
Biomimetic Nanopillar Silicon Surfaces Rupture Fungal Spores
 , , , and
, , , and
Abstract
:1. Introduction
2. Results
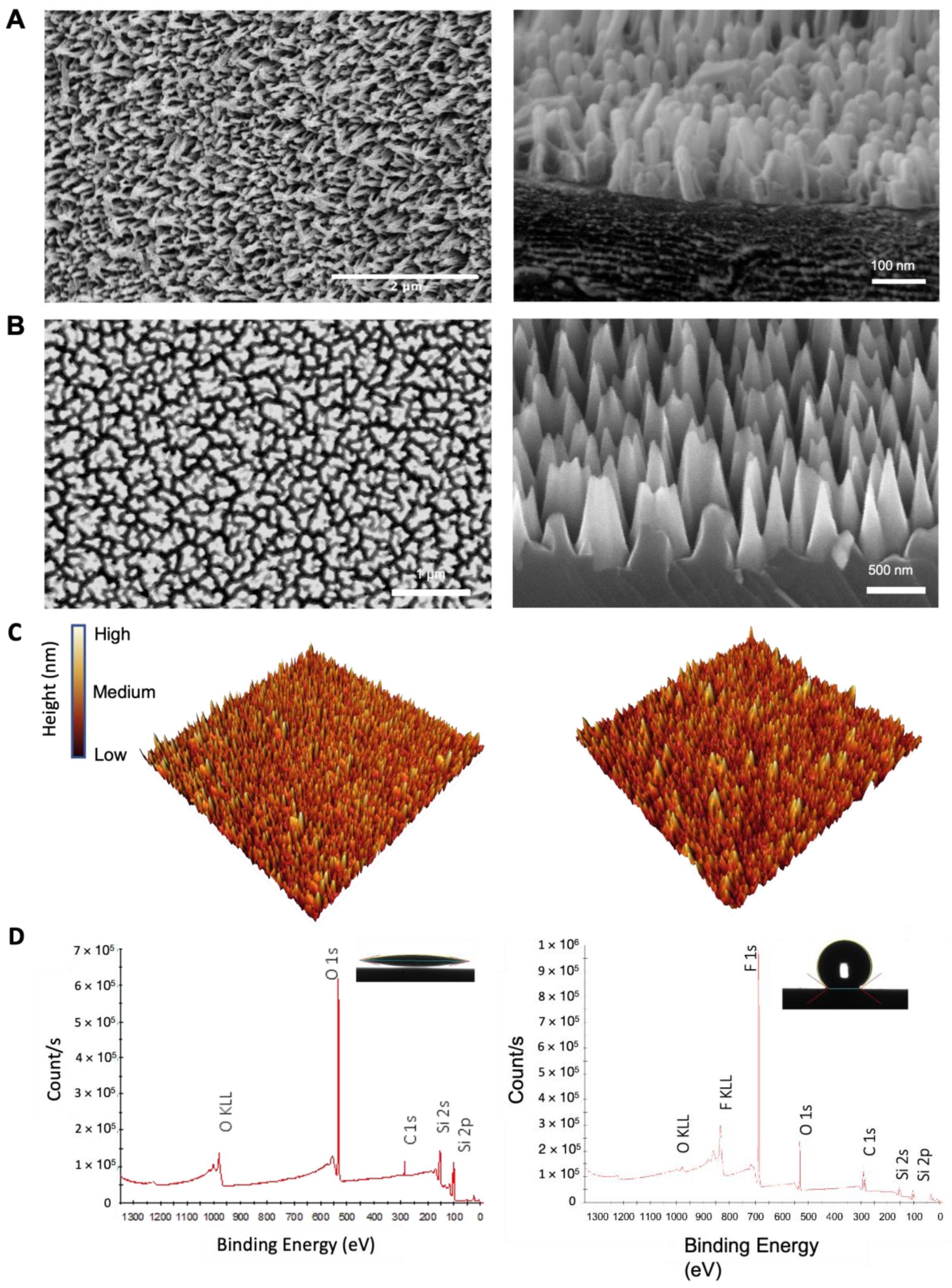
2.1. Surface Characterisation of Nano-Si Surfaces
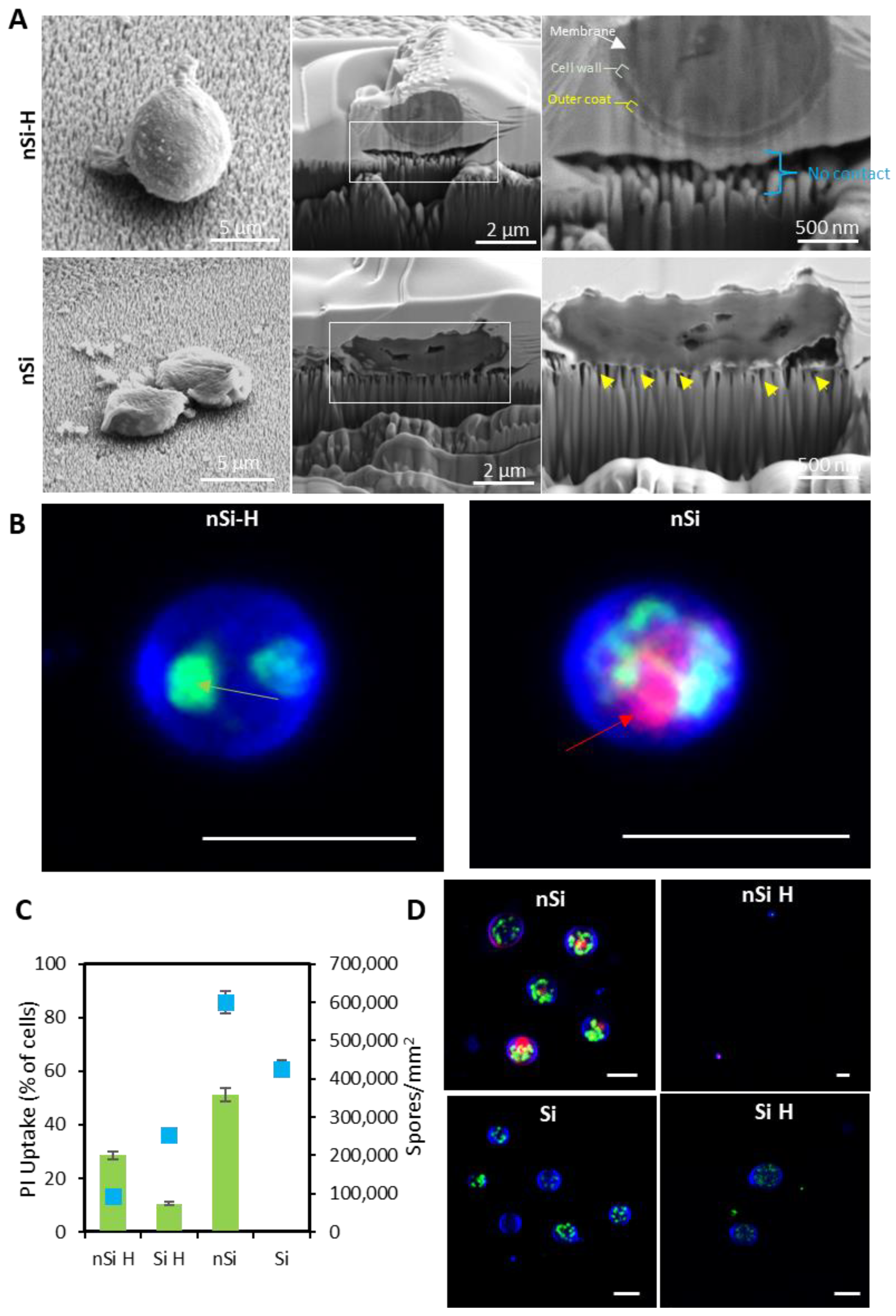
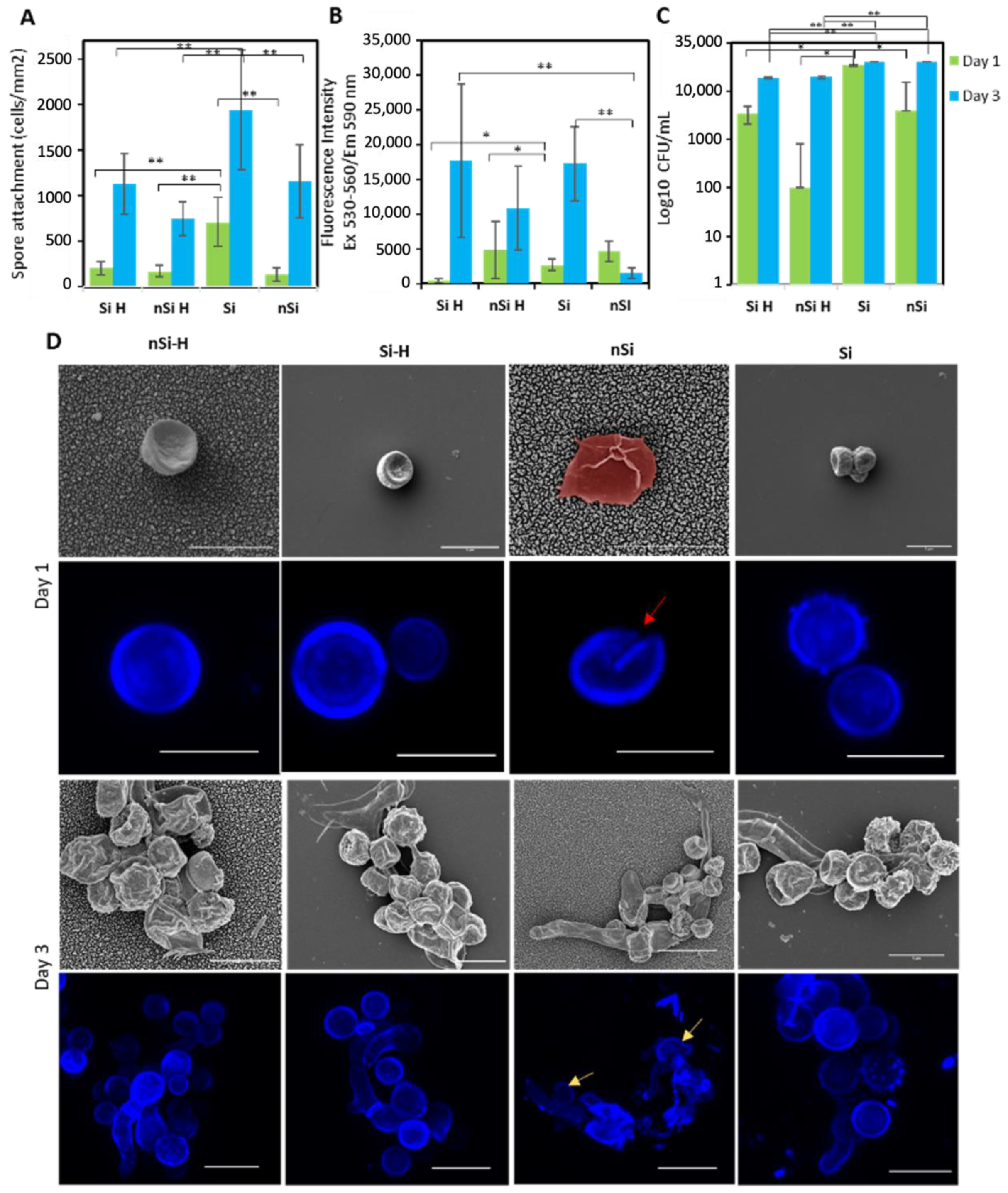
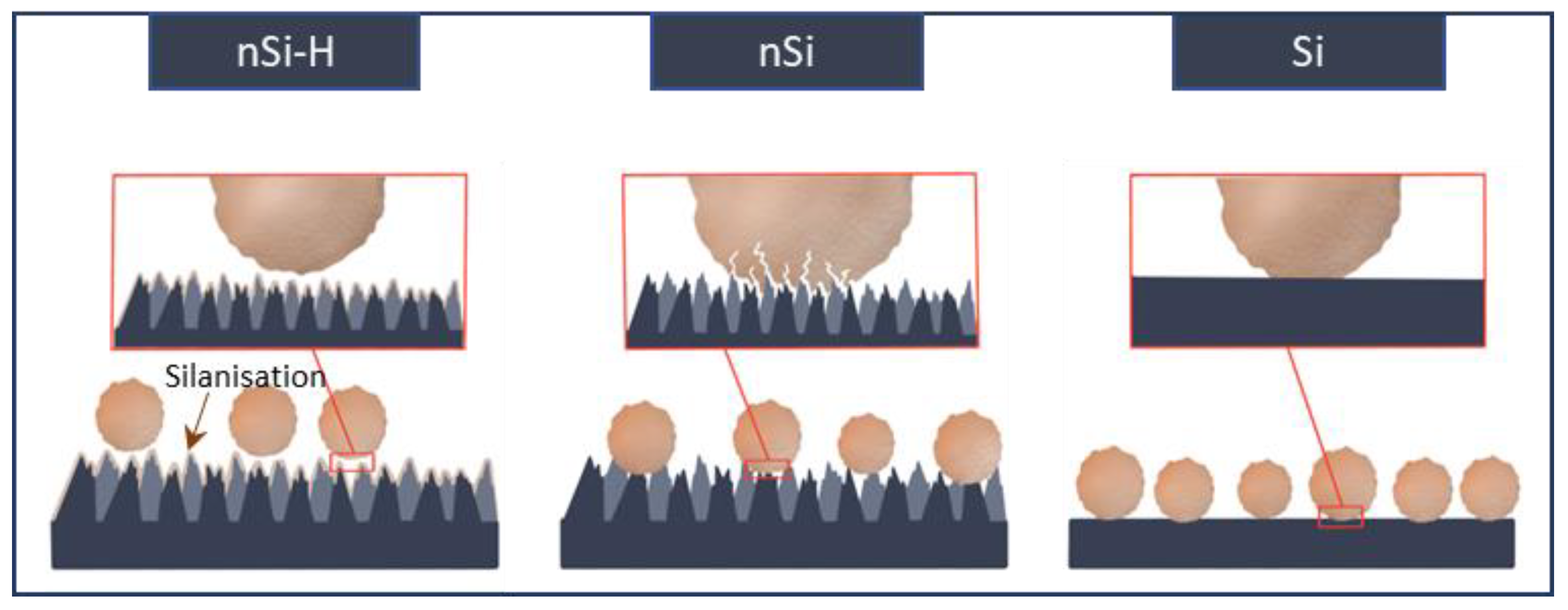
2.2. Interfacial Interactions of A. brasiliensis Spores with Hydrophilic and Hydrophobic Nano-Si Surfaces
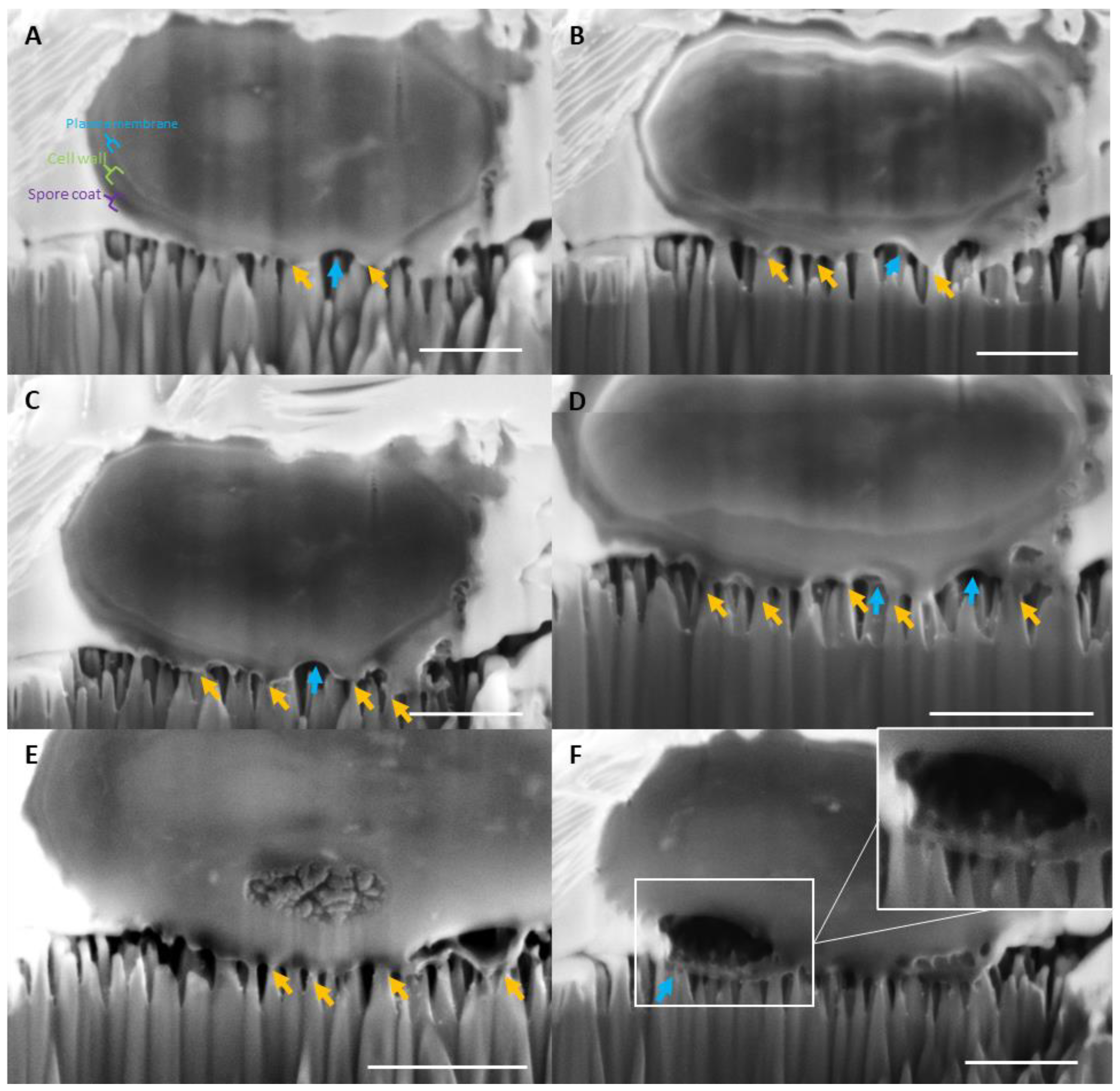
2.3. Assessment of the Structural Integrity, Morphology and Viability of the Fungal Spores during Interactions with Nano-Si Surfaces
3. Discussion
4. Materials and Methods
4.1. Fabrication of Nano-Si Surfaces
4.2. Atomic Force Microscopy (AFM)
4.3. Surface Wettability
4.4. X-ray Photoelectron Spectrometry
4.5. Microorganism, Growth Conditions and Experimental Set-Up
4.6. Confocal Laser Scanning Microscopy
4.7. Scanning Electron Microscopy (SEM)
4.8. Focused Ion Beam SEM
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Linklater, D.P.; Ivanova, E.P. Nanostructured antibacterial surfaces—What can be achieved? Nano Today 2022, 43, 101404. [Google Scholar] [CrossRef]
- Linklater, D.P.; Baulin, V.A.; Juodkazis, S.; Crawford, R.J.; Stoodley, P.; Ivanova, E.P. Mechano-bactericidal actions of nanostructured surfaces. Nat. Rev. Microbiol. 2020, 19, 8–22. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, E.P.; Linklater, D.P.; Werner, M.; Baulin, V.A.; Xu, X.; Vrancken, N.; Rubanov, S.; Hanssen, E.; Wandiyanto, J.; Truong, V.K.; et al. The multi-faceted mechano-bactericidal mechanism of nanostructured surfaces. Proc. Natl. Acad. Sci. USA 2020, 117, 12598–12605. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, E.P.; Hasan, J.; Webb, H.K.; Gervinskas, G.; Juodkazis, S.; Truong, V.K.; Wu, A.H.F.; Lamb, R.N.; Baulin, V.A.; Watson, G.S.; et al. Bactericidal activity of black silicon. Nat. Commun. 2013, 4, 2838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golan, J.J.; Pringle, A. Long-Distance Dispersal of Fungi. Microbiol. Spectr. 