. J. Agric. Food Chem. 2007, 55, 3741–3748. [Google Scholar] [CrossRef]
- Chuang, C.Y.; Hsu, C.; Chao, C.Y.; Wein, Y.-S.; Kuo, Y.-H.; Huanget, C. Fractionation and identification of 9c, 11t, 13t-conjugated linolenic acid as an activator of PPARalpha in bitter gourd (Momordica charantia L.). J. Biomed. Sci. 2006, 13, 763–772. [Google Scholar] [CrossRef] [Green Version]
- Lewis, S.N.; Brannan, L.; Guri, A.J.; Lu, P.; Hontecillas, R.; Bassaganya-Riera, J.; Bevanet, D.R. Dietary α-eleostearic acid ameliorates experimental inflammatory bowel disease in mice by activating peroxisome proliferator-activated receptor-γ. PLoS ONE 2011, 6, e24031. [Google Scholar] [CrossRef] [PubMed]
- Lepionka, T.; Białek, M.; Czauderna, M.; Białek, A. Pomegranate seed oil and bitter melon extract supplemented in diet influence the lipid profile and intensity of peroxidation in livers of SPRD rats exposed to a chemical carcinogen. Prostaglandins Other Lipid Mediat. 2021, 152, 106495. [Google Scholar] [CrossRef]
- Yuan, G.; Sinclair, A.J.; Xu, C.; Li, D. Incorporation and metabolism of punicic acid in healthy young humans. Mol. Nutr. Food Res. 2009, 53, 1336–1342. [Google Scholar] [CrossRef]
- Saha, S.S.; Ghosh, M. Protective effect of conjugated linolenic acid isomers present in vegetable oils against arsenite-induced renal toxicity in rat model. Nutrition 2013, 29, 903–910. [Google Scholar] [CrossRef]
- Saha, S.S.; Dasgupta, P.; Sengupta, B.S.; Ghosh, M. Synergistic effect of conjugated linolenic acid isomers against induced oxidative stress, inflammation and erythrocyte membrane disintegrity in rat model. Biochim. Biophys. Acta 2012, 1820, 1951–1970. [Google Scholar] [CrossRef] [PubMed]
- Yuan, G.; Wahlqvist, M.L.; Yuan, J.Q.; Wangband, Q.-M.; Li, D. Effect of punicic acid naturally occurring in food on lipid peroxidation in healthy young humans. J. Sci. Food Agric. 2009, 89, 2331–2335. [Google Scholar] [CrossRef]
- Olszanecki, R.; Jawien, J.; Gajda, M.; Mateuszuk, L.; Gebska, A.; Korabiowska, M.; Chłopicki, S.; Korbut, R. Effect of curcumin on atherosclerosis in apoE/LDLR-double knockout mice. J. Physiol. Pharmacol. 2005, 56, 627–635. [Google Scholar] [PubMed]
- Jawien, J.; Gajda, M.; Rudling, M.; Mateuszuk, L.; Olszanecki, R.; Guzik, T.J.; Cichocki, T.; Chlopicki, S.; Korbut, R. Inhibition of five lipoxygenase activating protein (FLAP) by MK-886 decreases atherosclerosis in apoE/LDLR-double knockout mice. Eur. J. Clin. Investig. 2006, 36, 141–146. [Google Scholar] [CrossRef]
- Jawien, J.; Csanyi, G.; Gajda, M.; Mateuszuk, L.; Lomnicka, M.; Korbut, R.; Chlopicki, S. Ticlopidine attenuates progression of atherosclerosis in apolipoprotein E and low density lipoprotein receptor double knockout mice. Eur. J. Pharmacol. 2006, 556, 129–135. [Google Scholar] [CrossRef]
