
Overexpression of Neuregulin-1 Type III Has Impact on Visual Function in Mice


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Visual Electrophysiology
2.3. OCT
2.4. Immunohistochemistry
2.5. Quantitative Analysis of Fluorescence in the Images of Histological Staining
2.6. Data Analysis
3. Results
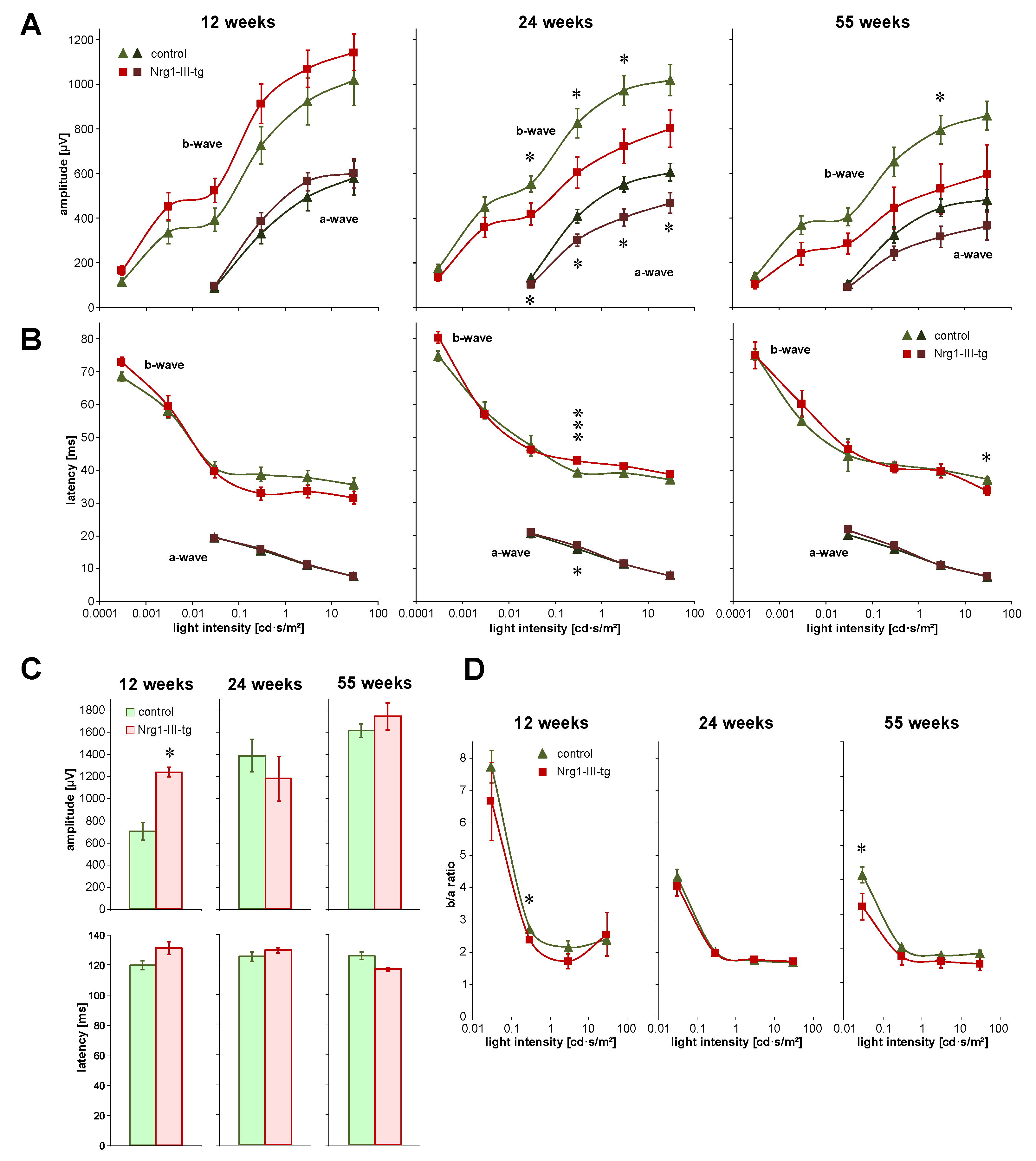
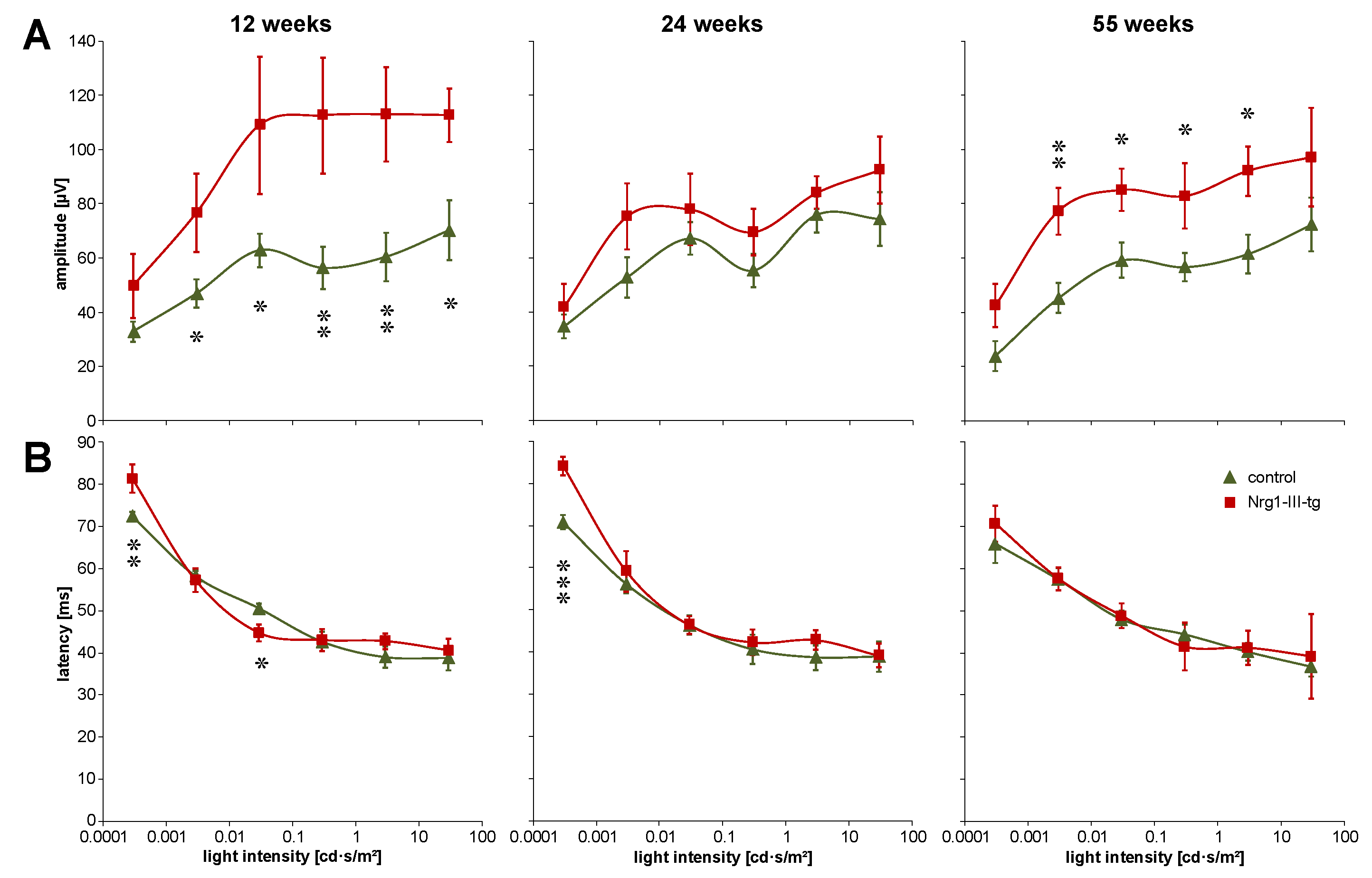
3.1. Investigation of the Function of the Visual System Measured by ERG and VEP
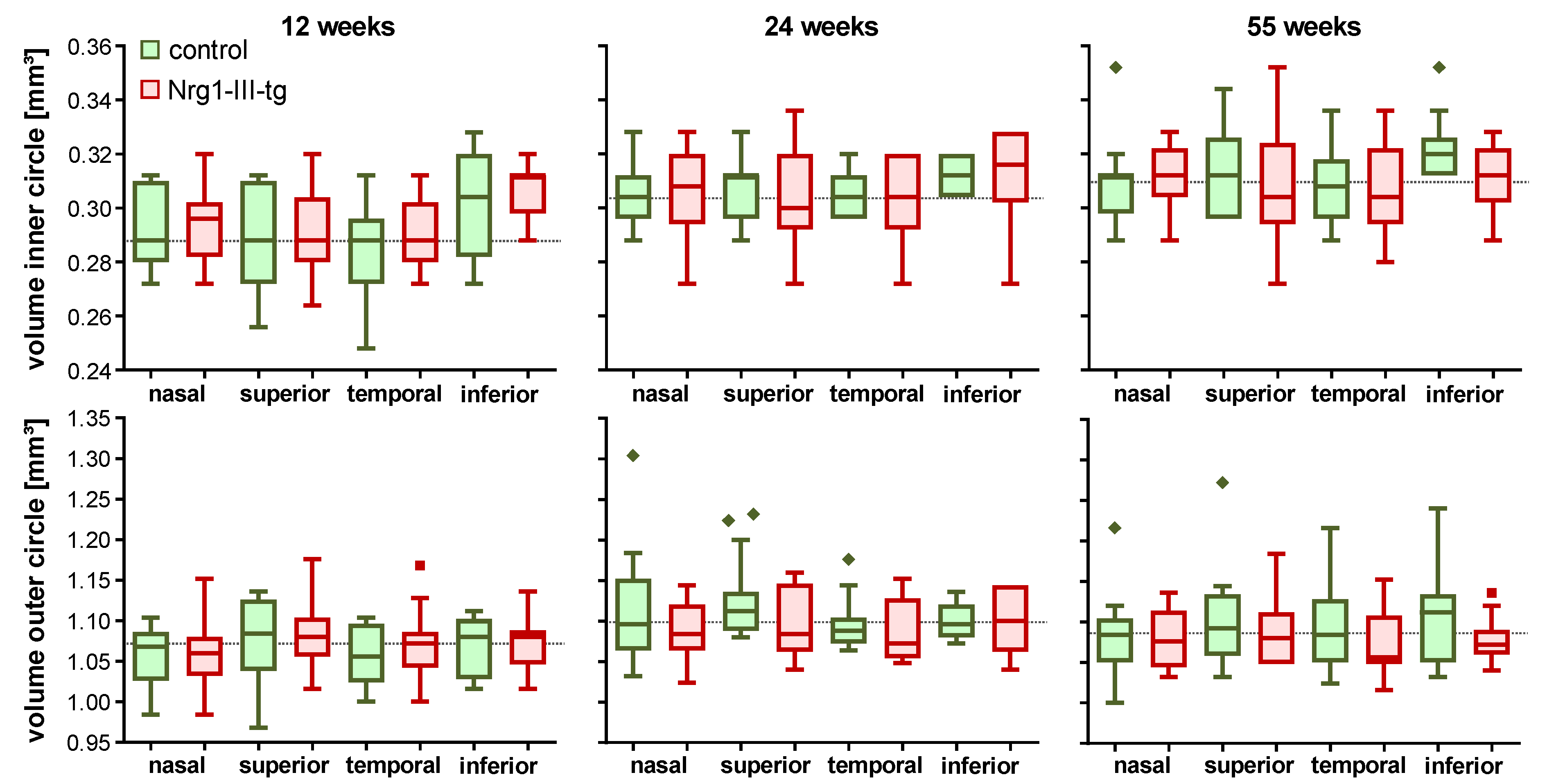
3.2. Investigation of the Thickness and Volume of the Mouse Retina Measured by OCT
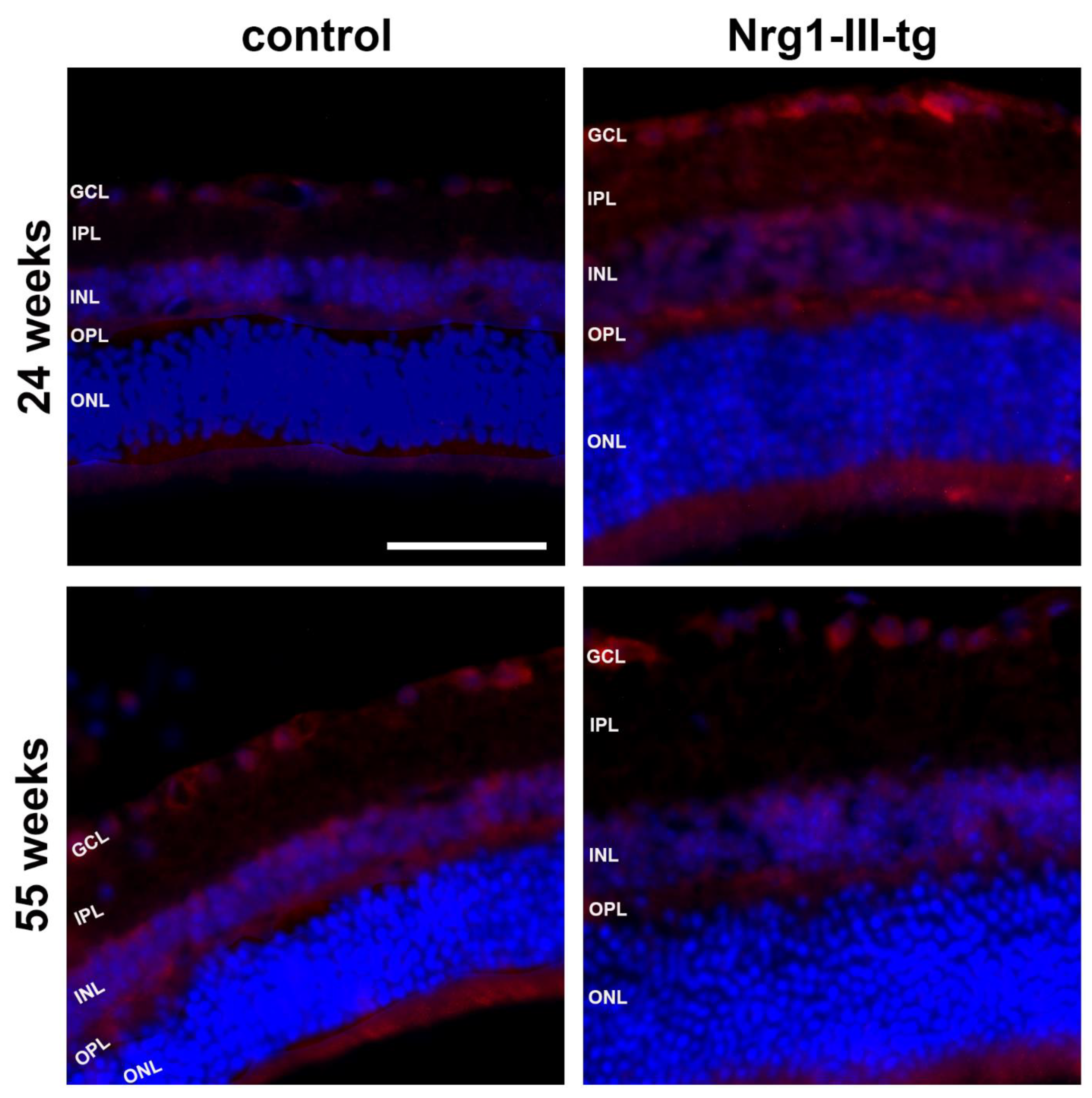
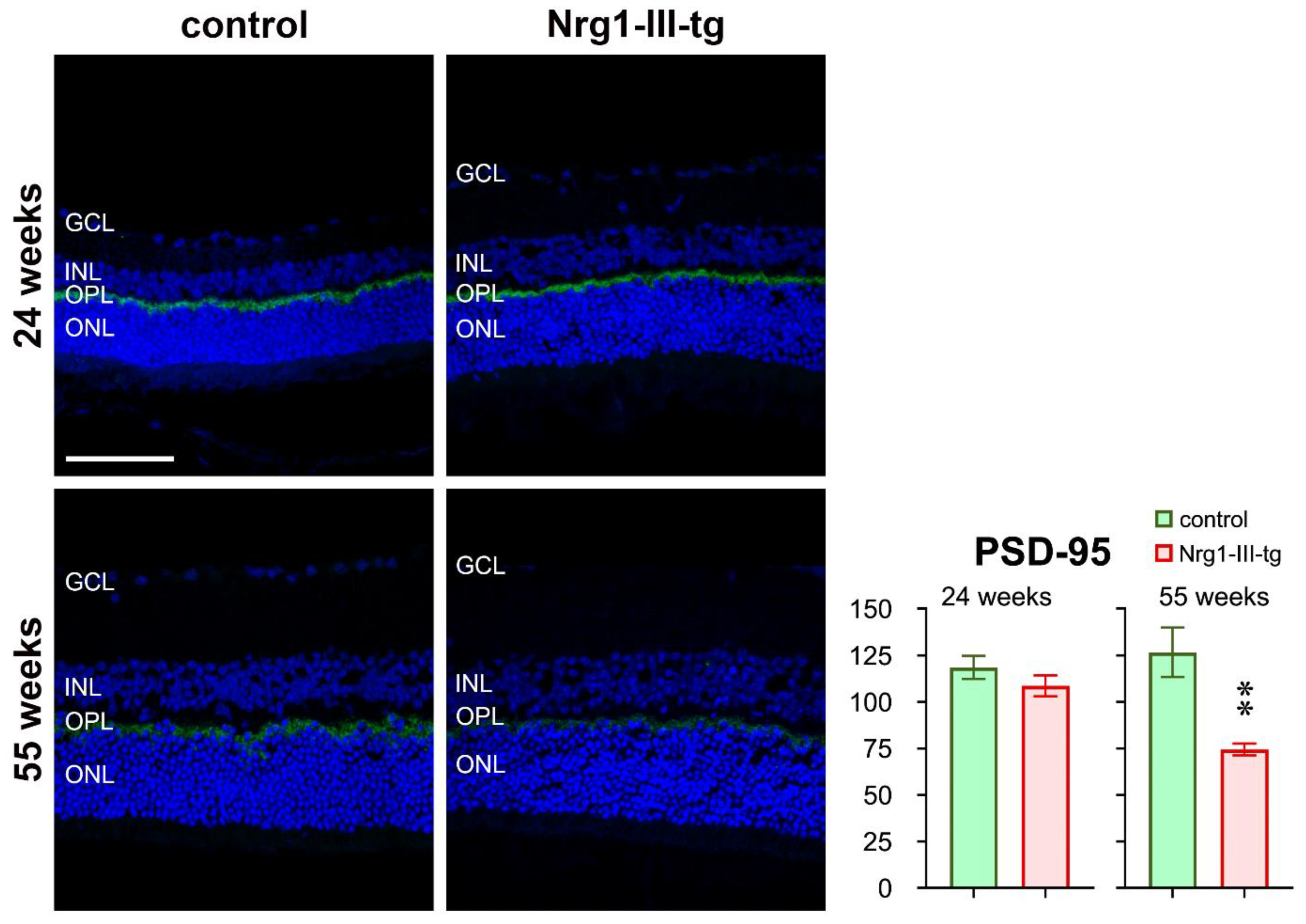
3.3. Semi-Quantitative Analysis of Histological Staining
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harrison, P.J.; Law, A.J. Neuregulin 1 and Schizophrenia: Genetics, Gene Expression, and Neurobiology. Biol. Psychiatry 2006, 60, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Falls, D.L. Neuregulins: Functions, forms, and signaling strategies. Exp. Cell Res. 2003, 284, 14–30. [Google Scholar] [CrossRef]
- Pan, P.; Dobrowsky, R.T. Differential expression of neuregulin-1 isoforms and downregulation of erbin are associated with Erb B2 receptor activation in diabetic peripheral neuropathy. Acta Neuropathol. Commun. 