2017, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valsecchi, I.; Dupres, V.; Stephen-Victor, E.; Guijarro, J.; Gibbons, J.; Beau, R.; Bayry, J.; Coppee, J.-Y.; Lafont, F.; Latgé, J.-P.; et al. Role of Hydrophobins in Aspergillus fumigatus. J. Fungi 2017, 4, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beauvais, A.; Latgé, J.-P. Special Issue: Fungal Cell Wall. J. Fungi 2018, 4, 91. [Google Scholar] [CrossRef] [Green Version]
- Binder, U.; Lass-Florl, C. Epidemiology of invasive fungal infections in the mediterranean area. Mediterr. J. Hematol. Infect. Dis. 2011, 3, e20110016. [Google Scholar] [CrossRef] [Green Version]
- Damasceno, J.L.; Santos, R.A.d.; Barbosa, A.H.; Casemiro, L.A.; Pires, R.H.; Martins, C.H.G. Risk of Fungal Infection to Dental Patients. Sci. World J. 2017, 2017, 2982478. [Google Scholar] [CrossRef] [Green Version]
- Van Dijck, P.; Sjollema, J.; Camue, B.P.A.; Lagrou, K.; Berman, J.; d’Enfert, C.; Andes, D.R.; Arendrup, M.C.; Brakhage, A.A.; Calderone, R.; et al. Methodologies for in vitro and in vivo evaluation of efficacy of antifungal and antibiofilm agents and surface coatings against fungal biofilms. Microb. Cell 2018, 5, 300–326. [Google Scholar] [CrossRef]
- Verweij, P.E.; Chowdhary, A.; Melchers, W.J.G.; Meis, J.F. Azole Resistance inAspergillus fumigatus: Can We Retain the Clinical Use of Mold-Active Antifungal Azoles? Clin. Infect. Dis. 2015, 62, 362–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, G.D.; Denning, D.W.; Levitz, S.M. Tackling human fungal infections. Science 2012, 336, 647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beyki, A.; Zardast, M.; Nasrollahi, Z. Maxillary sinus aspergillosis: A case report of the timely failure to treatment. Iran. J Microbiol. 2019, 11, 345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urs, A.B.; Singh, H.; Nunia, K.; Mohanty, S.; Gupta, S. Post endodontic Aspergillosis in an immunocompetent individual. J. Clin. Exp. Dent. 2015, 7, e535. [Google Scholar] [CrossRef] [PubMed]
