
The Thermodynamically Expensive Contribution of Three Calcium Sources to Somatic Release of Serotonin


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
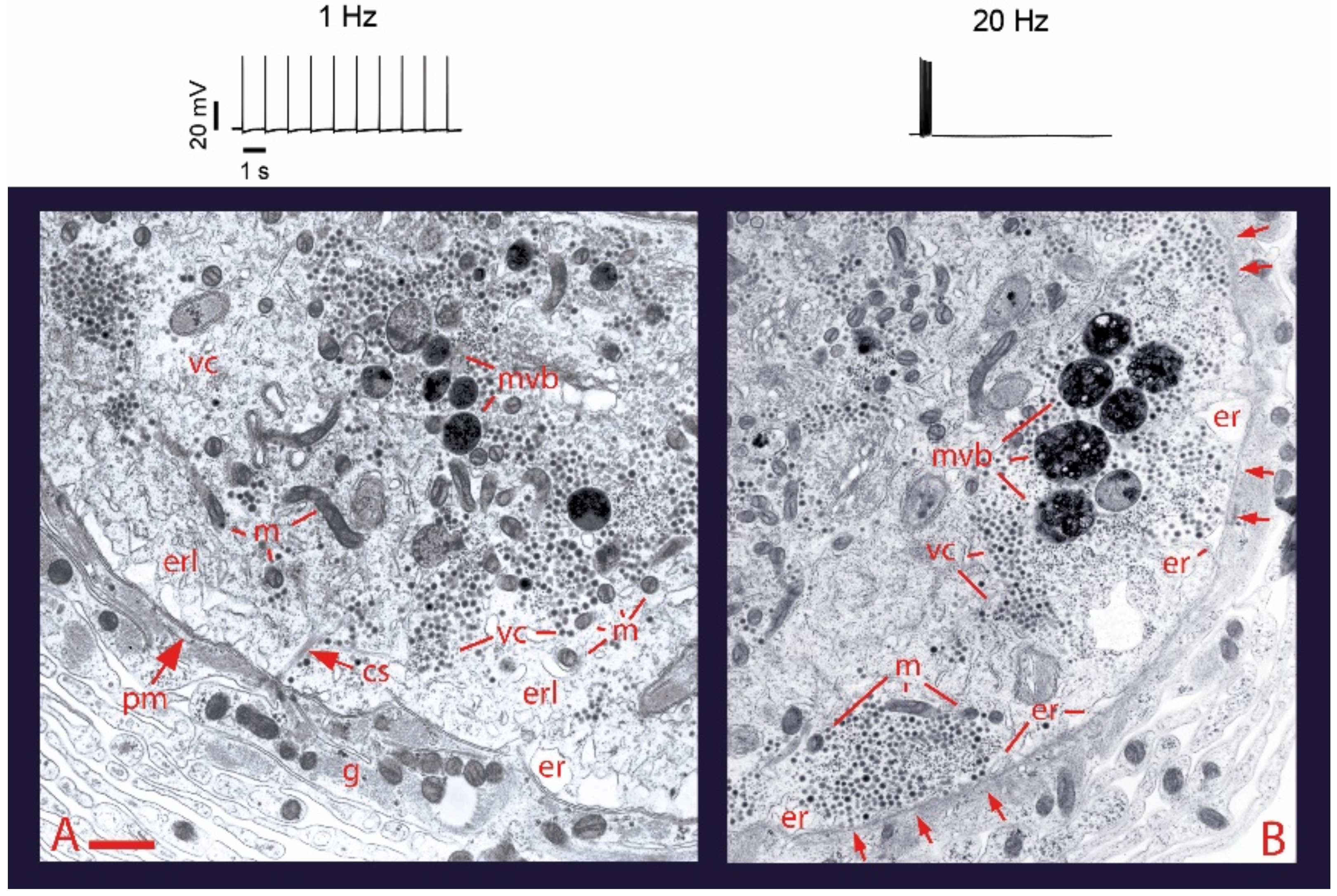
2. Electrical Activity and Calcium Promote Vesicle Transport and Somatic Exocytosis
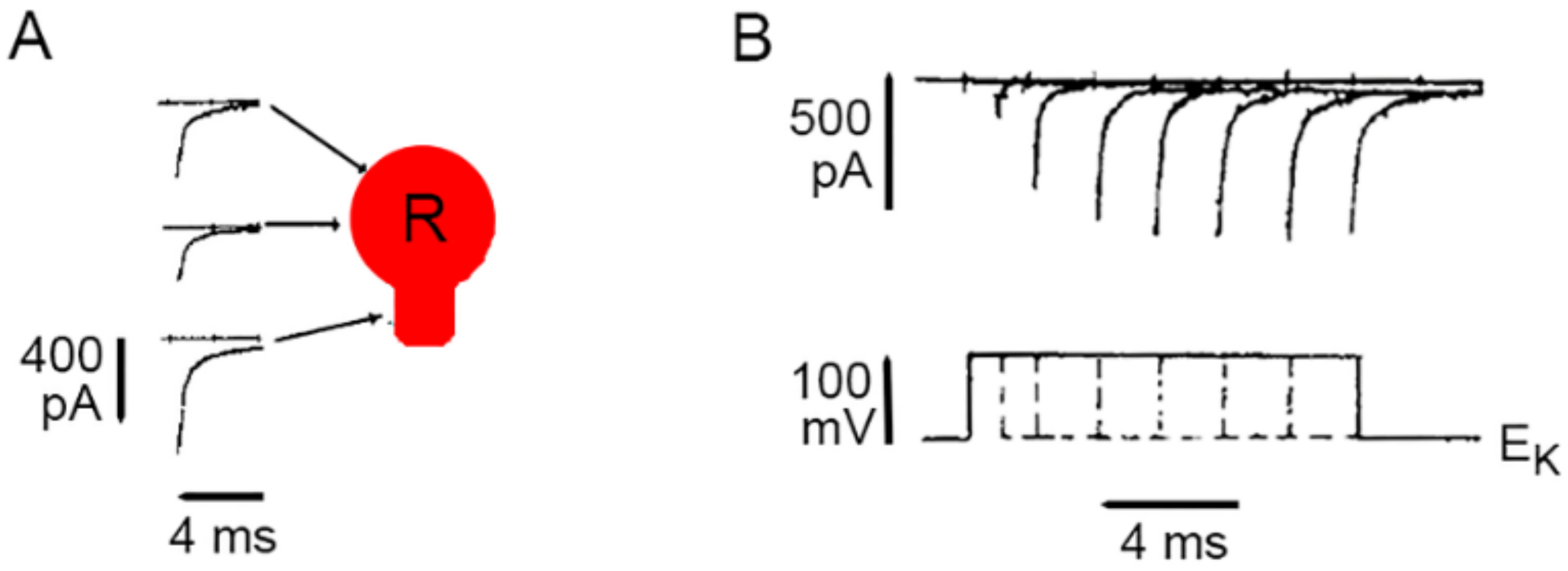
