
Reduced Glx and GABA Inductions in the Anterior Cingulate Cortex and Caudate Nucleus Are Related to Impaired Control of Attention in Attention-Deficit/Hyperactivity Disorder

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
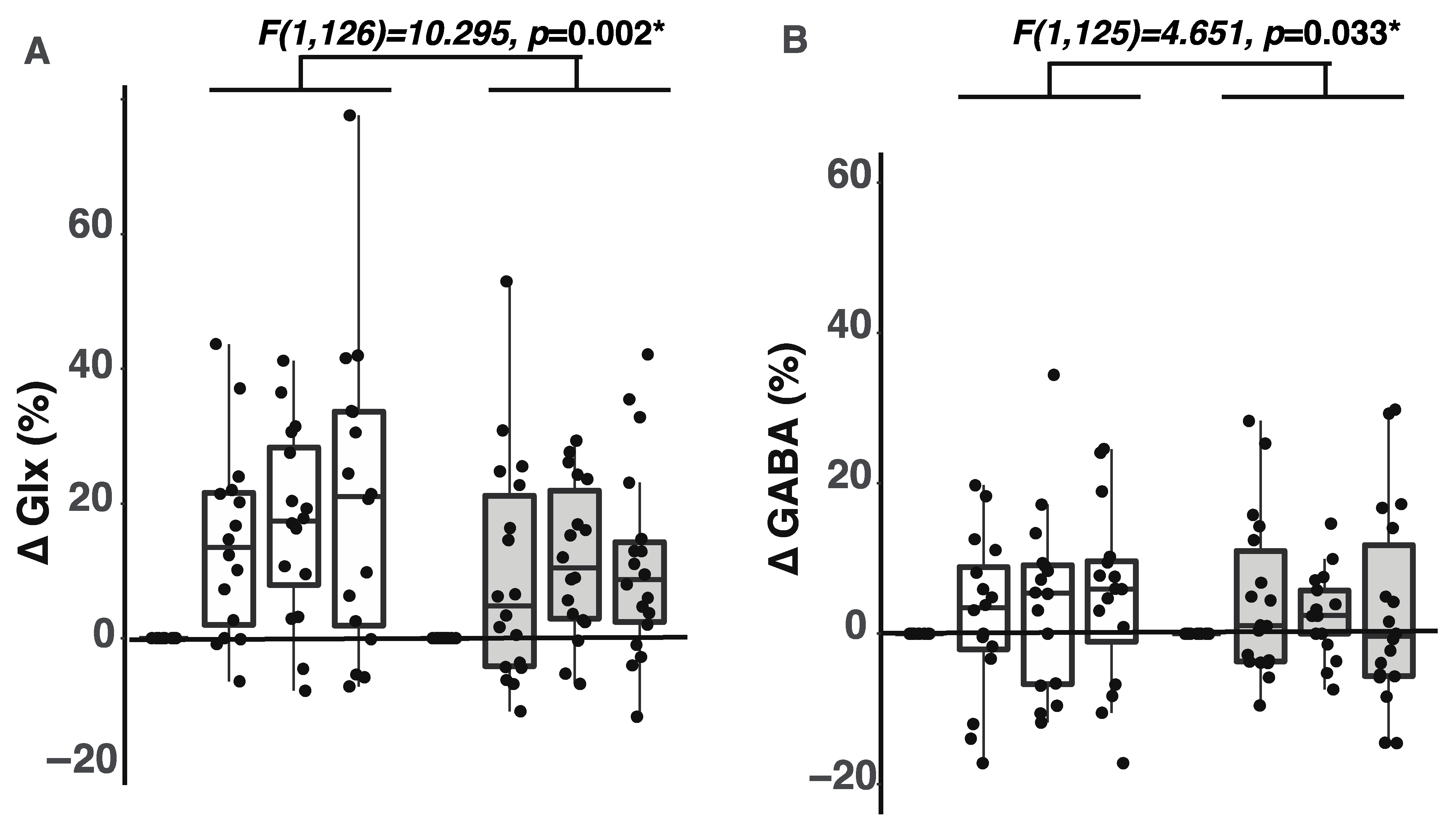
2.1. Participants with ADHD Showed Smaller Glx Increases during the Tasks
2.2. Participants with ADHD Showed Smaller GABA Increases during the Tasks
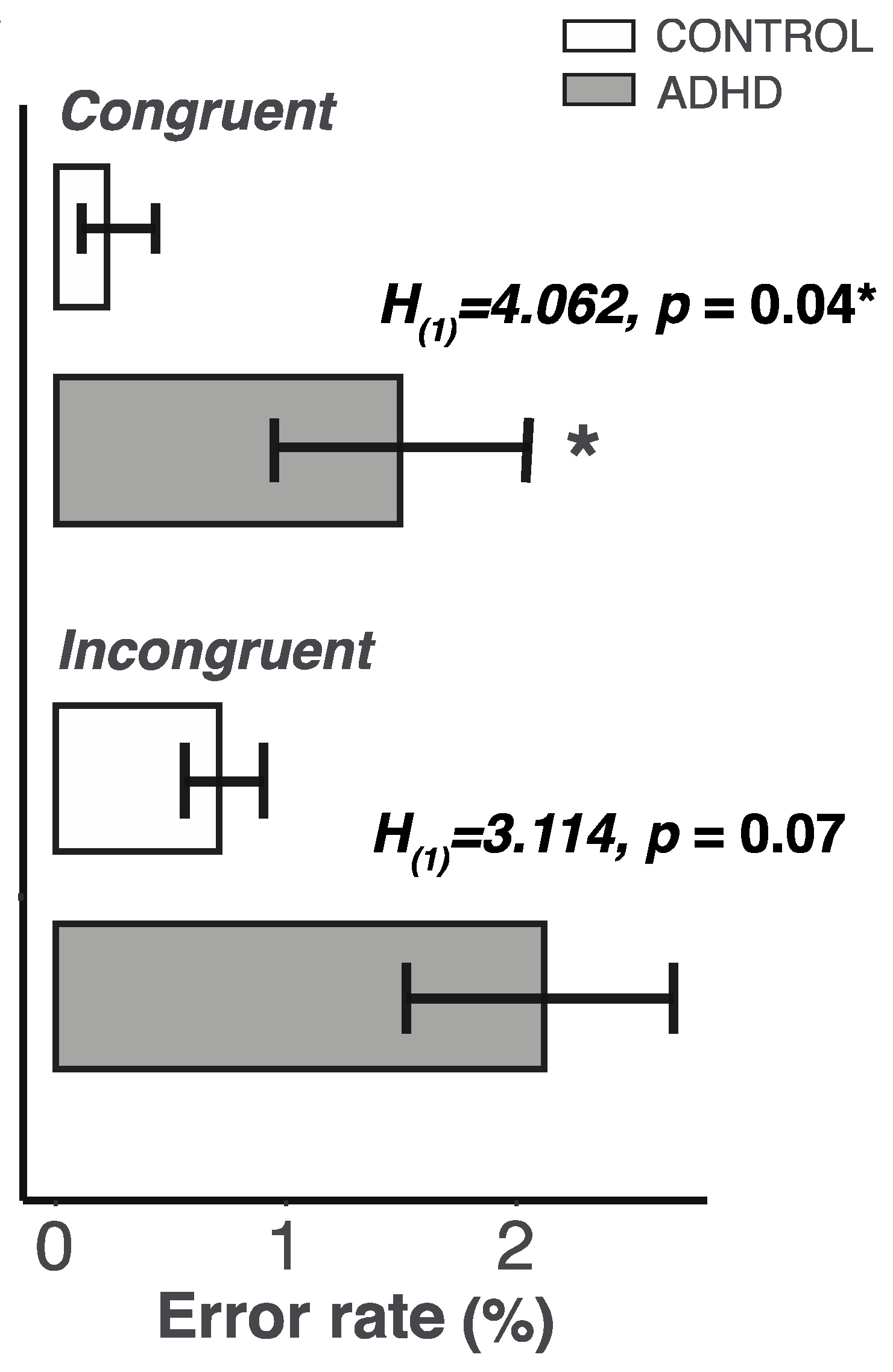
2.3. Lower Control of Attention in the ADHD Group
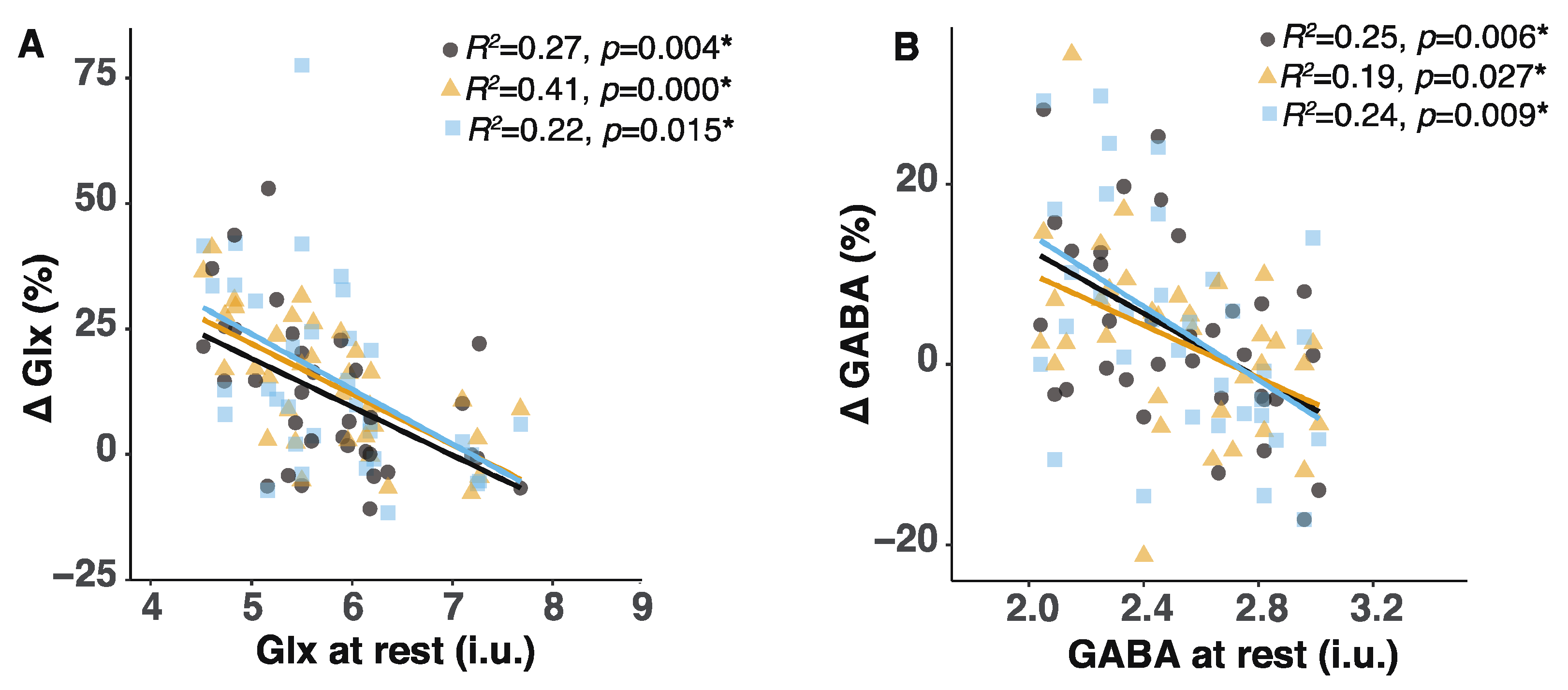
2.4. Lower Glx or GABA Content at Rest Predicted Greater Glx or GABA Increases during the Tasks
2.5. Volumes of Gray-Matter, White-Matter, and Cerebropinal Fluid Were Identical between ADHD and CONTROL Groups
3. Discussion
3.1. Reduced GABA and Glx Content during the Tasks Suggest Altered Glutamate-GABA Cycling in Subjects with ADHD
3.2. The Inverse Relationship between Basal Glx or GABA Content and the Extent of Glx or GABA Increases May Be Related to Reduced GAD Activity
3.3. Reduced GABA Increases during Tasks Contribute to Impaired Attention Control in ADHD
4. Methods and Materials
4.1. Study Participants
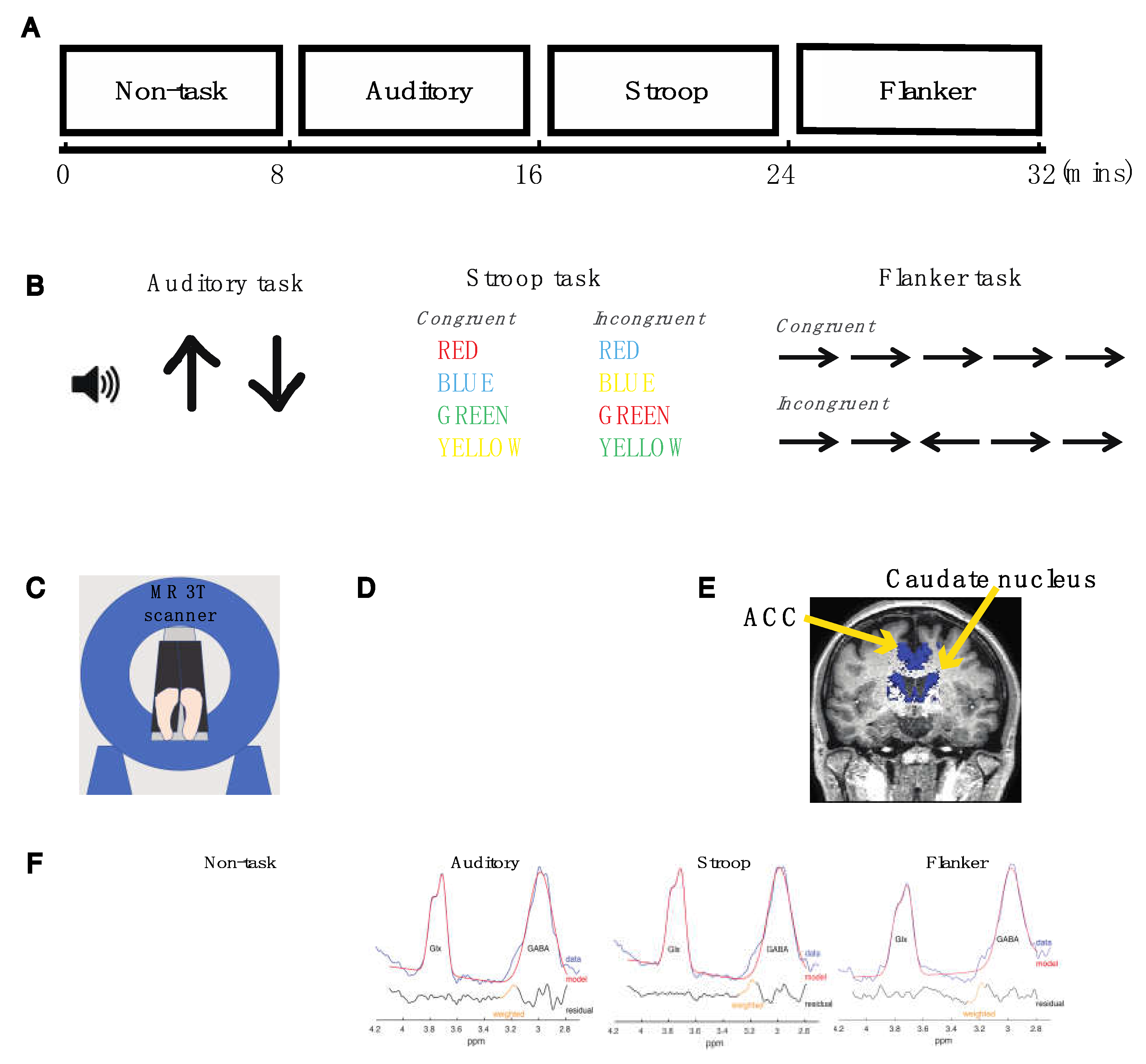
4.2. Experimental Design
4.3. Auditory Task
4.4. Color-Naming Version of the Stroop Task
4.5. Flanker Task
4.6. Magnetic Resonance (MR) Data Acquisition
4.7. MRS Data Analyses
4.8. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Polanczyk, G.V.; Willcutt, E.G.; Salum, G.A.; Kieling, C.; Rohde, L.A. ADHD prevalence estimates across three decades: An updated systematic review and meta-regression analysis. Int. J. Epidemiol. 2014, 43, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Faraone, S.V.; Biederman, J.; Mick, E. The age-dependent decline of attention deficit hyperactivity disorder: A meta-analysis of follow-up studies. Psychol. Med. 2006, 36, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Simon, V.; Czobor, P.; Bálint, S.; Mészáros, Á.; Bitter, I. Prevalence and correlates of adult attention-deficit hyperactivity disorder: Meta-analysis. Br. J. Psychiatry 2009, 194, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Franke, B.; on behalf of the International Multicentre persistent ADHD CollaboraTion (IMpACT); Faraone, S.V.; Asherson, P.; Buitelaar, J.; Bau, C.H.D.; Ramos-Quiroga, J.A.; Mick, E.; Grevet, E.H.; Johansson, S.; et al. The genetics of attention deficit/hyperactivity disorder in adults, a review. Mol. Psychiatry 2012, 17, 960–987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matte, B.; Anselmi, L.; Salum, G.A.; Kieling, C.; Gonçalves, H.; Menezes, A.; Grevet, E.H.; Rohde, L.A. ADHD