- Legent, F.; Billet, J.; Beauvillain, C.; Bonnet, J.; Miegeville, M. The role of dental canal fillings in the development of Aspergillus sinusitis. Arch. Oto-Rhino-Laryngol. 1989, 246, 318–320. [Google Scholar] [CrossRef]
- Mazari, W.; Boucherit-Otmani, Z.; Boucherit, K. In vitro susceptibility of amphotericin-B, voriconazole and caspofungin against Candida guilliermondii biofilms, isolated from dentals units water pipes, under different growth phases. J. Mycol. Méd. 2015, 25, 57–62. [Google Scholar] [CrossRef]
- ADA. Statement on dental unit waterlines. J. Am. Dent. Assoc. 1996, 127, 185–186. [Google Scholar] [CrossRef]
- Kadaifciler, D.G.; Ökten, S.; Sen, B. Mycological contamination in dental unit waterlines in Istanbul, Turkey. Braz. J. Microbiol. 2013, 44, 977–981. [Google Scholar] [CrossRef] [Green Version]
- Faustino, I.S.P.; Ramos, J.C.; Mariz, B.A.L.A.; Papadopoulou, E.; Georgaki, M.; Nikitakis, N.G.; Vargas, P.A.; Santos-Silva, A.R.; Lopes, M.A. A Rare Case of Mandibular Aspergillus Osteomyelitis in an Immunocompetent Patient. Dent. J. 2022, 10, 213. [Google Scholar] [CrossRef]
- Stacchi, C.; Del Lupo, V.; Berton, F.; Lombardi, T.; Bressan, R.; Di Lenarda, R.; Lagatolla, C. Aspergillus fumigatus biofilm formation on different bone substitutes used in maxillary sinus augmentation: An in vitro analysis. Int. J. Implant. Dent. 2019, 5, 22. [Google Scholar] [CrossRef]
- Pagella, F.; Matti, E.; Bernardi, F.D.; Semino, L.; Cavanna, C.; Marone, P.; Farina, C.; Castelnuovo, P. Paranasal sinus fungus ball: Diagnosis and management. Mycoses 2007, 50, 451–456. [Google Scholar] [CrossRef] [PubMed]
- Hill, B.R.; Watson Sr, T.F.; Triplett, B.L. Antimicrobial Microporous Coating. U.S. Patent No. 5,024,875, 18 June 1991. [Google Scholar]
- Khan, A.; Horner, C. Nanosized Metal and Metal Oxide Particles as a Biocides In roofing Coatings. U.S. Patent Application No. 11/454,335, 24 May 2007. [Google Scholar]
