
Identification of Key Genes and Pathways Associated with PIEZO1 in Bone-Related Disease Based on Bioinformatics


Abstract
:1. Introduction
2. Materials and Methods
2.1. Gene Expression Profiles in Piezo1 Mutant Tissues
2.2. Data Pre-Processing
2.3. Identifying DEGs
2.4. Identification of Overlapping DEGs in Mice Piezo1 Mutant Tissues
2.5. Gene Ontology (GO) Enrichment and Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway Analysis
2.6. Construction of the Protein–Protein Interaction Network
2.7. Module Analysis
2.8. In Vitro Chondrocyte Verification
Quantitative Real-Time Polymerase Chain Reaction
3. Results
3.1. Identification of DEGs in Piezo1 Mutant Tissues
3.2. Identification of Overlapping DEGs in Piezo1 Mutant Tissues
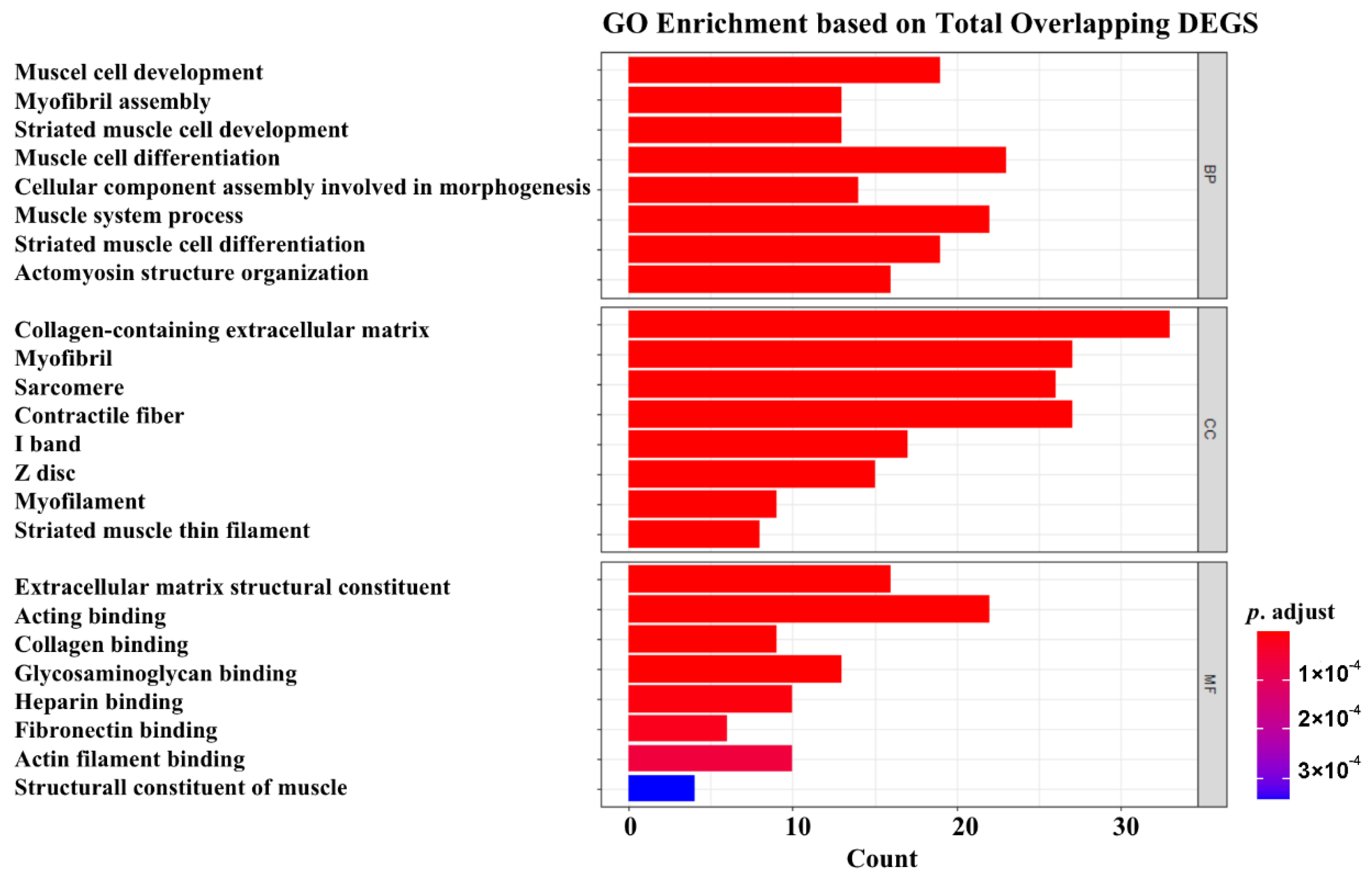
3.3. GO Enrichment Analysis
3.4. KEGG Pathway Analysis
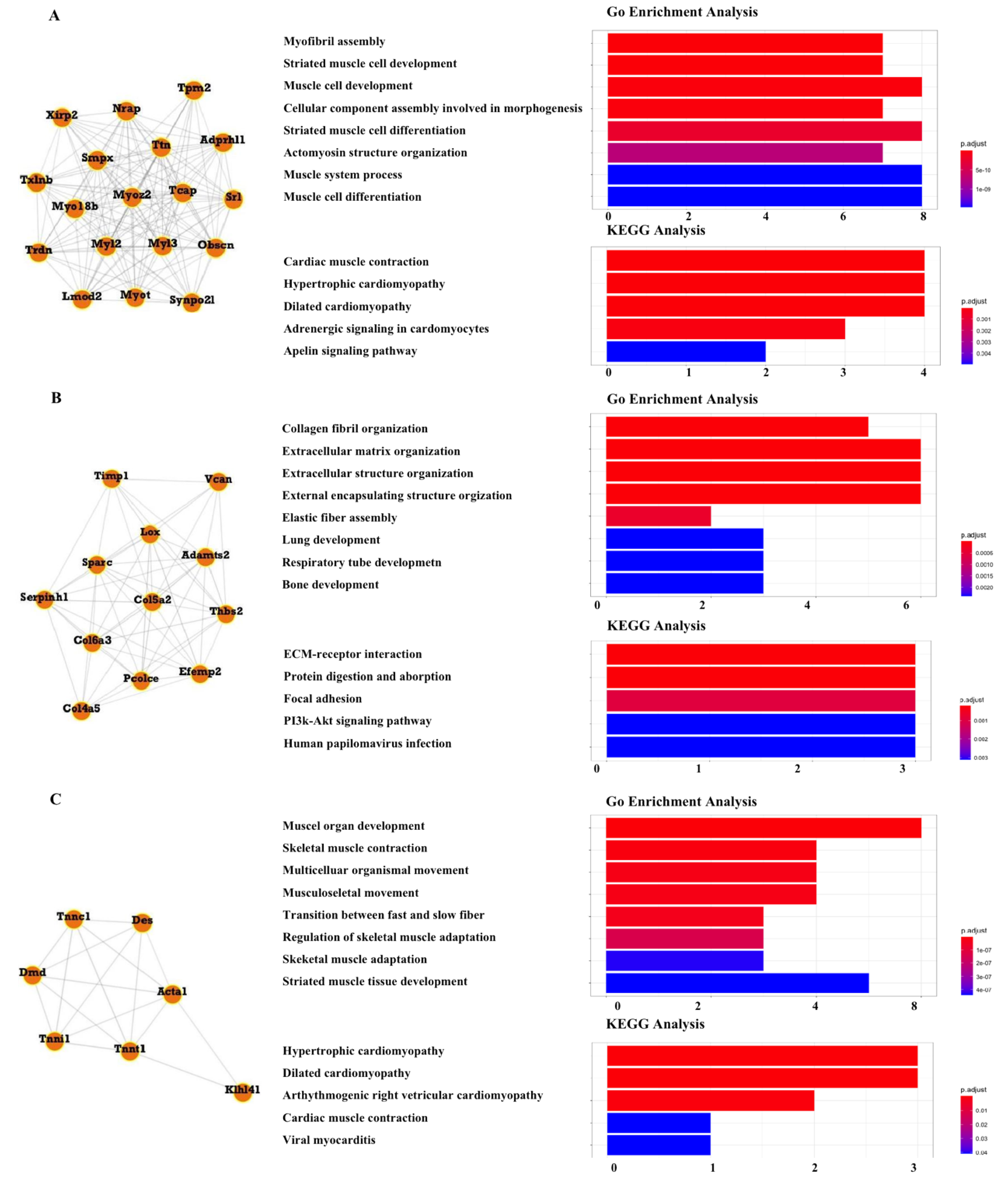
3.5. PPI Network Analysis
3.6. Module Analysis
3.7. In Vitro Chondrocyte Verification
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chalfie, M. Neurosensory mechanotransduction. Nat. Rev. Mol. Cell Biol. 2009, 10, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Hamill, O.P.; Martinac, B. Molecular Basis of Mechanotransduction in Living Cells. Physiol. Rev. 2001, 81, 685–740. [Google Scholar] [CrossRef] [PubMed]
- Price, C.; Zhou, X.; Li, W.; Wang, L. Real-time measurement of solute transport within the lacunar-canalicular system of mechanically loaded bone: Direct evidence for load-induced fluid flow. J. Bone Miner. Res. 2011, 26, 277–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coste, B.; Mathur, J.; Schmidt, M.; Earley, T.J.; Ranade, S.; Petrus, M.J.; Dubin, A.E.; Patapoutian, A. Piezo1 and Piezo2 Are Essential Components of Distinct Mechanically Activated Cation Channels. Science 2010, 330, 55–60. [Google Scholar] [CrossRef] [Green Version]
- Bagriantsev, S.N.; Gracheva, E.O.; Gallagher, P.G. Piezo Proteins: Regulators of Mechanosensation and Other Cellular Processes. J. Biol. Chem. 2014, 289, 31673–31681. [Google Scholar] [CrossRef] [Green Version]
- Coste, B.; Xiao, B.; Santos, J.S.; Syeda, R.; Grandl, J.; Spencer, K.S.; Kim, S.E.; Schmidt, M.; Mathur, J.; Dubin, A.E.; et al. Piezo proteins are pore-forming subunits of mechanically activated channels. Nature 2012, 483, 176–181. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Lewis, A.H.; Grandl, J. Touch, Tension, and Transduction—The Function and Regulation of Piezo Ion Channels. Trends Biochem. Sci. 2016, 42, 57–71. [Google Scholar] [CrossRef] [Green Version]