in DSM-5: A field trial in a large, representative sample of 18- to 19-year-old adults. Psychol. Med. 2015, 45, 361–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polanczyk, G.; Caspi, A.; Houts, R.; Kollins, S.H.; Rohde, L.A.; Moffitt, T.E. Implications of Extending the ADHD Age-of-Onset Criterion to Age 12: Results from a Prospectively Studied Birth Cohort. J. Am. Acad. Child Adolesc. Psychiatry 2010, 49, 210–216. [Google Scholar] [CrossRef]
- Faraone, S.; Biederman, J.; Spencer, T.; Wilens, T.; Seidman, L.J.; Mick, E.; Doyle, A.E. Attention-deficit/hyperactivity disorder in adults: An overview. Biol. Psychiatry 2000, 48, 9–20. [Google Scholar] [CrossRef]
- Feldman, H.M.; Reiff, M.I. Clinical practice. Attention deficit-hyperactivity disorder in children and adolescents. N. Engl. J. Med. 2014, 370, 838–846. [Google Scholar] [CrossRef]
- Demontis, D.; Walters, R.K.; Martin, J.; Mattheisen, M.; Als, T.D.; Agerbo, E.; Baldursson, G.; Belliveau, R.; Bybjerg-Grauholm, J.; Bækvad-Hansen, M.; et al. Discovery of the first genome-wide significant risk loci for attention deficit/hyperactivity disorder. Nat. Genet. 2019, 51, 63–75. [Google Scholar] [CrossRef] [Green Version]
- Barbaresi, W.J.; Colligan, R.C.; Weaver, A.L.; Voigt, R.G.; Killian, J.M.; Katusic, S.K.; Chawla, A.; Sprinz, P.G.; Welch, J.; Heeney, M.; et al. Mortality, ADHD, and Psychosocial Adversity in Adults With Childhood ADHD: A Prospective Study. Pediatrics 2013, 131, 637–644. [Google Scholar] [CrossRef] [Green Version]
- Visser, S.N.; Danielson, M.; Bitsko, R.H.; Holbrook, J.R.; Kogan, M.D.; Ghandour, R.M.; Perou, R.; Blumberg, S.J. Trends in the Parent-Report of Health Care Provider-Diagnosed and Medicated Attention-Deficit/Hyperactivity Disorder: United States, 2003–2011. J. Am. Acad. Child Adolesc. Psychiatry 2014, 53, 34–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbaresi, W.J.; Campbell, L.; Diekroger, E.A.; Froehlich, T.E.; Liu, Y.H.; O’Malley, E.; Pelham, W.E., Jr.; Power, T.J.; Zinner, S.H.; Chan, E. Society for Developmental and Behavioral Pediatrics Clinical Practice Guideline for the Assessment and Treatment of Children and Adolescents with Complex Attention-Deficit/Hyperactivity Disorder. J. Dev. Behav. Pediatr. 2020, 41 (Suppl. 2S), S35–S57. [Google Scholar] [CrossRef]
- Naaijen, J.; The IMAGE consortium; Bralten, J.; Poelmans, G.; Glennon, J.C.; Franke, B.; Buitelaar, J.K. Glutamatergic and GABAergic gene sets in attention-deficit/hyperactivity disorder: Association to overlapping traits in ADHD and autism. Transl. Psychiatry 2017, 7, e999. [Google Scholar] [CrossRef] [PubMed]
