

Ring Finger 149-Related Is an FGF/MAPK-Independent Regulator of Pharyngeal Muscle Fate Specification


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
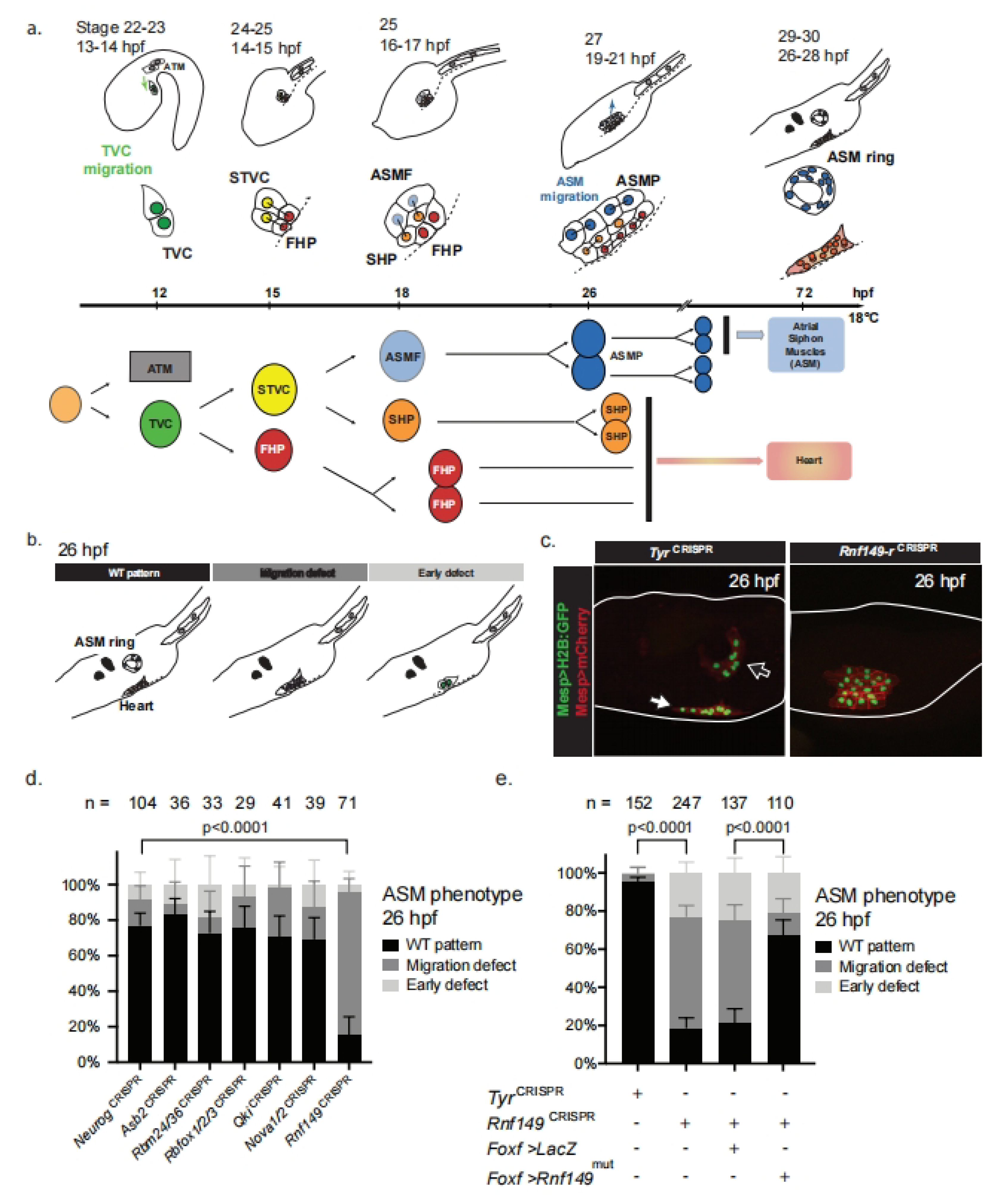
2.1. CRISPR/Cas9-Mediated Mutagenesis Identifies the Pharyngeal Muscle Determinant Rnf149-r
2.2. Rnf149-r Encodes an Atypical Ubiquitin Ligase-Related Protein
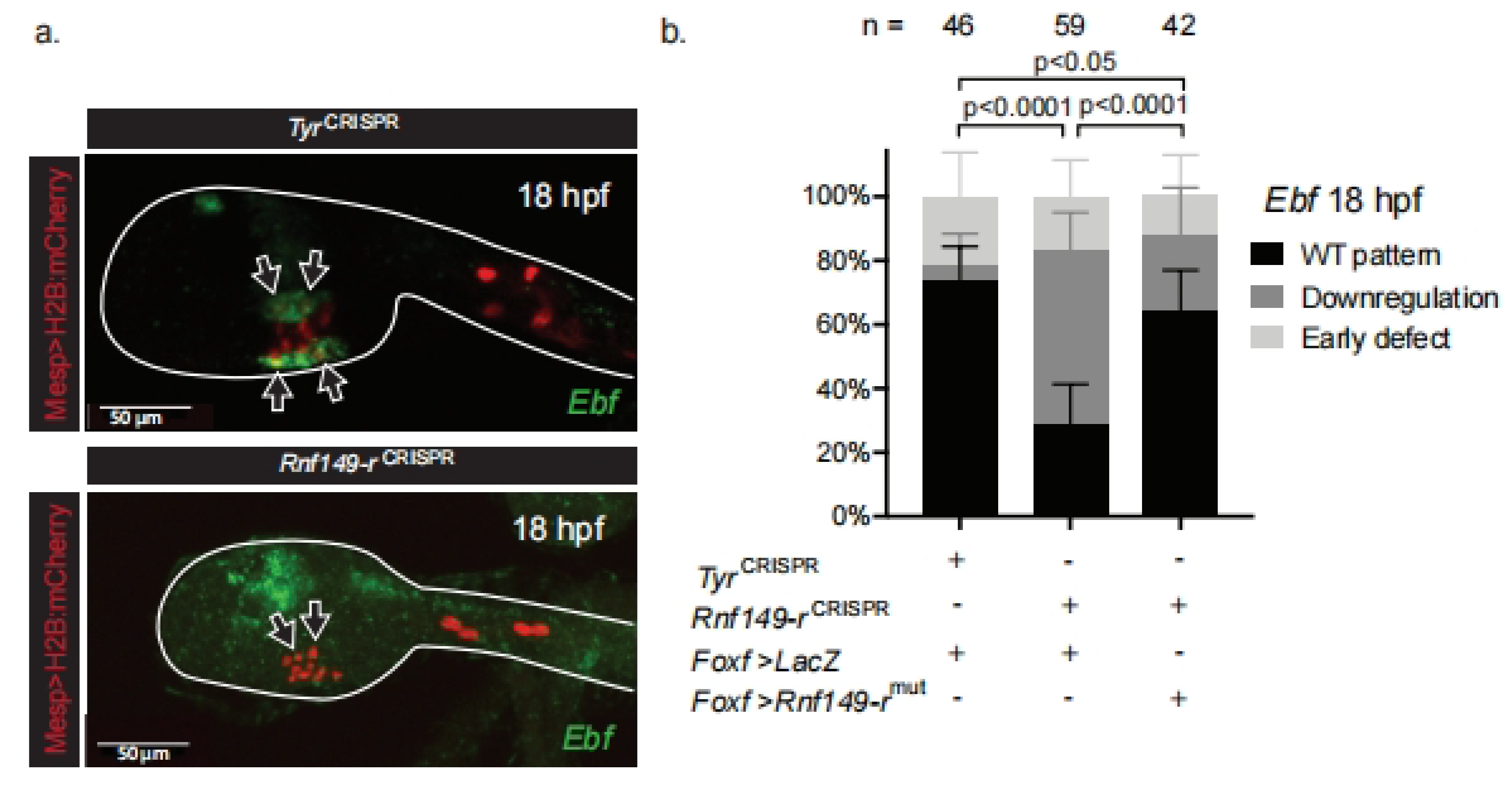
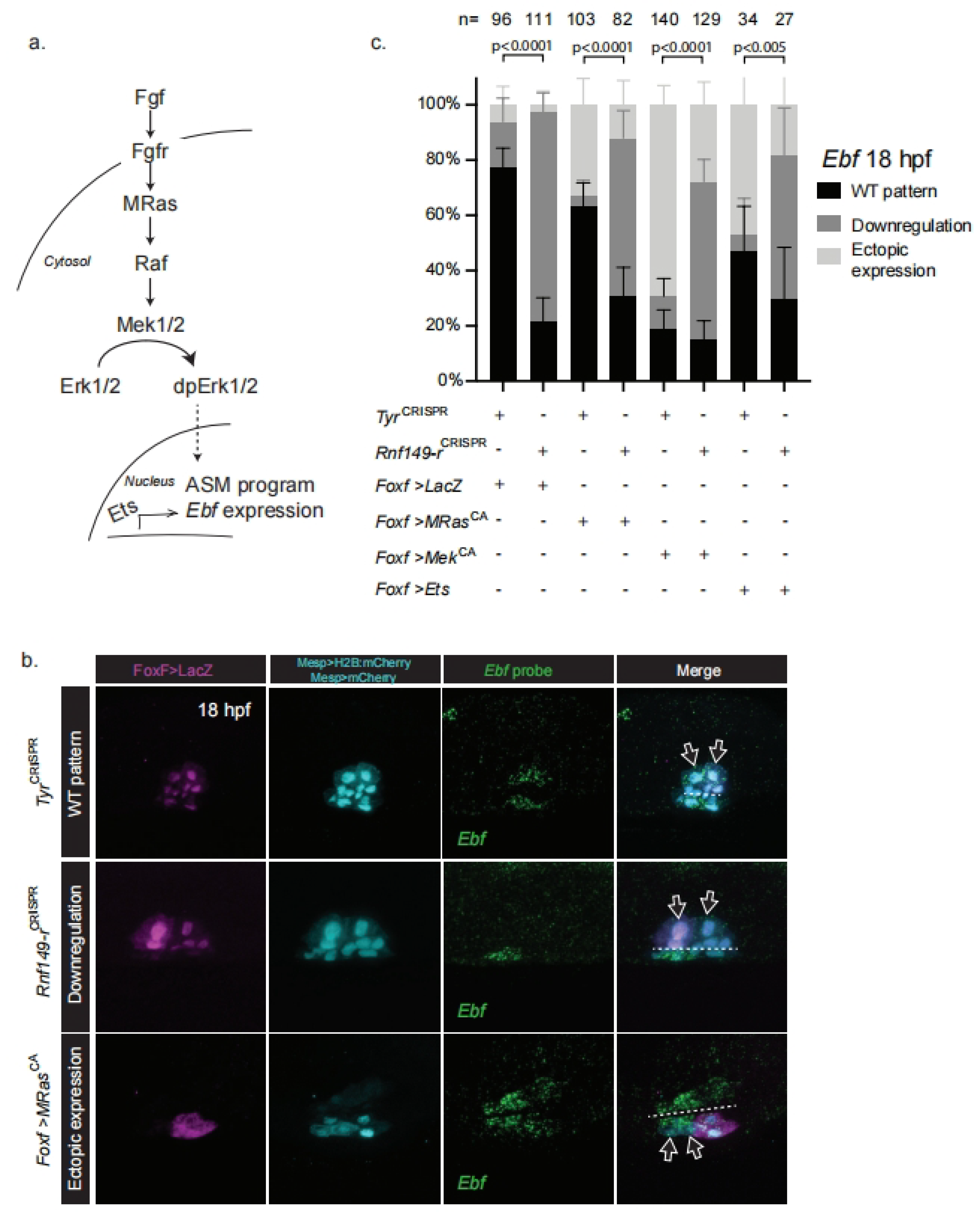
2.3. Rnf149-r Regulates Cardiopharyngeal Fates Independently of FGF/MAPK Signaling
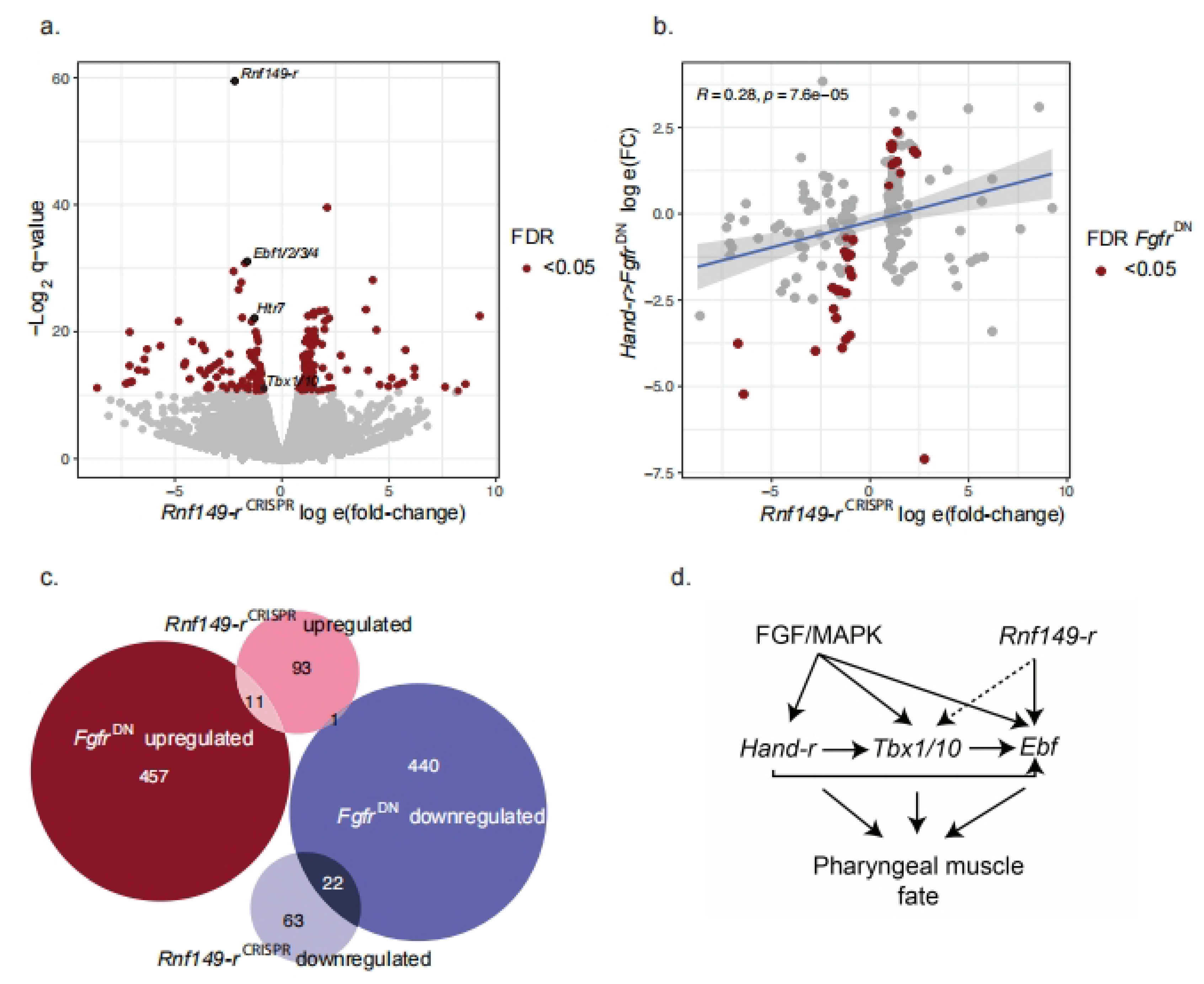
2.4. Rnf149-r Regulates Both MAPK-Dependent and Independent Genes
3. Discussion
4. Methods
4.1. Ciona Robusta Handling
4.2. CRISPR/Cas9-Mediated Mutagenesis
4.3. Molecular Cloning of Rnf149-rmut Rescue Construct
4.4. Fluorescent In Situ Hybridization Immunohistochemistry (FISH-IHC) of Ciona Embryos
4.5. Cell Dissociation and FACS-Purification of Ciona Robusta Cells
4.6. RNA-seq Library Preparation, Sequencing and Analysis
4.7. Data Availability
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kelly, R.G.; Buckingham, M.E.; Moorman, A.F. Heart Fields and Cardiac Morphogenesis. Cold Spring Harb. Perspect. Med. 2014, 4, a015750. [Google Scholar] [CrossRef] [PubMed]
- Kelly, R.G. Advances in the second heart field. In Molecular Mechanism of Congenital Heart Disease and Pulmonary Hypertension; Springer: Singapore, 2020; pp. 301–307. [Google Scholar] [CrossRef]
- Buckingham, M. First and Second Heart Field. In Heart Development and Regeneration; Academic Press: Cambridge, MA, USA, 2016; pp. 25–40. [Google Scholar] [CrossRef]