2014, 1, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Law, A.J.; Kleinman, J.E.; Weinberger, D.R.; Weickert, C.S. Disease-associated intronic variants in the ErbB4 gene are related to altered ErbB4 splice-variant expression in the brain in schizophrenia. Hum. Mol. Genet. 2007, 16, 129–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, D.A.; Sweet, R.A. Schizophrenia from a neural circuitry perspective: Advancing toward rational pharmacological therapies. J. Clin. Investig. 2009, 119, 706–716. [Google Scholar] [CrossRef] [PubMed]
- Chong, V.Z.; Thompson, M.; Beltaifa, S.; Webster, M.J.; Law, A.J.; Weickert, C.S. Elevated neuregulin-1 and ErbB4 protein in the prefrontal cortex of schizophrenic patients. Schizophr. Res. 2008, 100, 270–280. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, A.; Zhang, M.; Trembak-Duff, I.; Unterbarnscheidt, T.; Radyushkin, K.; Dibaj, P.; Martins-De-Souza, D.; Boretius, S.; Brzózka, M.M.; Steffens, H.; et al. Dysregulated Expression of Neuregulin-1 by Cortical Pyramidal Neurons Disrupts Synaptic Plasticity. Cell Rep. 2014, 8, 1130–1145. [Google Scholar] [CrossRef] [Green Version]
- Olaya, J.C.; Heusner, C.L.; Matsumoto, M.; Sinclair, D.; Kondo, M.; Karl, T.; Weickert, C.S. Overexpression of Neuregulin 1 Type III Confers Hippocampal mRNA Alterations and Schizophrenia-Like Behaviors in Mice. Schizophr. Bull. 2018, 44, 865–875. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Ikrar, T.; Davis, M.F.; Gong, N.; Zheng, X.; Luo, Z.D.; Lai, C.; Mei, L.; Holmes, T.; Gandhi, S.P.; et al. Neuregulin-1/ErbB4 Signaling Regulates Visual Cortical Plasticity. Neuron 2016, 92, 160–173. [Google Scholar] [CrossRef]