- Bollmann, S.; Ghisleni, C.; Poil, S.-S.; Martin, E.L.; Ball, J.S.; Eich-Höchli, D.; Edden, R.A.E.; Klaver, P.; Michels, L.; Brandeis, D.; et al. Developmental changes in gamma-aminobutyric acid levels in attention-deficit/hyperactivity disorder. Transl. Psychiatry 2015, 5, e589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dramsdahl, M.; Ersland, L.; Plessen, K.J.; Haavik, J.; Hugdahl, K.; Specht, K. Adults with Attention-Deficit/Hyperactivity Disorder ? A Brain Magnetic Resonance Spectroscopy Study. Front. Psychiatry 2011, 2, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edden, R.A.E.; Crocetti, D.; Zhu, H.; Gilbert, D.; Mostofsky, S.H. Reduced GABA Concentration in Attention-Deficit/Hyperactivity Disorder. Arch. Gen. Psychiatry 2012, 69, 750–753. [Google Scholar] [CrossRef]
- Ende, G.; Cackowski, S.; Van Eijk, J.; Sack, M.; Demirakca, T.; Kleindienst, N.; Bohus, M.; Sobanski, E.; Krause-Utz, A.; Schmahl, C. Impulsivity and Aggression in Female BPD and ADHD Patients: Association with ACC Glutamate and GABA Concentrations. Neuropsychopharmacology 2016, 41, 410–418. [Google Scholar] [CrossRef] [Green Version]
- MacMaster, F.; Carrey, N.; Sparkes, S.; Kusumakar, V. Proton spectroscopy in medication-free pediatric attention-deficit/hyperactivity disorder. Biol. Psychiatry 2003, 53, 184–187. [Google Scholar] [CrossRef]
- Maltezos, S.; Horder, J.; Coghlan, S.; Skirrow, C.; O’Gorman, R.; Lavender, T.J.; Mendez, M.A.; Mehta, M.; Daly, E.; Xenitidis, K.; et al. Glutamate/glutamine and neuronal integrity in adults with ADHD: A proton MRS study. Transl. Psychiatry 2014, 4, e373. [Google Scholar] [CrossRef]
- Perlov, E.; Philipsen, A.; Hesslinger, B.; Buechert, M.; Ahrendts, J.; Feige, B.; Bubl, E.; Hennig, J.; Ebert, D.; van Elst, L.T. Reduced cingulate glutamate/glutamine-to-creatine ratios in adult patients with attention deficit/hyperactivity disorder—A magnet resonance spectroscopy study. J. Psychiatr. Res. 2007, 41, 934–941. [Google Scholar] [CrossRef]
- Puts, N.A.; Ryan, M.; Oeltzschner, G.; Horska, A.; Edden, R.A.; Mahone, E.M. Reduced striatal GABA in unmedicated children with ADHD at 7T. Psychiatry Res. Neuroimaging 2020, 301, 111082. [Google Scholar] [CrossRef] [PubMed]
- Courvoisie, H.; Hooper, S.R.; Fine, C.; Kwock, L.; Castillo, M. Neurometabolic functioning and neuropsychological correlates in children with ADHD-H: Preliminary findings. J. Neuropsychiatry Clin. Neurosci. 2004, 16, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Tafazoli, S.; O’Neill, J.; Bejjani, A.; Ly, R.; Salamon, N.; McCracken, J.T.; Alger, J.R.; Levitt, J.G. 1H MRSI of middle frontal gyrus in pediatric ADHD. J. Psychiatr. Res. 2013, 47, 505–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gold, B.I.; Roth, R.H. Glutamate decarboxylase activity in striatal slices: Characterization of the increase following depolarization. J. Neurochem. 1979, 32, 883–888. [Google Scholar] [CrossRef] [PubMed]