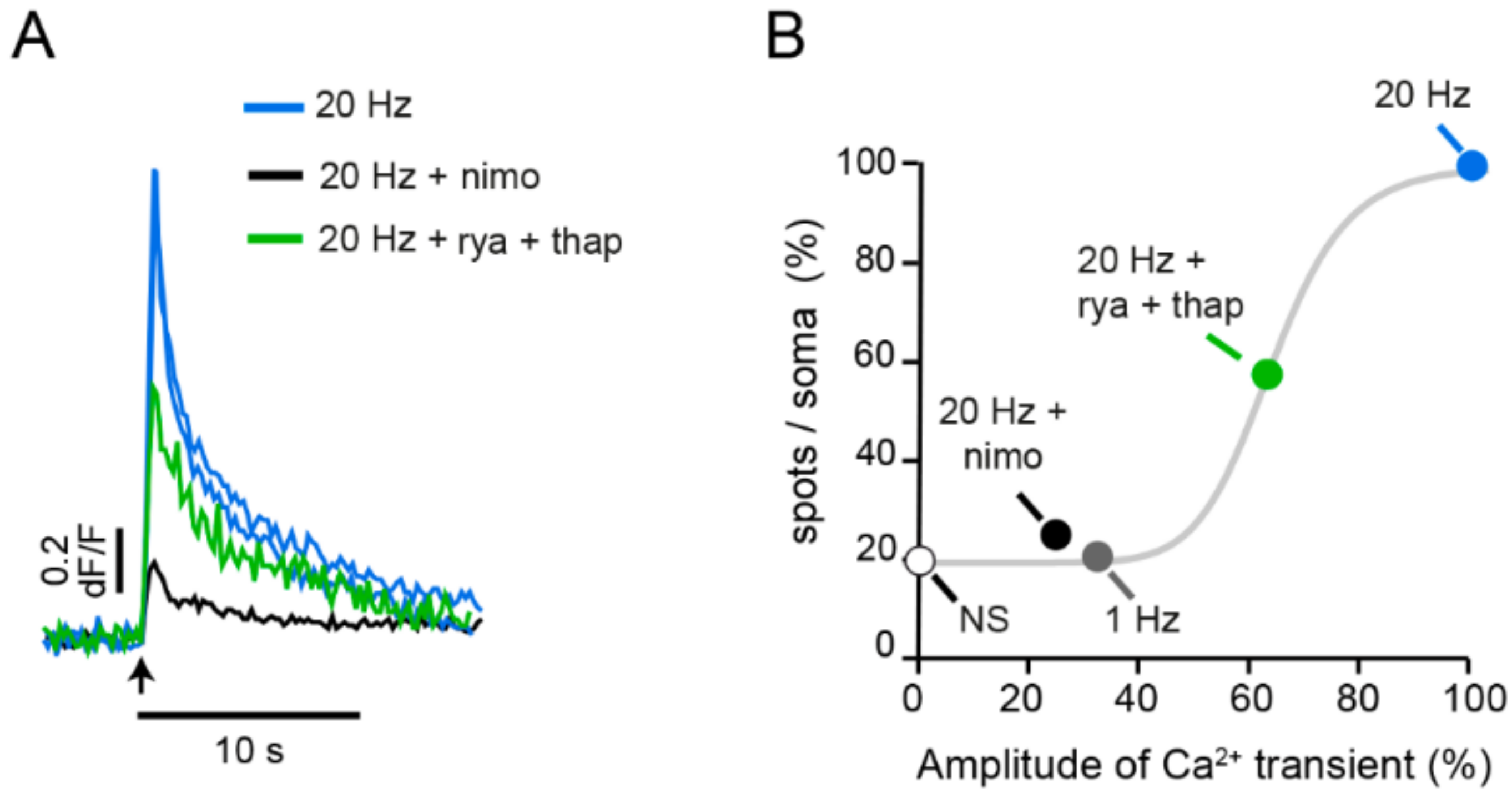
3. Calcium Enters the Neuronal Soma through L-Type Channels
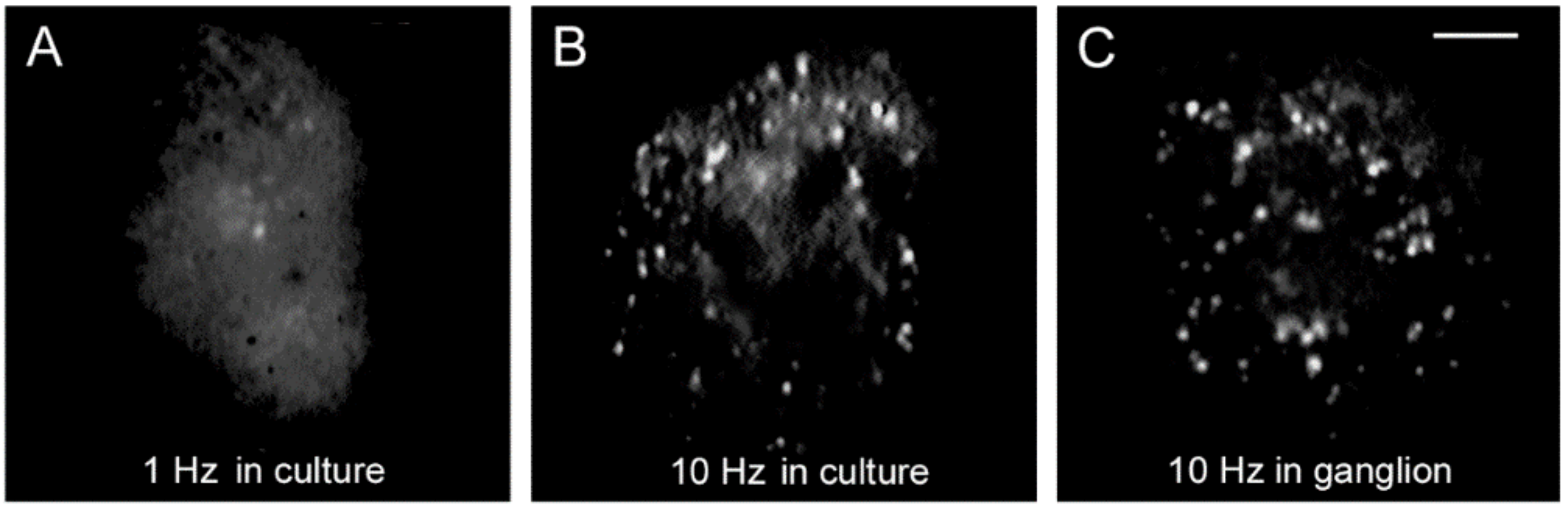
4. High-Frequency Activation of L-Type Channels Promotes Somatic Exocytosis
5. Analysis of the Calcium Dynamics
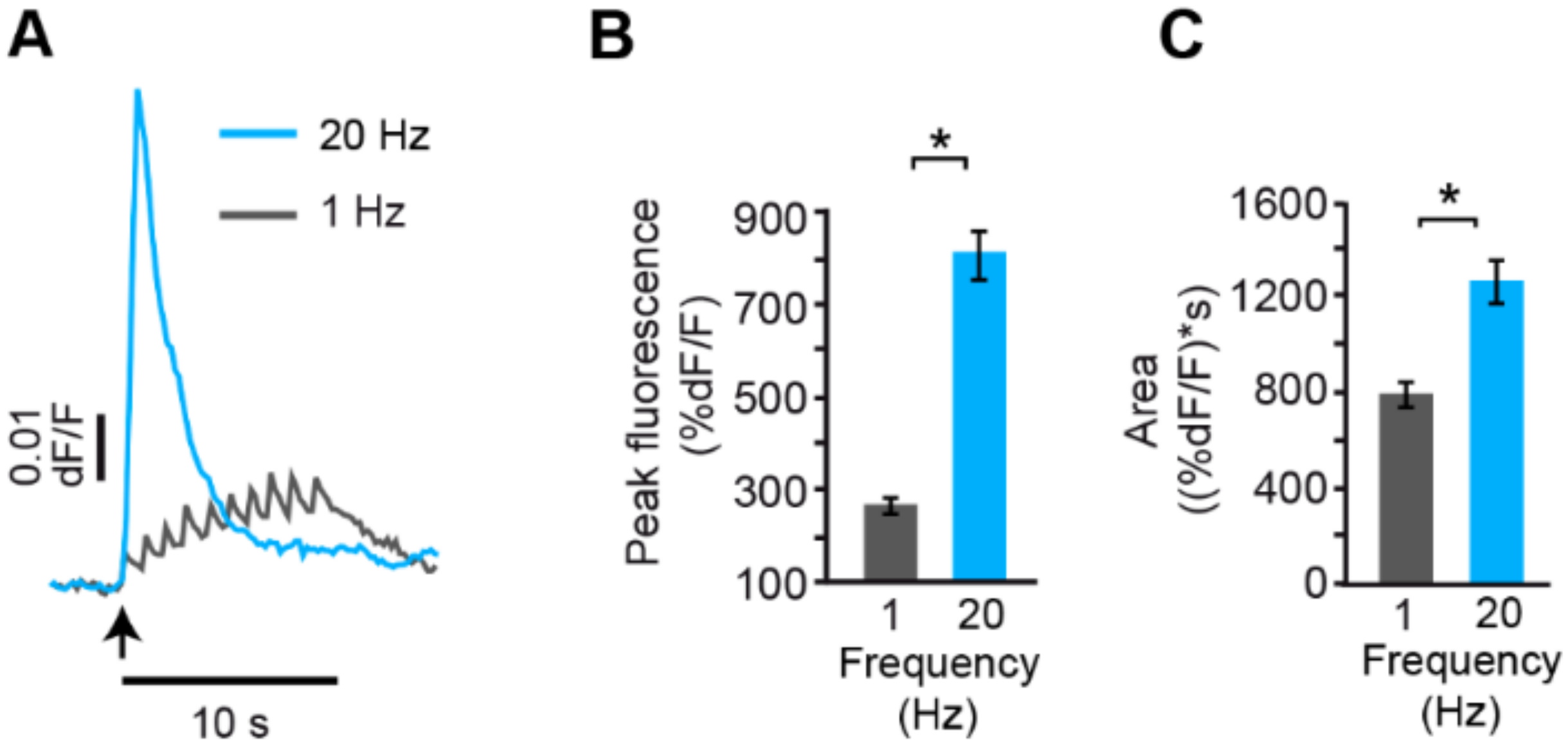
6. The Fast Calcium Transient Determines the Amount of Release