- Lescroart, F.; Chabab, S.; Lin, X.; Rulands, S.; Paulissen, C.; Rodolosse, A.; Auer, H.; Achouri, Y.; Dubois, C.; Bondue, A.; et al. Early lineage restriction in temporally distinct populations of Mesp1 progenitors during mammalian heart development. Nature 2014, 16, 829–840. [Google Scholar] [CrossRef] [PubMed]
- Devine, W.P.; Wythe, J.D.; George, M.; Koshiba-Takeuchi, K.; Bruneau, B.G. Early patterning and specification of cardiac progenitors in gastrulating mesoderm. eLife 2014, 3, e03848. [Google Scholar] [CrossRef] [PubMed]
- Lescroart, F.; Hamou, W.; Francou, A.; Théveniau-Ruissy, M.; Kelly, R.G.; Buckingham, M. Clonal analysis reveals a common origin between nonsomite-derived neck muscles and heart myocardium. Proc. Natl. Acad. Sci. USA 2015, 112, 1446–1451. [Google Scholar] [CrossRef] [PubMed]
- Lescroart, F.; Kelly, R.; le Garrec, J.-F.; Nicolas, J.-F.; Meilhac, S.; Buckingham, M. Clonal analysis reveals common lineage relationships between head muscles and second heart field derivatives in the mouse embryo. Development 2010, 137, 3269–3279. [Google Scholar] [CrossRef] [PubMed]
- Delsuc, F.; Brinkmann, H.; Chourrout, D.; Philippe, H. Tunicates and not cephalochordates are the closest living relatives of vertebrates. Nature 2006, 439, 965–968. [Google Scholar] [CrossRef]
- Putnam, N.H.; Butts, T.; Ferrier, D.E.K.; Furlong, R.F.; Hellsten, U.; Kawashima, T.; Robinson-Rechavi, M.; Shoguchi, E.; Terry, A.; Yu, J.-K.; et al. The amphioxus genome and the evolution of the chordate karyotype. Nature 2008, 453, 1064–1071. [Google Scholar] [CrossRef]
- Kaplan, N.; Razy-Krajka, F.; Christiaen, L. Regulation and evolution of cardiopharyngeal cell identity and behavior: Insights from simple chordates. Curr. Opin. Genet. Dev. 2015, 32, 119–128. [Google Scholar] [CrossRef]