- Frohlich, M.W.; Jaeger, A.; Weiss, D.G.; Kriehuber, R. Inhibition of BCL-2 leads to increased apoptosis and delayed neuronal differentiation in human ReNcell VM cells in vitro. Int. J. Dev. Neurosci. 2015, 48, 9–17. [Google Scholar] [CrossRef]
- Lee, W.; Leddy, H.A.; Chen, Y.; Lee, S.H.; Zelenski, N.A.; McNulty, A.L.; Wu, J.; Beicker, K.N.; Coles, J.; Zauscher, S.; et al. Synergy between Piezo1 and Piezo2 channels confers high-strain mechanosensitivity to articular cartilage. Proc. Natl. Acad. Sci. USA 2014, 111, E5114–E5122. [Google Scholar] [CrossRef] [Green Version]
- Sugimoto, A.; Miyazaki, A.; Kawarabayashi, K.; Shono, M.; Akazawa, Y.; Hasegawa, T.; Ueda-Yamaguchi, K.; Kitamura, T.; Yoshizaki, K.; Fukumoto, S.; et al. Piezo type mechanosensitive ion channel component 1 functions as a regulator of the cell fate determination of mesenchymal stem cells. Sci. Rep. 2017, 7, 17696. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.-L.; Liao, H.-Y. Piezo-type mechanosensitive ion channel component 1 (Piezo1) in human cancer. Biomed. Pharmacother. 2021, 140, 111692. [Google Scholar] [CrossRef]
- Sanchez-Adams, J.; Leddy, H.A.; McNulty, A.L.; O’Conor, C.J.; Guilak, F. The Mechanobiology of Articular Cartilage: Bearing the Burden of Osteoarthritis. Curr. Rheumatol. Rep. 2014, 16, 451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, W.; Nims, R.J.; Savadipour, A.; Zhang, Q.; Leddy, H.A.; Liu, F.; McNulty, A.L.; Chen, Y.; Guilak, F.; Liedtke, W.B. Inflammatory signaling sensitizes Piezo1 mechanotransduction in articular chondrocytes as a pathogenic feed-forward mechanism in osteoarthritis. Proc. Natl. Acad. Sci. USA 2021, 118, e2001611118. [Google Scholar] [CrossRef]
- Latourte, A.; Kloppenburg, M.; Richette, P. Emerging pharmaceutical therapies for osteoarthritis. Nat. Rev. Rheumatol. 2020, 16, 673–688. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; You, X.; Lotinun, S.; Zhang, L.; Wu, N.; Zou, W. Mechanical sensing protein PIEZO1 regulates bone homeostasis via osteoblast-osteoclast crosstalk. Nat. Commun. 2020, 11, 282. [Google Scholar] [CrossRef] [Green Version]
