
Translocation of Methionine Adenosyl Transferase MAT2A and Its Prognostic Relevance for Liver Hepatocellular Carcinoma

Abstract
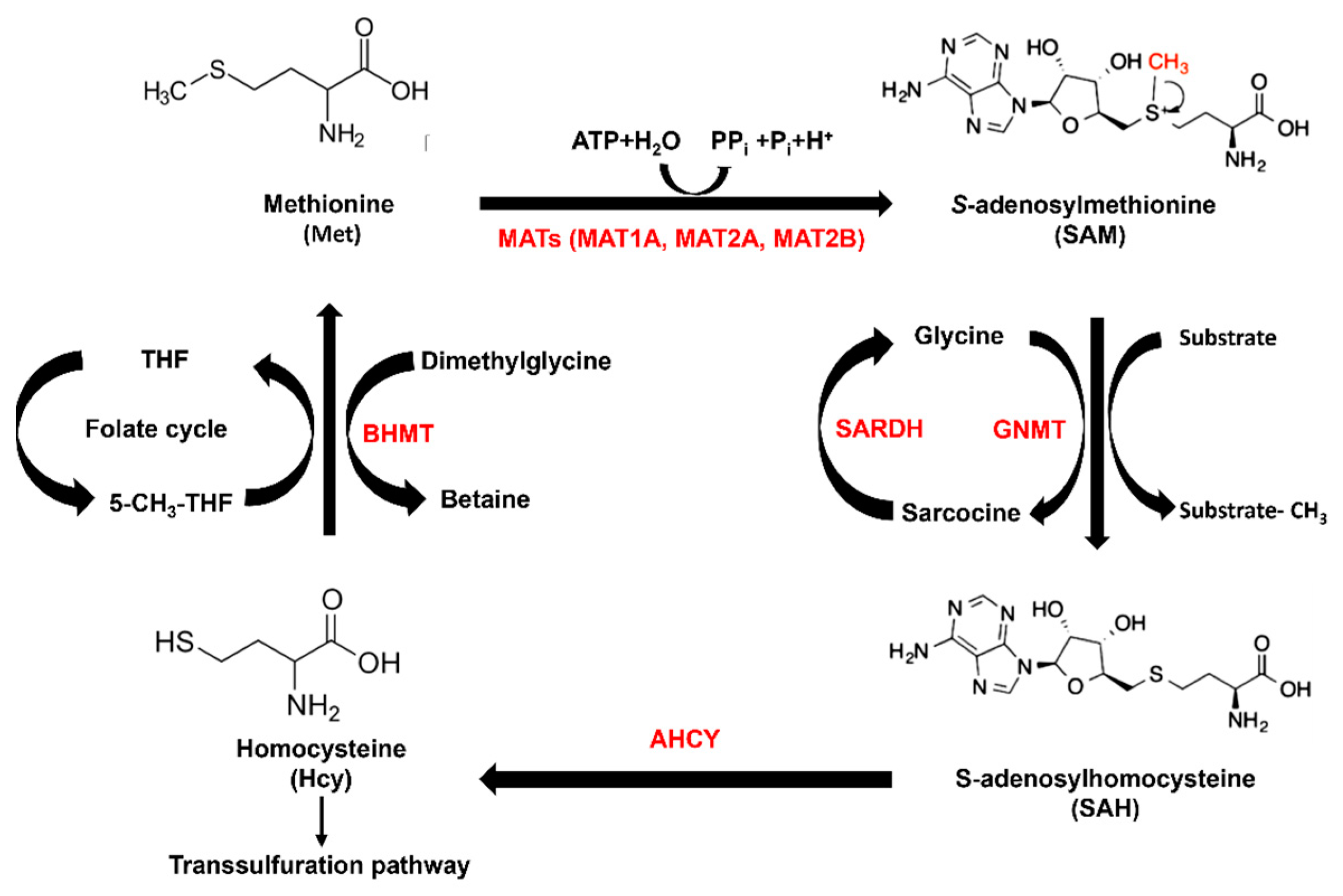
:1. Introduction
2. Results
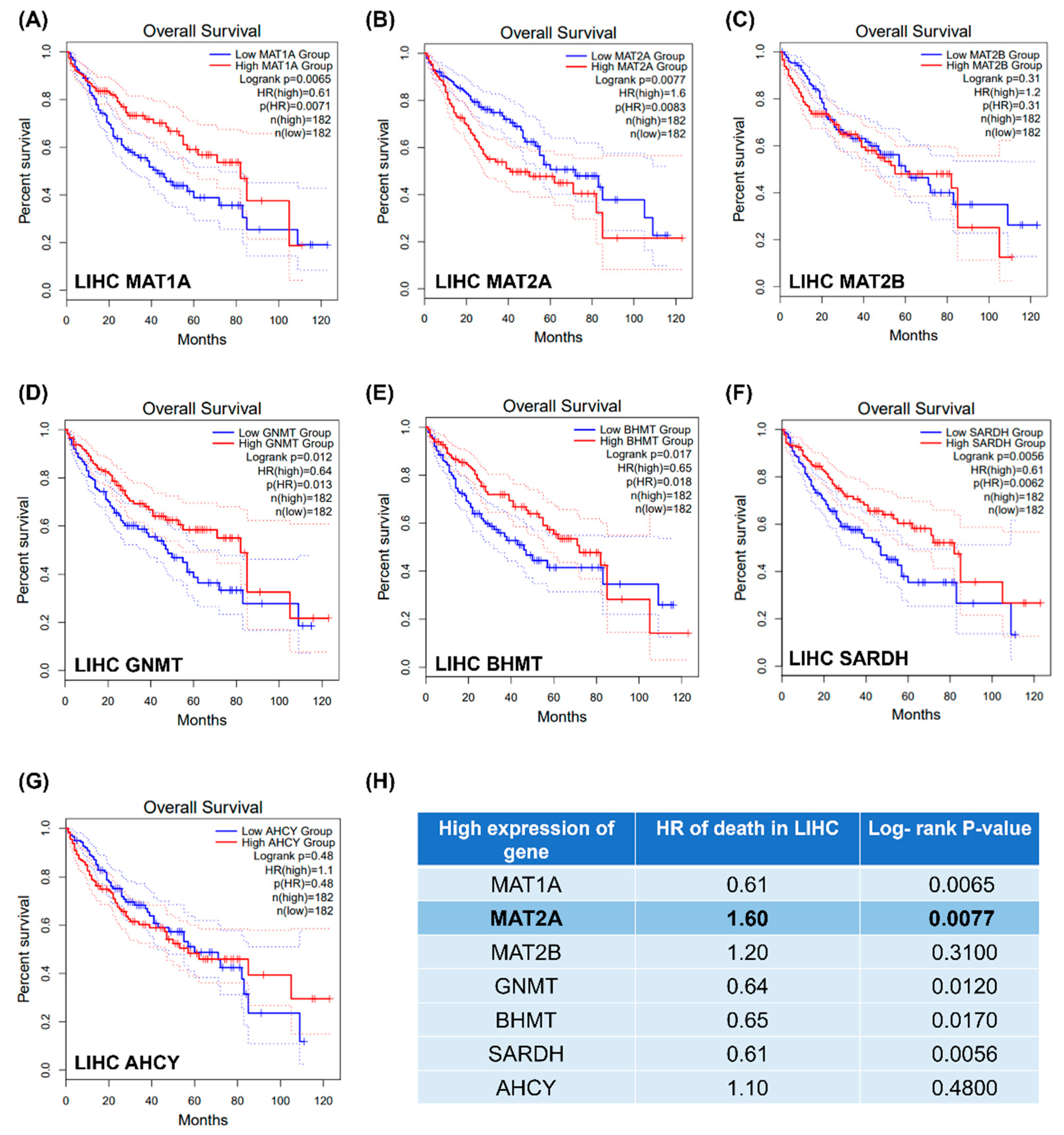
2.1. Gene Expression of Methionine Cycle Enzymes and LIHC Overall Survival
2.2. MAT2A mRNA Expression in Common Cancers
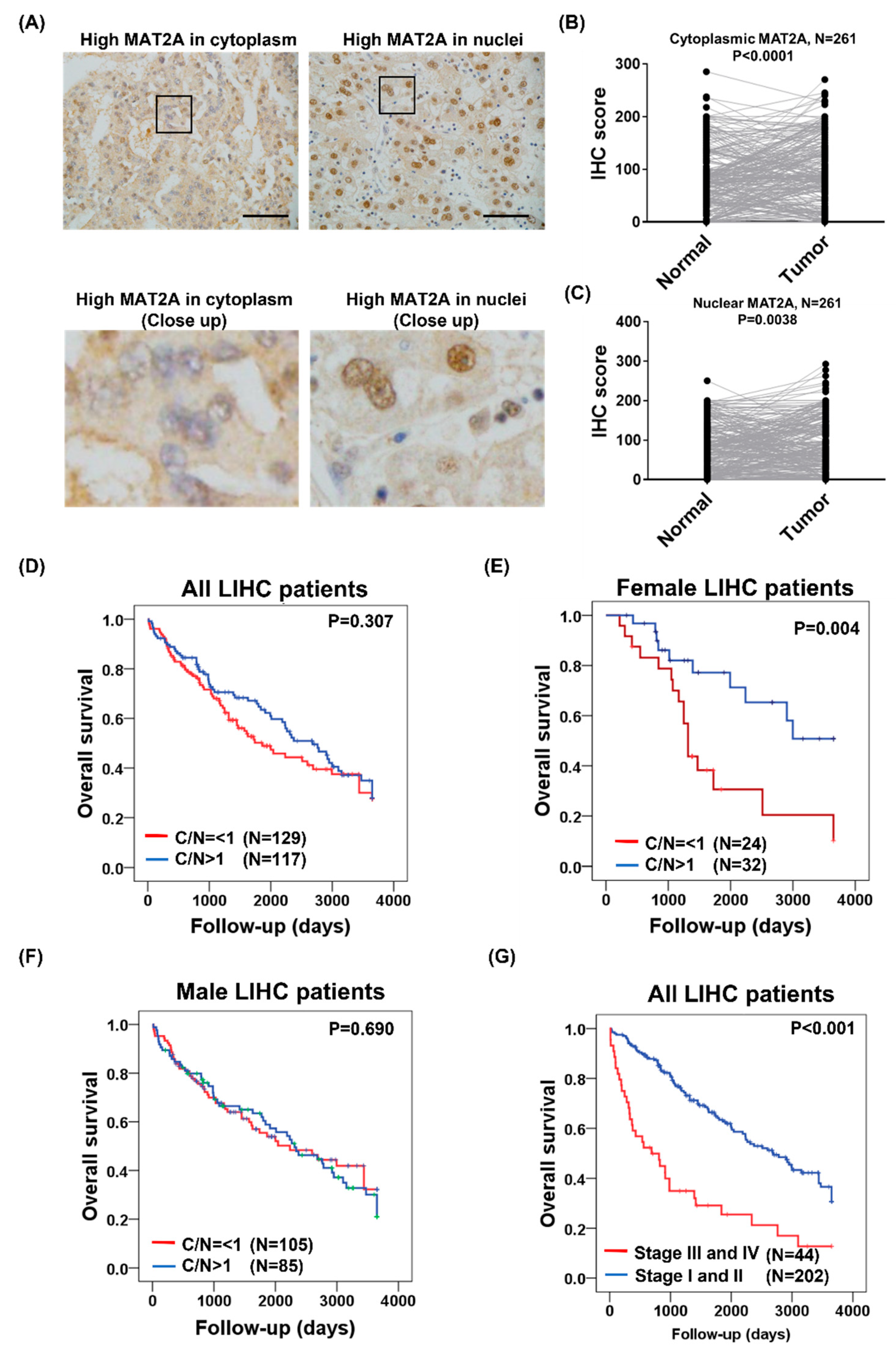
2.3. Subcellular MAT2A Distributions in Tumor and Adjacent Normal Tissues
2.4. Subcellular MAT2A Distributions Were Associated with LIHC Survival in Females
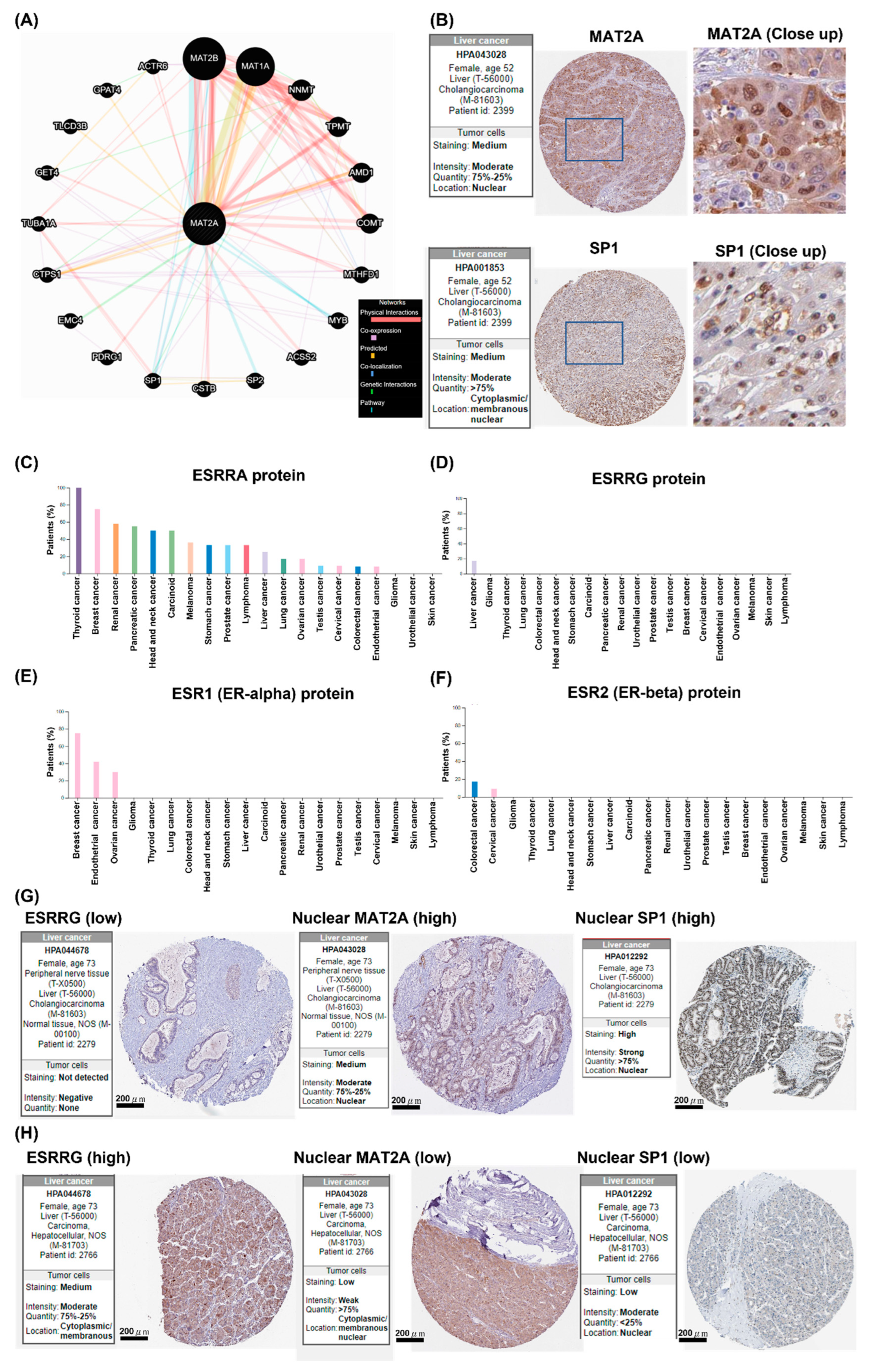
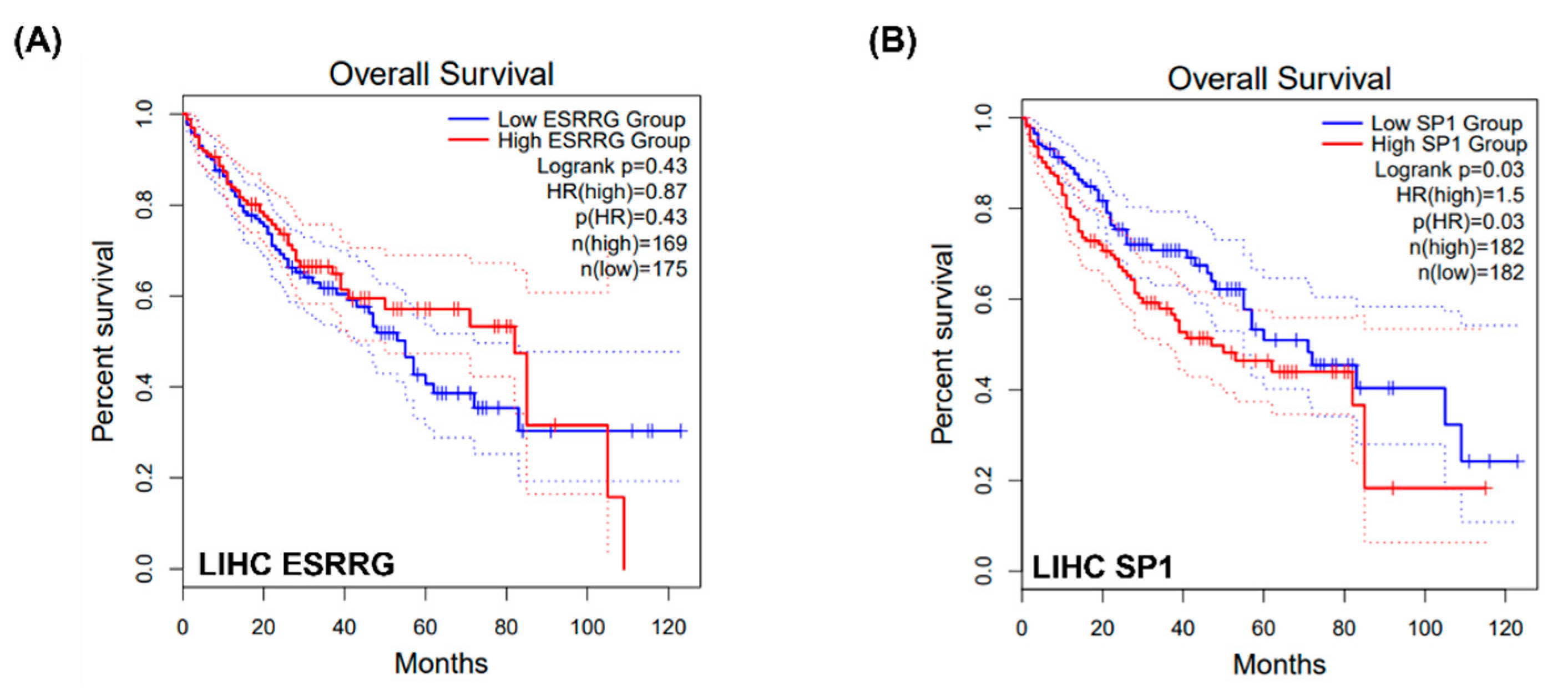
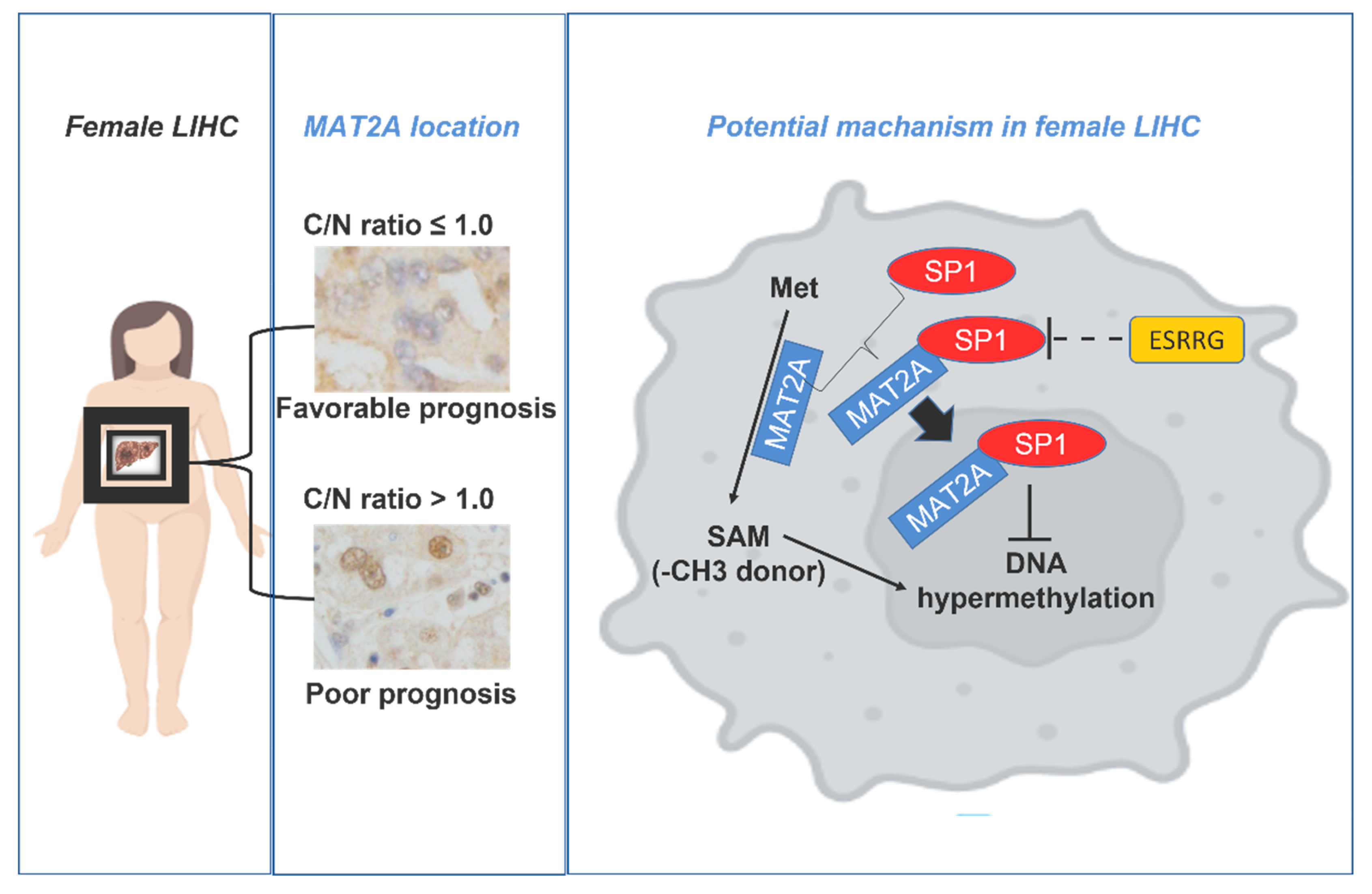
2.5. Potential Interactions between MAT2A and SP1, WhichIs Negatively Associated with ESSRG
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Immunohistochemistry and Scoring
4.3. Correlation Analysis
4.4. Protein Analysis
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| MAT | Methionine adenosyl transferases |
| SAM | Adenosylmethionine |
| GEPIA | Gene expression profiling interactive analysis |
| HPA | Human Protein Atlas |
| LIHC | Liver hepatocellular carcinoma |
| C/N | Cytoplasmic and nuclear protein expression ratio |
| BMHT | Methyl transferase enzyme |
| GNMT | Glycine N-methyltransferase |
| SARDH | Sarcosine dehydrogenase |