- Anderson, H.E.; Christiaen, L. Ciona as a Simple Chordate Model for Heart Development and Regeneration. J. Cardiovasc. Dev. Dis. 2016, 3, 25. [Google Scholar] [CrossRef]
- Christiaen, L.; Davidson, B.; Kawashima, T.; Powell, W.; Nolla, H.; Vranizan, K.; Levine, M. The transcription/migration interface in heart precursors of Ciona intestinalis. Science 2008, 320, 1349–1352. [Google Scholar] [CrossRef]
- Davidson, B.; Shi, W.; Levine, M. Uncoupling heart cell specification and migration in the simple chordate Ciona intestinalis. Development 2005, 132, 4811–4818. [Google Scholar] [CrossRef] [PubMed]
- Stolfi, A.; Gainous, T.B.; Young, J.J.; Mori, A.; Levine, M.; Christiaen, L. Early Chordate Origins of the Vertebrate Second Heart Field. Science 2010, 329, 565–568. [Google Scholar] [CrossRef] [PubMed]
- Satou, Y.; Imai, K.S.; Satoh, N. The ascidian Mesp gene specifies heart precursor cells. Development 2004, 131, 2533–2541. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Razy-Krajka, F.; Siu, E.; Ketcham, A.; Christiaen, L. NK4 antagonizes Tbx1/10 to promote cardiac versus pharyngeal muscle fate in the ascidian second heart field. PLoS Biol. 2013, 11, e1001725. [Google Scholar] [CrossRef]
- Razy-Krajka, F.; Lam, K.; Wang, W.; Stolfi, A.; Joly, M.; Bonneau, R.; Christiaen, L. Collier/OLF/EBF-Dependent Transcriptional Dynamics Control Pharyngeal Muscle Specification from Primed Cardiopharyngeal Progenitors. Dev. Cell 2014, 29, 263–276. [Google Scholar] [CrossRef]
