
Characterization of a Thermostable and Surfactant-Tolerant Chondroitinase B from a Marine Bacterium Microbulbifer sp. ALW1

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
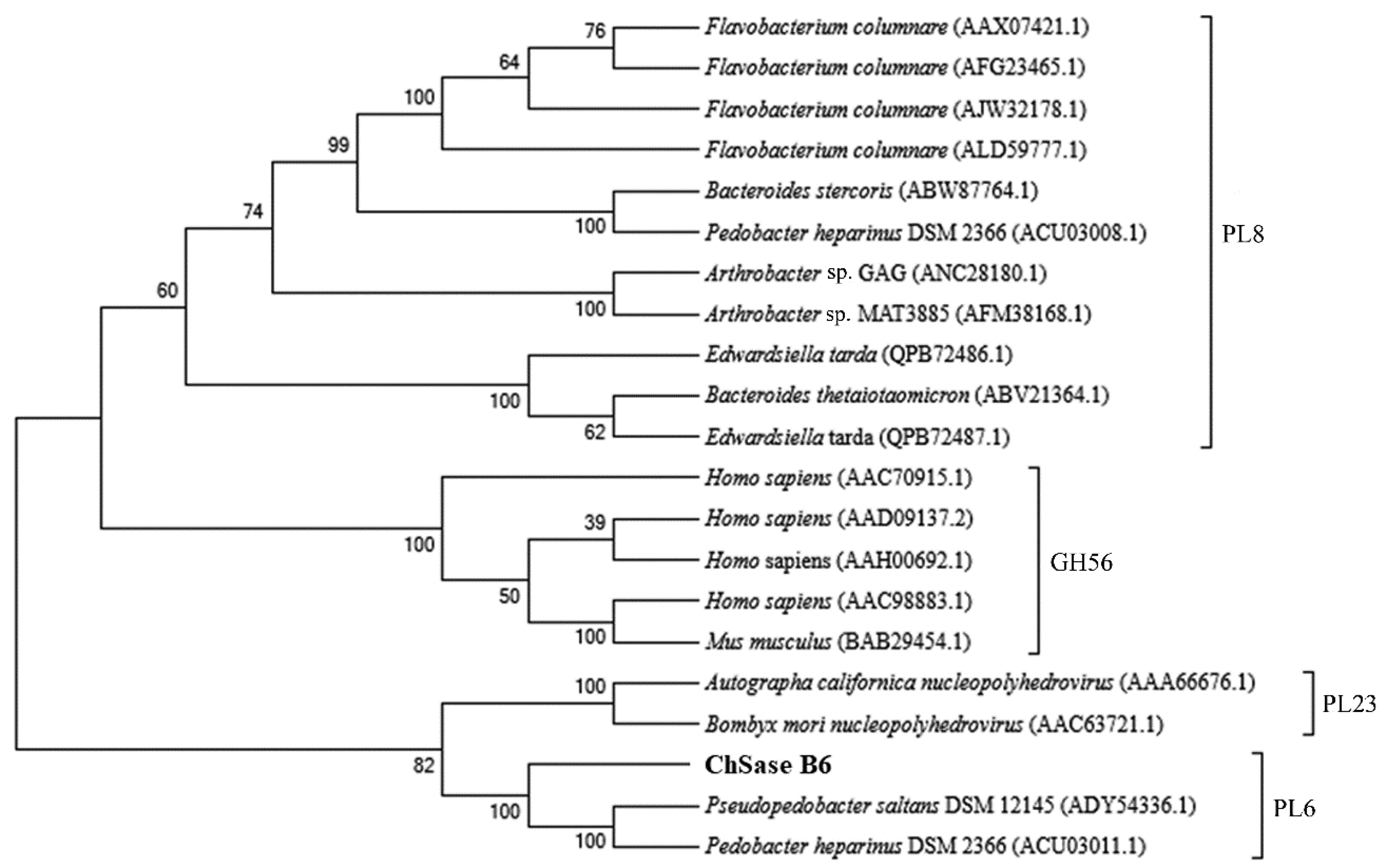
2.1. Sequence Analysis of Chondroitinase
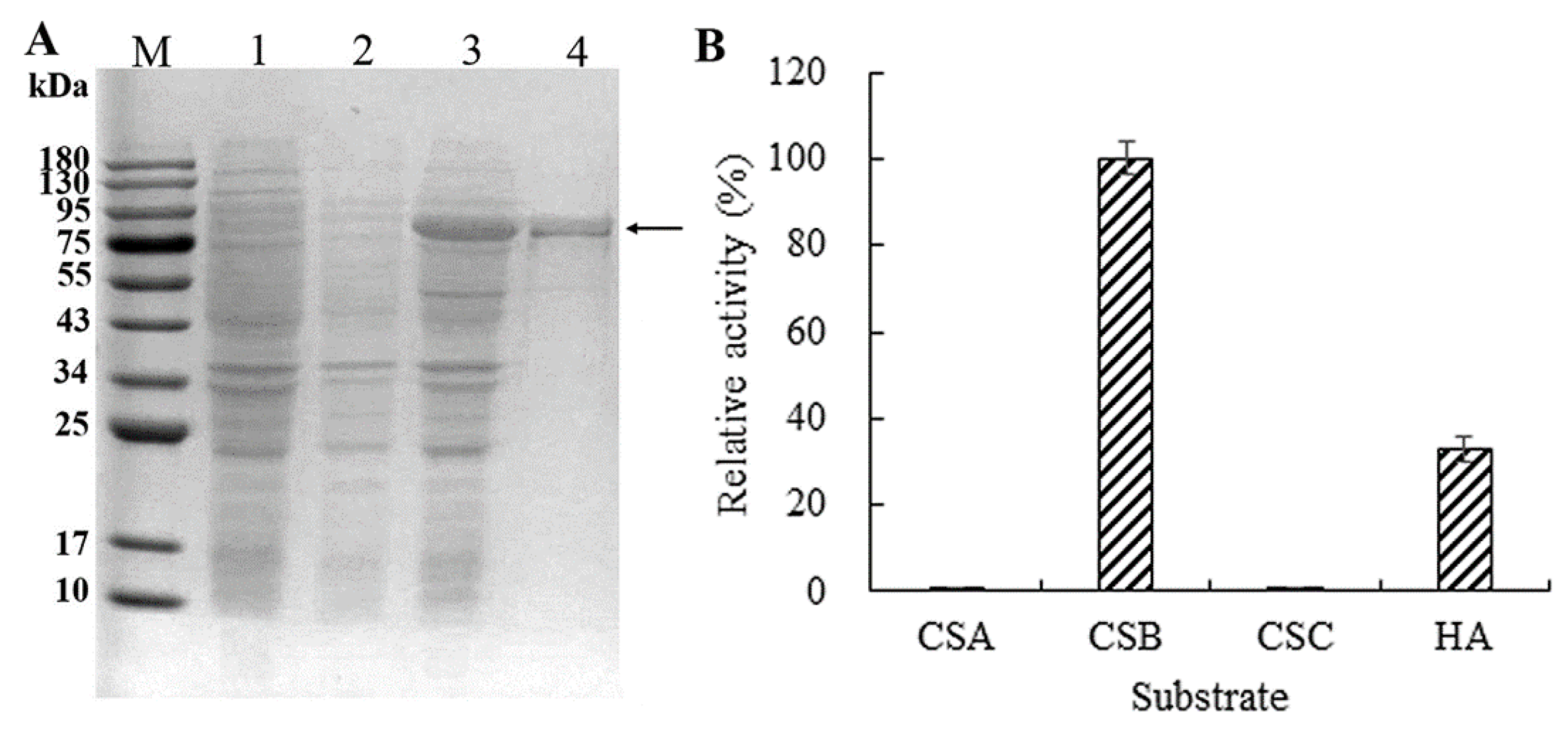
2.2. Chondroitinase Expression and Substrate Specificity
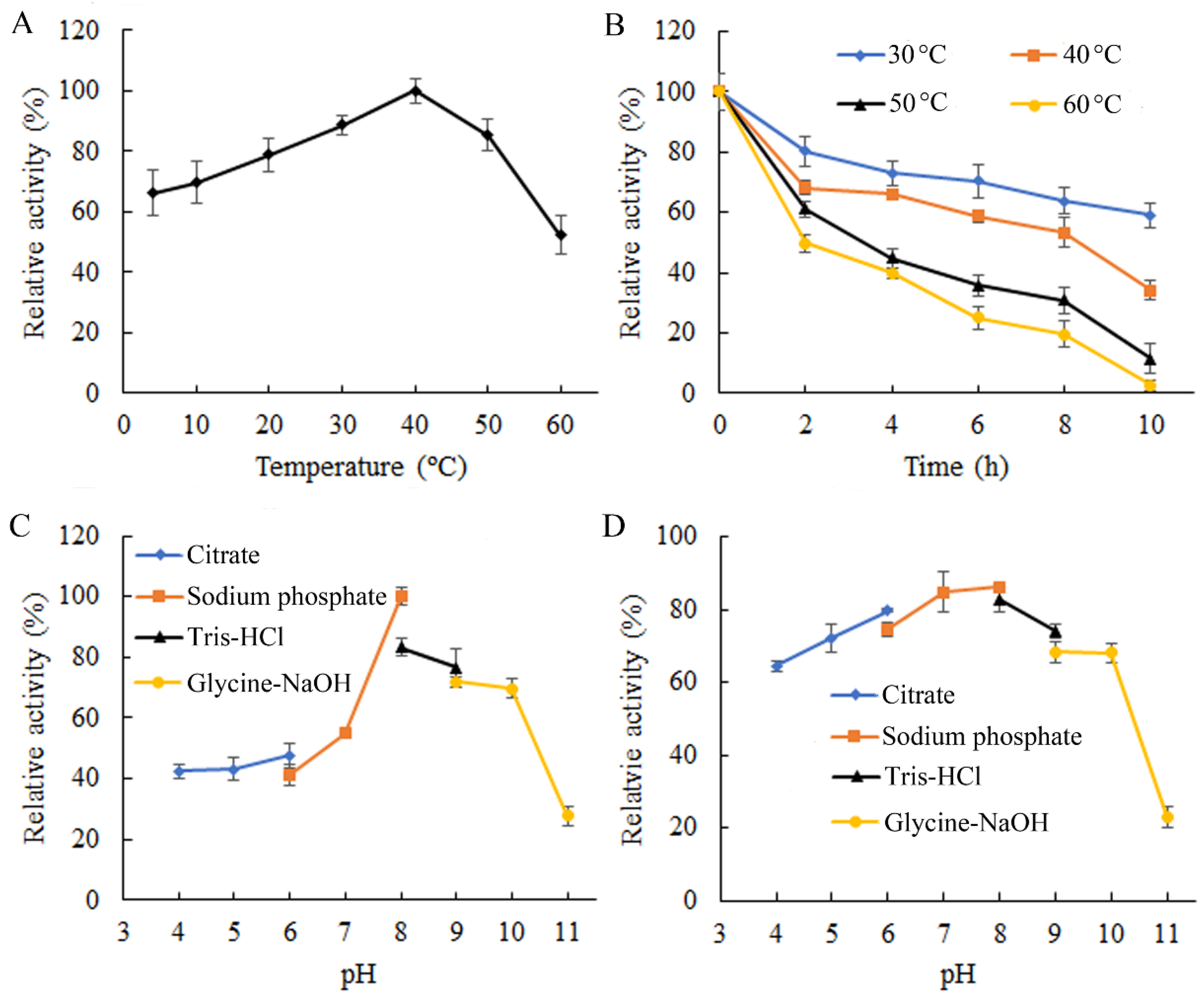
2.3. Enzymatic Properties of ChSase B6
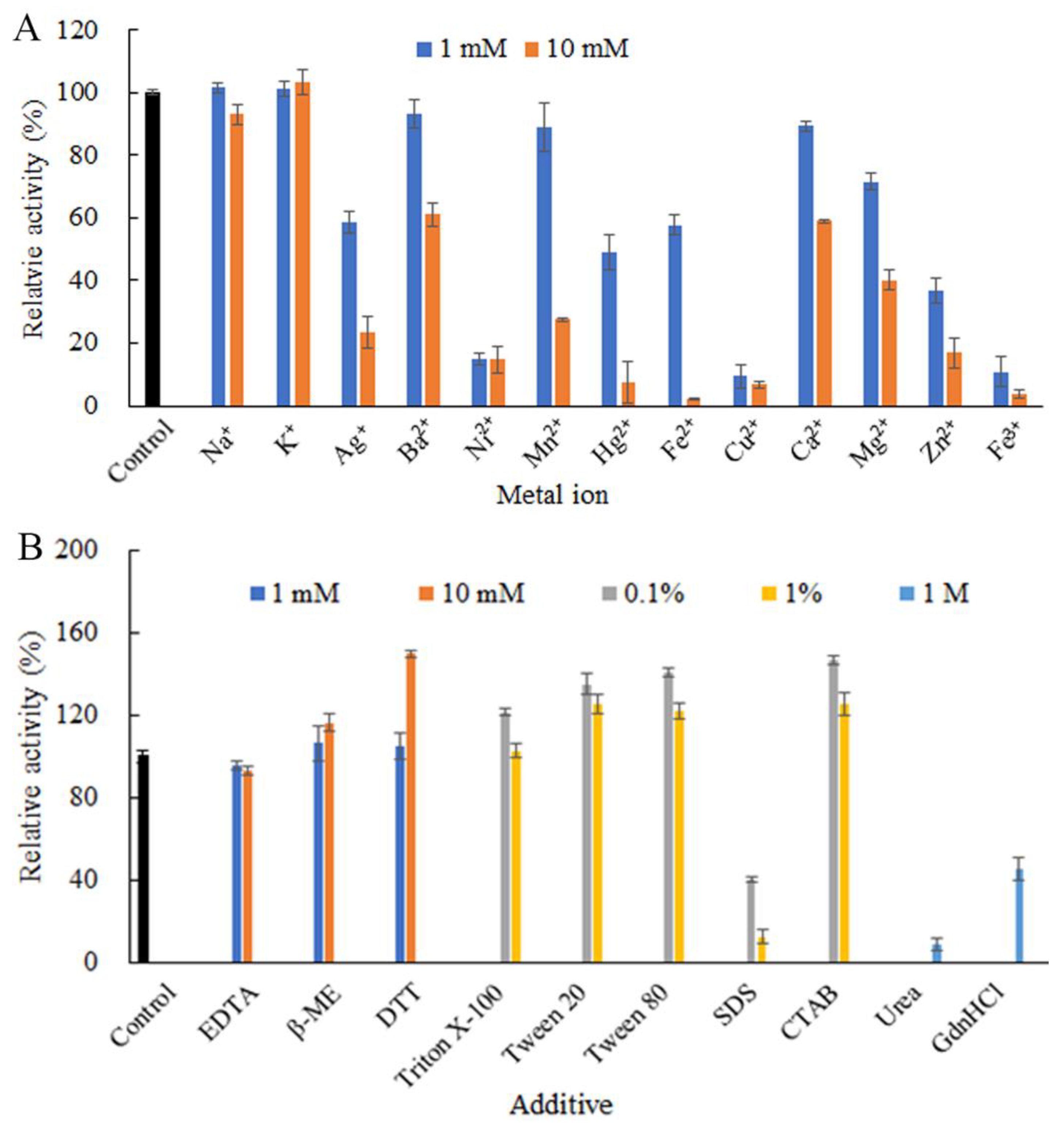
2.4. Effects of Chemicals on ChSase B6 Stability
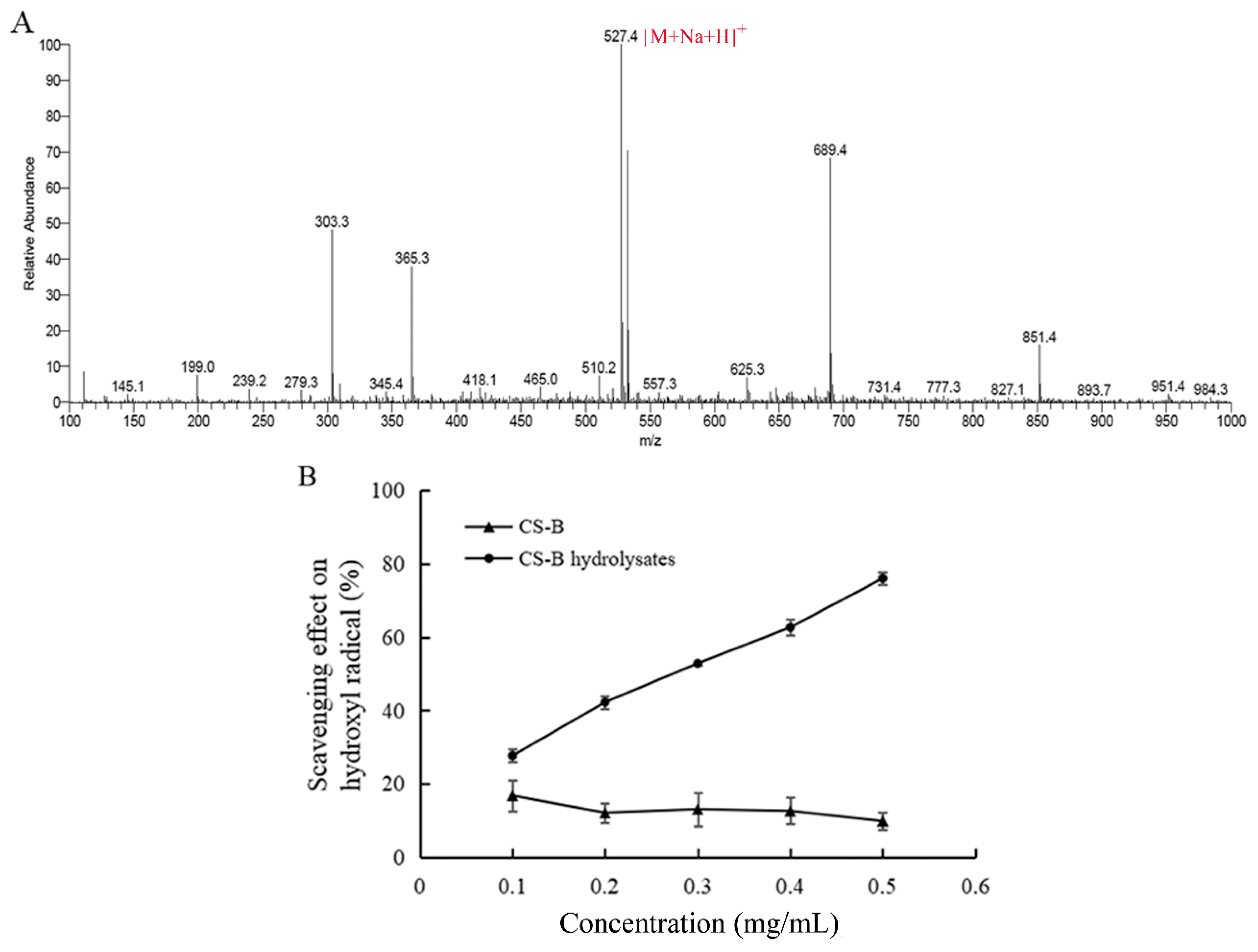
2.5. Hydroxyl Radical Scavenging Activity of the Enzymatic Products
2.6. CD Spectroscopy of ChSase B6
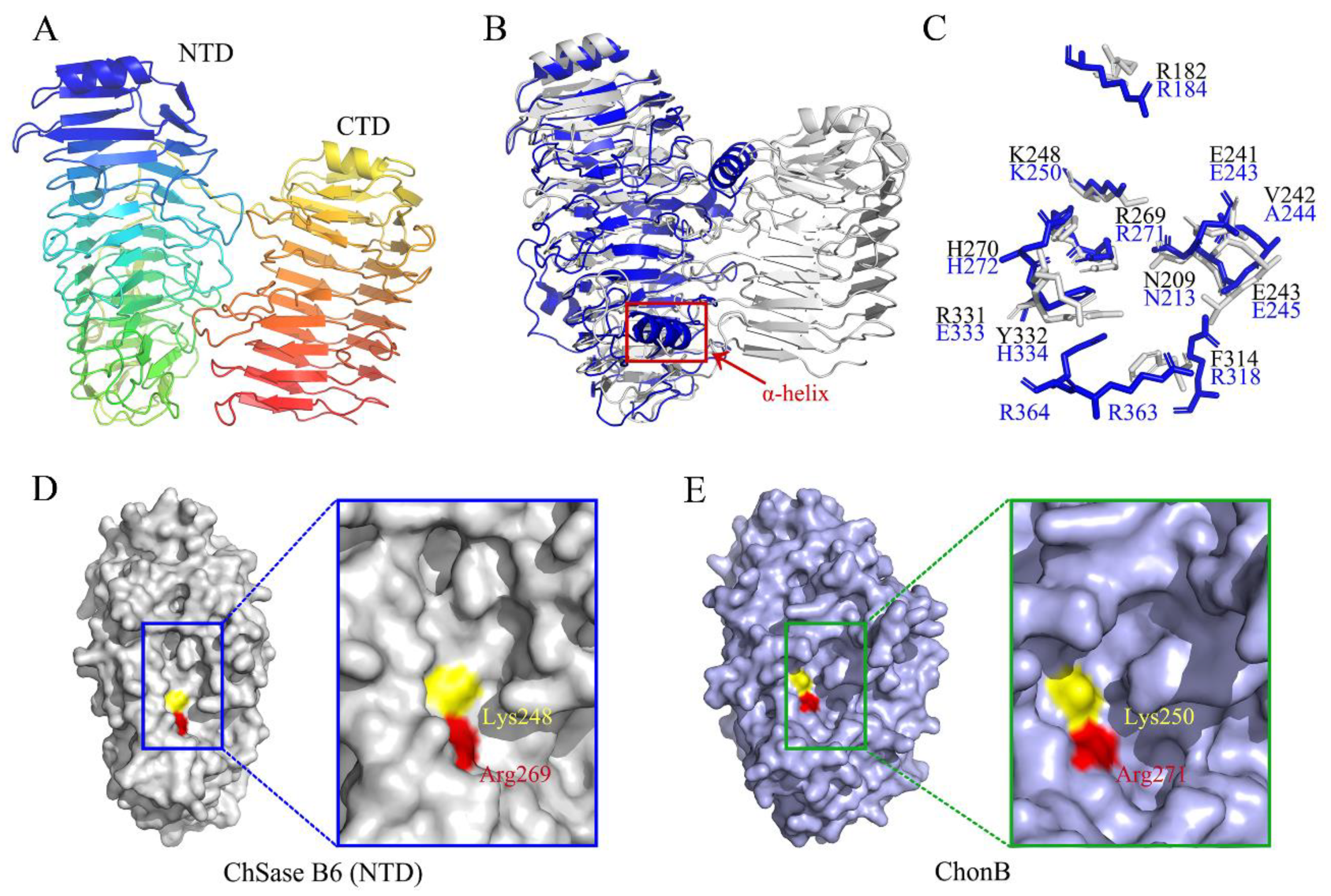
2.7. Structure Modeling
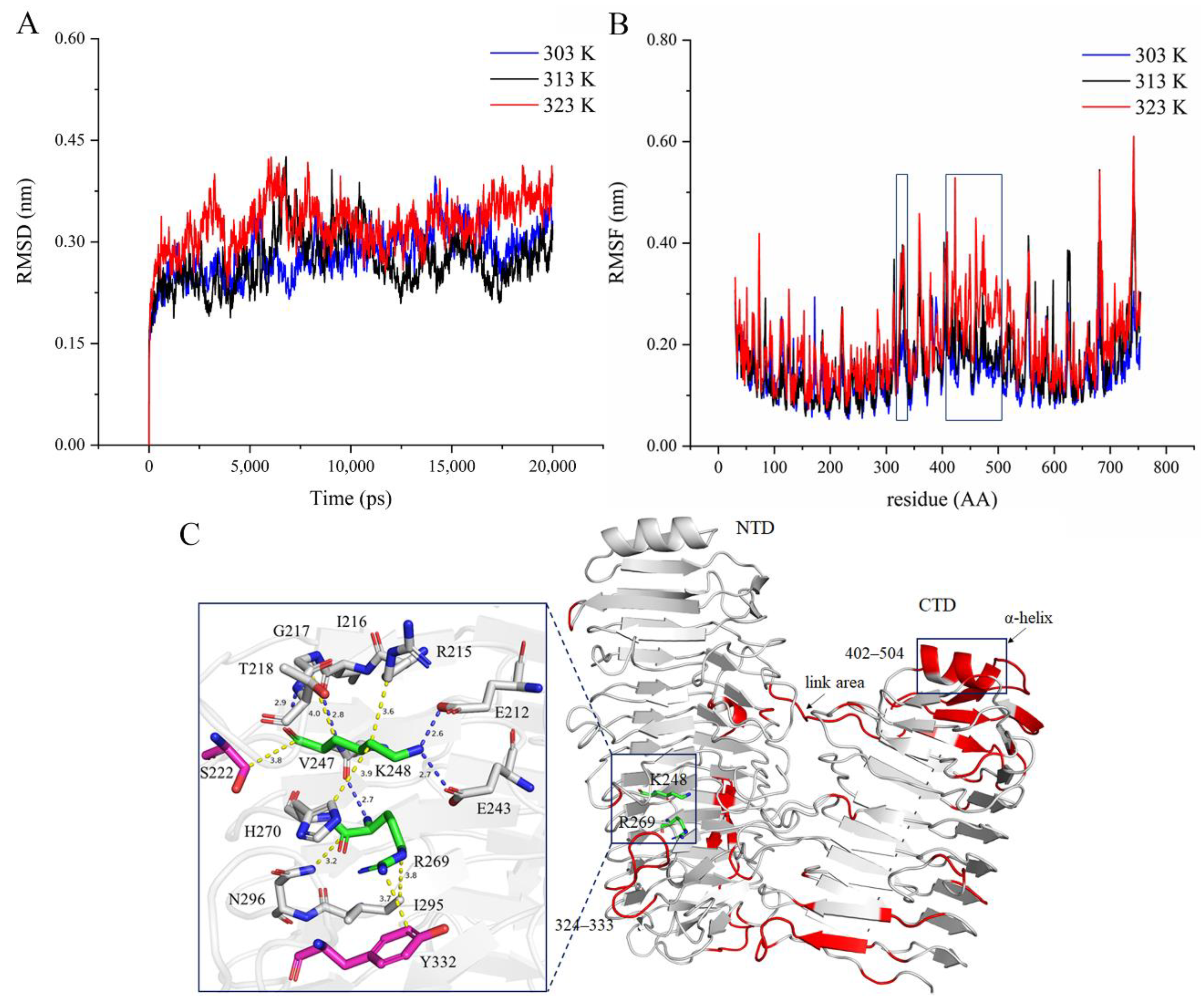
2.8. Thermal Adaptation Analysis by MD Simulation
3. Materials and Methods
3.1. Sequence and Structure Analysis
3.2. Recombinant Construction and Protein Expression
3.3. Circular Dichroism (CD) Spectroscopy
3.4. Enzyme Activity Assay
3.5. Substrate Specificity
3.6. Enzyme Property Assay
3.7. Effects of Chemicals on Enzyme Stability
3.8. Preparation and Identification of Enzymatic Products
3.9. Hydroxyl Radical Scavenging Assay
3.10. Site-Directed Mutagenesis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nikitovic, D.; Pérez, S. Preface for the special issue on the exploration of the multifaceted roles of glycosaminoglycans: GAGs. Biomolecules 2021, 11, 1630. [Google Scholar] [CrossRef] [PubMed]
- Köwitsch, A.; Zhou, G.; Groth, T. Medical application of glycosaminoglycans: A review. J. Tissue Eng. Regen. Med. 2018, 12, e23–e41. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Shi, L.; Qin, Y.; Li, F. Research and application of chondroitin sulfate/dermatan sulfate-degrading enzymes. Front. Cell Dev. Biol. 2020, 8, 560442. [Google Scholar] [CrossRef]