- De Almeida, R.C.; Ramos, Y.F.M.; Mahfouz, A.; Hollander, W.D.; Lakenberg, N.; Houtman, E.; Van Hoolwerff, M.; Suchiman, H.E.D.; Ruiz, A.R.; Slagboom, P.E.; et al. RNA sequencing data integration reveals an miRNA interactome of osteoarthritis cartilage. Ann. Rheum. Dis. 2018, 78, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Tuerlings, M.; van Hoolwerff, M.; Houtman, E.; Suchiman, E.H.E.D.; Lakenberg, N.; Mei, H.; van der Linden, E.; Nelissen, R.R.G.H.H.; Ramos, Y.Y.F.M.; de Almeida, R.C.; et al. RNA Sequencing Reveals Interacting Key Determinants of Osteoarthritis Acting in Subchondral Bone and Articular Cartilage: Identification of IL11 and CHADL as Attractive Treatment Targets. Arthritis Rheumatol. 2020, 73, 789–799. [Google Scholar] [CrossRef]
- Zheng, H.; Forgetta, V.; Hsu, Y.; Estrada, K.; Rosello-Diez, A.; Leo, P.J.; Dahia, C.L.; Park-Min, K.H.; Tobias, J.H.; Kooperberg, C.; et al. Whole-genome sequencing identifies EN1 as a determinant of bone density and fracture. Nat. 2015, 526, 112–117. [Google Scholar] [CrossRef] [Green Version]
- Kun-Peng, Z.; Xiao-Long‡, M.; Lei‡, Z.; Chun-Lin, Z.; Jian-Ping, H.; Tai-Cheng, Z. Screening circular RNA related to chemotherapeutic resistance in osteosarcoma by RNA sequencing. Epigenomics 2018, 10, 1327–1346. [Google Scholar] [CrossRef]
- Hasegawa, K.; Fujii, S.; Matsumoto, S.; Tajiri, Y.; Kikuchi, A.; Kiyoshima, T. YAP signaling induces PIEZO1 to promote oral squamous cell carcinoma cell proliferation. J. Pathol. 2020, 253, 80–93. [Google Scholar] [CrossRef]
- Li, J.; Hou, B.; Tumova, S.; Muraki, K.; Bruns, A.; Ludlow, M.J.; Sedo, A.; Hyman, A.J.; McKeown, L.; Young, R.S.; et al. Piezo1 integration of vascular architecture with physiological force. Nature 2014, 515, 279–282. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Gao, B.; Fan, Y.; Liu, Y.; Feng, S.; Cong, Q.; Zhang, X.; Zhou, Y.; Yadav, P.S.; Lin, J.; et al. Piezo1/2 mediate mechanotransduction essential for bone formation through concerted activation of NFAT-YAP1-ß-catenin. eLife 2020, 9, e52779. [Google Scholar] [CrossRef] [PubMed]
- Solis, A.G.; Bielecki, P.; Steach, H.R.; Sharma, L.; Harman, C.C.D.; Yun, S.; De Zoete, M.R.; Warnock, J.N.; To, S.D.F.; York, A.G.; et al. Mechanosensation of cyclical force by PIEZO1 is essential for innate immunity. Nature 2019, 573, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Park, C.; Bennett, C.; Thornton, M.; Kim, D. Rapid and accurate alignment of nucleotide conversion sequencing reads with HISAT-3N. Genome Res. 2021, 31, 1290–1295. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING database in 2021: Customizable protein–protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2020, 49, D605–D612. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Gosset, M.; Berenbaum, F.; Thirion, S.; Jacques, C. Primary culture and phenotyping of murine chondrocytes. Nat. Protoc. 2008, 3, 1253–1260. [Google Scholar] [CrossRef]
