
Changes in the Phytochemical Profile and Antioxidant Properties of Prunus persica Fruits after the Application of a Commercial Biostimulant Based on Seaweed and Yeast Extract

 , , and
, , and
Abstract
:1. Introduction
2. Results and Discussion
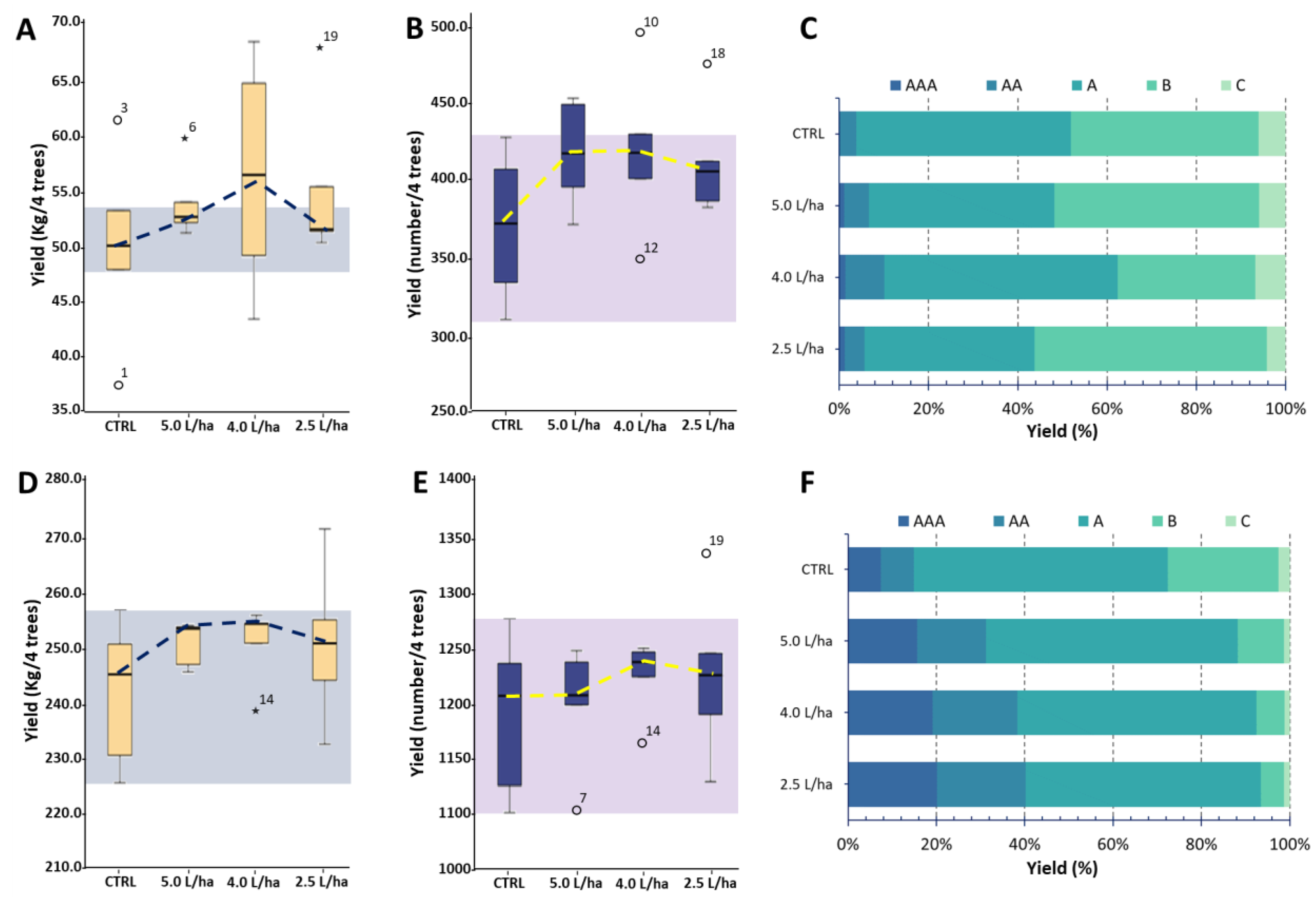
2.1. Biostimulant Application Contributes to Producing Homogeneous Yields and Improving Fruit Marketability
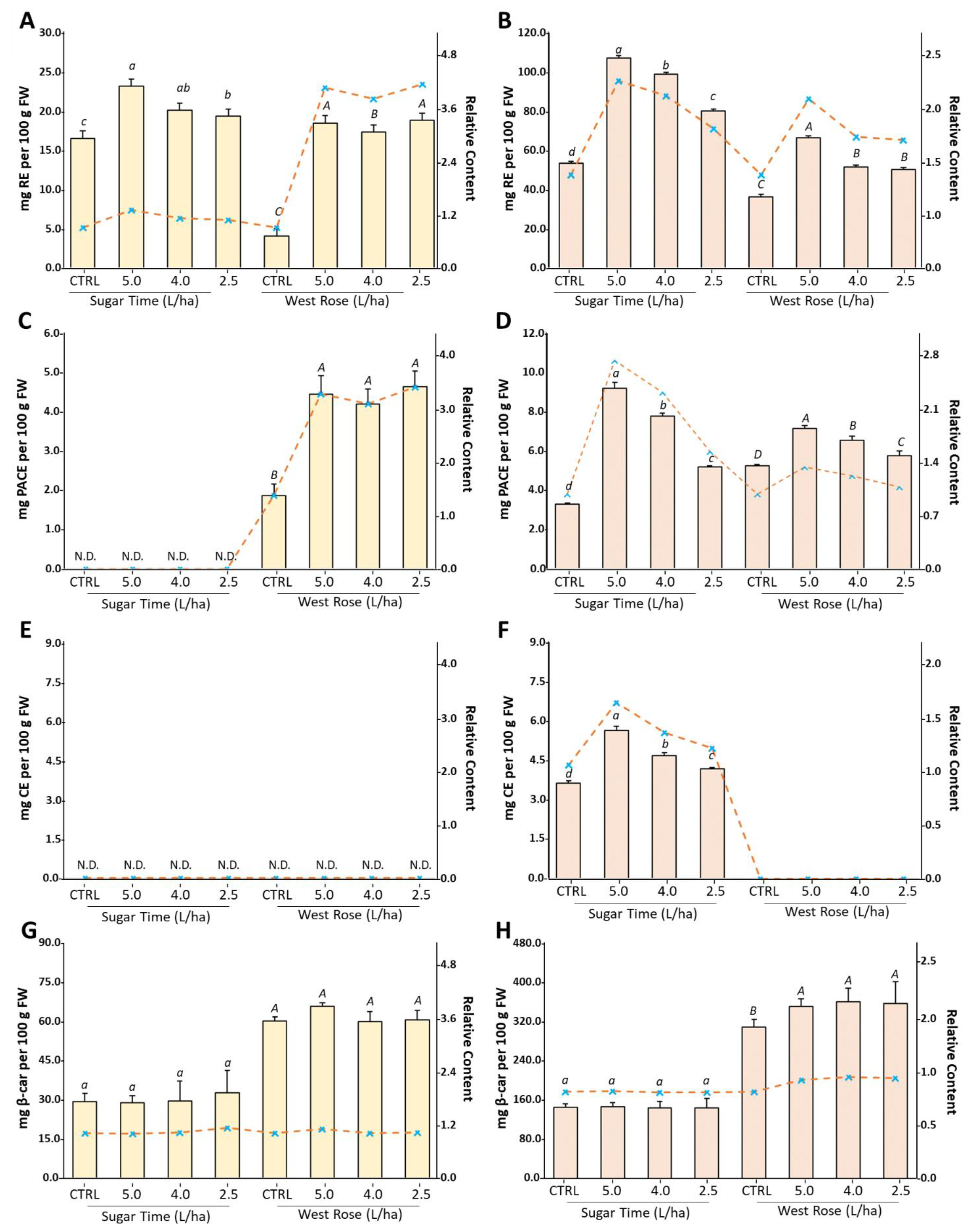
2.2. UV/Vis Screening Assays Suggest an Increase in Phytochemicals in Peach Fruits
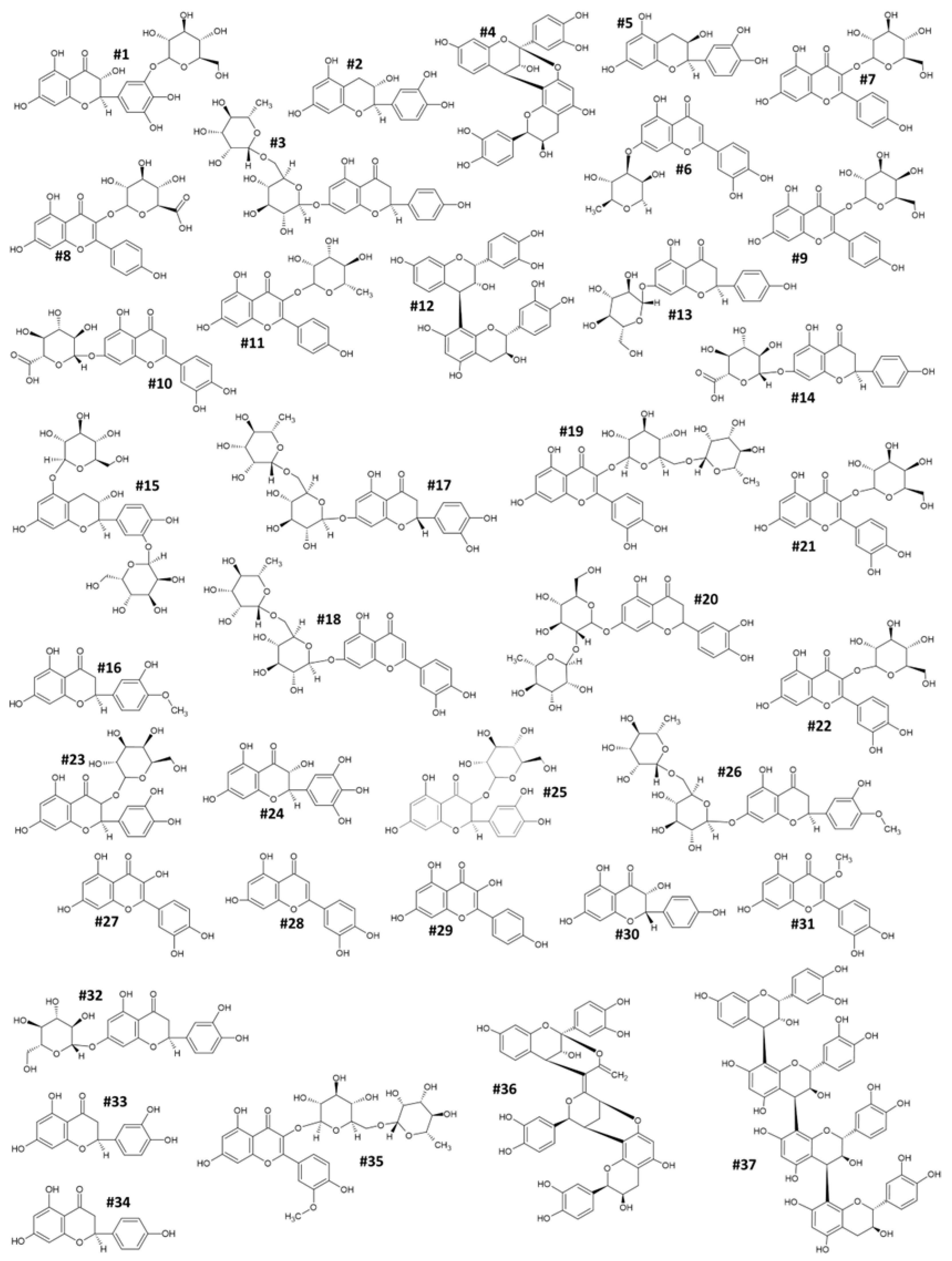
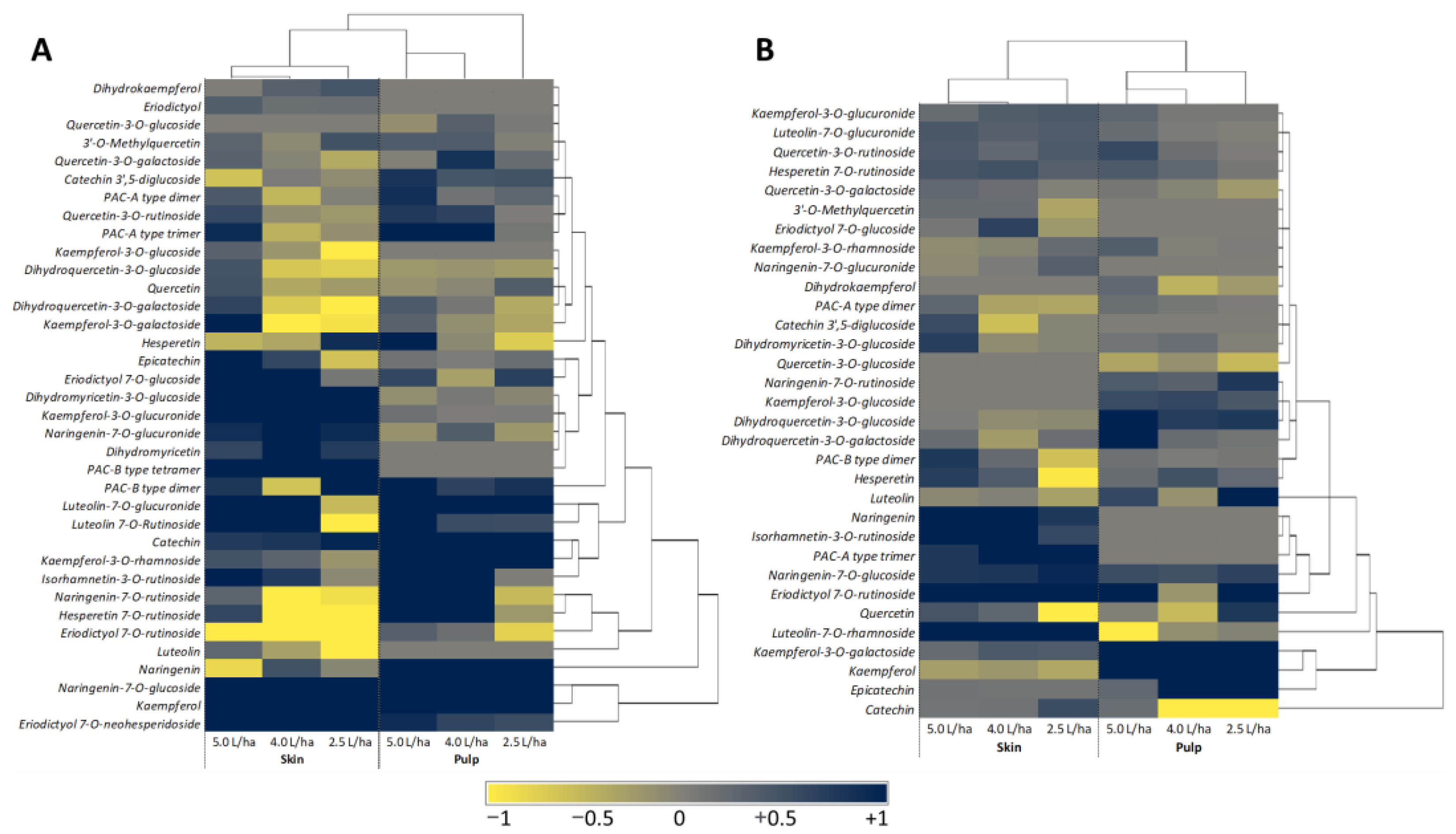
2.3. HPLC-DAD-MS/MS Analysis Identified Bioactive Compounds Responsible for the Nutraceutical Boost
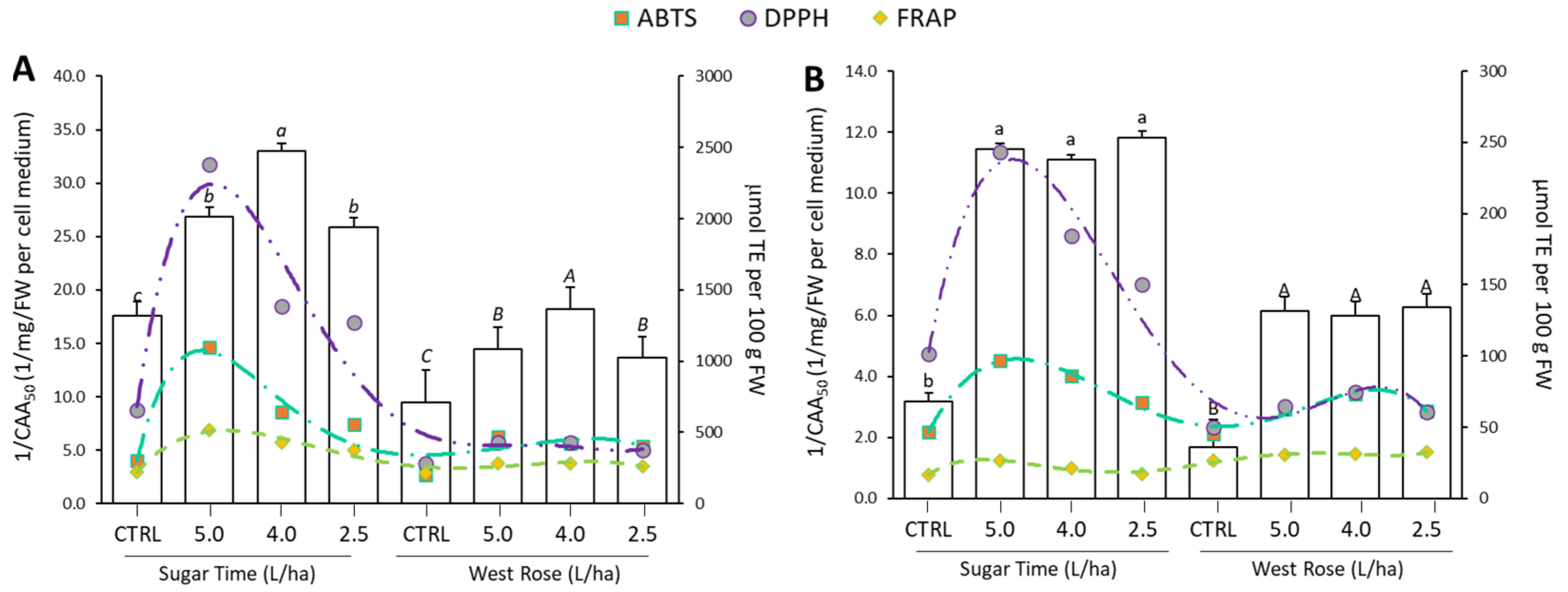
2.4. The Antioxidant Properties Were Enhanced by the Biostimulant Treatment
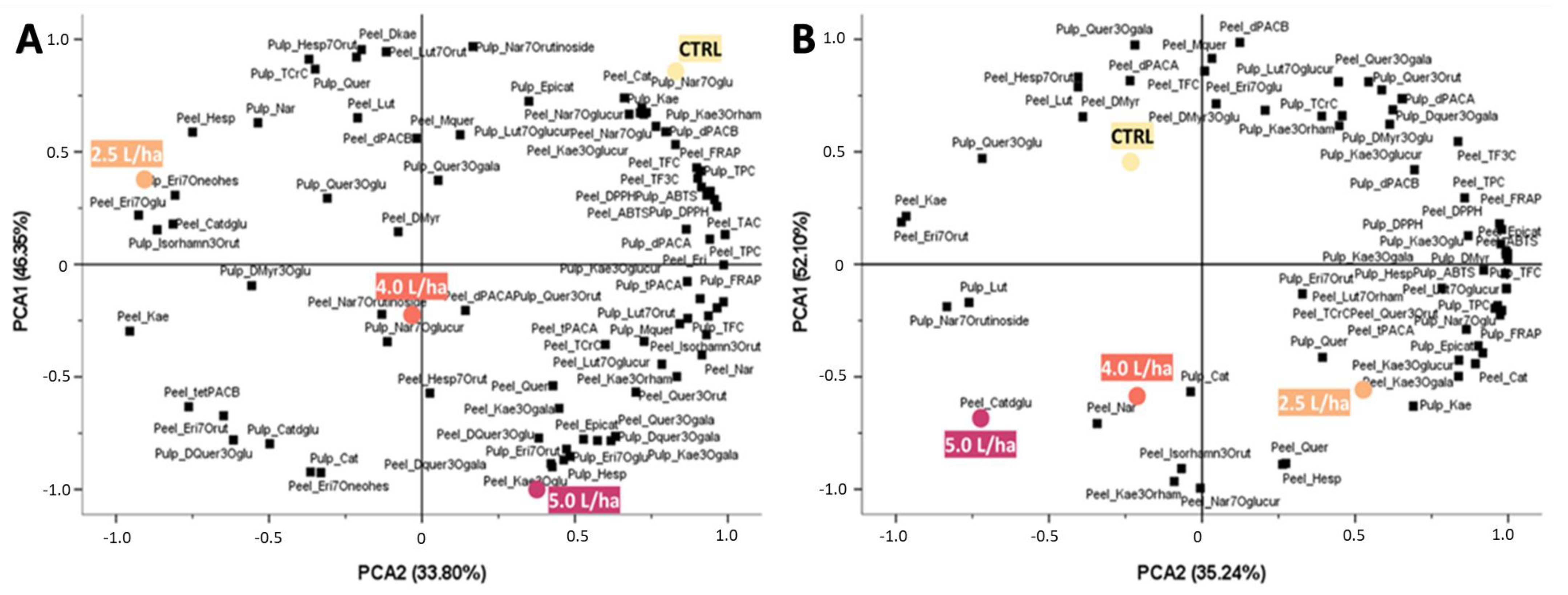
2.5. PCA Revealed a Statistical Change in the Nutraceutical Profile of the Fruits
3. Materials and Methods
3.1. Site Location and Description
3.2. Biostimulant Formulation and Trial Design
3.3. Preparation of Hydroalcoholic Extracts
3.4. Polyphenol Determination
3.5. Flavonoid Determination
3.6. Flavan-3-ol Determination
3.7. Anthocyanin Determination
3.8. Carotenoid Determination
3.9. Identification of Bioactive Compounds via HPLC-ESI-MS/MS
3.10. Evaluation of Antioxidant Properties
3.10.1. 2,2′-Azino-bis(3-ethylbenzothiazoline-6-sulphonic Acid) (ABTS) Assay
3.10.2. 2,2-Diphenyl-1-picrylhydrazyl (DPPH) Assay
3.10.3. Ferric Reducing Antioxidant Power (FRAP) Assay
3.10.4. Cellular Antioxidant Assay (CAA)
3.11. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Farina, V.; Tinebra, I.; Perrone, A.; Sortino, G.; Palazzolo, E.; Mannino, G.; Gentile, C. Physicochemical, nutraceutical and sensory traits of six papaya (Carica papaya L.) cultivars grown in greenhouse conditions in the Mediterranean climate. Agronomy 2020, 10, 501. [Google Scholar] [CrossRef] [Green Version]
- Kwasny, T.; Dobernig, K.; Riefler, P. Towards reduced meat consumption: A systematic literature review of intervention effectiveness, 2001–2019. Appetite 2022, 168, 105739. [Google Scholar] [CrossRef] [PubMed]
- Mannino, G.; Serio, G.; Bertea, C.M.; Chiarelli, R.; Lauria, A.; Gentile, C. Phytochemical profile and antioxidant properties of the edible and non-edible portions of black sapote (Diospyros digyna Jacq.). Food Chem. 2022, 380, 132137. [Google Scholar] [CrossRef]