- Davidson, B.; Shi, W.; Beh, J.; Christiaen, L.; Levine, M. FGF signaling delineates the cardiac progenitor field in the simple chordate, Ciona intestinalis. Genes Dev. 2006, 20, 2728–2738. [Google Scholar] [CrossRef]
- Razy-Krajka, F.; Gravez, B.; Kaplan, N.; Racioppi, C.; Wang, W.; Christiaen, L. An FGF-driven feed-forward circuit patterns the cardiopharyngeal mesoderm in space and time. eLife 2018, 7, e29656. [Google Scholar] [CrossRef]
- Hotta, K.; Dauga, D.; Manni, L. The ontology of the anatomy and development of the solitary ascidian Ciona: The swimming larva and its metamorphosis. Sci. Rep. 2020, 10, 17916. [Google Scholar] [CrossRef]
- Hotta, K.; Mitsuhara, K.; Takahashi, H.; Inaba, K.; Oka, K.; Gojobori, T.; Ikeo, K. A web-based interactive devel-opmental table for the ascidian Ciona intestinalis, including 3D real-image embryo reconstruc-tions: I. From fertilized egg to hatching larva. Dev. Dyn. 2007, 236, 1790–1805. [Google Scholar] [CrossRef]
- Wang, W.; Niu, X.; Stuart, T.; Jullian, E.; Mauck, W.M., III; Kelly, R.G.; Satija, R.; Christiaen, L. A single-cell transcriptional roadmap for cardiopharyngeal fate diversification. Nat. Cell Biol. 2019, 21, 674–686. [Google Scholar] [CrossRef]
- Yamada, L.; Kobayashi, K.; Satou, Y.; Satoh, N. Microarray analysis of localization of maternal transcripts in eggs and early embryos of the ascidian, Ciona intestinalis. Dev. Biol. 2005, 284, 536–550. [Google Scholar] [CrossRef]
- Brozovic, M.; Martin, C.; Dantec, C.; Dauga, D.; Mendez, M.; Simion, P.; Percher, M.; Laporte, B.; Scornavacca, C.; Di Gregorio, A.; et al. ANISEED 2015: A digital framework for the comparative developmental biology of ascidians. Nucleic Acids Res. 2016, 44, D808–D818. [Google Scholar] [CrossRef]