- Arbonés-Mainar, J.M.; Navarro, M.A.; Acín, S.; Guzmán, M.A.; Arnal, C.; Surra, J.C.; Carnicer, R.; Roche, H.M.; Osada, J. Trans-10, cis-12- and cis-9, trans-11-Conjugated Linoleic Acid Isomers Selectively Modify HDL-Apolipoprotein Composition in Apolipoprotein E Knockout Mice. J. Nutr. 2006, 136, 353–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nestel, P.; Fujii, A.; Allen, T. The cis-9,trans-11 isomer of conjugated linoleic acid (CLA) lowers plasma triglyceride and raises HDL cholesterol concentrations but does not suppress aortic atherosclerosis in diabetic apoE-deficient mice. Atherosclerosis 2006, 189, 282–287. [Google Scholar] [CrossRef]
- Franczyk-Żarów, M.; Kostogrys, R.B.; Szymczyk, B.; Jawień, J.; Gajda, M.; Cichocki, T.; Wojnar, L.; Chlopicki, S.; Pisulewski, P.M. Functional effects of eggs, naturally enriched with conjugated linoleic acid (CLA), on the blood lipid profile, development of atherosclerosis and composition of atherosclerotic plaque in apolipoprotein E and low density lipoprotein receptor double-knockout mice (apoE/LDLR−/−). Brit. J. Nutr. 2008, 99, 49–58. [Google Scholar] [PubMed] [Green Version]
- Franczyk-Żarów, M.; Czyżyńska, I.; Drahun, A.; Maślak, E.; Chłopicki, S.; Kostogrys, R.B. Margarine supplemented with conjugated linolenic acid (CLnA) has no effect on atherosclerosis but alleviates the liver steatosis and affects the expression of lipid metabolism genes in apoE/LDLR−/− mice. Eur. J. Lipid Sci. Technol. 2015, 117, 589–600. [Google Scholar] [CrossRef]
- Man, J.J.; Beckman, J.A.; Jaffe, I.Z. Sex as a Biological Variable in Atherosclerosis. Circ Res. 2020, 24, 1297–1319. [Google Scholar] [CrossRef] [PubMed]
- Csányi, G.; Gajda, M.; Franczyk-Zarow, M.; Kostogrys, R.; Gwoźdź, P.; Mateuszuk, Ł.; Sternak, M.; Wojcik, L.; Zalewska, T.; Walski, M.; et al. Functional alterations in endothelial NO, PGI₂ and EDHF pathways in aorta in ApoE/LDLR−/− mice. Prostaglandins Other Lipid Mediat. 2012, 98, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C. AIN-93 purified diets for laboratory rodents: Final report of the American Institute of Nutrition ad hoc Writing Committee on the reformulation of the AIN-76A rodent diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [CrossRef]
- Kiernan, J.A. Histological & Histochemical Methods. Theory & Practice, 2nd ed.; Pergamon Press, Great Britain BPCC, Wheatons Ltd.: Exeter, UK, 1990; pp. 413–421. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | LnA | CLnA | CLA | |
|---|---|---|---|---|
| Corn starch | 532.486 | 532.486 | 532.486 | 532.486 |
| Caseine | 200 | 200 | 200 | 200 |
| Sucrose | 100 | 100 | 100 | 100 |
| Soybean oil | 70 | 59.75 | 61.3 | 63.34 |