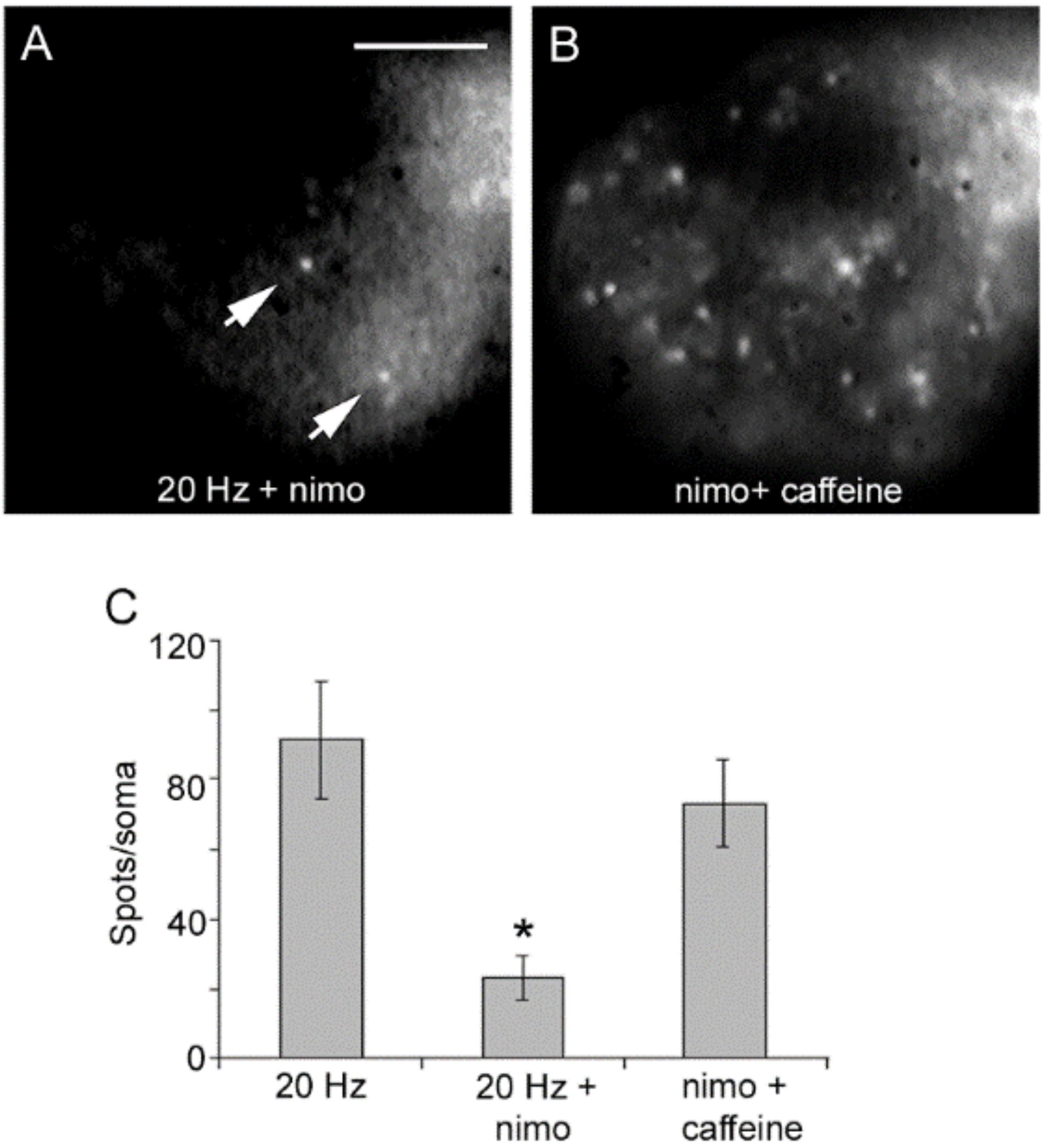
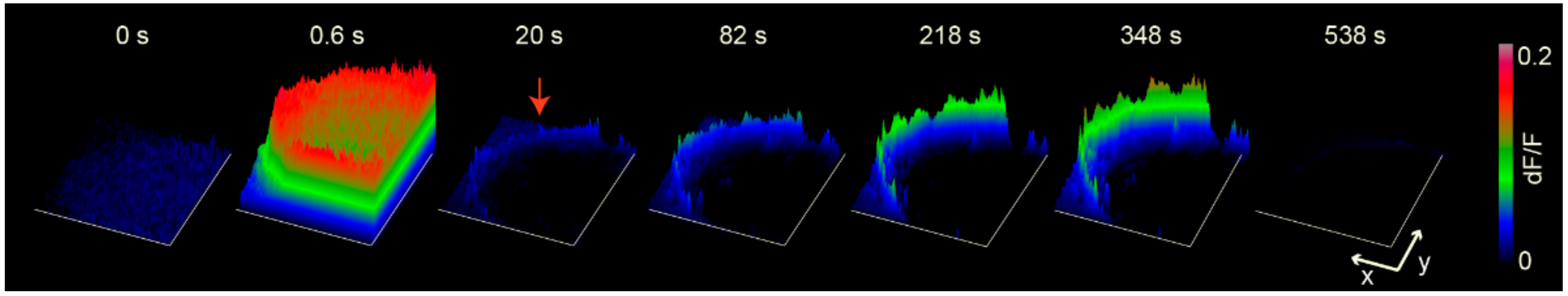
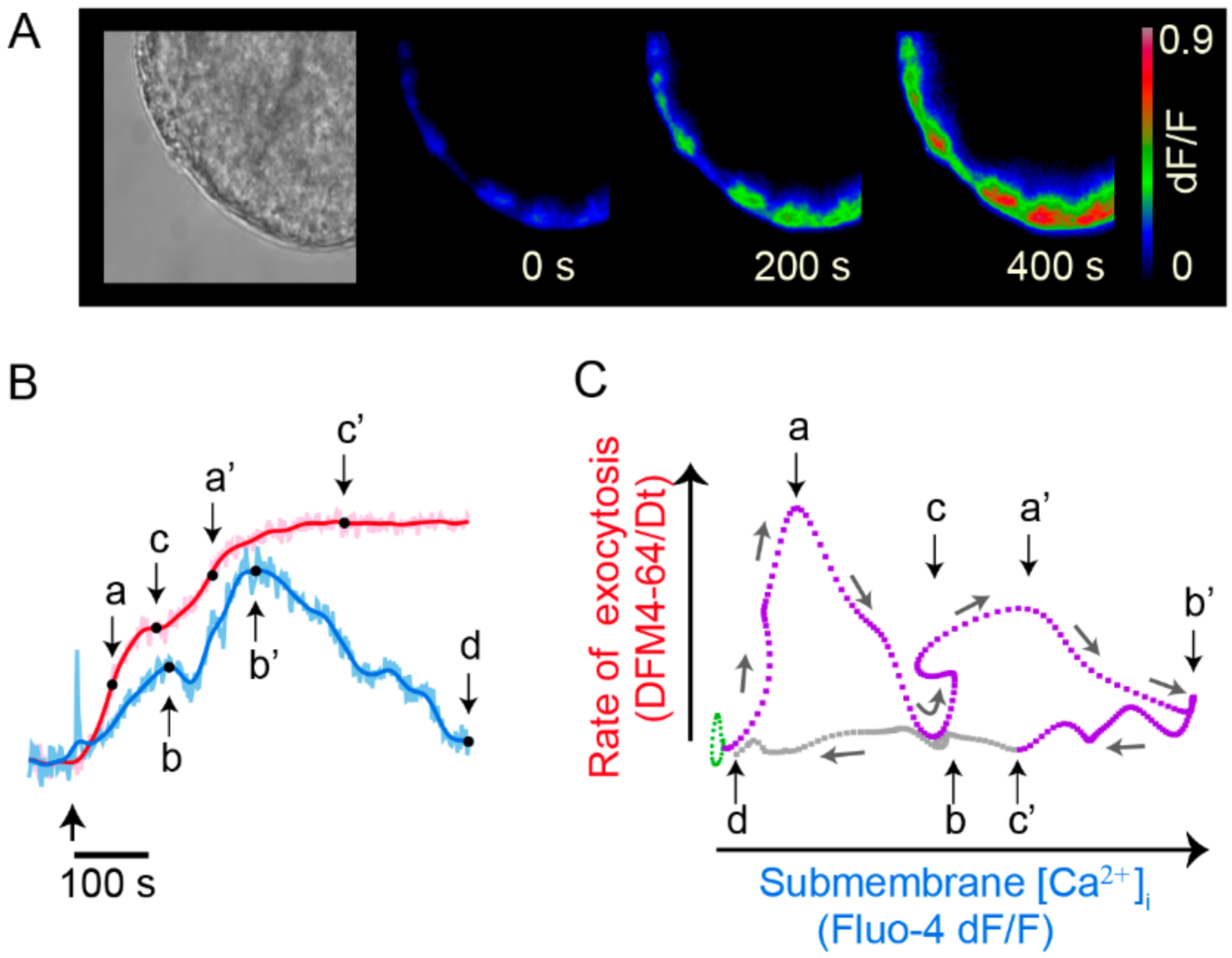
7. Exocytosis Is Sustained by a Positive Feedback Loop
8. Serotonin That Has Been Released Activates the Third Calcium Pool
9. The Possible Calcium Sensors for Somatic Exocytosis
10. On the High Energy Cost of the Use of Three Calcium Pools
