
Charged Residue Implantation Improves the Affinity of a Cross-Reactive Dengue Virus Antibody


Abstract
:1. Introduction
2. Results
2.1. Structural Analysis
2.2. Mutation Design to Improve Affinity
2.3. MD Simulation Validates the Feasibility of the Designs
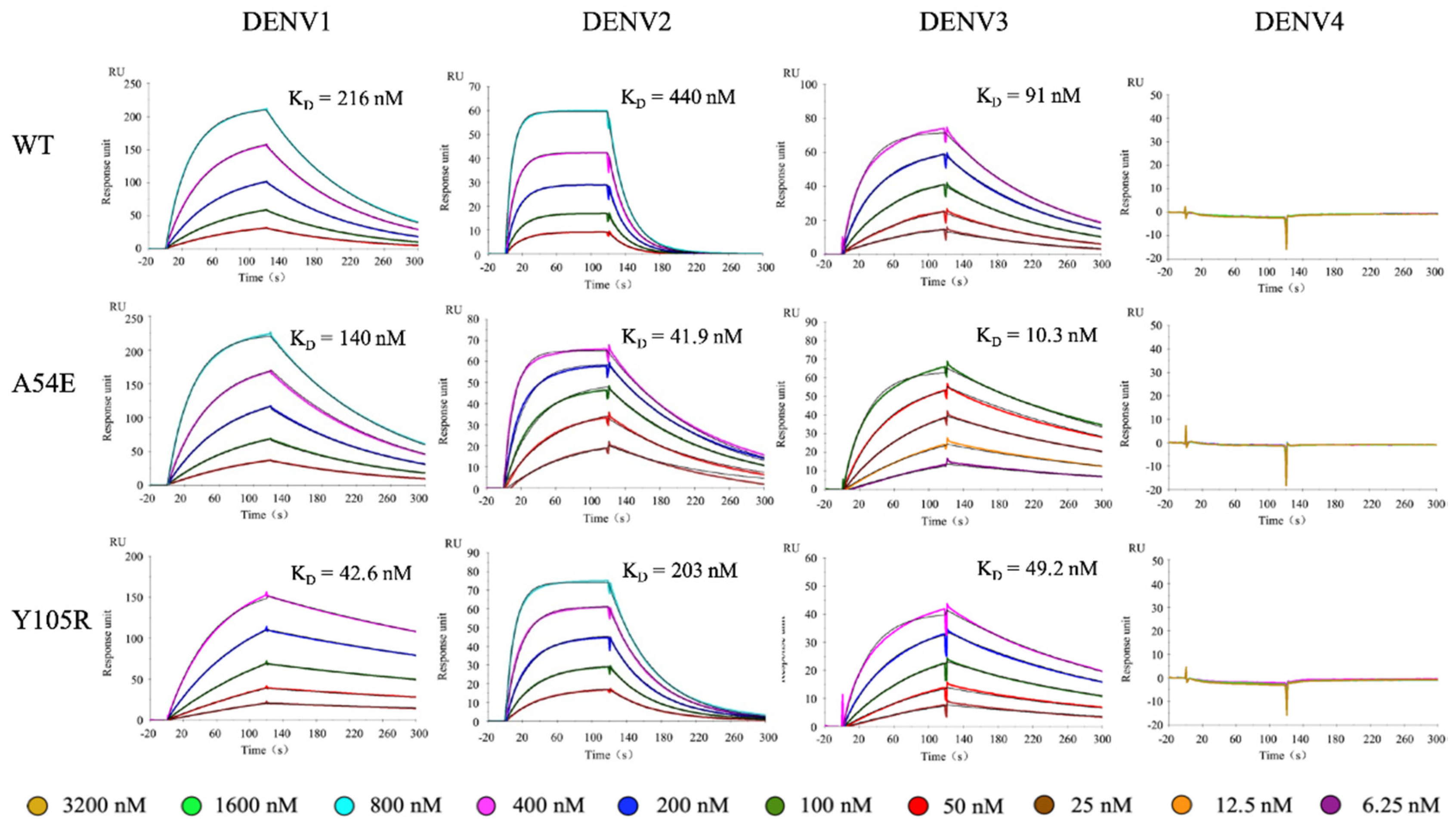
2.4. Binding Kinetics
2.5. Double Mutant Additively Improves 1A1D-2 Affinity
2.6. Molecular Mechanism of Affinity Improvement to DENV1 and DENV3
2.6.1. Affinity Improvement of A54E.H to DENV1 and 3
2.6.2. Affinity Improvement of Y105R.H to DENV1 and 3
3. Discussion
4. Materials and Methods
4.1. Cloning, Expression, and Purification of Four Recombinant EDIIIs
4.2. Cloning and Preparation of 1A1D-2 Fragment Antigen Binding (Fab)
4.3. Binding Kinetics by Surface Plasmon Resonance (SPR)
4.4. MD Simulation
4.5. In Silico Analysis of Binding Mechanisms
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Messina, J.P.; Brady, O.J.; Golding, N.; Kraemer, M.U.G.; Wint, G.R.W.; Ray, S.E.; Pigott, D.M.; Shearer, F.M.; Johnson, K.; Earl, L.; et al. The current and future global distribution and population at risk of dengue. Nat. Microbiol. 2019, 4, 1508–1515. [Google Scholar] [CrossRef] [PubMed]
- Dowd, K.A.; Pierson, T.C. Antibody-mediated neutralization of flaviviruses: A reductionist view. Virology 2011, 411, 306–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Bardelli, M.; Espinosa, D.A.; Pedotti, M.; Ng, T.-S.; Bianchi, S.; Simonelli, L.; Lim, E.X.; Foglierini, M.; Zatta, F.; et al. A Human Bi-specific Antibody against Zika Virus with High Therapeutic Potential. Cell 2017, 171, 229–241.e15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, X.; Deng, Y.; Wang, H.; Ji, G.; Tan, W.; Jiang, T.; Li, X.; Zhao, H.; Xia, T.; Meng, Y.; et al. A bispecific antibody effectively neutralizes all four serotypes of dengue virus by simultaneous blocking virus attachment and fusion. mAbs 2016, 8, 574–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dejnirattisai, W.; Jumnainsong, A.; Onsirisakul, N.; Fitton, P.; Vasanawathana, S.; Limpitikul, W.; Puttikhunt, C.; Edwards, C.; Duangchinda, T.; Supasa, S.; et al. Cross-Reacting Antibodies Enhance Dengue Virus Infection in Humans. Science 2010, 328, 745–748. [Google Scholar] [CrossRef] [Green Version]
- de Alwis, R.; Smith, S.A.; Olivarez, N.P.; Messer, W.B.; Huynh, J.P.; Wahala, W.M.P.B.; White, L.J.; Diamond, M.S.; Baric, R.S.; Crowe, J.E.; et al. Identification of human neutralizing antibodies that bind to complex epitopes on dengue virions. Proc. Natl. Acad. Sci. USA 2012, 109, 7439–7444. [Google Scholar] [CrossRef] [Green Version]