| Celulose powder | 50 | 50 | 50 | 50 |
| Mineral mixture a | 35 | 35 | 35 | 35 |
| Vitamin mixture b | 10 | 10 | 10 | 10 |
| Choline | 2.5 | 2.5 | 2.5 | 2.5 |
| tert-butylhydrochinon | 0.014 | 0.014 | 0.014 | 0.014 |
| Flaxseed oil c | - | 10.25 | - | - |
| Pomegranate seed oil d | - | - | 8.7 | - |
| cis-9,trans-11 CLA e | - | - | - | 6.66 |
| SHORT | LONG | |||||||
|---|---|---|---|---|---|---|---|---|
| Control | LnA | CLnA | CLA | Control | LnA | CLnA | CLA | |
| Initial body weight (g) | 23.06 ± 1.02 | 21.77 ± 1.68 | 22.91 ± 2.04 | 22.17 ± 1.62 | 17.82 ± 1.45 | 18.97 ± 1.06 | 18.13 ± 1.83 | 17.46 ± 1.51 |
| Final body weight (g) | 23.25 ± 0.74 a | 22.13 ± 2.32 a | 22.97 ± 2.44 a | 21.94 ± 0.97 a | 21.58 ± 1.56 A | 23.17 ± 1.25 B | 21.97 ± 1.20 AB | 21.24 ± 2.09 A |
| Liver weight (g/100g BW) | 4.45 ± 0.35 a | 4.19 ± 0.45 a | 4.70 ± 0.58 a | 4.58 ± 0.52 a | 4.28 ± 0.54 A | 4.36 ± 0.39 A | 4.61 ± 0.61 A | 4.37 ± 0.46 A |
| Final glucose (mg/dL) | 135.2 ± 12.5 a | 144.1 ± 25.2 a | 140.2 ± 19.3 a | 122.8 ± 15.4 a | 120.4 ± 11.6 A | 123.6 ± 14.8 A | 133.0 ± 13.7 A | 123.2 ± 9.4 A |
| TCh (mmol/L) | 28.59 ± 5.06 b | 23.67 ± 2.45 ab | 19.93 ± 5.29 a | 21.84 ± 0.58 a | 20.60 ± 1.24 A | 20.78 ± 2.49 A | 21.05 ± 2.45 A | 21.82 ± 1.69 A |
| LDL + VLDL (mmol/L) | 14.51 ± 1.97 c | 12.14 ± 2.19 bc | 6.94 ± 0.73 a | 10.57 ± 1.10 ab | 8.31 ± 0.76 A | 8.56 ± 1.51 A | 8.90 ± 1.50 A | 9.20 ± 1.27 A |
| HDL (mmol/L) | 0.94 ± 0.22 a | 0.73 ± 0.15 a | 0.78 ± 0.13 a | 0.65 ± 0.03 a | 0.74 ± 0.13 A | 0.78 ± 0.15 A | 0.78 ± 0.19 A | 0.80 ± 0.16 A |
| TAG (mmol/L) | 3.76 ± 1.59 a | 2.27 ± 0.30 a | 1.92 ± 0.09 a | 2.46 ± 0.80 a | 1.85 ± 0.52 A | 2.04 ± 0.81 A | 1.96 ± 0.74 A | 2.12 ± 0.95 A |
| SHORT | LONG | |||||||
|---|---|---|---|---|---|---|---|---|
| Control | LnA | CLnA | CLA | Control | LnA | CLnA | CLA | |
| PPARα (a.u.) | 1.00 | 1.32 | 3.77 | 1.94 | 1.00 | 1.97 | 0.93 | 2.58 |
| SREBP-1 (a.u.) | 1.00 | 1.49 | 3.25 | 1.23 | 1.00 | 0.73 | 0.78 | 3.14 |
| ACO (a.u.) | 1.00 | 0.72 | 2.83 | 0.66 | 1.00 | 1.01 | 1.11 | 1.71 |
| FAS (a.u.) | 1.00 | 0.72 | 2.83 | 0.66 | 1.00 | 0.69 | 0.56 | 1.16 |
| Fatty Acid | SHORT | LONG | ||||||
|---|---|---|---|---|---|---|---|---|
| Control | LnA | CLnA | CLA | Control | LnA | CLnA | CLA | |
| C 14:0 | 1.37 ± 0.24 a | 1.44 ± 0.20 a | 1.34 ± 0.27 a | 1.30 ± 0.30 a | 1.15 ± 0.09 A | 1.29 ± 0.15 A | 1.30 ± 0.11 AB | 1.47 ± 0.18 B |
| C 16:0 | 16.93 ± 1.32 a | 17.33 ± 1.29 a | 18.31 ± 1.26 a | 17.50 ± 1.17 a | 17.01 ± 1.04 AB | 15.71 ± 0.86 A | 16.74 ± 0.69 A | 18.74 ± 2.43 B |
| C 16:1 | 4.16 ± 0.61 a | 4.43 ± 0.80 ab | 4.63 ± 0.61 ab | 5.21 ± 0.78 ab | 4.13 ± 0.84 A | 3.94 ± 0.54 A | 4.64 ± 0.90 A | 5.62 ± 0.27 B |