- Blum, M.; Chang, H.-Y.; Chuguransky, S.; Grego, T.; Kandasaamy, S.; Mitchell, A.; Nuka, G.; Paysan-Lafosse, T.; Qureshi, M.; Raj, S.; et al. The InterPro protein families and domains database: 20 years on. Nucleic Acids Res. 2021, 49, D344–D354. [Google Scholar] [CrossRef] [PubMed]
- UniProt Consortium. UniProt: A worldwide hub of protein knowledge. Nucleic Acids Res. 2019, 47, D506–D515. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, S.; Razy-Krajka, F.; Christiaen, L.; Stolfi, A. CRISPR Knockouts in Ciona Embryos. Transgenic Ascidians 2018, 1029, 141–152. [Google Scholar] [CrossRef]
- Stolfi, A.; Gandhi, S.; Salek, F.; Christiaen, L. Tissue-specific genome editing in Ciona embryos by CRISPR/Cas9. Development 2014, 141, 4115–4120. [Google Scholar] [CrossRef] [PubMed]
- Haeussler, M.; Schönig, K.; Eckert, H.; Eschstruth, A.; Mianné, J.; Renaud, J.-B.; Schneider-Maunoury, S.; Shkumatava, A.; Teboul, L.; Kent, J.; et al. Evaluation of off-target and on-target scoring algorithms and integration into the guide RNA selection tool CRISPOR. Genome Biol. 2016, 17, 148. [Google Scholar] [CrossRef]
- Gandhi, S.; Haeussler, M.; Razy-Krajka, F.; Christiaen, L.; Stolfi, A. Evaluation and rational design of guide RNAs for efficient CRISPR/Cas9-mediated mutagenesis in Ciona. Dev. Biol. 2017, 425, 8–20. [Google Scholar] [CrossRef] [PubMed]
- Beh, J.; Shi, W.; Levine, M.; Davidson, B.; Christiaen, L. FoxF is essential for FGF-induced migration of heart pro-genitor cells in the ascidian Ciona intestinalis. Development 2007, 134, 3297–3305. [Google Scholar] [CrossRef]