- Yamada, S.; Matsushima, K.; Ura, H.; Miyamoto, N.; Sugahara, K. Mass preparation of oligosaccharides by the hydrolysis of chondroitin sulfate polysaccharides with a subcritical water microreaction system. Carbohydr. Res. 2013, 371, 16–21. [Google Scholar] [CrossRef]
- Yan, L.; Li, J.; Wang, D.; Ding, T.; Hu, Y.; Ye, X.; Linhardt, R.J.; Chen, S. Molecular size is important for the safety and selective inhibition of intrinsic factor Xase for fucosylated chondroitin sulfate. Carbohydr. Polym. 2017, 178, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Li, J.; Liu, C.; Song, C.; Li, P.; Yin, F.; Xiao, Y.; Li, J.; Jiang, W.; Zong, A.; et al. Protective effects of low molecuiar weight chondroitin sulfate on amyloid beta (Aβ)-induced damage in vitro and in vivo. Neuroscience 2015, 305, 169–182. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Nagarkatti, P.; Zhong, Y.; Nagarkatti, M. Immune modulation by chondroitin sulfate and its degraded disaccharide product in the development of an experimental model of multiple sclerosis. J. Neuroimmunol. 2010, 223, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Li, S.; Zhi, Z.; Yan, L.; Ye, X.; Ding, T.; Yan, L.; Linhardt, R.J.; Chen, S. Depolymerization of fucosylated chondroitin sulfate with a modified fenton-system and anticoagulant activity of the resulting fragments. Mar. Drugs 2016, 14, 170. [Google Scholar] [CrossRef] [Green Version]
- Wu, N.; Ye, X.; Guo, X.; Liao, N.; Yin, X.; Hu, Y.; Sun, Y.; Liu, D.; Chen, S. Depolymerization of fucosylated chondroitin sulfate from sea cucumber, Pearsonothuria graeffei, via 60Co irradiation. Carbohydr. Polym. 2013, 93, 604–614. [Google Scholar] [CrossRef]
- de Souza, J.F.; Lessa, E.F.; Nörnberg, A.; Gularte, M.S.; Quadrado, R.F.N.; Fajardo, A.R. Enzymatic depolymerization–An easy approach to reduce the chondroitin sulfate molecular weight. Process Biochem. 2018, 74, 118–124. [Google Scholar] [CrossRef]