- Syeda, R.; Xu, J.; Dubin, A.; Coste, B.; Mathur, J.; Huynh, T.; Matzen, J.T.; Lao, J.; Tully, D.C.; Engels, I.H.; et al. Chemical activation of the mechanotransduction channel Piezo1. eLife 2015, 4, e07369. [Google Scholar] [CrossRef]
- Mariani, T.J.; Budhraja, V.; Mecham, B.H.; Gu, C.C.; Watson, M.A.; Sadovsky, Y. A variable fold-change threshold determines significance for expression microarrays. FASEB J. 2002, 17, 321–323. [Google Scholar] [CrossRef] [Green Version]
- Vimal, V.; Angelo, Z.; Riccardo, P.; Suvro, C.; Nadia, R.; Anna, T.; Mattia, C. Interleukin-1β, lipocalin 2 and nitric oxide synthase 2 are mechano-responsive mediators of mouse and human endothelial cell-osteoblast crosstalk. Sci. Rep. 2016, 6, 29880. [Google Scholar] [CrossRef] [Green Version]
- Nadia, R.; Mattia, C.; Sara Gemini, P.; Alfredo, C.; Patrick, L.; Petra, F.-M.; Martina, H.; Anna, T. Lipocalin 2: A new mechanoresponding gene regulating bone homeostasis. J. Bone Miner. Res. 2014, 30, 357–368. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.-J.; Yoon, H.-J.; Yoon, K.-A.; Gwon, M.-R.; Seong, S.J.; Suk, K.; Kim, S.-Y.; Yoon, Y.-R. Lipocalin-2 inhibits osteoclast formation by suppressing the proliferation and differentiation of osteoclast lineage cells. Exp. Cell Res. 2015, 334, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Barbara, J.-S.; Michael, E.B.; Cheng-Chi, C.; Erin, E.M.; Wendy, M.B.; Iannis, E.A.; Paul, G.H.; Svetlana, A.; Edward, P.B.; Terrill, K.M.; et al. Myeloid DAP12-associating lectin (MDL)-1 regulates synovial inflammation and bone erosion associated with autoimmune arthritis. J. Exp. Med. 2010, 207, 579–589. [Google Scholar] [CrossRef]
- Ooiean, T.; Szu-Ting, C.; Tsui-Ling, H.; Sin Fun, S.; Suzanne, C.; Sophie, A.V.; Tzu-Yun, H.; Jian Teddy, Z.; Wenwei, T.; Roberto, B.; et al. CLEC5A-Mediated Enhancement of the Inflammatory Response in Myeloid Cells Contributes to Influenza Virus Pathogenicity In Vivo. J. Virol. 2016, 91, e01813-16. [Google Scholar] [CrossRef] [Green Version]
- Fujita, K.-I.; Janz, S. Attenuation of WNT signaling by DKK-1 and -2 regulates BMP2-induced osteoblast differentiation and expression of OPG, RANKL and M-CSF. Mol. Cancer 2007, 6, 71. [Google Scholar] [CrossRef] [Green Version]