11. Summary
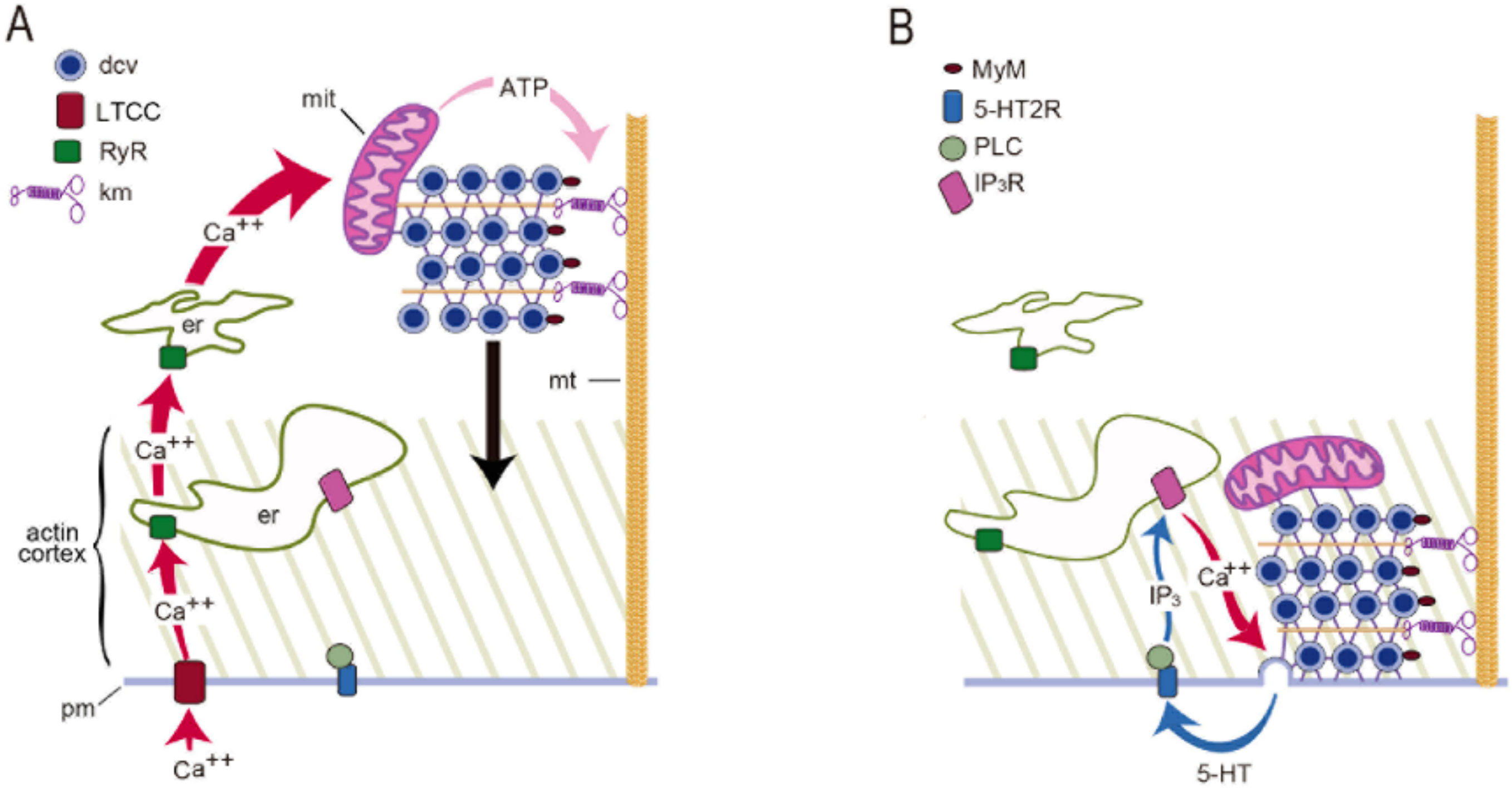
- Somatic exocytosis by Retzius neurons is regulated by a sequential activation of three calcium sources. The first two sources cooperate to regulate the amount of transmitter transported to the plasma membrane; the third source activates vesicle fusion.