- Ivanov-Omskii, V.I.; Panina, L.K.; Yastrebov, S.G. Amorphous hydrogenated carbon doped with copper as antifungal protective coating. Carbon 2000, 38, 495–499. [Google Scholar] [CrossRef]
- Coad, B.R.; Griesser, H.J.; Peleg, A.Y.; Traven, A.J.P.p. Anti-infective surface coatings: Design and therapeutic promise against device-associated infections. PLoS Pathog. 2016, 12, e1005598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eversdijk, J.; Erich, S.; Hermanns, S.; Adan, O.; De Bolle, M.; De Meyer, K.; Bylemans, D.; Bekker, M.; Ten Cate, A.J.P.i.O.C. Development and evaluation of a biocide release system for prolonged antifungal activity in finishing materials. Prog. Org. Coat. 2012, 74, 640–644. [Google Scholar] [CrossRef]
- Griesser, S.S.; Jasieniak, M.; Coad, B.R.; Griesser, H.J. Antifungal coatings by caspofungin immobilization onto biomaterials surfaces via a plasma polymer interlayer. Biointerphases 2015, 10, 04A307. [Google Scholar] [CrossRef] [Green Version]
- Arreche, R.A.; Igal, K.; Bellotti, N.; Deyá, C.; Vázquez, P.G. Functionalized zirconia compounds as antifungal additives for hygienic waterborne coatings. Prog. Org. Coat. 2019, 128, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Knowles, B.R.; Yang, D.; Wagner, P.; Maclaughlin, S.; Higgins, M.J.; Molino, P.J. Zwitterion Functionalized Silica Nanoparticle Coatings: The Effect of Particle Size on Protein, Bacteria, and Fungal Spore Adhesion. Langmuir 2019, 35, 1335–1345. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-W.; Hwang, W. Fabrication of a superhydrophobic surface with fungus-cleaning properties on brazed aluminum for industrial application in heat exchangers. Appl. Surf. Sci. 2018, 442, 461–466. [Google Scholar] [CrossRef]
- Li, G.; Zhao, H.; Hong, J.; Quan, K.; Yuan, Q.; Wang, X. Antifungal graphene oxide-borneol composite. Colloids Surf. B 2017, 160, 220–227. [Google Scholar] [CrossRef]
- Aburto-Medina, A.; Le, P.H.; MacLaughlin, S.; Ivanova, E. Diversity of experimental designs for the fabrication of antifungal surfaces for the built environment. Appl. Microbiol. Biotechnol. 2021, 105, 2663–2674. [Google Scholar] [CrossRef]