- Fu, J.; Jiang, Z.; Chang, J.; Han, B.; Liu, W.; Peng, Y. Purification, characterization of Chondroitinase ABC from Sphingomonas paucimobilis and in vitro cardiocytoprotection of the enzymatically degraded CS-A. Int. J. Biol. Macromol. 2018, 115, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Valcarcel, J.; García, M.R.; Sampayo, L.F.; Antonio Vázquez, J. Marine chondroitin sulfate of defined molecular weight by enzymatic depolymerization. Carbohydr. Polym. 2020, 229, 115450. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, J.; Li, F. Hyaluronidase and Chondroitinase. Adv. Exp. Med. Biol. 2017, 925, 75–87. [Google Scholar] [PubMed]
- Garron, M.-L.; Cygler, M. Structural and mechanistic classification of uronic acid-containing polysaccharide lyases. Glycobiology 2010, 20, 1547–1573. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Lu, D.; Wang, S.; Wei, L.; Wang, W.; Li, F. Identification and biochemical characterization of a novel chondroitin sulfate/dermantan sulfate lyase from Photobacterium sp. Int. J. Biol. Macromol. 2020, 165, 2314–2325. [Google Scholar] [CrossRef] [PubMed]
- Kale, V.; Friðjónsson, Ó.; Jónsson, J.Ó.; Kristinsson, H.G.; Ómarsdóttir, S.; Hreggviðsson, G.Ó. Chondroitin lyase from a marine Arthrobacter sp. MAT3885 for the production of chondroitin sulfate disaccharides. Mar. Biotechnol. 2015, 17, 479–492. [Google Scholar] [CrossRef]
- Li, F.; Nandini, C.D.; Hattori, T.; Bao, X.; Murayama, D.; Nakamura, T.; Fukushima, N.; Sugahara, K. Structure of pleiotrophin- and hepatocyte growth factor-binding sulfated hexasaccharide determined by biochemical and computational approaches. J. Biol. Chem. 2010, 285, 27673–27685. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Cai, X.; Han, N.; Han, W.; Sugahara, K.; Li, F. Sequencing of chondroitin sulfate oligosaccharides using a novel exolyase from a marine bacterium that degrades hyaluronan and chondroitin sulfate/dermatan sulfate. Biochem. J. 2017, 474, 3831–3848. [Google Scholar] [CrossRef]