| AHCY | Adenosylhomocysteinase |
| SP1 | Specificity protein 1 |
| PDRG1 | P53 and DNA damage-regulated 1 |
| ESSRG | Estrogen-related receptor G |
References
- Forner, A.; Reig, M.; Bruix, J. Hepatocellular carcinoma. Lancet 2018, 391, 1301–1314. [Google Scholar] [CrossRef] [PubMed]
- Sakata, S.F.; Okumura, S.; Matsuda, K.; Horikawa, Y.; Maeda, M.; Kawasaki, K.; Chou, J.Y.; Tamaki, N. Effect of fasting on methionine adenosyltransferase expression and the methionine cycle in the mouse liver. J. Nutr. Sci. Vitaminol. 2005, 51, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Nordgren, K.K.; Chai, Y.; Hebbring, S.J.; Jenkins, G.D.; Abo, R.P.; Peng, Y.; Pelleymounter, L.L.; Moon, I.; Eckloff, B.W.; et al. Human liver methionine cycle: MAT1A and GNMT gene resequencing, functional genomics, and hepatic genotype-phenotype correlation. Drug Metab. Dispos. 2012, 40, 1984–1992. [Google Scholar] [CrossRef] [PubMed]
- Martinov, M.V.; Vitvitsky, V.M.; Banerjee, R.; Ataullakhanov, F.I. The logic of the hepatic methionine metabolic cycle. Biochim. Biophys. Acta 2010, 1804, 89–96. [Google Scholar] [CrossRef]
- Palmisano, B.T.; Zhu, L.; Stafford, J.M. Role of Estrogens in the Regulation of Liver Lipid Metabolism. Adv. Exp. Med. Biol. 2017, 1043, 227–256. [Google Scholar] [CrossRef]