- Luca, S.V.; Macovei, I.; Bujor, A.; Miron, A.; Skalicka-Woźniak, K.; Aprotosoaie, A.C.; Trifan, A. Bioactivity of dietary polyphenols: The role of metabolites. Crit. Rev. Food Sci. Nutr. 2020, 60, 626–659. [Google Scholar] [CrossRef] [PubMed]
- Vigliante, I.; Mannino, G.; Maffei, M.E. OxiCyan®, a phytocomplex of bilberry (Vaccinium myrtillus) and spirulina (Spirulina platensis), exerts both direct antioxidant activity and modulation of ARE/Nrf2 pathway in HepG2 cells. J. Funct. Foods 2019, 61, 103508. [Google Scholar] [CrossRef]
- Mannino, G.; Iovino, P.; Lauria, A.; Genova, T.; Asteggiano, A.; Notarbartolo, M.; Porcu, A.; Serio, G.; Chinigò, G.; Occhipinti, A.; et al. Bioactive Triterpenes of Protium heptaphyllum Gum Resin Extract Display Cholesterol-Lowering Potential. Int. J. Mol. Sci. 2021, 22, 2664. [Google Scholar] [CrossRef]
- Caradonna, F.; Consiglio, O.; Luparello, C.; Gentile, C. Science and healthy meals in the world: Nutritional epigenomics and nutrigenetics of the mediterranean diet. Nutrients 2020, 12, 1748. [Google Scholar] [CrossRef]
- Mason-D’Croz, D.; Bogard, J.R.; Sulser, T.B.; Cenacchi, N.; Dunston, S.; Herrero, M.; Wiebe, K. Gaps between fruit and vegetable production, demand, and recommended consumption at global and national levels: An integrated modelling study. Lancet Planet. Health 2019, 3, e318–e329. [Google Scholar] [CrossRef] [Green Version]
- Organization, W.H. Healthy Diet; World Health Organization Regional Office for the Eastern Mediterranean: Cairo, Egypt, 2019. [Google Scholar]
- Jacob-Lopes, E.; Maroneze, M.M.; Deprá, M.C.; Sartori, R.B.; Dias, R.R.; Zepka, L.Q. Bioactive food compounds from microalgae: An innovative framework on industrial biorefineries. Curr. Opin. Food Sci. 2019, 25, 1–7. [Google Scholar] [CrossRef]
- Boccia, F.; Covino, D.; Sarnacchiaro, P. Genetically modified food versus knowledge and fear: A Noumenic approach for consumer behaviour. Food Res. Int. 2018, 111, 682–688. [Google Scholar] [CrossRef]
- Rizvi, R.F.; Wang, Y.; Nguyen, T.; Vasilakes, J.; Bian, J.; He, Z.; Zhang, R. Analyzing social media data to understand consumer information needs on dietary supplements. Stud. Health Technol. Inform. 2019, 264, 323. [Google Scholar] [PubMed]
- Homer, P.M.; Mukherjee, S. The impact of dietary supplement form and dosage on perceived efficacy. J. Consum. Mark. 2018, 35, 228–238. [Google Scholar] [CrossRef]
- Boon, H.; Bozinovski, N. A systematic narrative review of the evidence for labeling of natural health products and dietary supplements. J. Altern. Complement. Med. 2019, 25, 777–788. [Google Scholar] [CrossRef] [PubMed]