- Han, W.; Wang, W.; Zhao, M.; Sugahara, K.; Li, F. A novel eliminase from a marine bacterium that degrades hyaluronan and chondroitin sulfate. J. Biol. Chem. 2014, 289, 27886–27898. [Google Scholar] [CrossRef] [Green Version]
- Linhardt, R.J.; Avci, F.Y.; Toida, T.; Kim, Y.S.; Cygler, M. CS lyases: Structure, activity, and applications in analysis and the treatment of diseases. Adv. Pharmacol. 2006, 53, 187–215. [Google Scholar]
- Rani, A.; Patel, S.; Goyal, A. Chondroitin sulfate (CS) lyases: Structure, function and application in therapeutics. Curr. Protein. Pept. Sci. 2018, 19, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Wu, L.; Chen, Y.; Ni, H.; Xiao, A.; Cai, H. Characterization of an extracellular biofunctional alginate lyase from marine Microbulbifer sp. ALW1 and antioxidant activity of enzymatic hydrolysates. Microbiol. Res. 2016, 182, 49–58. [Google Scholar] [CrossRef]
- Cox, J.; Mann, M. Quantitative, high-resolution proteomics for data-driven systems biology. Annu. Rev. Biochem. 2011, 80, 273–299. [Google Scholar] [CrossRef] [PubMed]
- Spliid, C.B.; Toledo, A.G.; Salanti, A.; Esko, J.D.; Clausen, T.M. Beware, commercial chondroitinases vary in activity and substrate specificity. Glycobiology 2021, 31, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Su, H.; Wang, X.; Tang, L.; Hu, J.; Yu, W.; Han, F. Cloning and characterization of a novel chondroitinase ABC categorized into a new subfamily of polysaccharide lyase family 8. Int. J. Biol. Macromol. 2020, 164, 3762–3770. [Google Scholar] [CrossRef] [PubMed]
- Michel, G.; Pojasek, K.; Li, Y.; Sulea, T.; Linhardt, R.J.; Raman, R.; Prabhakar, V.; Sasisekharan, R.; Cygler, M. The structure of chondroitin B lyase complexed with glycosaminoglycan oligosaccharides unravels a calcium-dependent catalytic machinery. J. Biol. Chem. 2004, 279, 32882–32896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tkalec, A.L.; Fink, D.; Blain, F.; Zhang-Sun, G.; Laliberte, M.; Bennett, D.C.; Gu, K.; Zimmermann, J.J.; Su, H. Isolation and expression in Escherichia coli of cslA and cslB, genes coding for the chondroitin sulfate-degrading enzymes chondroitinase AC and chondroitinase B, respectively, from Flavobacterium heparinum. Appl. Environ. Microbiol. 