- London, A.; Benhar, I.; Schwartz, M. The retina as a window to the brain—From eye research to CNS disorders. Nat. Rev. Neurol. 2013, 9, 44–53. [Google Scholar] [CrossRef]
- Silverstein, S.M.; Rosen, R. Schizophrenia and the eye. Schizophr. Res. Cogn. 2015, 2, 46–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granö, N.; Salmijärvi, L.; Karjalainen, M.; Kallionpää, S.; Roine, M.; Taylor, P. Early signs of worry: Psychosis risk symptom visual distortions are independently associated with suicidal ideation. Psychiatry Res. 2015, 225, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Gordon-Lipkin, E.; Chodkowski, B.; Reich, D.S.; Smith, S.A.; Pulicken, M.; Balcer, L.J.; Frohman, E.M.; Cutter, G.; Calabresi, P.A. Retinal nerve fiber layer is associated with brain atrophy in multiple sclerosis. Neurology 2007, 69, 1603–1609. [Google Scholar] [CrossRef] [PubMed]
- Ong, Y.-T.; Hilal, S.; Cheung, C.Y.; Venketasubramanian, N.; Niessen, W.J.; Vrooman, H.; Anuar, A.R.; Chew, M.; Chen, C.; Wong, T.Y.; et al. Retinal neurodegeneration on optical coherence tomography and cerebral atrophy. Neurosci. Lett. 2015, 584, 12–16. [Google Scholar] [CrossRef]