- Teoh, E.P.; Kukkaro, P.; Teo, E.W.; Lim, A.P.C.; Tan, T.T.; Yip, A.; Schul, W.; Aung, M.; Kostyuchenko, V.A.; Leo, Y.S.; et al. The Structural Basis for Serotype-Specific Neutralization of Dengue Virus by a Human Antibody. Sci. Transl. Med. 2012, 4, 139ra83. [Google Scholar] [CrossRef]
- Fibriansah, G.; Ibarra, K.D.; Ng, T.-S.; Smith, S.A.; Tan, J.L.; Lim, X.-N.; Ooi, J.S.G.; Kostyuchenko, V.A.; Wang, J.; de Silva, A.M.; et al. Cryo-EM structure of an antibody that neutralizes dengue virus type 2 by locking E protein dimers. Science 2015, 349, 88–91. [Google Scholar] [CrossRef] [Green Version]
- Dejnirattisai, W.; Wongwiwat, W.; Supasa, S.; Zhang, X.; Dai, X.; Rouvinski, A.; Jumnainsong, A.; Edwards, C.; Quyen, N.T.H.; Duangchinda, T.; et al. A new class of highly potent, broadly neutralizing antibodies isolated from viremic patients infected with dengue virus. Nat. Immunol. 2014, 16, 170–177. [Google Scholar] [CrossRef] [Green Version]
- Beltramello, M.; Williams, K.L.; Simmons, C.P.; Macagno, A.; Simonelli, L.; Quyen, N.T.H.; Sukupolvi-Petty, S.; Navarro-Sanchez, E.; Young, P.; de Silva, A.M.; et al. The Human Immune Response to Dengue Virus Is Dominated by Highly Cross-Reactive Antibodies Endowed with Neutralizing and Enhancing Activity. Cell Host Microbe 2010, 8, 271–283. [Google Scholar] [CrossRef] [Green Version]
- Edeling, M.A.; Austin, S.K.; Shrestha, B.; Dowd, K.A.; Mukherjee, S.; Nelson, C.A.; Johnson, S.; Mabila, M.N.; Christian, E.; Rucker, J.; et al. Potent Dengue Virus Neutralization by a Therapeutic Antibody with Low Monovalent Affinity Requires Bivalent Engagement. PLoS Pathog. 2014, 10, e1004072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Midgley, C.M.; Flanagan, A.; Tran, H.B.; Dejnirattisai, W.; Chawansuntati, K.; Jumnainsong, A.; Wongwiwat, W.; Duangchinda, T.; Mongkolsapaya, J.; Grimes, J.M.; et al. Structural Analysis of a Dengue Cross-Reactive Antibody Complexed with Envelope Domain III Reveals the Molecular Basis of Cross-Reactivity. J. Immunol. 2012, 188, 4971–4979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lok, S.-M.; Kostyuchenko, V.; Nybakken, G.E.; Holdaway, H.A.; Battisti, A.J.; Sukupolvi-Petty, S.; Sedlak, D.; Fremont, D.H.; Chipman, P.R.; Roehrig, J.; et al. Binding of a neutralizing antibody to dengue virus alters the arrangement of surface glycoproteins. Nat. Struct. Mol. Biol. 2008, 15, 312–317. [Google Scholar] [CrossRef] [PubMed]