- Racioppi, C.; Wiechecki, A.K.; Christiaen, L. Combinatorial chromatin dynamics foster accurate cardiopharyngeal fate choices. eLife 2019, 8, e49921. [Google Scholar] [CrossRef]
- Mahon, P.; Bateman, A. The PA domain: A protease-associated domain. Protein Sci. 2000, 9, 1930–1934. [Google Scholar] [CrossRef]
- Hong, S.-W.; Jin, D.-H.; Shin, J.-S.; Moon, J.-H.; Na, Y.-S.; Jung, K.-A.; Kim, S.-M.; Kim, J.C.; Kim, K.-P.; Hong, Y.S.; et al. Ring Finger Protein 149 Is an E3 Ubiquitin Ligase Active on Wild-type v-Raf Murine Sarcoma Viral Oncogene Homolog B1 (BRAF). J. Biol. Chem. 2012, 287, 24017–24025. [Google Scholar] [CrossRef]
- Keduka, E.; Kaiho, A.; Hamada, M.; Watanabe-Takano, H.; Takano, K.; Ogasawara, M.; Satou, Y.; Satoh, N.; Endo, T. M-Ras evolved independently of R-Ras and its neural function is conserved between mammalian and ascidian, which lacks classical Ras. Gene 2009, 429, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Miya, T.; Nishida, H. An Ets transcription factor, HrEts, is target of FGF signaling and involved in induction of notochord, mesenchyme, and brain in ascidian embryos. Dev. Biol. 2003, 261, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Christiaen, L.; Wagner, E.; Shi, W.; Levine, M. Isolation of sea squirt (Ciona) gametes, fertilization, dechorionation, and development. Cold Spring Harb. Protoc. 2009, 2009, pdb-prot5344. [Google Scholar] [CrossRef] [PubMed]
- Racioppi, C.; Kamal, A.K.; Razy-Krajka, F.; Gambardella, G.; Zanetti, L.; di Bernardo, D.; Sanges, R.; Christiaen, L.A.; Ristoratore, F. Fibroblast growth factor signalling controls nervous system patterning and pigment cell formation in Ciona intestinalis. Nat. Commun. 2014, 5, 4830. [Google Scholar] [CrossRef]