- Ivanova, E.P.; Linklater, D.P.; Medina, A.A.; Le, P.; Baulin, V.A.; Khuong Duy Nguyen, H.; Curtain, R.; Hanssen, E.; Gervinskas, G.; Hock Ng, S.; et al. Antifungal versus antibacterial defence of insect wings. J. Colloid Interface Sci. 2021, 603, 886–897. [Google Scholar] [CrossRef] [PubMed]
- Elbourne, A.; Dupont, M.F.; Collett, S.; Truong, V.K.; Xu, X.; Vrancken, N.; Baulin, V.; Ivanova, E.P.; Crawford, R.J. Imaging the air-water interface: Characterising biomimetic and natural hydrophobic surfaces using in situ atomic force microscopy. J. Colloid Interface Sci. 2019, 536, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Byun, D.; Hong, J.; Saputra; Ko, J.H.; Lee, Y.J.; Park, H.C.; Byun, B.-K.; Lukes, J.R. Wetting Characteristics of Insect Wing Surfaces. J. Bionic Eng. 2009, 6, 63–70. [Google Scholar] [CrossRef]
- Truong, V.K.; Geeganagamage, N.M.; Baulin, V.A.; Vongsvivut, J.; Tobin, M.J.; Luque, P.; Crawford, R.J.; Ivanova, E.P. The susceptibility of Staphylococcus aureus CIP 65.8 and Pseudomonas aeruginosa ATCC 9721 cells to the bactericidal action of nanostructured Calopteryx haemorrhoidalis damselfly wing surfaces. Appl. Microbiol. Biotechnol. 2017, 101, 4683–4690. [Google Scholar] [CrossRef]
- Linklater, D.P.; Juodkazis, S.; Rubanov, S.; Ivanova, E.P. Comment on “Bactericidal Effects of Natural Nanotopography of Dragonfly Wing on Escherichia coli”. ACS Appl. Mater. Interfaces 2017, 9, 29387–29393. [Google Scholar] [CrossRef] [Green Version]
- Vellwock, A.E.; Yao, H. Biomimetic and bioinspired surface topographies as a green strategy for combating biofouling: A review. Bioinspir. Biomim. 2021, 16, 041003. [Google Scholar] [CrossRef]
- Vellwock, A.E.; Su, P.; Zhang, Z.; Feng, D.; Yao, H. Reconciling the Conflict between Optical Transparency and Fouling Resistance with a Nanowrinkled Surface Inspired by Zebrafish’s Cornea. ACS Appl. Mater. Interfaces 2022, 14, 7617–7625. [Google Scholar] [CrossRef]