- Shen, Z.; Hu, Y.; Zhou, C.; Yuan, J.; Xu, J.; Hao, W.; Deng, H.; Ye, D. ESRRG promoter hypermethylation as a diagnostic and prognostic biomarker in laryngeal squamous cell carcinoma. J. Clin. Lab. Anal. 2019, 33, e22899. [Google Scholar] [CrossRef]
- Klein Geltink, R.I.; Pearce, E.L. The importance of methionine metabolism. Elife 2019, 8, e47221. [Google Scholar] [CrossRef]
- Murray, B.; Barbier-Torres, L.; Fan, W.; Mato, J.M.; Lu, S.C. Methionine adenosyltransferases in liver cancer. World J. Gastroenterol. 2019, 25, 4300–4319. [Google Scholar] [CrossRef]
- LeGros, L.; Halim, A.B.; Chamberlin, M.E.; Geller, A.; Kotb, M. Regulation of the human MAT2B gene encoding the regulatory beta subunit of methionine adenosyltransferase, MAT II. J. Biol. Chem. 2001, 276, 24918–24924. [Google Scholar] [CrossRef]
- Bailey, J.; Douglas, H.; Masino, L.; de Carvalho, L.P.S.; Argyrou, A. Human Mat2A Uses an Ordered Kinetic Mechanism and Is Stabilized but Not Regulated by Mat2B. Biochemistry 2021, 60, 3621–3632. [Google Scholar] [CrossRef]
- Cai, J.; Sun, W.M.; Hwang, J.J.; Stain, S.C.; Lu, S.C. Changes in S-adenosylmethionine synthetase in human liver cancer: Molecular characterization and significance. Hepatology 1996, 24, 1090–1097. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.W.; Jiao, J.Y.; Chen, Z.; Zhu, X.Y.; Cheng, C.S. Keep a watchful eye on methionine adenosyltransferases, novel therapeutic opportunities for hepatobiliary and pancreatic tumours. Biochim. Biophys. Acta Rev. Cancer 2022, 1877, 188793. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.M.; Tsai, C.H.; Huang, C.C.; Hwang, H.H.; Li, J.R.; Liu, C.C.; Ko, H.A.; Chiang, E.I. Downregulation of Methionine Cycle Genes MAT1A and GNMT Enriches Protein-Associated Translation Process and Worsens Hepatocellular Carcinoma Prognosis. Int. J. Mol. Sci. 2022, 23, 481. [Google Scholar] [CrossRef] [PubMed]
- Perez, C.; Perez-Zuniga, F.J.; Garrido, F.; Reytor, E.; Portillo, F.; Pajares, M.A. The Oncogene PDRG1 Is an Interaction Target of Methionine Adenosyltransferases. PLoS ONE 2016, 11, e0161672. [Google Scholar] [CrossRef]
- Huang, Z.Z.; Mao, Z.; Cai, J.; Lu, S.C. Changes in methionine adenosyltransferase during liver regeneration in the rat. Am. J. Physiol. 1998, 275, G14–G21. [Google Scholar] [CrossRef]
- Chu, P.Y.; Wu, H.J.; Wang, S.M.; Chen, P.M.; Tang, F.Y.; Chiang, E.I. MAT2A Localization and Its Independently Prognostic Relevance in Breast Cancer Patients. Int. J. Mol. Sci. 2021, 22, 5382. [Google Scholar] [CrossRef]
- An, J.; Na, S.K.; Shim, J.H.; Park, Y.S.; Jun, M.J.; Lee, J.H.; Song, G.W.; Lee, H.C.; Yu, E. Histological expression of methionine adenosyl transferase (MAT) 2A as a post-surgical prognostic surrogate in patients with hepatocellular carcinoma. J. Surg. Oncol. 2018, 117, 892–901. [Google Scholar] [CrossRef]