- Ferrari, L.; Huang, S.-C.; Magnani, G.; Ambrosi, A.; Comi, G.; Leocani, L. Optical Coherence Tomography Reveals Retinal Neuroaxonal Thinning in Frontotemporal Dementia as in Alzheimer’s Disease. J. Alzheimer’s Dis. 2017, 56, 1101–1107. [Google Scholar] [CrossRef]
- Hébert, M.; Mérette, C.; Gagné, A.-M.; Paccalet, T.; Moreau, I.; Lavoie, J.; Maziade, M. The Electroretinogram May Differentiate Schizophrenia From Bipolar Disorder. Biol. Psychiatry 2020, 87, 263–270. [Google Scholar] [CrossRef]
- Schönfeldt-Lecuona, C.; Kregel, T.; Schmidt, A.; Kassubek, J.; Dreyhaupt, J.; Freudenmann, R.W.; Connemann, B.J.; Gahr, M.; Pinkhardt, E.H. Retinal single-layer analysis with optical coherence tomography (OCT) in schizophrenia spectrum disorder. Schizophr. Res. 2020, 219, 5–12. [Google Scholar] [CrossRef]
- Velanac, V.; Unterbarnscheidt, T.; Hinrichs, W.; Gummert, M.N.; Fischer, T.M.; Rossner, M.; Trimarco, A.; Brivio, V.; Taveggia, C.; Willem, M.; et al. Bace1 processing of NRG1 type III produces a myelin-inducing signal but is not essential for the stimulation of myelination. Glia 2012, 60, 203–217. [Google Scholar] [CrossRef] [Green Version]
- Schubert, T.; Gleiser, C.; Heiduschka, P.; Franz, C.; Nagel-Wolfrum, K.; Sahaboglu, A.; Weisschuh, N.; Eske, G.; Rohbock, K.; Rieger, N.; et al. Deletion of myosin VI causes slow retinal optic neuropathy and age-related macular degeneration (AMD)-relevant retinal phenotype. Cell. Mol. Life Sci. 2015, 72, 3953–3969. [Google Scholar] [CrossRef] [Green Version]