- Yizhar, O.; Fenno, L.E.; Prigge, M.; Schneider, F.; Davidson, T.; O’Shea, D.J.; Sohal, V.; Goshen, I.; Finkelstein, J.; Paz, J.T.; et al. Neocortical excitation/inhibition balance in information processing and social dysfunction. Nature 2011, 477, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.; Bridi, M.; Koh, M.T.; Gallagher, M.; Kirkwood, A. Reduced cognitive performance in aged rats correlates with increased excitation/inhibition ratio in the dentate gyrus in response to lateral entorhinal input. Neurobiol. Aging 2019, 82, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.; Neufeld, R.W.J.; Schaefer, B.; Densmore, M.; Rajakumar, N.; Osuch, E.A.; Williamson, P.C.; Theberge, J. Functional magnetic resonance spectroscopy of glutamate in schizophrenia and major depressive disorder: Anterior cingulate activity during a color-word Stroop task. NPJ Schizophr. 2015, 1, 15028. [Google Scholar] [CrossRef]
- Taylor, R.; Schaefer, B.; Densmore, M.; Neufeld, R.W.; Rajakumar, N.; Williamson, P.C.; Théberge, J. Increased glutamate levels observed upon functional activation in the anterior cingulate cortex using the Stroop Task and functional spectroscopy. NeuroReport 2015, 26, 107–112. [Google Scholar] [CrossRef] [Green Version]
- Bezalel, V.; Paz, R.; Tal, A. Inhibitory and excitatory mechanisms in the human cingulate-cortex support reinforcement learning: A functional Proton Magnetic Resonance Spectroscopy study. NeuroImage 2019, 184, 25–35. [Google Scholar] [CrossRef]
- Cai, K.; Nanga, R.P.; Lamprou, L.; Schinstine, C.; Elliott, M.A.; Hariharan, H.; Reddy, R.; Epperson, C.N. The Impact of Gabapentin Administration on Brain GABA and Glutamate Concentrations: A 7T 1H-MRS Study. Neuropsychopharmacology 2012, 37, 2764–2771. [Google Scholar] [CrossRef] [Green Version]
- Akhtar, N.D.; Land, P.W. Activity-dependent regulation of glutamic acid decarboxylase in the rat barrel cortex: Effects of neonatal versus adult sensory deprivation. J. Comp. Neurol. 1991, 307, 200–213. [Google Scholar] [CrossRef] [PubMed]
- Benson, D.; Huntsman, M.M.; Jones, E.G. Activity-dependent Changes in GAD and Preprotachykinin mRNAs in Visual Cortex of Adult Monkeys. Cereb. Cortex 1994, 4, 40–51. [Google Scholar] [CrossRef] [PubMed]