- Hassan, M.Q.; Maeda, Y.; Taipaleenmaki, H.; Zhang, W.; Jafferji, M.; Gordon, J.A.R.; Li, Z.; Croce, C.M.; van Wijnen, A.J.; Stein, J.L.; et al. miR-218 Directs a Wnt Signaling Circuit to Promote Differentiation of Osteoblasts and Osteomimicry of Metastatic Cancer Cells. J. Biol. Chem. 2012, 287, 42084–42092. [Google Scholar] [CrossRef] [Green Version]
- Nie, B.; Zhang, S.-Y.; Guan, S.-M.; Zhou, S.-Q.; Fang, X. Role of Wnt/β-Catenin Pathway in the Arterial Medial Calcification and Its Effect on the OPG/RANKL System. Curr. Med. Sci. 2019, 39, 28–36. [Google Scholar] [CrossRef]
- Fernando, A.; Masakiyo, S.; Mikiro, T.; Yasutomo, N.; Kyouhei, K.; Shin, E.; Masahiro, M.; Masayoshi, N.; Hiromi, K.; Nam-ho, H. Adenovirus-mediated overexpression of REIC/Dkk-3 selectively induces apoptosis in human prostate cancer cells through activation of c-Jun-NH2-kinase. Cancer Res. 2005, 65, 9617–9622. [Google Scholar] [CrossRef] [Green Version]
- Jun, Y.; Qian, T.; Yuen, Y.C.; Kwan, Y.L.; Simon, S.M.N.; Kin, F.C.; Linwei, T.; Sun, Y.R.; Ulf, N.; Christoph, R.; et al. Promoter methylation of the Wnt/beta-catenin signaling antagonist Dkk-3 is associated with poor survival in gastric cancer. Cancer 2008, 115, 49–60. [Google Scholar] [CrossRef]
- Tingxiu, X.; Lili, L.; Xuedong, Y.; Lan, Z.; Weiyan, P.; Zhu, Q.; Guosheng, R.; Qian, T. Epigenetic silencing of the WNT antagonist Dickkopf 3 disrupts normal Wnt/β-catenin signalling and apoptosis regulation in breast cancer cells. J. Cell. Mol. Med. 2013, 17, 1236–1246. [Google Scholar] [CrossRef]
- Yu, M.; Douglas, A.; Yusuke, H.; Ryu, Y.; Masayuki, K.; Eiji, I.; David, W.R. Identification of a progenitor cell population destined to form fracture fibrocartilage callus in Dickkopf-related protein 3-green fluorescent protein reporter mice. J. Bone Miner. Metab. 2016, 34, 606–614. [Google Scholar] [CrossRef]
- Changming, Z.; Yulin, G.; Yan, W.; Qiaozhen, Q.; Ting, W.; Meng, H.; Heyang, Z.; Yannv, Q.; Jingwen, Z.; Zhangzhen, D.; et al. miR-129-5p Promotes Osteogenic Differentiation of BMSCs and Bone Regeneration via Repressing Dkk3. Stem Cells Int. 2021, 2021, 7435605. [Google Scholar] [CrossRef]
- Tomohiko, M.; Atsushi, S.; Shin-ichiro, H.; Shinichi, K.; Soshi, K.; Kazuyasu, C.; Hiroshi, S.; Kenji, T.; Kimiko, O.; Kazuya, Y.; et al. Signalling mediated by the endoplasmic reticulum stress transducer OASIS is involved in bone formation. Nat. Cell Biol. 2009, 11, 1205–1211. [Google Scholar] [CrossRef]