- Stefansson, H.; Steinthorsdottir, V.; Thorgeirsson, T.; Gulcher, J.; Stefansson, K. Neuregulin 1 and schizophrenia. Ann. Med. 2004, 36, 62–71. [Google Scholar] [CrossRef]
- Weickert, C.S.; Tiwari, Y.; Schofield, P.; Mowry, B.J.; Fullerton, J.M. Schizophrenia-associated HapICE haplotype is associated with increased NRG1 type III expression and high nucleotide diversity. Transl. Psychiatry 2012, 2, e104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samani, N.N.; Proudlock, F.A.; Siram, V.; Suraweera, C.; Hutchinson, C.; Nelson, C.P.; Al-Uzri, M.; Gottlob, I. Retinal Layer Abnormalities as Biomarkers of Schizophrenia. Schizophr. Bull. 2018, 44, 876–885. [Google Scholar] [CrossRef] [PubMed]
- Adams, S.A.; Nasrallah, H.A. Multiple retinal anomalies in schizophrenia. Schizophr. Res. 2018, 195, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Pinto, L.H.; Invergo, B.; Shimomura, K.; Takahashi, J.S.; Troy, J.B. Interpretation of the mouse electroretinogram. Doc. Ophthalmol. 2007, 115, 127–136. [Google Scholar] [CrossRef] [Green Version]
- Penn, R.D.; Hagins, W.A. Signal Transmission along Retinal Rods and the Origin of the Electroretinographic a-Wave. Nature 1969, 223, 201–205. [Google Scholar] [CrossRef]
- Robson, J.; Maeda, H.; Saszik, S.; Frishman, L. In vivo studies of signaling in rod pathways of the mouse using the electroretinogram. Vis. Res. 2004, 44, 3253–3268. [Google Scholar] [CrossRef] [Green Version]