| C 18:0 | 4.42 ± 0.42 b | 4.39 ± 0.37 b | 3.44 ± 0.57 a | 2.88 ± 0.65 a | 3.37 ± 0.58 A | 4.12 ± 0.51 A | 3.51 ± 0.62 A | 2.48 ± 0.79 B |
| C 18:1 | 35.45 ± 1.01 b | 34.73 ± 1.34 ab | 33.41 ± 1.04 a | 33.44 ± 1.61 a | 37.23 ± 0.95 BC | 38.09 ± 1.03 C | 35.77 ± 1.01 AB | 35.57 ± 1.69 A |
| C 18:2 | 34.09 ± 1.92 b | 31.25 ± 1.56 a | 32.52 ± 2.17 ab | 32.78 ± 1.95 ab | 33.79 ± 1.46 B | 30.77 ± 0.89 A | 30.74 ± 1.01 A | 30.47 ± 2.14 A |
| C 20:0 | 0.30 ± 0.05 a | 0.28 ± 0.07 a | 0.24 ± 0.06 a | 0.22 ± 0.08 a | 0.23 ± 0.04 A | 0.30 ± 0.05 C | 0.23 ± 0.06 A | 0.17 ± 0.03 B |
| C 20:1 | 1.12 ± 0.19 b | 0.88 ± 0.17 a | 0.82 ± 0.09 a | 0.83 ± 0.11 a | 0.83 ± 0.15 A | 1.03 ± 0.10 C | 0.85 ± 0.18 A | 0.62 ± 0.08 B |
| C 18:3 | 2.17 ± 0.32 b | 5.28 ± 0.48 c | 1.64 ± 0.11 a | 1.89 ± 0.22 ab | 1.85 ± 0.19 A | 4.77 ± 0.74 B | 1.79 ± 0.33 A | 1.37 ± 0.17 A |
| CLA cis-9,trans-11 | 1.76 ± 0.16 a | 3.33 ± 0.30 b | 0.37 ± 0.14 A | 2.24 ± 0.24 B | 2.89 ± 0.26 C | |||
| CLA trans-10,cis-12 | 0.52 ± 0.11 a | 0.65 ± 0.10 b | 0.16 ± 0.04 B | 0.69 ± 0.16 A | 0.59 ± 0.09 A | |||
| CLnA cis-9,trans-11,cis-13 | 1.37 ± 0.27 | 1.51 ± 0.47 | ||||||
| SFA | 23.01 ± 1.70 a | 23.43 ± 1.50 a | 23.33 ± 1.73 a | 21.87 ± 1.85 a | 21.76 ± 1.39 A | 21.41 ± 1.25 A | 21.77 ± 0.89 A | 22.86 ± 3.50 A |
| MUFA | 40.72 ± 2.69 a | 40.04 ± 2.77 ab | 38.85 ± 2.77 ab | 39.48 ± 2.17 b | 42.19 ± 2.83 A | 43.06 ± 2.10 AB | 41.25 ± 1.65 AB | 41.81 ± 2.03 B |
| PUFA | 36.27 ± 1.97 a | 36.54 ± 1.58 ab | 37.82 ± 2.13 ab | 38.65 ± 1.82 b | 36.16 ± 1.41 A | 35.53 ± 1.49 A | 36.97 ± 0.93 A | 35.33 ± 2.43 A |
| 16:1/16:0 | 0.25 ± 0.02 a | 0.26 ± 0.05 ab | 0.25 ± 0.03 ab | 0.30 ± 0.04 b | 0.24 ± 0.05 A | 0.25 ± 0.04 AB | 0.28 ± 0.05 AB | 0.30 ± 0.04 B |
| 18:1/18:0 | 8.03 ± 0.77 a | 7.91 ± 0.72 ab | 9.72 ± 1.41 b | 11.60 ± 2.52 c | 11.05 ± 1.80 A | 9.25 ± 1.07 A | 10.20 ± 1.71 A | 14.34 ± 4.21 B |
| 16:0/18:2 | 0.50 ± 0.06 a | 0.55 ± 0.06 a | 0.56 ± 0.06 a | 0.53 ± 0.06 a | 0.50 ± 0.04 A | 0.51 ± 0.04 A | 0.54 ± 0.03 A | 0.62 ± 0.13 B |
| Fatty Acid | SHORT | LONG | ||||||
|---|---|---|---|---|---|---|---|---|
| Control | LnA | CLnA | CLA | Control | LnA | CLnA | CLA | |
| C 14:0 | 0.56 ± 0.11 ab | 0.48 ± 0.05 ab | 0.38 ± 0.07 a | 0.61 ± 0.09 b | 0.48 ± 0.05 A | 0.45 ± 0.07 A | 0.52 ± 0.13 A | 0.52 ± 0.17 A |
| C 16:0 | 25.80 ± 0.48 ab | 25.83 ± 1.74 ab | 24.67 ± 1.11 a | 28.00 ± 0.41 b | 24.51 ± 2.74 A | 24.41 ± 0.88 A | 22.95 ± 2.13 A | 24.33 ± 4.32 A |
| C 16:1 | 2.21 ± 0.21 bc | 1.74 ± 0.18 ab | 1.25 ± 0.24 a | 2.39 ± 0.21 c | 1.61 ± 0.23 A | 1.63 ± 0.58 A | 2.13 ± 0.56 A | 2.54 ± 0.48 A |
| C 18:0 | 4.23 ± 0.51 a | 4.26 ± 0.74 a | 3.84 ± 0.52 a | 3.47 ± 0.27 a | 4.31 ± 1.06 A | 3.47 ± 0.87 A | 5.15 ± 0.49 A | 3.97 ± 0.88 A |
| C 18:1 | 30.64 ± 0.30 a | 31.63 ± 3.44 a | 35.72 ± 2.42 a | 30.46 ± 0.85 a | 34.52 ± 3.53 A | 31.44 ± 2.15 A | 33.10 ± 1.81 A | 32.34 ± 1.23 A |