- Majeed, A. Application of agrochemicals in agriculture: Benefits, risks and responsibility of stakeholders. J. Food Sci. Toxicol. 2018, 2, 3. [Google Scholar]
- Srivastav, A.L. Chemical fertilizers and pesticides: Role in groundwater contamination. In Agrochemicals Detection, Treatment and Remediation; Elsevier: Amsterdam, The Netherlands, 2020; pp. 143–159. [Google Scholar]
- Pandey, S.; Joshi, N.; Kumar, M. Agrochemicals and human well-being: A review in context of Indian agriculture. Int. J. Cosmet. Sci. 2020, 8, 1539–1543. [Google Scholar] [CrossRef] [Green Version]
- Aleinik, S.N.; Dorofeev, A.F.; Akinchin, A.V.; Linkov, S.A.; Melentiev, A.A. Agriculture development in the context of technological and ecology problems. J. Crit. Rev. 2020, 7, 2174–2182. [Google Scholar]
- Sarker, M.N.I.; Ahmads, M.S.; Islam, M.S.; Syed, M.M.M.A.; Memon, N.H. Potential food safety risk in fruit production from the extensive use of fluorine-containing agrochemicals. Fluoride 2020, 53, 499–520. [Google Scholar]
- Mapelli, F.; Carullo, D.; Farris, S.; Ferrante, A.; Bacenetti, J.; Ventura, V.; Frisio, D.; Borin, S. Food waste-derived biomaterials enriched by biostimulant agents for sustainable horticultural practices: A possible circular solution. Front. Sustain. 2022, 3, 1–7. [Google Scholar] [CrossRef]
- Montoneri, E.; Baglieri, A.; Fascella, G. Biostimulant effects of waste derived biobased products in the cultivation of ornamental and food plants. Agriculture 2022, 12, 994. [Google Scholar] [CrossRef]
- Puglisi, I.; Barone, V.; Sidella, S.; Coppa, M.; Broccanello, C.; Gennari, M.; Baglieri, A. Biostimulant activity of humic-like substances from agro-industrial waste on Chlorella vulgaris and Scenedesmus quadricauda. Eur. J. Phycol. 2018, 53, 433–442. [Google Scholar] [CrossRef]
- Kaur, M.; Bhari, R.; Singh, R.S. Chicken feather waste-derived protein hydrolysate as a potential biostimulant for cultivation of mung beans. Biologia 2021, 76, 1807–1815. [Google Scholar] [CrossRef]
- Mironenko, G.A.; Zagorskii, I.A.; Bystrova, N.A.; Kochetkov, K.A. The Effect of a Biostimulant Based on a Protein Hydrolysate of Rainbow Trout (Oncorhynchus mykiss) on the Growth and Yield of Wheat (Triticum aestivum L.). Molecules 2022, 27, 6663. [Google Scholar] [CrossRef] [PubMed]
- Castiglione, A.M.; Mannino, G.; Contartese, V.; Bertea, C.M.; Ertani, A. Microbial Biostimulants as Response to Modern Agriculture Needs: Composition, Role and Application of These Innovative Products. Plants 2021, 10, 1533. [Google Scholar] [CrossRef] [PubMed]