- Flames, N.; Long, J.E.; Garratt, A.; Fischer, T.M.; Gassmann, M.; Birchmeier, C.; Lai, C.; Rubenstein, J.L.; Marin, O. Short- and Long-Range Attraction of Cortical GABAergic Interneurons by Neuregulin-1. Neuron 2004, 44, 251–261. [Google Scholar] [CrossRef]
- Ting, A.K.; Chen, Y.; Wen, L.; Yin, N.-M.; Shen, C.; Tao, Y.; Liu, X.; Xiong, W.-C.; Mei, L. Neuregulin 1 Promotes Excitatory Synapse Development and Function in GABAergic Interneurons. J. Neurosci. 2011, 31, 15–25. [Google Scholar] [CrossRef] [Green Version]
- Soghomonian, J.-J.; Martin, D.L. Two isoforms of glutamate decarboxylase: Why? Trends Pharmacol. Sci. 1998, 19, 500–505. [Google Scholar] [CrossRef]
- Hensch, T.K.; Fagiolini, M.; Mataga, N.; Stryker, M.P.; Baekkeskov, S.; Kash, S.F. Local GABA Circuit Control of Experience-Dependent Plasticity in Developing Visual Cortex. Science 1998, 282, 1504–1508. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Kaplan, I.V.; Cooper, N.G.; Mower, G.D. Expression of two forms of glutamic acid decarboxylase (GAD67 and GAD65) during postnatal development of cat visual cortex. Dev. Brain Res. 1997, 103, 127–141. [Google Scholar] [CrossRef]
- Christgau, S.; Aanstoot, H.J.; Schierbeck, H.; Begley, K.; Tullin, S.; Hejnaes, K.; Baekkeskov, S. Membrane anchoring of the autoantigen GAD65 to microvesicles in pancreatic beta-cells by palmitoylation in the NH2-terminal domain. J. Cell Biol. 1992, 118, 309–320. [Google Scholar] [CrossRef] [PubMed]