2000, 66, 29–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.Y.; Li, Y.; Feng, Y.; Chen, L.; Yuan, Q.P. Enzyme activity enhancement of chondroitinase ABC I from Proteus vulgaris by site-directed mutagenesis. RSC Adv. 2015, 5, 76040–76047. [Google Scholar] [CrossRef]
- Andreini, C.; Bertini, I.; Cavallaro, G.; Holliday, G.L.; Thornton, J.M. Metal ions in biological catalysis: From enzyme databases to general principles. J. Biol. Inorg. Chem. 2008, 13, 1205–1218. [Google Scholar] [CrossRef]
- Holmberg, K. Interactions between surfactants and hydrolytic enzymes. Colloids Surf. B Biointerfaces 2018, 168, 169–177. [Google Scholar] [CrossRef]
- Schiller, J.; Arnhold, J.; Benard, S.; Reichl, S.; Arnold, K. Cartilage degradation by hyaluronate lyase and chondroitin ABC lyase: A MALDI-TOF mass spectrometric study. Carbohydr. Res. 1999, 318, 116–122. [Google Scholar] [CrossRef]
- Sun, Y.; Yang, B.; Wu, Y.; Liu, Y.; Gu, X.; Zhang, H.; Wang, C.; Cao, H.; Huang, L.; Wang, Z. Structural characterization and antioxidant activities of κ-carrageenan oligosaccharides degraded by different methods. Food Chem. 2015, 178, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Matte, A.; Li, Y.; Kim, Y.S.; Linhardt, R.J.; Su, H.; Cygler, M. Crystal structure of chondroitinase B from Flavobacterium heparinum and its complex with a disaccharide product at 1.7 Å resolution. J. Mol. Biol. 1999, 294, 1257–1269. [Google Scholar] [CrossRef]
- Bernardi, R.C.; Cann, I.; Schulten, K. Molecular dynamics study of enhanced Man5B enzymatic activity. Biotechnol. Biofuels 2014, 7, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parra-Cruz, R.; Jäger, C.M.; Lau, P.L.; Gomes, R.L.; Pordea, A. Rational design of thermostable carbonic anhydrase mutants using molecular dynamics simulations. J. Phys. Chem. B 2018, 122, 8526–8536. [Google Scholar] [CrossRef] [PubMed]