- Increases in the frequency of electrical activity allow for the summation of transmembrane calcium transients generated upon activation of L-type calcium channels. As the stimulation frequency increases, summation increases the free calcium concentration and activates a second calcium source: calcium-induced calcium release.
- Calcium-induced calcium release increases the amplitude of the intracellular calcium transient. A resulting calcium tsunami invades the mitochondria and makes them produce ATP for the vesicle transport to the plasma membrane. The amplitude of the calcium transient in the mitochondria determines the number of vesicles transported and fused.
- Serotonin that is released maintains a large-scale exocytosis for hundreds of seconds through a third calcium source: the activation of 5-HT2 receptors coupled to phospholipase C and production of IP3 produces calcium release from endoplasmic reticulum adjacent to the plasma membrane.
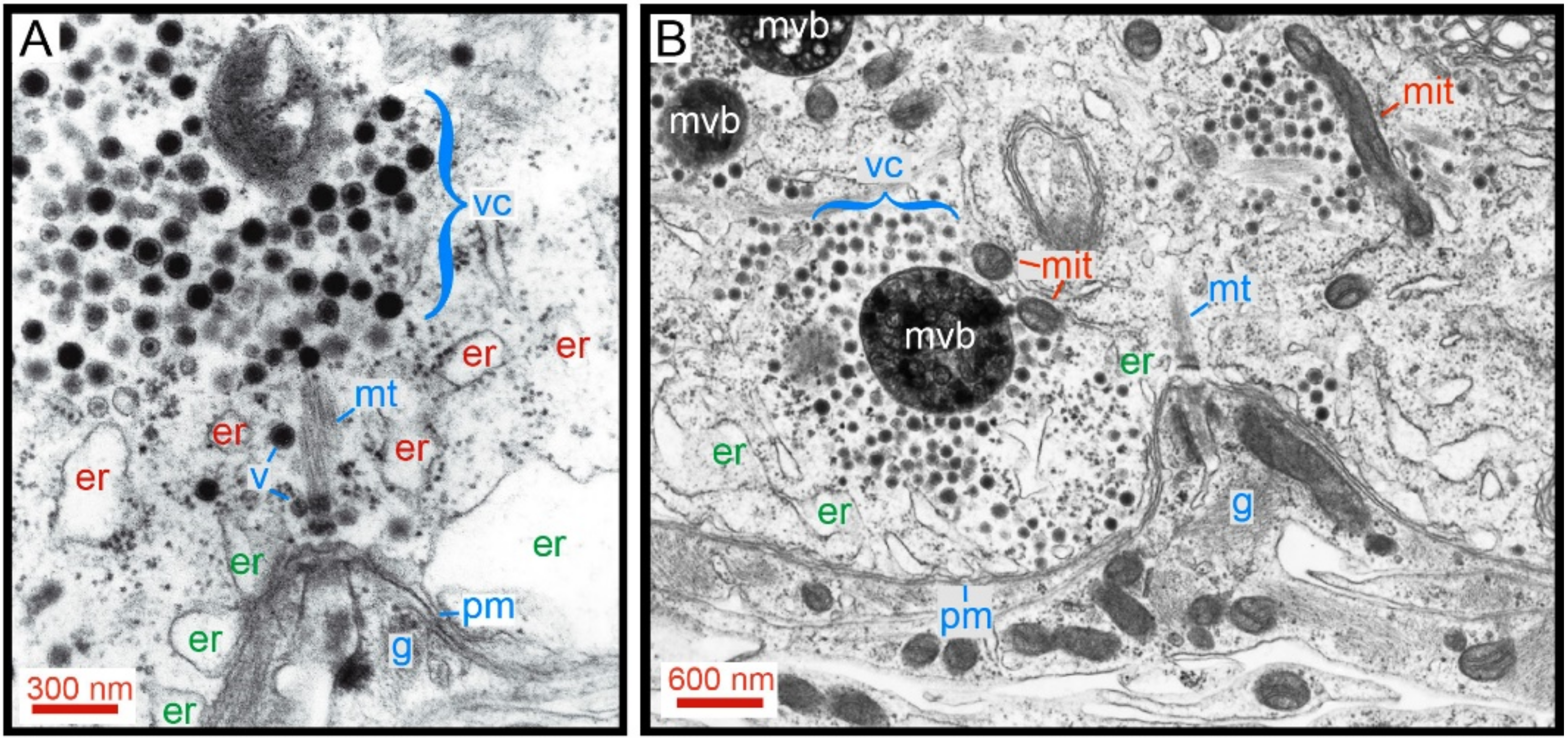
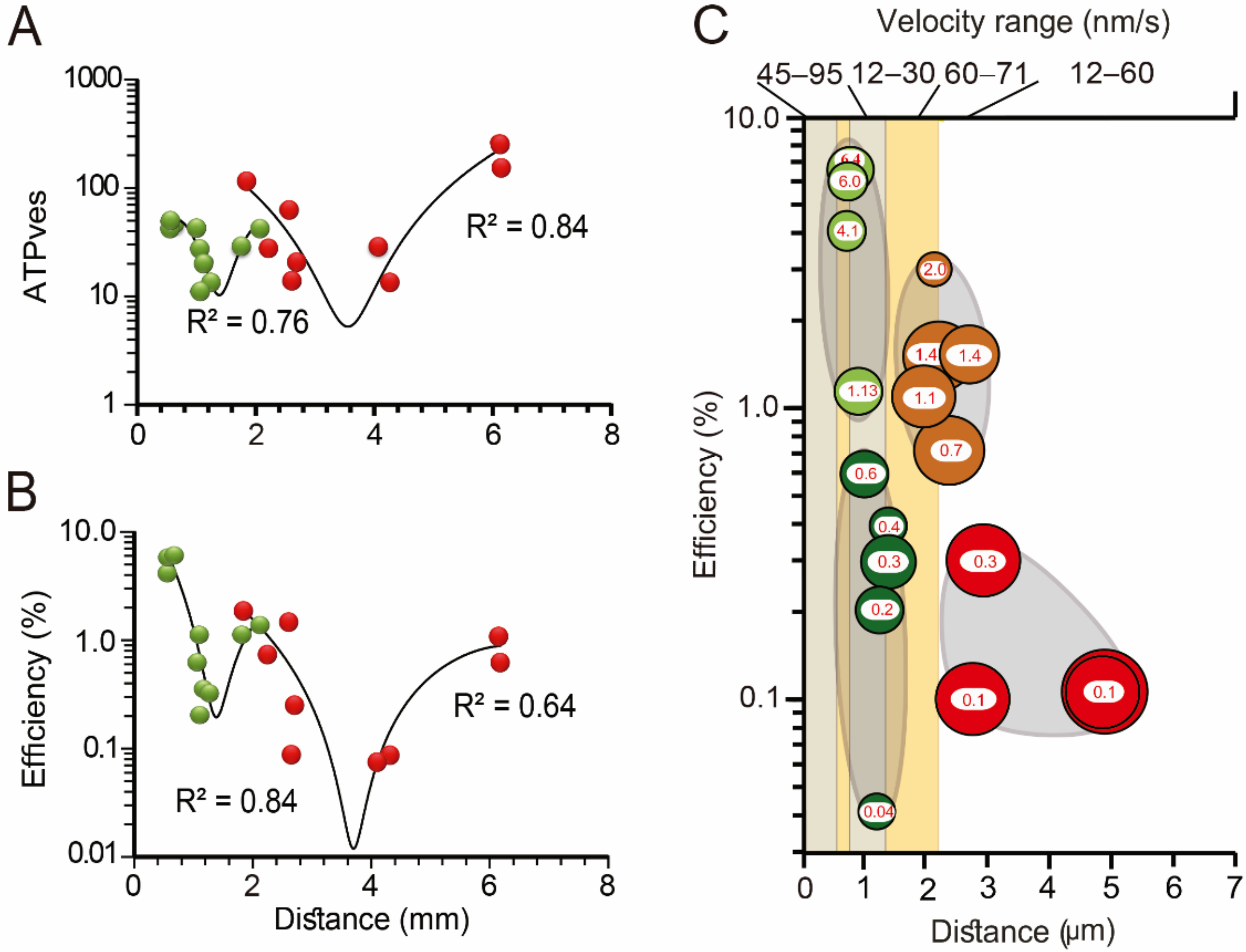
- The calcium-releasing endoplasmic reticulum and the actin cortex generate obstacles to the transport pathway of vesicles. The resulting friction forces reduce the thermodynamic efficiency of the ATP usage, adding energy cost to somatic exocytosis.