- Katarina, L.; Eva, Å.; Anca, D.; Sofie, S.; Paul, C.; Sune, L.; Eleftherios, P.; Paul, R.; Sonja, G.; Klaus, K.; et al. Homozygosity for CREB3L1 premature stop codon in first case of recessive osteogenesis imperfecta associated with OASIS-deficiency to survive infancy. Bone 2018, 114, 268–277. [Google Scholar] [CrossRef]
- Forlino, A.; Marini, J.C. Osteogenesis Imperfecta: Prospects for Molecular Therapeutics. Mol. Genet. Metab. 2000, 71, 225–232. [Google Scholar] [CrossRef] [Green Version]
- Moradifard, S.; Hoseinbeyki, M.; Emam, M.M.; Parchiniparchin, F.; Ebrahimi-Rad, M. Association of the Sp1 binding site and -1997 promoter variations in COL1A1 with osteoporosis risk: The application of meta-analysis and bioinformatics approaches offers a new perspective for future research. Mutat. Res. Mutat. Res. 2020, 786, 108339. [Google Scholar] [CrossRef]
- Tomohiko, M.; Shinichi, K.; Maiko, O.; Soshi, K.; Atsushi, S.; Akio, W.; Kazunori, I. Cleavage of the membrane-bound transcription factor OASIS in response to endoplasmic reticulum stress. J. Neurochem. 2006, 96, 1090–1100. [Google Scholar] [CrossRef]
- Masamine, T.; Yumiko, K. Severe osteogenesis imperfecta caused by CREB3L1 mutation in a cat. J. Vet. Diagn. Investig. 2022, 34, 558–563. [Google Scholar] [CrossRef]
- Ramesh, G.; Devika, N.; Srividya, G.; Venkat, R.G.; Prathap, R.K.; Ayesha, I. Disrupted expression of genes essential for skeletal muscle fibre integrity and energy metabolism in Vitamin D deficient rats. J. Steroid Biochem. Mol. Biol. 2019, 197, 105525. [Google Scholar] [CrossRef]
- Gong, Z.; Da, W.; Tian, Y.; Zhao, R.; Qiu, S.; Wu, Q.; Wen, K.; Shen, L.; Zhou, R.; Tao, L.; et al. Exogenous melatonin prevents type 1 diabetes mellitus–induced bone loss, probably by inhibiting senescence. Osteoporos. Int. 2021, 33, 453–466. [Google Scholar] [CrossRef]
- Lu, Y.; Ling, C.; Ruo-xi, L.; Yuan-yuan, L.; Yan, J.; Ou, W.; Mei, L.; Xiao-Ping, X.; Qian-Qian, P.; Ruizhi, J.; et al. A Common Mutation and a Novel Mutation in the HPGD Gene in Nine Patients with Primary Hypertrophic Osteoarthropathy. Calcif. Tissue Int. 2015, 97, 336–342. [Google Scholar] [CrossRef]
- Li, N.; Ma, Y.; Jiang, Y.; You, L.; Huang, Y.; Peng, Y.; Ding, X.; Zhao, L. Characterization of Mineral and Bone Metabolism Biomarkers in a Chinese Consanguineous Twin Family with Primary Hypertrophic Osteoarthropathy. Int. J. Endocrinol. 2020, 2020, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Yuan, G.; Li, Y.; Ling, S.; Xu, B.; Zhao, J.; Li, Y. Review of Drug Protection of Bone Loss Induced by Microgravity. Manned Spacefl. 2021, 27, 298–304. [Google Scholar]