- Alam, M.; Shima, H.; Matsuo, Y.; Long, N.C.; Matsumoto, M.; Ishii, Y.; Sato, N.; Sugiyama, T.; Nobuta, R.; Hashimoto, S.; et al. mTORC1-independent translation control in mammalian cells by methionine adenosyltransferase 2A and S-adenosylmethionine. J. Biol. Chem. 2022, 298, 102084. [Google Scholar] [CrossRef]
- Zhao, L.; Su, H.; Liu, X.; Wang, H.; Feng, Y.; Wang, Y.; Chen, H.; Dai, L.; Lai, S.; Xu, S.; et al. mTORC1-c-Myc pathway rewires methionine metabolism for HCC progression through suppressing SIRT4 mediated ADP ribosylation of MAT2A. Cell Biosci. 2022, 12, 183. [Google Scholar] [CrossRef]
- Guo, J.; Yang, Y.; Buettner, R.; Rosen, S.T. Targeting the methionine-methionine adenosyl transferase 2A- S -adenosyl methionine axis for cancer therapy. Curr. Opin. Oncol. 2022, 34, 546–551. [Google Scholar] [CrossRef]
- Wang, Y.C.; Chiang, E.P. Low-dose methotrexate inhibits methionine S-adenosyltransferase in vitro and in vivo. Mol. Med. 2012, 18, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, J.L.; Boukaba, A.; Sandoval, J.; Georgieva, E.I.; Latasa, M.U.; Garcia-Trevijano, E.R.; Serviddio, G.; Nakamura, T.; Avila, M.A.; Sastre, J.; et al. Transcription of the MAT2A gene, coding for methionine adenosyltransferase, is up-regulated by E2F and Sp1 at a chromatin level during proliferation of liver cells. Int. J. Biochem. Cell. Biol. 2007, 39, 842–850. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, M.H.; Pitot, H.C.; Chung, S.H.; Lambert, P.F.; Drinkwater, N.R.; Bilger, A. Estrogen Receptor-alpha Suppresses Liver Carcinogenesis and Establishes Sex-Specific Gene Expression. Cancers 2021, 13, 2355. [Google Scholar] [CrossRef] [PubMed]
- Iyer, J.K.; Kalra, M.; Kaul, A.; Payton, M.E.; Kaul, R. Estrogen receptor expression in chronic hepatitis C and hepatocellular carcinoma pathogenesis. World J. Gastroenterol. 2017, 23, 6802–6816. [Google Scholar] [CrossRef]
- Kang, M.H.; Choi, H.; Oshima, M.; Cheong, J.H.; Kim, S.; Lee, J.H.; Park, Y.S.; Choi, H.S.; Kweon, M.N.; Pack, C.G.; et al. Estrogen-related receptor gamma functions as a tumor suppressor in gastric cancer. Nat. Commun. 2018, 9, 1920. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Choi, Y.K.; Byun, J.K.; Kim, M.K.; Kang, Y.N.; Kim, S.H.; Lee, S.; Jang, B.K.; Park, K.G. Estrogen-related receptor gamma is upregulated in liver cancer and its inhibition suppresses liver cancer cell proliferation via induction of p21 and p27. Exp. Mol. Med. 2016, 48, e213. [Google Scholar] [CrossRef]