| C 18:2 | 35.13 ± 1.17 b | 32.97 ± 0.65 ab | 31.63 ± 1.66 a | 32.34 ± 0.77 ab | 32.92 ± 2.23 A | 34.60 ± 2.26 A | 31.72 ± 1.96 A | 33.12 ± 2.68 A |
| C 18:3 | 1.44 ± 0.03 a | 3.08 ± 0.59 b | 1.18 ± 0.23 a | 1.27 ± 0.04 a | 1.66 ± 0.47 A | 4.01 ± 0.52 B | 1.82 ± 0.17 A | 1.79 ± 0.32 A |
| CLA cis-9,trans-11 | 1.34 ± 0.30 a | 1.31 ± 0.03 a | 1.62 ± 0.18 B | 1.20 ± 0.22 A | ||||
| CLA trans-10,cis-12 | 0.20 ± 0.05 A | 0.20 ± 0.02 A | ||||||
| CLnA cis-9,trans-11,cis-13 | 0.81 ± 0.11 | |||||||
| SFA | 30.59 ± 0.83 a | 30.57 ± 2.40 a | 28.88 ± 0.71 a | 32.08 ± 0.34 a | 29.30 ± 2.49 A | 28.33 ± 1.54 B | 28.62 ± 2.19 A | 28.82 ± 4.01 A |
| MUFA | 32.84 ± 0.50 a | 33.37 ± 3.29 a | 36.97 ± 2.18 a | 32.85 ± 1.05 a | 36.13 ± 3.57 A | 33.06 ± 1.83 A | 35.22 ± 1.80 A | 34.88 ± 0.87 A |
| PUFA | 36.57 ± 1.14 a | 36.06 ± 1.24 a | 34.15 ± 1.73 a | 34.92 ± 0.79 a | 34.58 ± 2.47 A | 38.61 ± 2.34 A | 36.17 ± 2.24 A | 36.30 ± 3.20 A |
| 16:1/16:0 | 0.09 ± 0.01 b | 0.07 ± 0.00 a | 0.05 ± 0.01 a | 0.09 ± 0.01 b | 0.07 ± 0.01 A | 0.07 ± 0.02 A | 0.09 ± 0.02 AB | 0.10 ± 0.01 B |
| 18:1/18:0 | 7.31 ± 0.97 a | 7.42 ± 1.93 a | 9.36 ± 0.70 a | 8.82 ± 0.90 a | 8.55 ± 2.94 A | 9.56 ± 2.95 A | 6.49 ± 0.88 A | 8.45 ± 1.85 A |
| 16:0/18:2 | 0.74 ± 0.04 a | 0.78 ± 0.05 ab | 0.78 ± 0.03 ab | 0.87 ± 0.03 b | 0.75 ± 0.10 A | 0.71 ± 0.07 A | 0.73 ± 0.11 A | 0.75 ± 0.18 A |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Franczyk-Żarów, M.; Tarko, T.; Drahun-Misztal, A.; Czyzynska-Cichon, I.; Kus, E.; Kostogrys, R.B. Pomegranate Seed Oil as a Source of Conjugated Linolenic Acid (CLnA) Has No Effect on Atherosclerosis Development but Improves Lipid Profile and Affects the Expression of Lipid Metabolism Genes in apoE/LDLR−/− Mice. Int. J. Mol. Sci. 2023, 24, 1737. https://doi.org/10.3390/ijms24021737
Franczyk-Żarów M, Tarko T, Drahun-Misztal A, Czyzynska-Cichon I, Kus E, Kostogrys RB. Pomegranate Seed Oil as a Source of Conjugated Linolenic Acid (CLnA) Has No Effect on Atherosclerosis Development but Improves Lipid Profile and Affects the Expression of Lipid Metabolism Genes in apoE/LDLR−/− Mice. International Journal of Molecular Sciences. 2023; 24(2):1737. https://doi.org/10.3390/ijms24021737
Chicago/Turabian StyleFranczyk-Żarów, Magdalena, Tomasz Tarko, Anna Drahun-Misztal, Izabela Czyzynska-Cichon, Edyta Kus, and Renata B. Kostogrys. 2023. "Pomegranate Seed Oil as a Source of Conjugated Linolenic Acid (CLnA) Has No Effect on Atherosclerosis Development but Improves Lipid Profile and Affects the Expression of Lipid Metabolism Genes in apoE/LDLR−/− Mice" International Journal of Molecular Sciences 24, no. 2: 1737. https://doi.org/10.3390/ijms24021737