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Huang, L.Y.; Neher, E. Ca(2+)-dependent exocytosis in the somata of dorsal root ganglion neurons. Neuron 1996, 17, 135–145. [Google Scholar] [CrossRef] [Green Version]
- Jaffe, E.H.; Marty, A.; Schulte, A.; Chow, R.H. Extrasynaptic vesicular transmitter release from the somata of substantia nigra neurons in rat midbrain slices. J. Neurosci. 1998, 18, 3548–3553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puopolo, M.; Hochstetler, S.E.; Gustincich, S.; Wightman, R.M.; Raviola, E. Extrasynaptic release of dopamine in a retinal neuron: Activity dependence and transmitter modulation. Neuron 2001, 30, 211–225. [Google Scholar] [CrossRef] [Green Version]
- Kaushalya, S.K.; Desai, R.; Arumugam, S.; Ghosh, H.; Balaji, J.; Maiti, S. Three-photon microscopy shows that somatic release can be a quantitatively significant component of serotonergic neurotransmission in the mammalian brain. J. Neurosci. Res. 2008, 86, 3469–8340. [Google Scholar] [CrossRef]
- Huang, H.P.; Zhu, F.P.; Chen, X.W.; Xu, Z.Q.; Zhang, C.X.; Zhou, Z. Physiology of quantal norepinephrine release from somatodendritic sites of neurons in locus coeruleus. Front. Mol. Neurosci. 2012, 5, 29. [Google Scholar] [CrossRef] [Green Version]
- Trueta, C.; De-Miguel, F.F. Extrasynaptic exocytosis and its mechanisms: A source of molecules mediating volume transmission in the nervous system. Front. Physiol. 2012, 3, 319. [Google Scholar] [CrossRef] [Green Version]
- Sulzer, D.; Cragg, S.J.; Rice, M.E. Striatal dopamine neurotransmission: Regulation of release and uptake. Basal Ganglia 2016, 6, 123–148. [Google Scholar] [CrossRef] [Green Version]
- Grinevich, V.; Ludwig, M. The multiple faces of the oxytocin and vasopressin systems in the brain. J. Neuroendocr. 2021, 33, e13004. [Google Scholar] [CrossRef]
- Borroto-Escuela, D.O.; Perez De La Mora, M.; Manger, P.; Narváez, M.; Beggiato, S.; Crespo-Ramírez, M.; Navarro, G.; Wydra, K.; Díaz-Cabiale, Z.; Rivera, A.; et al. Brain Dopamine Transmission in Health and Parkinson’s Disease: Modulation of Synaptic Transmission and Plasticity Through Volume Transmission and Dopamine Heteroreceptors. Front. Syn. Neurosci. 2018, 10, 20. [Google Scholar] [CrossRef]
- Newman, E.A. Glial cell regulation of neuronal activity and blood flow in the retina by release of gliotransmitters. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2015, 370, 20140195. [Google Scholar] [CrossRef]
- Igelhorst, B.A.; Niederkinkhaus, V.; Karus, C.; Lange, M.D.; Dietzel, I.D. Regulation of neuronal excitability by release of proteins from glial cells. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2015, 370, 20140194. [Google Scholar] [CrossRef] [Green Version]
- Kravitz, E.A. Hormonal control of behavior: Amines and the biasing of behavioral output in lobsters. Science 1988, 241, 1775–1781. [Google Scholar] [CrossRef]
- Hirasawa, H.; Contini, M.; Raviola, E. Extrasynaptic release of GABA and dopamine by retinal dopaminergic neurons. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370, 20140186. [Google Scholar] [CrossRef] [Green Version]
- Ludwig, M.; Apps, D.; Menzies, J.; Patel, J.C.; Rice, M.E. Dendritic Release of Neurotransmitters. Comp. Physiol. 2016, 7, 235–252. [Google Scholar]
- Quentin, E.; Belmer, A.; Maroteaux, L. Somato-Dendritic Regulation of Raphe Serotonin Neurons; A Key to Antidepressant Action. Front. Neurosci. 2018, 12, 982. [Google Scholar] [CrossRef] [Green Version]
- Hökfelt, T.; Barde, S.; Xu, Z.D.; Kuteeva, E.; Rüegg, J.; Le Maitre, E.; Risling, M.; Kehr, J.; Ihnatko, R.; Theodorsson, E.; et al. Neuropeptide and Small Transmitter Coexistence: Fundamental Studies and Relevance to Mental Illness. Front. Neural. Circ. 2018, 12, 106. [Google Scholar] [CrossRef] [Green Version]
- Delgado-Lezama, R.; Bravo-Hernández, M.; Franco-Enzástiga, Ú.; De la Luz-Cuellar, Y.E.; Alvarado-Cervantes, N.S.; Raya-Tafolla, G.; Martínez-Zaldivar, L.A.; Vargas-Parada, A.; Rodríguez-Palma, E.J.; Vidal-Cantú, G.C.; et al. The role of spinal cord extrasynaptic α5 GABAA receptors in chronic pain. Physl. Rep. 2021, 16, e14984. [Google Scholar]
- Wacker, D.; Ludwig, M. The role of vasopressin in olfactory and visual processing. Cell Tis. Res. 2021, 375, 201–215. [Google Scholar] [CrossRef]
- Brown, C.H.; Ludwig, M.; Tasker, J.G.; Stern, J.E. Somato-dendritic vasopressin and oxytocin secretion in endocrine and autonomic regulation. J. Neuroendocr. 2020, 32, e12856. [Google Scholar] [CrossRef]
- Livero, G. Somatostatin, a Presynaptic Modulator of Glutamatergic Signal in the Central Nervous System. Int. J. Mol. Sci. 2021, 22, 5864. [Google Scholar]
- Tobin, V.A.; Ludwig, M. The role of the actin cytoskeleton in oxytocin and vasopressin release from rat supraoptic nucleus neurons. J. Physiol. 2007, 582, 1337–1348. [Google Scholar] [CrossRef]
- De-Miguel, F.F.; Leon-Pinzon, C.; Torres-Platas, S.G.; Del-Pozo, V.; Hernández-Mendoza, G.A.; Aguirre-Olivas, D.; Mendez, B.; Moore, S.; Sánchez-Sugía, C.; García-Aguilera, M.A.; et al. Extrasynaptic Communication. Front. Mol. Neurosci. 2021, 14, 638858. [Google Scholar] [CrossRef] [PubMed]
- De-Miguel, F.F.; Leon-Pinzon, C.; Noguez, P.; Mendez, B. Serotonin release from the neuronal cell body and its long-lasting effects on the nervous system. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370, 20140196. [Google Scholar] [CrossRef] [Green Version]