- Bowers, G.; Cullinan, W.E.; Herman, J. Region-Specific Regulation of Glutamic Acid Decarboxylase (GAD) mRNA Expression in Central Stress Circuits. J. Neurosci. 1998, 18, 5938–5947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlson, V.C.C.; Mathur, B.; Davis, M.; Lovinger, D.M. Subsets of spiny striosomal striatal neurons revealed in the Gad1-GFP BAC transgenic mouse. Basal Ganglia 2011, 1, 201–211. [Google Scholar] [CrossRef] [Green Version]
- Hendry, S.H.; Jones, E.G. Activity-dependent regulation of GABA expression in the visual cortex of adult monkeys. Neuron 1988, 1, 701–712. [Google Scholar] [CrossRef]
- Li, Y.; Blanco, G.D.; Lei, Z.; Xu, Z.C. Increased GAD expression in the striatum after transient cerebral ischemia. Mol. Cell. Neurosci. 2010, 45, 370–377. [Google Scholar] [CrossRef] [Green Version]
- Charpier, S.; Deniau, J.M. In vivo activity-dependent plasticity at cortico-striatal connections: Evidence for physiological long-term potentiation. Proc. Natl. Acad. Sci. USA 1997, 94, 7036–7040. [Google Scholar] [CrossRef] [Green Version]
- Gierdalski, M.; Jablonska, B.; Siucinska, E.; Lech, M.; Skibinska, A.; Kossut, M. Rapid regulation of GAD67 mRNA and protein level in cortical neurons after sensory learning. Cereb. Cortex 2001, 11, 806–815. [Google Scholar] [CrossRef] [Green Version]
- Besse, A.; Wu, P.; Bruni, F.; Donti, T.; Graham, B.; Craigen, W.J.; McFarland, R.; Moretti, P.; Lalani, S.; Scott, K.L.; et al. The GABA Transaminase, ABAT, Is Essential for Mitochondrial Nucleoside Metabolism. Cell Metab. 2015, 21, 417–427. [Google Scholar] [CrossRef] [Green Version]
- Mahmoud, S.; Gharagozloo, M.; Simard, C.; Gris, D. Astrocytes Maintain Glutamate Homeostasis in the CNS by Controlling the Balance between Glutamate Uptake and Release. Cells 2019, 8, 184. [Google Scholar] [CrossRef] [Green Version]
- Rae, C.D. A Guide to the Metabolic Pathways and Function of Metabolites Observed in Human Brain 1H Magnetic Resonance Spectra. Neurochem. Res. 2014, 39, 1–36. [Google Scholar] [CrossRef] [PubMed]
- Siucinska, E. Gamma-Aminobutyric acid in adult brain: An update. Behav. Brain Res. 2019, 376, 112224. [Google Scholar] [CrossRef] [PubMed]
- Hyder, F.; Fulbright, R.K.; Shulman, R.G.; Rothman, D.L. Glutamatergic Function in the Resting Awake Human Brain is Supported by Uniformly High Oxidative Energy. J. Cereb. Blood Flow Metab. 2013, 33, 339–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruxel, E.M.; Akutagava-Martins, G.C.; Salatino-Oliveira, A.; Genro, J.P.; Zeni, C.P.; Polanczyk, G.V.; Chazan, R.; Schmitz, M.; Rohde, L.A.; Hutz, M.H. GAD1gene polymorphisms are associated with hyperactivity in Attention-Deficit/Hyperactivity Disorder. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2016, 171, 1099–1104. [Google Scholar] [CrossRef]