- Linklater, D.P.; Nguyen, H.K.D.; Bhadra, C.M.; Juodkazis, S.; Ivanova, E.P. Influence of nanoscale topology on bactericidal efficiency of black silicon surfaces. Nanotechnology 2017, 28, 245301. [Google Scholar] [CrossRef]
- Žukauskas, A.; Malinauskas, M.; Kadys, A.; Gervinskas, G.; Seniutinas, G.; Kandasamy, S.; Juodkazis, S. Black silicon: Substrate for laser 3D micro/nano-polymerization. Opt. Express 2013, 21, 6901–6909. [Google Scholar] [CrossRef]
- Gervinskas, G.; Seniutinas, G.; Hartley, J.S.; Kandasamy, S.; Stoddart, P.R.; Fahim, N.F.; Juodkazis, S. Surface-enhanced Raman scattering sensing on black silicon. Ann. Phys. 2013, 525, 907–914. [Google Scholar] [CrossRef]
- Fan, H.; Guo, Z. Bioinspired surfaces with wettability: Biomolecule adhesion behaviors. Biomater. Sci. 2020, 8, 1502–1535. [Google Scholar] [CrossRef] [PubMed]
- Mosquera, S.; Stergiopoulos, I.; Leveau, J.H.J. Interruption of Aspergillus niger spore germination by the bacterially produced secondary metabolite collimomycin. Environ. Microbiol. Rep. 2020, 12, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Firstencel, H.; Butt, T.M.; Carruthers, R.I. A fluorescence microscopy method for determining the viability of entomophthoralean fungal spores. J. Invertebr. Pathol. 1990, 55, 258–264. [Google Scholar] [CrossRef]
- Rampersad, S.N. Multiple applications of Alamar Blue as an indicator of metabolic function and cellular health in cell viability bioassays. Sensors 2012, 12, 12347–12360. [Google Scholar] [CrossRef]
- Fung, D.Y.C. Biochemical and modern identification techniques—Introduction. In Encyclopedia of Food Microbiology, 2nd ed.; Batt, C.A., Tortorello, M.L., Eds.; Academic Press: Oxford, UK, 2014; pp. 223–231. [Google Scholar]
- Heedy, S.; Marshall, M.E.; Pineda, J.J.; Pearlman, E.; Yee, A.F. Synergistic Antimicrobial Activity of a Nanopillar Surface on a Chitosan Hydrogel. ACS Appl. Bio Mater. 2020, 3, 8040–8048. [Google Scholar] [CrossRef]
- Riduan, S.N.; Zhang, Y. Recent Advances of Zinc-based Antimicrobial Materials. Chem. Asian J. 2021, 16, 2588–2595. [Google Scholar] [CrossRef]