- Tharakaraman, K.; Robinson, L.N.; Hatas, A.; Chen, Y.-L.; Siyue, L.; Raguram, S.; Sasisekharan, V.; Wogan, G.N.; Sasisekharan, R. Redesign of a cross-reactive antibody to dengue virus with broad-spectrum activity and increased in vivo potency. Proc. Natl. Acad. Sci. USA 2013, 110, E1555–E1564. [Google Scholar] [CrossRef] [Green Version]
- Schoeder, C.T.; Schmitz, S.; Adolf-Bryfogle, J.; Sevy, A.M.; Finn, J.A.; Sauer, M.F.; Bozhanova, N.G.; Mueller, B.K.; Sangha, A.K.; Bonet, J.; et al. Modeling Immunity with Rosetta: Methods for Antibody and Antigen Design. Biochemistry 2021, 60, 825–846. [Google Scholar] [CrossRef]
- Baker, N.A.; Sept, D.; Joseph, S.; Holst, M.J.; McCammon, J.A. Electrostatics of nanosystems: Application to microtubules and the ribosome. Proc. Natl. Acad. Sci. USA 2001, 98, 10037–10041. [Google Scholar] [CrossRef] [Green Version]
- DeLano, W.L.; Lam, J.W. PyMOL: A communications tool for computational models. In Abstracts of Papers of the American Chemical Society; American Chemical Society: Washington, DC, USA, 2005; Volume 230, pp. U1371–U1372. [Google Scholar]
- Valdés-Tresanco, M.S.; Valiente, P.A.; Moreno, E. gmx_MMPBSA: A New Tool to Perform End-State Free Energy Calculations with GROMACS. J. Chem. Theory Comput. 2021, 17, 6281–6291. [Google Scholar] [CrossRef]
- Schmidt, A.G.; Xu, H.; Khan, A.R.; O’Donnell, T.; Khurana, S.; King, L.R.; Manischewitz, J.; Golding, H.; Suphaphiphat, P.; Carfi, A.; et al. Preconfiguration of the antigen-binding site during affinity maturation of a broadly neutralizing influenza virus antibody. Proc. Natl. Acad. Sci. USA 2013, 110, 264–269. [Google Scholar] [CrossRef] [Green Version]
- Willis, J.R.; Sapparapu, G.; Murrell, S.; Julien, J.-P.; Singh, V.; King, H.G.; Xia, Y.; Pickens, J.A.; Labranche, C.C.; Slaughter, J.; et al. Redesigned HIV antibodies exhibit enhanced neutralizing potency and breadth. J. Clin. Investig. 2015, 125, 2523–2531. [Google Scholar] [CrossRef] [Green Version]
- Warszawski, S.; Katz, A.B.; Lipsh, R.; Khmelnitsky, L.; Ben Nissan, G.; Javitt, G.; Dym, O.; Unger, T.; Knop, O.; Albeck, S.; et al. Optimizing antibody affinity and stability by the automated design of the variable light-heavy chain interfaces. PLoS Comput. Biol. 2019, 15, e1007207. [Google Scholar] [CrossRef] [Green Version]
- Lippow, S.M.; Wittrup, K.D.; Tidor, B. Computational design of antibody-affinity improvement beyond in vivo maturation. Nat. Biotechnol. 2007, 25, 1171–1176. [Google Scholar] [CrossRef] [Green Version]