- Rutecki, P.A. Neuronal excitability: Voltage-dependent currents and synaptic transmission. J. Clin. Neurophysiol. 1992, 9, 195–211. [Google Scholar] [CrossRef] [PubMed]
- Lanore, F.; Silver, R.A. Extracting Quantal Properties of Transmission at Central Synapses. Neuromethods 2016, 113, 193–211. [Google Scholar] [CrossRef]
- Dodson, P.; Forsythe, I. Presynaptic K+ channels: Electrifying regulators of synaptic terminal excitability. Trends Neurosci. 2004, 27, 210–217. [Google Scholar] [CrossRef]
- Ikematsu, N.; Dallas, M.L.; Ross, F.A.; Lewis, R.W.; Rafferty, J.N.; David, J.A.; Suman, R.; Peers, C.; Hardie, D.G.; Evans, A.M. Phosphorylation of the voltage-gated potassium channel Kv2.1 by AMP-activated protein kinase regulates membrane excitability. Proc. Natl. Acad. Sci. USA 2011, 108, 18132–18137. [Google Scholar] [CrossRef] [Green Version]
- Asi, H.; Perlman, I. Relationships between the electroretinogram a-wave, b-wave and oscillatory potentials and their application to clinical diagnosis. Doc. Ophthalmol. 1992, 79, 125–139. [Google Scholar] [CrossRef]
- Ridder, W.; Nusinowitz, S. The visual evoked potential in the mouse—Origins and response characteristics. Vis. Res. 2006, 46, 902–913. [Google Scholar] [CrossRef] [Green Version]
- Barnstable, C.; Dräger, U.C. Thy-1 antigen: A ganglion cell specific marker in rodent retina. Neuroscience 1984, 11, 847–855. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary Antibodies | |||||