- Campobenedetto, C.; Agliassa, C.; Mannino, G.; Vigliante, I.; Contartese, V.; Secchi, F.; Bertea, C.M. A biostimulant based on seaweed (Ascophyllum nodosum and Laminaria digitata) and yeast extracts mitigates water stress effects on tomato (Solanum lycopersicum L.). Agriculture 2021, 11, 557. [Google Scholar] [CrossRef]
- Agliassa, C.; Mannino, G.; Molino, D.; Cavalletto, S.; Contartese, V.; Bertea, C.M.; Secchi, F. A new protein hydrolysate-based biostimulant applied by fertigation promotes relief from drought stress in Capsicum annuum L. Plant Physiol. Biochem. 2021, 166, 1076–1086. [Google Scholar] [CrossRef]
- Bulgari, R.; Franzoni, G.; Ferrante, A. Biostimulants Application in Horticultural Crops under Abiotic Stress Conditions. Agronomy 2019, 9, 306. [Google Scholar] [CrossRef] [Green Version]
- Pardo-García, A.I.; Martínez-Gil, A.M.; Cadahía, E.; Pardo, F.; Alonso, G.L.; Salinas, M.R. Oak extract application to grapevines as a plant biostimulant to increase wine polyphenols. Food Res. Int. 2014, 55, 150–160. [Google Scholar] [CrossRef]
- Zarzecka, K.; Gugała, M.; Sikorska, A.; Mystkowska, I.; Baranowska, A.; Niewęgłowski, M.; Dołęga, H. The effect of herbicides and biostimulants on polyphenol content of potato (Solanum tuberosum L.) tubers and leaves. J. Saudi Soc. Agric. Sci. 2019, 18, 102–106. [Google Scholar] [CrossRef]
- Mannino, G.; Campobenedetto, C.; Vigliante, I.; Contartese, V.; Gentile, C.; Bertea, C.M. The application of a plant biostimulant based on seaweed and yeast extract improved tomato fruit development and quality. Biomolecules 2020, 10, 1662. [Google Scholar] [CrossRef]
- Mystkowska, I.; Zarzecka, K.; Gugała, M.; Sikorska, A. The Polyphenol Content in Three Edible Potato Cultivars Depending on the Biostimulants Used. Agriculture 2020, 10, 269. [Google Scholar] [CrossRef]
- Ziosi, V.; Giovannetti, G.; Vitali, F.; Di Nardo, A.; Porro, D. SUNRED®, a botanical extract-based biostimulant, enhances polyphenols accumulation and improves quality of musts. In Proceedings of the I World Congress on the Use of Biostimulants in Agriculture 1009, Strassbourg, France, 26–29 November 2012; pp. 67–70. [Google Scholar]
- Montesano, V.; Negro, D.; Sonnante, G.; Laghetti, G.; Urbano, M. Polyphenolic Compound Variation in Globe Artichoke Cultivars as Affected by Fertilization and Biostimulants Application. Plants 2022, 11, 2067. [Google Scholar] [CrossRef] [PubMed]