- Kheirollahi, A.; Khajeh, K.; Golestani, A. Rigidifying flexible sites: An approach to improve stability of chondroitinase ABC I. Int. J. Biol. Macromol. 2017, 97, 270–278. [Google Scholar] [CrossRef]
- Piovesan, D.; Minervini, G.; Tosatto, S.C.E. The RING 2.0 web server for high quality residue interaction networks. Nucleic Acids Res. 2016, 44, W367–W374. [Google Scholar] [CrossRef] [PubMed]
- Schmid, N.; Eichenberger, A.P.; Choutko, A.; Riniker, S.; Winger, M.; Mark, A.E.; van Gunsteren, W.F. Definition and testing of the GROMOS force-field versions 54A7 and 54B7. Eur. Biophys. J. 2011, 40, 843–856. [Google Scholar] [CrossRef]
- Li, L.; Li, W.; Gong, J.; Xu, Y.; Wu, Z.; Jiang, Z.; Cheng, Y.; Li, Q.; Ni, H. An effective computational-screening strategy for simultaneously improving both catalytic activity and thermostability of α-L-rhamnosidase. Biotechnol. Bioeng. 2021, 118, 3409–3419. [Google Scholar] [CrossRef]
- Jiang, Z.; Guo, Y.; Wang, X.; Li, H.; Ni, H.; Li, L.; Xiao, A.; Zhu, Y. Molecular cloning and characterization of AlgL17, a new exo-oligoalginate lyase from Microbulbifer sp. ALW1. Protein Expr. Purif. 2019, 161, 17–27. [Google Scholar] [CrossRef]
- Li, Z.; Huang, X.; Guo, Y.; Zhang, C.; Yang, L.; Du, X.; Ni, H.; Wang, X.; Zhu, Y. Toward understanding the alginate catabolism in Microbulbifer sp. ALW1 by proteomics profiling. Front. Bioeng. Biotechnol. 2022, 10, 829428. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sequence | Length (aa) | Position (aa) | Score | Mass (Da) | Fraction Average | Retention Time (Min) |
|---|---|---|---|---|---|---|
| KLEAGDSVR | 9 | 48–56 | 99 | 487.7671 | 2 | 6.95 |
| LANGVWK | 7 | 57–63 | 96.27 | 394.7161 | 2 | 17.56667 |
| DFEILFTGNGEK | 12 | 64–75 | 99 | 685.8295 | 2 | 5.36 × 100.01 |