|---|---|---|---|---|---|
| Specificity | Host | Supplier | Catalogue No. | Dilution | |
| Kv-2.1 | Rabbit | Synaptic Systems | 231 002 | 1:500 | |
| GAD 2/GAD 65 | Guinea pig | Synaptic Systems | 198104 | 1:500 | |
| Neuregulin-1 | Rabbit | GeneTex | GTX133355 | 1:1000 | |
| PSD-95 | Rat | Santa Cruz Biotechnology | sc-32290 | 1:300 | |
| Secondary Antibodies | |||||
| Specificity | Host | dye | Supplier | Catalogue No. | Dilution |
| Anti-Rabbit | goat | Texas Red | Abcam | ab150080 | 1:800 |
| Anti-Guinea pig | goat | Alexa Fluor 488 | Abcam | ab150185 | 1:600 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, N.; Zhang, W.; Eter, N.; Heiduschka, P.; Zhang, M. Overexpression of Neuregulin-1 Type III Has Impact on Visual Function in Mice. Int. J. Mol. Sci. 2022, 23, 4489. https://doi.org/10.3390/ijms23094489
Su N, Zhang W, Eter N, Heiduschka P, Zhang M. Overexpression of Neuregulin-1 Type III Has Impact on Visual Function in Mice. International Journal of Molecular Sciences. 2022; 23(9):4489. https://doi.org/10.3390/ijms23094489
Chicago/Turabian StyleSu, Nan, Weiqi Zhang, Nicole Eter, Peter Heiduschka, and Mingyue Zhang. 2022. "Overexpression of Neuregulin-1 Type III Has Impact on Visual Function in Mice" International Journal of Molecular Sciences 23, no. 9: 4489. https://doi.org/10.3390/ijms23094489