- Bartucca, M.L.; Cerri, M.; Del Buono, D.; Forni, C. Use of biostimulants as a new approach for the improvement of phytoremediation performance—A Review. Plants 2022, 11, 1946. [Google Scholar] [CrossRef] [PubMed]
- Manganaris, G.A.; Minas, I.; Cirilli, M.; Torres, R.; Bassi, D.; Costa, G. Peach for the future: A specialty crop revisited. Sci. Hortic. 2022, 305, 111390. [Google Scholar] [CrossRef]
- Anthony, B.M.; Minas, I.S. Redefining the impact of preharvest factors on peach fruit quality development and metabolism: A review. Sci. Hortic. 2022, 297, 110919. [Google Scholar] [CrossRef]
- Rudke, C.R.M.; Zielinski, A.A.F.; Ferreira, S.R.S. From Biorefinery to Food Product Design: Peach (Prunus persica) By-Products Deserve Attention. Food Bioprocess Technol. 2022, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Lamichhane, J.R.; Debaeke, P.; Steinberg, C.; You, M.P.; Barbetti, M.J.; Aubertot, J.-N. Abiotic and biotic factors affecting crop seed germination and seedling emergence: A conceptual framework. Plant Soil 2018, 432, 1–28. [Google Scholar] [CrossRef]
- Matthews, S.; Ali, A.; Siddiqui, Y.; Supramaniam, C.V. Plant Bio-stimulant: Prospective, Safe and Natural Resources. J. Soil Sci. Plant Nutr. 2022, 22, 2570–2586. [Google Scholar] [CrossRef]
- Ali, O.; Ramsubhag, A.; Jayaraman, J. Biostimulant properties of seaweed extracts in plants: Implications towards sustainable crop production. Plants 2021, 10, 531. [Google Scholar] [CrossRef]
- El Boukhari, M.E.M.; Barakate, M.; Bouhia, Y.; Lyamlouli, K. Trends in seaweed extract based biostimulants: Manufacturing process and beneficial effect on soil-plant systems. Plants 2020, 9, 359. [Google Scholar] [CrossRef] [Green Version]
- Tarantino, A.; Lops, F.; Disciglio, G.; Lopriore, G. Effects of plant biostimulants on fruit set, growth, yield and fruit quality attributes of ‘Orange rubis®’ apricot (Prunus armeniaca L.) cultivar in two consecutive years. Sci. Hortic. 2018, 239, 26–34. [Google Scholar] [CrossRef]
- Chouliaras, V.; Tasioula, M.; Chatzissavvidis, C.; Therios, I.; Tsabolatidou, E. The effects of a seaweed extract in addition to nitrogen and boron fertilization on productivity, fruit maturation, leaf nutritional status and oil quality of the olive (Olea europaea L.) cultivar Koroneiki. J. Sci. Food Agric. 2009, 89, 984–988. [Google Scholar] [CrossRef]
- Chouliaras, V.; Gerascapoulos, D.; Lionakis, S. Effects of seaweed extract on fruit growth, weight and maturation of’Hayward’kiwifruit. In Proceedings of the III International Symposium on Kiwifruit 444, Thessalonaki, Greece, 19–22 September 1995; pp. 485–492. [Google Scholar]