- Mesce, K.A.; Esch, T.; Kristan, W.B., Jr. Cellular substrates of action selection: A cluster of higher-order descending neurons shapes body posture and locomotion. J. Comp. Physiol. A 2008, 194, 469–481. [Google Scholar] [CrossRef]
- Ashaber, M.; Tomina, Y.; Kassraian, P.; Bushong, E.A.; Kristan, W.B.; Ellisman, M.H.; Wagenaar, D.A. Anatomy and activity patterns in a multifunctional motor neuron and its surrounding circuits. eLife 2021, 10, e61881. [Google Scholar] [CrossRef]
- De-Miguel, F.F.; Santamaría-Holek, I.; Noguez, P.; Bustos, C.; Hernández-Lemus, E.; Rubí, J.M. Biophysics of active vesicle transport, an intermediate step that couples excitation and exocytosis of serotonin in the neuronal soma. PLoS ONE 2012, 460, e45454. [Google Scholar]
- Ma, P.M.; Beltz, B.S.; Kravitz, E.A. Serotonin-containing neurons in lobsters: Their role as gain-setters in postural control mechanisms. J. Neurophysiol. 1992, 68, 36–54. [Google Scholar] [CrossRef]
- García-Pérez, E.; Vargas-Caballero, M.; Velazquez-Ulloa, N.; Minzoni, A.; De-Miguel, F.F. Synaptic integration in electrically coupled neurons. Biophys. J. 2004, 86, 646–655. [Google Scholar] [CrossRef] [Green Version]
- Aghajanian, G.K.; Foote, W.E.; Sheard, M.H. Lysergic acid diethylamide: Sensitive neuronal units in the midbrain raphe. Science 1968, 161, 706–708. [Google Scholar] [CrossRef]
- Mosko, S.S.; Jacobs, B.L. Midbrain raphe neurons: Spontaneous activity and response to light. Phys. Behav. 1974, 13, 589–593. [Google Scholar] [CrossRef]
- Trueta, C.; Kuffler, D.P.; De-Miguel, F.F. Cycling of dense core vesicles involved in somatic exocytosis of serotonin by leech neurons. Front. Physiol. 2012, 3, 175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trueta, C.; Méndez, B.; De-Miguel, F.F. Somatic exocytosis of serotonin mediated by L-type calcium channels in cultured leech neurones. J. Physiol. 2003, 547, 405–416. [Google Scholar] [CrossRef] [PubMed]
- Trueta, C.; Sánchez-Armass, S.; Morales, M.A.; De-Miguel, F.F. Calcium-induced calcium release contributes to somatic secretion of serotonin in leech Retzius neurons. J. Neurobiol. 2004, 61, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.W.; Nurse, C.A.; Wong, V.; Cutz, E. Hypoxia-induced secretion of serotonin from intact pulmonary neuroepithelial bodies in neonatal rabbit. J. Physiol. 2002, 539, 503–510. [Google Scholar] [CrossRef]
- Simmons, M.L.; Terman, G.W.; Gibbs, S.M.; Chavkin, C. L-type calcium channels mediate dynorphin neuropeptide release from dendrites but not axons of hippocampal granule cells. Neuron 1995, 14, 1265–1272. [Google Scholar] [CrossRef] [Green Version]
- Shibuya, I.; Noguchi, J.; Tanaka, K.; Harayama, N.; Inoue, U.; Kabashima, N.; Ueta, Y.; Hattori, Y.; Yamashita, H. PACAP increases the cytosolic Ca2+ concentration and stimulates somatodendritic vasopressin release in rat supraoptic neurons. J. Neuroendocr. 1998, 10, 31–42. [Google Scholar] [CrossRef]
- De Kock, C.P.; Burnashev, N.; Lodder, J.C.; Mansvelder, H.D.; Brussaard, A.B. NMDA receptors induce somatodendritic secretion in hypothalamic neurones of lactating female rats. J. Physiol. 2004, 561, 53–64. [Google Scholar] [CrossRef]
- De Kock, C.P.; Cornelisse, L.N.; Burnashev, N.; Lodder, J.C.; Timmerman, A.J.; Couey, J.J.; Mansvelder, H.D.; Brussaard, A.B. NMDA receptors trigger neurosecretion of 5-HT within dorsal raphe nucleus of the rat in the absence of action potential firing. J. Physiol. 2006, 577, 891–905. [Google Scholar] [CrossRef]
- Fernandez de Miguel, F.; Cooper, R.; Adams, W.B. Synaptogenesis and calcium current distribution in cultured leech neurons. Proc. R. Soc. Lond. B 1992, 247, 215–221. [Google Scholar]
- Leon-Pinzon, C.; Cercós, M.G.; Noguez, P.; Trueta, C.; De-Miguel, F.F. Exocytosis of serotonin from the neuronal soma is sustained by a serotonin and calcium-dependent feedback loop. Front. Cell Neurosci. 2014, 8, 169. [Google Scholar] [CrossRef] [Green Version]
- Dreifuss, J.J.; Kalnins, I.; Kelly, J.S.; Ruf, K.B. Action potentials and release of neurohypophysial hormones in vitro. J. Physiol. 1971, 215, 805–817. [Google Scholar] [CrossRef]
- Hörner, M.; Weiger, W.A.; Edwards, D.H.; Kravitz, E.A. Excitation of identified serotonergic neurons by escape command neurons in lobsters. J. Exp. Biol. 1997, 200, 2017–2033. [Google Scholar] [CrossRef]
- Kirischuk, S.; Voitenko, N.; Kostyuk, P.; Verkhratsky, A. Calcium signalling in granule neurones studied in cerebellar slices. Cell Calcium 1996, 19, 59–71. [Google Scholar] [CrossRef]
- Shmigol, A.; Svichar, N.; Kostyuk, P.; Verkhratsky, A. Gradual caffeine-induced Ca2+ release in mouse dorsal root ganglion neurons is controlled by cytoplasmic and luminal Ca2+. Neuroscience 1996, 73, 1061–1067. [Google Scholar] [CrossRef]
- Thayer, S.A.; Hirning, L.D.; Miller, R.J. The role of caffeine-sensitive calcium stores in the regulation of the intracellular free calcium concentration in rat sympathetic neurons in vitro. Mol. Pharm. 1988, 34, 664–673. [Google Scholar]
- Usachev, Y.; Shmigol, A.; Pronchuk, N.; Kostyuk, P.; Verkhratsky, A. Caffeine-induced calcium release from internal stores in cultured rat sensory neurons. Neuroscience 1993, 57, 845–859. [Google Scholar] [CrossRef]
- Ludwig, M.; Sabatier, N.; Bull, P.M.; Landgraf, R.; Dayanithi, G.; Leng, G. Intracellular calcium stores regulate activity-dependent neuropeptide release from dendrites. Nature 2002, 418, 85–89. [Google Scholar] [CrossRef]
- Patel, J.C.; Witkovsky, P.; Avshalumov, M.V.; Rice, M.E. Mobilization of calcium from intracellular stores facilitates somatodendritic dopamine release. J. Neurosci. 2009, 29, 6568–6579. [Google Scholar] [CrossRef]