- Hayles, A.; Bright, R.; Wood, J.; Palms, D.; Zilm, P.; Brown, T.; Barker, D.; Vasilev, K. Spiked Nanostructures Disrupt Fungal Biofilm and Impart Increased Sensitivity to Antifungal Treatment. Adv. Mater. Interfaces 2022, 9, 2102353. [Google Scholar] [CrossRef]
- Valdez-Salas, B.; Beltran-Partida, E.; Nedev, N.; Ibarra-Wiley, R.; Salinas, R.; Curiel-Alvarez, M.; Valenzuela-Ontiveros, Y.; Perez, G. Controlled antifungal behavior on Ti6Al4V nanostructured by chemical nanopatterning. Mater. Sci. Eng. C Mater. Biol. Appl. 2019, 96, 677–683. [Google Scholar] [CrossRef]
- Beltrán-Partida, E.; Valdez-Salas, B.; Curiel-Álvarez, M.; Castillo-Uribe, S.; Escamilla, A.; Nedev, N. Enhanced antifungal activity by disinfected titanium dioxide nanotubes via reduced nano-adhesion bonds. Mater. Sci. Eng. C 2017, 76, 59–65. [Google Scholar] [CrossRef]
- Le, P.H.; Nguyen, D.H.K.; Medina, A.A.; Linklater, D.P.; Loebbe, C.; Crawford, R.J.; MacLaughlin, S.; Ivanova, E.P. Surface Architecture Influences the Rigidity of Candida albicans Cells. Nanomaterials 2022, 12, 567. [Google Scholar] [CrossRef]
- Xie, Y.; Pan, Y.; Cai, P. Cellulose-based antimicrobial films incroporated with ZnO nanopillars on surface as biodegradable and antimicrobial packaging. Food Chem. 2022, 368, 130784. [Google Scholar] [CrossRef] [PubMed]
- Najibi Ilkhechi, N.; Mozammel, M.; Yari Khosroushahi, A. Antifungal effects of ZnO, TiO2 and ZnO-TiO2 nanostructures on Aspergillus flavus. Pestic. Biochem. Physiol. 2021, 176, 104869. [Google Scholar] [CrossRef] [PubMed]
- Basu, P.; Chakraborty, J.; Ganguli, N.; Mukherjee, K.; Acharya, K.; Satpati, B.; Khamrui, S.; Mandal, S.; Banerjee, D.; Goswami, D.; et al. Defect-Engineered MoS2 Nanostructures for Reactive Oxygen Species Generation in the Dark: Antipollutant and Antifungal Performances. ACS Appl. Mater. Interfaces 2019, 11, 48179–48191. [Google Scholar] [CrossRef] [PubMed]