- Fornes, F.; Sanchez-Perales, M.; Guardiola, J.L. Effect of a seaweed extract on the productivity of’de Nules’ clementine mandarin and navelina orange. Bot. Mar. 2002, 45, 486–489. [Google Scholar] [CrossRef]
- Taskos, D.; Stamatiadis, S.; Yvin, J.-C.; Jamois, F. Effects of an Ascophyllum nodosum (L.) Le Jol. extract on grapevine yield and berry composition of a Merlot vineyard. Sci. Hortic. 2019, 250, 27–32. [Google Scholar] [CrossRef]
- Sabir, A.; Yazar, K.; Sabir, F.; Kara, Z.; Yazici, M.A.; Goksu, N. Vine growth, yield, berry quality attributes and leaf nutrient content of grapevines as influenced by seaweed extract (Ascophyllum nodosum) and nanosize fertilizer pulverizations. Sci. Hortic. 2014, 175, 1–8. [Google Scholar] [CrossRef]
- Khan, A.S.; Ahmad, B.; Jaskani, M.J.; Ahmad, R.; Malik, A.U. cal. Int. J. Agric. Biol 2012, 14, 383–388. [Google Scholar]
- Naqash, S.; Naik, H. Peach Wastes and By-Products: Chemistry, Processing, and Utilization. In Handbook of Fruit Wastes and By-Products; CRC Press: Boca Raton, FL, USA, 2022; pp. 161–174. ISBN 1003164463. [Google Scholar]
- Basile, B.; Rouphael, Y.; Colla, G.; Soppelsa, S.; Andreotti, C. Appraisal of emerging crop management opportunities in fruit trees, grapevines and berry crops facilitated by the application of biostimulants. Sci. Hortic. 2020, 267, 109330. [Google Scholar] [CrossRef]
- Graziani, G.; Ritieni, A.; Cirillo, A.; Cice, D.; Di Vaio, C. Effects of biostimulants on annurca fruit quality and potential nutraceutical compounds at harvest and during storage. Plants 2020, 9, 775. [Google Scholar] [CrossRef]
- Ganugi, P.; Martinelli, E.; Lucini, L. Microbial biostimulants as a sustainable approach to improve the functional quality in plant-based foods: A review. Curr. Opin. Food Sci. 2021, 41, 217–223. [Google Scholar] [CrossRef]
- Mannino, G.; Gentile, C.; Porcu, A.; Agliassa, C.; Caradonna, F.; Bertea, C.M. Chemical Profile and Biological Activity of Cherimoya (Annona cherimola Mill.) and Atemoya (Annona atemoya) Leaves. Molecules 2020, 25, 2612. [Google Scholar] [CrossRef]
- Rahman, M.; Rahman, M.; Sabir, A.A.; Mukta, J.A.; Khan, M.M.A.; Mohi-Ud-Din, M.; Miah, M.G.; Islam, M.T. Plant probiotic bacteria Bacillus and Paraburkholderia improve growth, yield and content of antioxidants in strawberry fruit. Sci. Rep. 2018, 8, 2504. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Seco, D.; Zhang, Y.; Gutierrez-Mañero, F.J.; Martin, C.; Ramos-Solano, B. Application of Pseudomonas fluorescens to blackberry under field conditions improves fruit quality by modifying flavonoid metabolism. PLoS ONE 2015, 10, e0142639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagy, P.T.; Csihon, Á.; Szabó, A. Effects of Algae Products on Nutrient Uptake and Fruit Quality of Apple. Nat. Resour. Sustain. Dev. 2019, 9, 80–91. [Google Scholar] [CrossRef]
- Mannino, G.; Chinigò, G.; Serio, G.; Genova, T.; Gentile, C.; Munaron, L.; Bertea, C.M. Proanthocyanidins and Where to Find Them: A Meta-Analytic Approach to Investigate Their Chemistry, Biosynthesis, Distribution and Effect on Human Health. Antioxidants 2021, 10, 1229. [Google Scholar] [CrossRef] [PubMed]
- Mannino, G.; Gentile, C.; Ertani, A.; Serio, G.; Bertea, C.M. Anthocyanins: Biosynthesis, Distribution, Ecological Role, and Use of Biostimulants to Increase Their Content in Plant Foods—A Review. Agriculture 2021, 11, 212. [Google Scholar] [CrossRef]
- Krga, I.; Milenkovic, D. Anthocyanins: From sources and bioavailability to cardiovascular-health benefits and molecular mechanisms of action. J. Agric. Food Chem. 2019, 67, 1771–1783. [Google Scholar] [CrossRef]
- Chaves-Silva, S.; Dos Santos, A.L.; Chalfun-Júnior, A.; Zhao, J.; Peres, L.E.P.; Benedito, V.A. Understanding the genetic regulation of anthocyanin biosynthesis in plants–tools for breeding purple varieties of fruits and vegetables. Phytochemistry 2018, 153, 11–27. [Google Scholar] [CrossRef]
- Frioni, T.; Sabbatini, P.; Tombesi, S.; Norrie, J.; Poni, S.; Gatti, M.; Palliotti, A. Effects of a biostimulant derived from the brown seaweed Ascophyllum nodosum on ripening dynamics and fruit quality of grapevines. Sci. Hortic. 2018, 232, 97–106. [Google Scholar] [CrossRef]
- Todeschini, V.; Aitlahmidi, N.; Mazzucco, E.; Marsano, F.; Gosetti, F.; Robotti, E.; Bona, E.; Massa, N.; Bonneau, L.; Marengo, E.; et al. Impact of beneficial microorganisms on strawberry growth, fruit production, nutritional quality, and volatilome. Front. Plant Sci. 2018, 9, 1611. [Google Scholar] [CrossRef]
- Vigliante, I.; Mannino, G.; Maffei, M.E. Chemical Characterization and DNA Fingerprinting of Griffonia simplicifolia Baill. Molecules 2019, 24, 1032. [Google Scholar] [CrossRef] [Green Version]
- Mannino, G.; Pernici, C.; Serio, G.; Gentile, C.; Bertea, C.M. Melatonin and phytomelatonin: Chemistry, biosynthesis, metabolism, distribution and bioactivity in plants and animals—An overview. Int. J. Mol. Sci. 2021, 22, 9996. [Google Scholar] [CrossRef]