| VILSGQSNLR | 10 | 89–98 | 98.61 | 544.3088 | 2 | 19.11667 |
| LAGDHLVVSGLVFK | 14 | 99–112 | 99 | 485.617 | 3 | 44.71667 |
| DHLANHSRVTEVVIEDFSKPER | 22 | 129–150 | 32.56 | 868.4452 | 3 | 67.6 |
| MESDYWVGIY | 10 | 151–160 | 99 | 631.7729 | 2 | 61.46667 |
| FDHSHLAGK | 9 | 166–174 | 99 | 506.2507 | 2 | 7.133333 |
| GVTMAVR | 7 | 178–184 | 99 | 375.2045 | 2 | 7.016667 |
| LDSEQSQENR | 10 | 184–193 | 99 | 603.2728 | 2 | 5.333333 |
| PVLGSNGGETLR | 12 | 206–217 | 99 | 600.814 | 2 | 19.96667 |
| IGTSHYSLTDSMTLVENNFFDR | 22 | 218–239 | 99 | 855.0704 | 3 | 50.56667 |
| NNTFYESR | 8 | 258–265 | 99 | 516.2231 | 2 | 16.15 |
| IFLGNGVDHTGGIR | 14 | 282–295 | 99 | 728.8798 | 2 | 28.83333 |
| VINADQVIR | 9 | 296–304 | 99 | 535.801 | 2 | 40.43333 |
| NNYLEGLTGYR | 11 | 305–315 | 61.11 | 572.2689 | 2 | 51.91667 |
| FGSGF | 5 | 316–320 | 25.78 | 514.2279 | 1 | 24.9 |
| TVMNGVPNSPINR | 13 | 321–333 | 99 | 700.352 | 2 | 22.88333 |
| YHQVVNAK | 8 | 334–341 | 99 | 479.7429 | 2 | 5.85 |
| IEHNSFINVEHIYL | 14 | 342–355 | 79.68 | 864.4382 | 2 | 49.95 |
| DSTISDNIFYTNNGK | 15 | 369–383 | 99 | 845.3834 | 2 | 38.58333 |
| SPFSVFDDISGITF | 14 | 384–397 | 98.53 | 766.3783 | 2 | 70.1 |
| AKNGLLYPIDSAAASK | 16 | 424–439 | 99 | 810.4335 | 2 | 29.9 |
| DFEILFTGNGEK | 12 | 449–460 | 99 | 685.8273 | 2 | 53.75 |
| IHGGINK | 7 | 450–456 | 18.2 | 369.7809 | 2 | 12.733 |
| EMTGVSWYPK | 10 | 463–472 | 99 | 599.2839 | 2 | 33.36666 |
| SEPVTPFDSGK | 11 | 473–483 | 99 | 582.2825 | 2 | 18.85 |
| AIAGAADGDTLMLENGTYNARK | 22 | 497–518 | 24.37 | 708.9996 | 3 | 37.35 |
| TAVWGMLK | 8 | 570–577 | 95.37 | 777.3964 | 1 | 60.08333 |
| FSMSNSR | 7 | 581–587 | 97.18 | 415.1802 | 2 | 11.58333 |
| VENLDINHSYHFFDSGHR | 18 | 588–605 | 99 | 730.0019 | 3 | 32.3 |
| ISLSNNL | 7 | 612–618 | 98.04 | 380.7134 | 2 | 32.15 |
| EVVIEDFSKPER | 12 | 619–630 | 99 | 724.3733 | 2 | 32.1 |
| NITGDLLK | 8 | 632–639 | 99 | 437.2512 | 2 | 26.7 |