- Rosenzweig, R.; Marshall, M.; Parivar, A.; Ly, V.K.; Pearlman, E.; Yee, A.F. Biomimetic Nanopillared Surfaces Inhibit Drug Resistant Filamentous Fungal Growth. ACS Appl. Bio Mater. 2019, 2, 3159–3163. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Rubio, R.; de Oliveira, H.C.; Rivera, J.; Trevijano-Contador, N. The Fungal Cell Wall: Candida, Cryptococcus, and Aspergillus Species. Front. Microbiol. 2019, 10, 2993. [Google Scholar] [CrossRef] [PubMed]
- Kang, X.; Kirui, A.; Muszynski, A.; Widanage, M.C.D.; Chen, A.; Azadi, P.; Wang, P.; Mentink-Vigier, F.; Wang, T. Molecular architecture of fungal cell walls revealed by solid-state NMR. Nat. Commun. 2018, 9, 2747. [Google Scholar] [CrossRef] [Green Version]
- Krasowska, A.; Sigler, K. How microorganisms use hydrophobicity and what does this mean for human needs? Front. Cell. Infect. Microbiol. 2014, 4, 112. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Surface Roughness Parameters | Sa | Sq | Smax |
|---|---|---|---|
| nSi-H | 29.16 ± 0.42 | 36.97 ± 0.59 | 323.02 ± 10.71 |
| Si-H | 7.60 ± 2.50 | 9.64 ± 3.10 | 86.94 ± 32.75 |
| nSi | 35.03 ± 0.59 | 44.12 ± 0.56 | 380.18 ± 32.75 |
| Si | 14.25 ± 4.54 | 17.58 ± 4.31 | 130.52 ± 34.80 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Linklater, D.P.; Le, P.H.; Aburto-Medina, A.; Crawford, R.J.; Maclaughlin, S.; Juodkazis, S.; Ivanova, E.P. Biomimetic Nanopillar Silicon Surfaces Rupture Fungal Spores. Int. J. Mol. Sci. 2023, 24, 1298. https://doi.org/10.3390/ijms24021298
Linklater DP, Le PH, Aburto-Medina A, Crawford RJ, Maclaughlin S, Juodkazis S, Ivanova EP. Biomimetic Nanopillar Silicon Surfaces Rupture Fungal Spores. International Journal of Molecular Sciences. 2023; 24(2):1298. https://doi.org/10.3390/ijms24021298
Chicago/Turabian StyleLinklater, Denver P., Phuc H. Le, Arturo Aburto-Medina, Russell J. Crawford, Shane Maclaughlin, Saulius Juodkazis, and Elena P. Ivanova. 2023. "Biomimetic Nanopillar Silicon Surfaces Rupture Fungal Spores" International Journal of Molecular Sciences 24, no. 2: 1298. https://doi.org/10.3390/ijms24021298