- Robinson, L.N.; Tharakaraman, K.; Rowley, K.; Costa, V.V.; Chan, K.R.; Wong, Y.H.; Ong, L.C.; Tan, H.C.; Koch, T.; Cain, D.; et al. Structure-Guided Design of an Anti-dengue Antibody Directed to a Non-immunodominant Epitope. Cell 2015, 162, 493–504. [Google Scholar] [CrossRef] [Green Version]
- James, L.C.; Roversi, P.; Tawfik, D.S. Antibody Multispecificity Mediated by Conformational Diversity. Science 2003, 299, 1362–1367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selzer, T.; Albeck, S.; Schreiber, G. Rational design of faster associating and tighter binding protein complexes. Nat. Genet. 2000, 7, 537–541. [Google Scholar] [CrossRef]
- Wahala, W.; Kraus, A.A.; Haymore, L.B.; Accavitti-Loper, M.A.; de Silva, A.M. Dengue virus neutralization by human immune sera: Role of envelope protein domain III-reactive antibody. Virology 2009, 392, 103–113. [Google Scholar] [CrossRef] [Green Version]
- Cockburn, J.J.; Sanchez, M.E.N.; Fretes, N.; Urvoas, A.; Staropoli, I.; Kikuti, C.M.; Coffey, L.L.; Seisdedos, F.A.; Bedouelle, H.; Rey, F.A. Mechanism of Dengue Virus Broad Cross-Neutralization by a Monoclonal Antibody. Structure 2012, 20, 303–314. [Google Scholar] [CrossRef] [Green Version]
- Li, X.Q.; Qiu, L.W.; Chen, Y.; Wen, K.; Cai, J.P.; Chen, J.; Pan, Y.X.; Li, J.; Hu, D.M.; Huang, Y.F.; et al. Dengue virus envelope domain III immunization elicits predominantly cross-reactive, poorly neutralizing an-tibodies localized to the AB loop: Implications for dengue vaccine design. J. Gen. Virol. 2013, 94 Pt 10, 2191–2201. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Watterson, D.; Chang, C.W.; Che, X.Y.; Li, X.Q.; Ericsson, D.J.; Qiu, L.W.; Cai, J.P.; Chen, J.; Fry, S.R.; et al. Structural and Functional Characterization of a Cross-Reactive Dengue Virus Neutralizing Antibody that Recog-nizes a Cryptic Epitope. Structure 2018, 26, 51–59.e4. [Google Scholar] [CrossRef] [Green Version]
- Lim, X.-N.; Shan, C.; Marzinek, J.; Dong, H.; Ng, T.S.; Ooi, J.; Fibriansah, G.; Wang, J.; Verma, C.S.; Bond, P.J.; et al. Molecular basis of dengue virus serotype 2 morphological switch from 29 °C to 37 °C. PLoS Pathog. 2019, 15, e1007996. [Google Scholar] [CrossRef] [Green Version]
- Fibriansah, G.; Ng, T.-S.; Kostyuchenko, V.A.; Lee, J.; Lee, S.; Wang, J.; Lok, S.-M. Structural Changes in Dengue Virus When Exposed to a Temperature of 37 °C. J. Virol. 2013, 87, 7585–7592. [Google Scholar] [CrossRef] [Green Version]