| EQDDLGIYNAE | 11 | 643–653 | 35.16 | 633.7799 | 2 | 32.41667 |
| TFDQVQGALVDLYR | 14 | 661–674 | 99 | 812.9194 | 2 | 54.75 |
| GGTDESTFGPHIDFSK | 16 | 675–690 | 99 | 565.5933 | 3 | 32.38334 |
| NKAQALIK | 8 | 702–709 | 25.37 | 414.7721 | 2 | 17.31667 |
| LHGTQVADIQDNR | 13 | 711–723 | 99 | 489.5826 | 3 | 10.81667 |
| APLPEVTELVAEGPHTAVLANNEVTN | 26 | 750–775 | 99 | 895.7964 | 3 | 5.87 × 100.01 |
| Source | Substrate Specificity | Optimal Temperature (°C) | Optimal pH | Thermal Stability | Main Products | Reference |
|---|---|---|---|---|---|---|
| Vibrio sp. QY108 | CSA/CSB/CSC/CSD | 30 | 7.6 | completely inactivated after 40 °C for 1 h | disaccharides | [25] |
| Pedobacter heparinus | DS | 30 | 8.0 | ND | disaccharides | [26] |
| Flavobacterium heparinum | CSB | 30 | 8.0 | ND | ND | [27] |
| Proteus vulgaris | CSA/CSB | 37 | 7.4 | ND | ND | [28] |
| Microbulbifer sp. ALW1 | CSB | 40 | 8.0 | 49.7% residual activity after 60 °C for 2 h | disaccharides | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mou, M.; Hu, Q.; Li, H.; Long, L.; Li, Z.; Du, X.; Jiang, Z.; Ni, H.; Zhu, Y. Characterization of a Thermostable and Surfactant-Tolerant Chondroitinase B from a Marine Bacterium Microbulbifer sp. ALW1. Int. J. Mol. Sci. 2022, 23, 5008. https://doi.org/10.3390/ijms23095008
Mou M, Hu Q, Li H, Long L, Li Z, Du X, Jiang Z, Ni H, Zhu Y. Characterization of a Thermostable and Surfactant-Tolerant Chondroitinase B from a Marine Bacterium Microbulbifer sp. ALW1. International Journal of Molecular Sciences. 2022; 23(9):5008. https://doi.org/10.3390/ijms23095008
Chicago/Turabian StyleMou, Mingjing, Qingsong Hu, Hebin Li, Liufei Long, Zhipeng Li, Xiping Du, Zedong Jiang, Hui Ni, and Yanbing Zhu. 2022. "Characterization of a Thermostable and Surfactant-Tolerant Chondroitinase B from a Marine Bacterium Microbulbifer sp. ALW1" International Journal of Molecular Sciences 23, no. 9: 5008. https://doi.org/10.3390/ijms23095008