- Zerbes, M.; Clark, C.L.; Powis, D.A. Neurotransmitter release from bovine adrenal chromaffin cells is modulated by capacitative Ca(2+)entry driven by depleted internal Ca(2+)stores. Cell Calcium 2001, 29, 49–58. [Google Scholar] [CrossRef]
- Yang, L.; Fomina, A.F. Ca2+ influx and clearance at hyperpolarized membrane potentials modulate spontaneous and stimulated exocytosis in neuroendocrine cells. Cell Calcium 2020, 87, 102184. [Google Scholar] [CrossRef]
- Wotjak, C.T.; Ludwig, M.; Landgraf, R. Vasopressin facilitates its own release within the rat supraoptic nucleus in vivo. Neuroreport 1994, 5, 1181–1184. [Google Scholar] [CrossRef]
- Bao, L.; Jin, S.X.; Zhang, C.; Wang, L.H.; Xu, Z.Z.; Zhang, F.X.; Wang, L.C.; Ning, F.S.; Cai, H.J.; Guan, J.S.; et al. Activation of delta opioid receptors induces receptor insertion and neuropeptide secretion. Neuron 2003, 37, 121–133. [Google Scholar] [CrossRef] [Green Version]
- Harlow, M.L.; Ress, D.; Stoschek, A.; Marshall, R.M.; McMahan, U.J. The architecture of active zone material at the frog’s neuromuscular junction. Nature 2001, 409, 479–484. [Google Scholar] [CrossRef]
- Neher, E.; Sakaba, T. Multiple roles of calcium ions in the regulation of neurotransmitter release. Neuron 2008, 59, 861–872. [Google Scholar] [CrossRef] [Green Version]
- Nagwaney, S.; Harlow, M.L.; Jung, J.H.; Szule, J.A.; Ress, D.; Xu, J.; Marshall, R.M.; McMahan, U.J. Macromolecular connections of active zone material to docked synaptic vesicles and presynaptic membrane at neuromuscular junctions of mouse. J. Comp. Neurol. 2009, 513, 457–468. [Google Scholar] [CrossRef] [Green Version]
- Szule, J.A.; Harlow, M.L.; Jung, J.H.; De-Miguel, F.F.; Marshall, R.M.; McMahan, U.J. Regulation of synaptic vesicle docking by different classes of macromolecules in active zone material. PLoS ONE 2012, 7, e33333. [Google Scholar]
- Südhof, T.C. Neurotransmitter release: The last millisecond in the life of a synaptic vesicle. Neuron 2013, 80, 675–690. [Google Scholar] [CrossRef] [Green Version]
- Pang, Z.P.; Melicoff, E.; Padgett, D.; Liu, Y.; Teich, A.F.; Dickey, B.F.; Lin, W.; Adachi, R.; Südhof, T.C. Synaptotagmin-2 is essential for survival and contributes to Ca2+ triggering of neurotransmitter release in central and neuromuscular synapses. J. Neurosci. 2006, 26, 13493–13504. [Google Scholar] [CrossRef]
- Witkovsky, P.; Patel, J.C.; Lee, C.R.; Rice, M.E. Immunocytochemical identification of proteins involved in dopamine release from the somatodendritic compartment of nigral dopaminergic neurons. Neuroscience 2009, 164, 488–496. [Google Scholar] [CrossRef] [Green Version]
- Mendez, J.A.; Bourque, M.J.; Fasano, C.; Kortleven, C.; Trudeau, L.E. Somatodendritic dopamine release requires synaptotagmin 4 and 7 and the participation of voltage-gated calcium channels. J. Biol. Chem. 2011, 286, 23928–23937. [Google Scholar] [CrossRef] [Green Version]
- Noguez, P.; Rubí, J.M.; De-Miguel, F.F. Thermodynamic Efficiency of Somatic Exocytosis of Serotonin. Front. Physiol. 2019, 10, 473. [Google Scholar] [CrossRef] [PubMed]
- Vitale, M.L.; Seward, E.P.; Trifaró, J.M. Chromaffin cell cortical actin network dynamics control the size of the release-ready vesicle pool and the initial rate of exocytosis. Neuron 1995, 14, 353–363. [Google Scholar] [CrossRef] [Green Version]
- Oheim, M.; Stühmer, W. Tracking chromaffin granules on their way through the actin cortex. Eur. Biophys. J. 2000, 29, 67–89. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.F.; Hatton, G.I. Mechanisms underlying oxytocin-induced excitation of supraoptic neurons: Prostaglandin mediation of actin polymerization. J. Neurophysiol. 2006, 95, 3933–3947. [Google Scholar] [CrossRef] [Green Version]
- Torregrosa-Hetland, C.J.; Villanueva, J.; Giner, D.; Lopez-Font, I.; Nadal, A.; Quesada, I.; Viniegra, S.; Expósito-Romero, G.; Gil, A.; Gonzalez-Velez, V.; et al. The F-actin cortical network is a major factor influencing the organization of the secretory machinery in chromaffin cells. J. Cell Sci. 2011, 124, 727–734. [Google Scholar] [CrossRef] [Green Version]
- Trifaró, J.M.; Lejen, T.; Rosé, S.D.; Pene, T.D.; Barkar, N.D.; Seward, E.P. Pathways that control cortical F-actin dynamics during secretion. Neurochem. Res. 2002, 27, 1371–1385. [Google Scholar] [CrossRef]
- Villanueva, J.; Gimenez-Molina, Y.; Viniegra, S.; Gutiérrez, L.M. F-actin cytoskeleton and the fate of organelles in chromaffin cells. J. Neurochem. 2016, 137, 860–866. [Google Scholar] [CrossRef] [Green Version]
- Lervik, A.; Bresme, F.; Kjelstrup, S.; Rubí, J.M. On the thermodynamic efficiency of Ca2+-ATPase molecular machines. Biophys. J. 2012, 103, 1218–1226. [Google Scholar] [CrossRef] [Green Version]












Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De-Miguel, F.F. The Thermodynamically Expensive Contribution of Three Calcium Sources to Somatic Release of Serotonin. Int. J. Mol. Sci. 2022, 23, 1495. https://doi.org/10.3390/ijms23031495
De-Miguel FF. The Thermodynamically Expensive Contribution of Three Calcium Sources to Somatic Release of Serotonin. International Journal of Molecular Sciences. 2022; 23(3):1495. https://doi.org/10.3390/ijms23031495
Chicago/Turabian StyleDe-Miguel, Francisco F. 2022. "The Thermodynamically Expensive Contribution of Three Calcium Sources to Somatic Release of Serotonin" International Journal of Molecular Sciences 23, no. 3: 1495. https://doi.org/10.3390/ijms23031495