- Zhu, X.; Cao, Y. Sclerostin monoclonal antibody: A future new drug for the treatment of osteoporosis. Chin. J. Osteoporos. 2012, 18, 568–572,541. [Google Scholar]
- Chow, Y.Y.; Chin, K.-Y. The Role of Inflammation in the Pathogenesis of Osteoarthritis. Mediat. Inflamm. 2020, 2020, 8293921. [Google Scholar] [CrossRef] [Green Version]
- Herrero-Beaumont, G.; Pérez-Baos, S.; Pernaute, O.S.; Roman-Blas, J.A.; Lamuedra, A.; Largo, R. Targeting chronic innate inflammatory pathways, the main road to prevention of osteoarthritis progression. Biochem. Pharmacol. 2019, 165, 24–32. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence (5′–3′) | |
|---|---|---|
| Forward | Reverse | |
| Creb 311 | AGCCTTGTGCTTCGTTCTGGTG | CATCGTAGAACAGTAGGCTTCGG |
| Mt 1 | AGGCTCTGGGGTTCATCTCT | GCGTGGTAACCTCAGCTTCT |
| Dkk | GGAGGAAGCTACGCTCAATGAG | TTGCCAGGTTCACCTCAGAGGA |
| Icn 2 | ATGTCACCTCCATCCTGGTCAG | GCCACTTGCACATTGTAGCTCTG |
| Clec 5a | GGTTTGGTACGTCAGCCTGGAG | GACACAGTCGAAGTTCTGGTCC |
| Ipgd | AAGCAAAACGGAGGTGAAGGCG | GAGCGTGTGAATCCGATGATGC |
| Tnnt | GAGCAGAGGATGACGCCAAGAA | TTCATCTCCCGACCAGTCTGTC |
| Gapdh | AAATGGTGAAGGTCGGTGTGAAC | AACAATCTCCACTTTGCCACTG |
| Gene | Fold Change | Gene | Fold Change | Gene | Fold Change |
|---|---|---|---|---|---|
| Mrln | 6.18 | Dupd1 | 4.54 | Mybph | 4.06 |
| Nctc1 | 5.82 | Scrg1 | 4.48 | Xirp2 | 4.04 |
| Saa3 | 5.37 | Mpo | 4.43 | Ucma | 4.04 |
| Acta1 | 4.94 | Tpsab1 | 4.36 | Tcap | 3.95 |
| Myot | 4.81 | Prr33 | 4.34 | Tmem52 | 3.93 |
| Eef1a2 | 4.79 | Retnla | 4.15 | Plac8 | 3.89 |
| Mylk2 | 4.71 | Clec3a | 4.14 | Otub2 | 3.78 |
| 2310002L09Rik | 4.71 | Hapln1 | 4.12 | Myoz2 | 3.74 |
| Smpx | 4.59 | Cox8b | 4.12 | Ano5 | 3.67 |
| Trdn | 4.56 | Asb12 | 4.10 | Mpz | 3.65 |
| Gene | Locations |
|---|---|
| Lcn2 | Bone-marrow-derived macrophages, femur and humerus, tendon |
| Creb3l1 | Bone-marrow-derived macrophages, femur and humerus, tendon |
| Dkk3 | Bone-marrow-derived macrophages, femur and humerus, tendon |
| Clec5a | Bone-marrow-derived macrophages, femur and humerus, tendon |
| Tspan32 | Bone-marrow-derived macrophages, femur and humerus, tendon |
| Mt1 | Bone-marrow-derived macrophages, femur and humerus, tendon |
| Hpgd | Bone-marrow-derived macrophages, femur and humerus, tendon |
| Fxyd2 | Bone-marrow-derived macrophages, cortical bone, tendon |
| Tnnt1 | Bone-marrow-derived macrophages, cortical bone, tendon |
| Cox8b | Femur and humerus, cortical bone, tendon |
| Art1 | Femur and humerus, cortical bone, tendon |