- Hu, D.; Zhu, Z.; Li, S.; Deng, Y.; Wu, Y.; Zhang, N.; Puri, V.; Wang, C.; Zou, P.; Lei, C.; et al. A broadly neutralizing germline-like human monoclonal antibody against dengue virus envelope domain III. PLoS Pathog. 2019, 15, e1007836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jazii, F.R.; Karkhane, A.; Yakhchali, B.; Fatemi, S.; Deezagi, A. A simplified purification procedure for recombinant human granulocyte macrophage-colony stimulating factor from periplasmic space of Escherichia coli. J. Chromatogr. B 2007, 856, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Lindorff-Larsen, K.; Piana, S.; Palmo, K.; Maragakis, P.; Klepeis, J.L.; Dror, R.O.; Shaw, D.E. Improved side-chain torsion potentials for the Amber ff99SB protein force field. Proteins 2010, 78, 1950–1958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onufriev, A.; Bashford, D.; Case, D.A. Exploring protein native states and large-scale conformational changes with a modified generalized born model. Proteins Struct. Funct. Bioinform. 2004, 55, 383–394. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ka (1/Ms) | kd (1/s) | KD (nM) | KD Error (nM) | |
|---|---|---|---|---|
| DENV1 | ||||
| WT | 3.49 × 104 | 7.55 × 10−3 | 216 | 7.23 |
| A54E | 4.43 × 104 | 6.17 × 10−3 | 140 | 4.93 |
| Y105R | 4.35 × 104 | 1.85 × 10−3 | 42.6 | 1.44 |
| A54E-Y105R | 4.39 × 104 | 1.20 × 10−3 | 27.4 | 0.51 |
| DENV2 | ||||
| WT | 1.07 × 105 | 4.71 × 10−2 | 440 | 5.51 |
| A54E | 1.97 × 105 | 8.22 × 10−3 | 41.9 | 2.84 |
| Y105R | 9.62 × 104 | 1.95 × 10−2 | 203 | 3.21 |
| A54E-Y105R | 2.22 × 105 | 2.74 × 10−3 | 12.3 | 0.41 |
| DENV3 | ||||
| WT | 8.70 × 104 | 7.92 × 10−3 | 91 | 3.67 |
| A54E | 3.54 × 105 | 3.64 × 10−3 | 10.3 | 0.52 |
| Y105R | 8.52 × 104 | 4.19 × 10−3 | 49.2 | 2.87 |
| A54E-Y105R | 2.22 × 105 | 1.58 × 10−3 | 7.11 | 0.16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, H.; Tan, J.; Zhou, B.; Guan, X.; Zhong, Q.; Wang, J. Charged Residue Implantation Improves the Affinity of a Cross-Reactive Dengue Virus Antibody. Int. J. Mol. Sci. 2022, 23, 4197. https://doi.org/10.3390/ijms23084197
Wei H, Tan J, Zhou B, Guan X, Zhong Q, Wang J. Charged Residue Implantation Improves the Affinity of a Cross-Reactive Dengue Virus Antibody. International Journal of Molecular Sciences. 2022; 23(8):4197. https://doi.org/10.3390/ijms23084197
Chicago/Turabian StyleWei, Huiling, Jie Tan, Bingjie Zhou, Xiaotong Guan, Qiaoxian Zhong, and Jiaqi Wang. 2022. "Charged Residue Implantation Improves the Affinity of a Cross-Reactive Dengue Virus Antibody" International Journal of Molecular Sciences 23, no. 8: 4197. https://doi.org/10.3390/ijms23084197