- Marenco, S.; Savostyanova, A.A.; Van Der Veen, J.W.; Geramita, M.; Stern, A.J.; Barnett, A.S.; Kolachana, B.S.; Radulescu, E.; Zhang, F.; Callicott, J.; et al. Genetic Modulation of GABA Levels in the Anterior Cingulate Cortex by GAD1 and COMT. Neuropsychopharmacology 2010, 35, 1708–1717. [Google Scholar] [CrossRef]
- Sheikh, S.; Martin, S.; Martin, D. Regional distribution and relative amounts of glutamate decarboxylase isoforms in rat and mouse brain. Neurochem. Int. 1999, 35, 73–80. [Google Scholar] [CrossRef]
- Adler, L.A.; Kroon, R.A.; Stein, M.; Shahid, M.; Tarazi, F.I.; Szegedi, A.; Schipper, J.; Cazorla, P. A Translational Approach to Evaluate the Efficacy and Safety of the Novel AMPA Receptor Positive Allosteric Modulator Org 26576 in Adult Attention-Deficit/Hyperactivity Disorder. Biol. Psychiatry 2012, 72, 971–977. [Google Scholar] [CrossRef]
- Cheng, J.; Liu, A.; Shi, M.Y.; Yan, Z. Disrupted Glutamatergic Transmission in Prefrontal Cortex Contributes to Behavioral Abnormality in an Animal Model of ADHD. Neuropsychopharmacology 2017, 42, 2096–2104. [Google Scholar] [CrossRef]
- Morello, F.; Voikar, V.; Parkkinen, P.; Panhelainen, A.; Rosenholm, M.; Makkonen, A.; Rantamäki, T.; Piepponen, P.; Aitta-Aho, T.; Partanen, J. ADHD-like behaviors caused by inactivation of a transcription factor controlling the balance of inhibitory and excitatory neuron development in the mouse anterior brainstem. Transl. Psychiatry 2020, 10, 357. [Google Scholar] [CrossRef]
- Chen, L.; Yang, X.; Zhou, X.; Wang, C.; Gong, X.; Chen, B.; Chen, Y. Hyperactivity and impaired attention in Gamma aminobutyric acid transporter subtype 1 gene knockout mice. Acta Neuropsychiatr. 2015, 27, 368–374. [Google Scholar] [CrossRef]
- Yuan, F.-F.; Gu, X.; Huang, X.; Zhong, Y.; Wu, J. SLC6A1 gene involvement in susceptibility to attention-deficit/hyperactivity disorder: A case-control study and gene-environment interaction. Prog. Neuro-Psychopharmacology Biol. Psychiatry 2017, 77, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.; Atallah, B.; Scanziani, M. Equalizing excitation–inhibition ratios across visual cortical neurons. Nature 2014, 511, 596–600. [Google Scholar] [CrossRef]
- Petroff, O.; Rothman, D.L.; Behar, K.L.; Collins, T.L.; Mattson, R.H. Human brain GABA levels rise rapidly after initiation of vigabatrin therapy. Neurology 1996, 47, 1567–1571. [Google Scholar] [CrossRef]