| Obscn | Femur and humerus, cortical bone, tendon |
| Pathway | Enriched Genes |
|---|---|
| Hypertrophic cardiomyopathy | Dmd, Des, Cacng1, Myl2, Myl3, Tnnc1, Ttn, Igf1, Prkaa2, Tpm2 |
| Cardiac muscle contraction | Cox8b, Fxyd2, Cacng1, Atp1a2, Myl2, Myl3, Tnnc1, Trdn, Tpm2 |
| Dilated cardiomyopathy | Dmd, Des, Cacng1, Myl2, Myl3, Tnnc1, Ttn, Igf1, Tpm2 |
| ECM-receptor interaction | Col6a3, Comp, Chad, Thbs4, Sdc1, Col4a5, Thbs2 |
| Protein digestion and absorption | Col6a3, Col10a1, Col8a2, Fxyd2, Atp1a2, Col5a2, Col4a5 |
| Adrenergic signaling in cardiomyocytes | Creb3l1, Fxyd2, Cacng1, Atp1a2, Myl2, Myl3, Tnnc1, Tpm2 |
| Focal adhesion | Col6a3, Comp, Chad, Mylk2, Myl2, Thbs4, Igf1, Col4a5, Thbs2 |
| Mineral absorption | Mt1, Vdr, Fxyd2, Atp1a2, Mt2 |
| Phagosome | Mpo, Mrc2, Comp, Thbs4, Nos1, Coro1a, Thbs2 |
| Complement and coagulation cascades | Cd59a, C1qa, F13a1, Serping1, Cfb |
| Gene | Degree | Gene | Degree | Gene | Degree | Gene | Degree |
|---|---|---|---|---|---|---|---|
| Ttn | 34 | Nrap | 21 | Klhl41 | 16 | Efemp2 | 13 |
| Acta1 | 31 | Synpo2l | 20 | Col6a3 | 16 | Pcolce | 13 |
| Tcap | 30 | Lmod2 | 20 | Timp1 | 16 | Fcer1g | 12 |
| Myoz2 | 29 | Col5a2 | 19 | Lox | 15 | Acan | 12 |
| Myl2 | 28 | Xirp2 | 19 | Des | 15 | Apobec2 | 11 |
| Obscn | 27 | Tpm2 | 18 | Thbs2 | 15 | Smyd1 | 11 |
| Myl3 | 27 | Sparc | 18 | Adprhl1 | 15 | Eef1a2 | 11 |
| Srl | 25 | Txlnb | 17 | Cd68 | 15 | Vcan | 11 |
| Smpx | 25 | Myo18b | 17 | Tnnc1 | 15 | Col4a5 | 11 |
| Trdn | 25 | Igf1 | 17 | C1qa | 15 | Sdc1 | 10 |
| Myot | 24 | Tnni1 | 17 | Serpinh1 | 14 | Ankrd23 | 10 |
| Dmd | 22 | Tnnt1 | 16 | Adamts2 | 13 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Zhang, C.; Zhou, Z.; Zhang, C.; Wang, J. Identification of Key Genes and Pathways Associated with PIEZO1 in Bone-Related Disease Based on Bioinformatics. Int. J. Mol. Sci. 2022, 23, 5250. https://doi.org/10.3390/ijms23095250
Zhou Y, Zhang C, Zhou Z, Zhang C, Wang J. Identification of Key Genes and Pathways Associated with PIEZO1 in Bone-Related Disease Based on Bioinformatics. International Journal of Molecular Sciences. 2022; 23(9):5250. https://doi.org/10.3390/ijms23095250
Chicago/Turabian StyleZhou, Yuanyuan, Chen Zhang, Zhongguo Zhou, Chao Zhang, and Jiali Wang. 2022. "Identification of Key Genes and Pathways Associated with PIEZO1 in Bone-Related Disease Based on Bioinformatics" International Journal of Molecular Sciences 23, no. 9: 5250. https://doi.org/10.3390/ijms23095250