- Gao, Y.; Gan, K.; Liu, K.; Xu, B.; Chen, M. SP1 Expression and the Clinicopathological Features of Tumors: A Meta-Analysis and Bioinformatics Analysis. Pathol. Oncol. Res. 2021, 27, 581998. [Google Scholar] [CrossRef]
- Liao, Y.; Wang, C.; Yang, Z.; Liu, W.; Yuan, Y.; Li, K.; Zhang, Y.; Wang, Y.; Shi, Y.; Qiu, Y.; et al. Dysregulated Sp1/miR-130b-3p/HOXA5 axis contributes to tumor angiogenesis and progression of hepatocellular carcinoma. Theranostics 2020, 10, 5209–5224. [Google Scholar] [CrossRef]
- Herr, P.; Bostrom, J.; Rullman, E.; Rudd, S.G.; Vesterlund, M.; Lehtio, J.; Helleday, T.; Maddalo, G.; Altun, M. Cell Cycle Profiling Reveals Protein Oscillation, Phosphorylation, and Localization Dynamics. Mol. Cell. Proteom. 2020, 19, 608–623. [Google Scholar] [CrossRef]
- Li, T.; Fu, J.; Zeng, Z.; Cohen, D.; Li, J.; Chen, Q.; Li, B.; Liu, X.S. TIMER2.0 for analysis of tumor-infiltrating immune cells. Nucleic Acids Res. 2020, 48, W509–W514. [Google Scholar] [CrossRef]
- Wu, M.T.; Ye, W.T.; Wang, Y.C.; Chen, P.M.; Liu, J.Y.; Tai, C.K.; Tang, F.Y.; Li, J.R.; Liu, C.C.; Chiang, E.I. MTHFR Knockdown Assists Cell Defense against Folate Depletion Induced Chromosome Segregation and Uracil Misincorporation in DNA. Int. J. Mol. Sci. 2021, 22, 9392. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MAT2A | ||||
|---|---|---|---|---|