- Christiaen, L.; Wagner, E.; Shi, W.; Levine, M. Isolation of Individual Cells and Tissues from Electroporated Sea Squirt (Ciona) Embryos by Fluorescence-Activated Cell Sorting (FACS). Cold Spring Harb. Protoc. 2009, 2009, prot5349. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Racioppi, C.; Gravez, B.; Christiaen, L. Purification of Fluorescent Labeled Cells from Dissociated Ciona Embryos. Adv. Exp. Med. Biol. 2018, 1029, 101–107. [Google Scholar] [CrossRef]
- Zhang, P.; Hung, L.-H.; Lloyd, W.; Yeung, K.Y. Hot-starting software containers for STAR aligner. Gigascience 2018, 7, giy092. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. The R package Rsubread is easier, faster, cheaper and better for alignment and quantification of RNA sequencing reads. Nucleic Acids Res. 2019, 47, e47. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vitrinel, B.; Vogel, C.; Christiaen, L. Ring Finger 149-Related Is an FGF/MAPK-Independent Regulator of Pharyngeal Muscle Fate Specification. Int. J. Mol. Sci. 2023, 24, 8865. https://doi.org/10.3390/ijms24108865
Vitrinel B, Vogel C, Christiaen L. Ring Finger 149-Related Is an FGF/MAPK-Independent Regulator of Pharyngeal Muscle Fate Specification. International Journal of Molecular Sciences. 2023; 24(10):8865. https://doi.org/10.3390/ijms24108865
Chicago/Turabian StyleVitrinel, Burcu, Christine Vogel, and Lionel Christiaen. 2023. "Ring Finger 149-Related Is an FGF/MAPK-Independent Regulator of Pharyngeal Muscle Fate Specification" International Journal of Molecular Sciences 24, no. 10: 8865. https://doi.org/10.3390/ijms24108865