- Kurcyus, K.; Annac, E.; Hanning, N.M.; Harris, A.D.; Oeltzschner, G.; Edden, R.; Riedl, V. Opposite Dynamics of GABA and Glutamate Levels in the Occipital Cortex during Visual Processing. J. Neurosci. 2018, 38, 9967–9976. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.H.; Grandelis, A.; Maddock, R.J. Dorsolateral Prefrontal Cortex GABA Concentration in Humans Predicts Working Memory Load Processing Capacity. J. Neurosci. 2016, 36, 11788–11794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ajram, L.A.; Horder, J.; Mendez, M.A.; Galanopoulos, A.; Brennan, L.P.; Wichers, R.H.; Robertson, D.M.; Murphy, C.M.; Zinkstok, J.; Ivin, G.; et al. Shifting brain inhibitory balance and connectivity of the prefrontal cortex of adults with autism spectrum disorder. Transl. Psychiatry 2017, 7, e1137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrmann, M.J.; Mader, K.; Schreppel, T.; Jacob, C.; Heine, M.; Boreatti-Hümmer, A.; Ehlis, A.-C.; Scheuerpflug, P.; Pauli, P.; Fallgatter, A.J. Neural correlates of performance monitoring in adult patients with attention deficit hyperactivity disorder (ADHD). World J. Biol. Psychiatry 2010, 11, 457–464. [Google Scholar] [CrossRef]
- Johnson, K.A.; Robertson, I.H.; Barry, E.; Mulligan, A.; Dáibhis, A.; Daly, M.; Watchorn, A.; Gill, M.; Bellgrove, M. Impaired conflict resolution and alerting in children with ADHD: Evidence from the Attention Network Task (ANT). J. Child Psychol. Psychiatry 2008, 49, 1339–1347. [Google Scholar] [CrossRef]
- King, J.A.; Colla, M.; Brass, M.; Heuser, I.; Von Cramon, D. Inefficient cognitive control in adult ADHD: Evidence from trial-by-trial Stroop test and cued task switching performance. Behav. Brain Funct. 2007, 3, 42. [Google Scholar] [CrossRef] [Green Version]
- Mullane, J.C.; Corkum, P.V.; Klein, R.M.; McLaughlin, E.N.; Lawrence, M.A. Alerting, Orienting, and Executive Attention in Children With ADHD. J. Atten. Disord. 2011, 15, 310–320. [Google Scholar] [CrossRef]
- Schoenfelder, E.N.; Chronis-Tuscano, A.; Strickland, J.; Almirall, D.; Stein, M.A. Piloting a Sequential, Multiple Assignment, Randomized Trial for Mothers with Attention-Deficit/Hyperactivity Disorder and Their At-Risk Young Children. J. Child Adolesc. Psychopharmacol. 2019, 29, 256–267. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, S.A.; Larson, E.; Lee, A.K.C. Neural Switch Asymmetry in Feature-Based Auditory Attention Tasks. J. Assoc. Res. Otolaryngol. 2019, 20, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Boersma, P.; Weenik, D. Praat: Doing Phonetics by Computer (Version 5.1. 03) [Computer program]. Retrieved 21 March 2009. Available online: https://www.fon.hum.uva.nl/praat/ (accessed on 7 March 2022).