| Characteristics | No. | C/N ≦ 1 (N = 132) | C/N > 1 (N = 129) | p-Value |
| Diagnostic age | ||||
| <65 | 134 | 66 | 68 | 0.565 |
| ≧65 | 123 | 65 | 58 | |
| Missing | 4 | |||
| Gender | ||||
| Female | 63 | 25 | 38 | 0.047 |
| Male | 198 | 107 | 91 | |
| Stage | ||||
| I, II | 214 | 112 | 102 | 0.173 |
| III, IV | 47 | 20 | 27 | |
| 10-year follow-up | ||||
| Live | 119 | 61 | 58 | 0.928 |
| Dead | 137 | 69 | 68 | |
| Loss of follow-up | 15 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chu, P.-Y.; Chou, D.-A.; Chen, P.-M.; Chiang, E.-P.I. Translocation of Methionine Adenosyl Transferase MAT2A and Its Prognostic Relevance for Liver Hepatocellular Carcinoma. Int. J. Mol. Sci. 2023, 24, 9103. https://doi.org/10.3390/ijms24109103
Chu P-Y, Chou D-A, Chen P-M, Chiang E-PI. Translocation of Methionine Adenosyl Transferase MAT2A and Its Prognostic Relevance for Liver Hepatocellular Carcinoma. International Journal of Molecular Sciences. 2023; 24(10):9103. https://doi.org/10.3390/ijms24109103
Chicago/Turabian StyleChu, Pei-Yi, Dev-Aur Chou, Po-Ming Chen, and En-Pei Isabel Chiang. 2023. "Translocation of Methionine Adenosyl Transferase MAT2A and Its Prognostic Relevance for Liver Hepatocellular Carcinoma" International Journal of Molecular Sciences 24, no. 10: 9103. https://doi.org/10.3390/ijms24109103