- Mamiya, P.C.; Richards, T.L.; Kuhl, P.K. Right Forceps Minor and Anterior Thalamic Radiation Predict Executive Function Skills in Young Bilingual Adults. Front. Psychol. 2018, 9, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mamiya, P.C.; Richards, T.; Corrigan, N.M.; Kuhl, P.K. Strength of Ventral Tegmental Area Connections With Left Caudate Nucleus Is Related to Conflict Monitoring. Front. Psychol. 2019, 10, 2869. [Google Scholar] [CrossRef] [Green Version]
- Edden, R.A.; Barker, P.B. Spatial effects in the detection of gamma-aminobutyric acid: Improved sensitivity at high fields using inner volume saturation. Magn. Reson. Med. 2007, 58, 1276–1282. [Google Scholar] [CrossRef]
- Mescher, M.; Merkle, H.; Kirsch, J.; Garwood, M.; Gruetter, R. Simultaneous in vivo spectral editing and water suppression. NMR BioMed 1998, 11, 266–272. [Google Scholar] [CrossRef] [Green Version]
- Tkáč, I.; Starčuk, Z.; Choi, I.; Gruetter, R. In vivo 1H NMR spectroscopy of rat brain at 1 ms echo time. Magn. Reson. Med. 1999, 41, 649–656. [Google Scholar] [CrossRef] [Green Version]
- Edden, R.A.; Puts, N.A.; Harris, A.D.; Barker, P.B.; Evans, C.J. Gannet: A batch-processing tool for the quantitative analysis of gamma-aminobutyric acid-edited MR spectroscopy spectra. J. Magn. Reson. Imaging 2014, 40, 1445–1452. [Google Scholar] [CrossRef]
- Harris, A.D.; Puts, N.A.J.; Edden, R.A.E. Tissue correction for GABA-edited MRS: Considerations of voxel composition, tissue segmentation, and tissue relaxations. J. Magn. Reson. Imaging 2015, 42, 1431–1440. [Google Scholar] [CrossRef] [Green Version]
- Ashburner, J.; Friston, K. Unified segmentation. NeuroImage 2005, 26, 839–851. [Google Scholar] [CrossRef]
- Provencher, S.W. Estimation of metabolite concentrations from localized in vivo proton NMR spectra. Magn. Reson. Med. 1993, 30, 672–679. [Google Scholar] [CrossRef] [PubMed]
- Rowland, L.M.; Krause, B.W.; Wijtenburg, S.A.; McMahon, R.P.; Chiappelli, J.; Nugent, K.L.; Nisonger, S.J.; Korenic, S.A.; Kochunov, P.; Hong, L.E. Medial frontal GABA is lower in older schizophrenia: A MEGA-PRESS with macromolecule suppression study. Mol. Psychiatry 2016, 21, 198–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, F.; Edden, R.A.; Li, M.; Puts, N.A.; Wang, G.; Liu, C.; Zhao, B.; Wang, H.; Bai, X.; Zhao, C.; et al. Edited magnetic resonance spectroscopy detects an age-related decline in brain GABA levels. NeuroImage 2013, 78, 75–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mamiya, P.C.; Richards, T.L.; Edden, R.A.E.; Lee, A.K.C.; Stein, M.A.; Kuhl, P.K. Reduced Glx and GABA Inductions in the Anterior Cingulate Cortex and Caudate Nucleus Are Related to Impaired Control of Attention in Attention-Deficit/Hyperactivity Disorder. Int. J. Mol. Sci. 2022, 23, 4677. https://doi.org/10.3390/ijms23094677
Mamiya PC, Richards TL, Edden RAE, Lee AKC, Stein MA, Kuhl PK. Reduced Glx and GABA Inductions in the Anterior Cingulate Cortex and Caudate Nucleus Are Related to Impaired Control of Attention in Attention-Deficit/Hyperactivity Disorder. International Journal of Molecular Sciences. 2022; 23(9):4677. https://doi.org/10.3390/ijms23094677
Chicago/Turabian StyleMamiya, Ping C., Todd L. Richards, Richard A. E. Edden, Adrian K. C. Lee, Mark A. Stein, and Patricia K. Kuhl. 2022. "Reduced Glx and GABA Inductions in the Anterior Cingulate Cortex and Caudate Nucleus Are Related to Impaired Control of Attention in Attention-Deficit/Hyperactivity Disorder" International Journal of Molecular Sciences 23, no. 9: 4677. https://doi.org/10.3390/ijms23094677