- Tomás-Barberán, F.A.; Gil, M.I.; Cremin, P.; Waterhouse, A.L.; Hess-Pierce, B.; Kader, A.A. HPLC-DAD-ESIMS analysis of phenolic compounds in nectarines, peaches, and plums. J. Agric. Food Chem. 2001, 49, 4748–4760. [Google Scholar] [CrossRef] [PubMed]
- Di Vaio, C.; Marallo, N.; Graziani, G.; Ritieni, A.; Di Matteo, A. Evaluation of fruit quality, bioactive compounds and total antioxidant activity of flat peach cultivars. J. Sci. Food Agric. 2015, 95, 2124–2131. [Google Scholar] [CrossRef] [PubMed]
- Campbell, O.E.; Padilla-Zakour, O.I. Phenolic and carotenoid composition of canned peaches (Prunus persica) and apricots (Prunus armeniaca) as affected by variety and peeling. Food Res. Int. 2013, 54, 448–455. [Google Scholar] [CrossRef]
- Sidhu, V.; Nandwani, D.; Wang, L.; Wu, Y. A study on organic tomatoes: Effect of a biostimulator on phytochemical and antioxidant activities. J. Food Qual. 2017, 2017, 5020742. [Google Scholar] [CrossRef]
- Pourhadi, M.; Badi, H.N.; Mehrafarin, A.; Omidi, H.; Hajiaghaee, R. Phytochemical and growth responses of Mentha piperita to foliar application of biostimulants under greenhouse and field conditions. Herba Pol. 2018, 64, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Parađiković, N.; Vinković, T.; Vinković Vrček, I.; Žuntar, I.; Bojić, M.; Medić-Šarić, M. Effect of natural biostimulants on yield and nutritional quality: An example of sweet yellow pepper (Capsicum annuum L.) plants. J. Sci. Food Agric. 2011, 91, 2146–2152. [Google Scholar] [CrossRef]
- Lu, W.; Shi, Y.; Wang, R.; Su, D.; Tang, M.; Liu, Y.; Li, Z. Antioxidant activity and healthy benefits of natural pigments in fruits: A review. Int. J. Mol. Sci. 2021, 22, 4945. [Google Scholar] [CrossRef]
- García-Sánchez, F.; Simón-Grao, S.; Navarro-Pérez, V.; Alfosea-Simón, M. Scientific Advances in Biostimulation Reported in the 5th Biostimulant World Congress. Horticulturaeorticulturae 2022, 8, 665. [Google Scholar] [CrossRef]
- Kellett, M.E.; Greenspan, P.; Pegg, R.B. Modification of the cellular antioxidant activity (CAA) assay to study phenolic antioxidants in a Caco-2 cell line. Food Chem. 2018, 244, 359–363. [Google Scholar] [CrossRef]
- Liu, G.; Zhu, W.; Zhang, J.; Song, D.; Zhuang, L.; Ma, Q.; Yang, X.; Liu, X.; Zhang, J.; Zhang, H. Antioxidant capacity of phenolic compounds separated from tea seed oil in vitro and in vivo. Food Chem. 2022, 371, 131122. [Google Scholar] [CrossRef]
- Martinelli, E.; Granato, D.; Azevedo, L.; Gonçalves, J.E.; Lorenzo, J.M.; Munekata, P.E.S.; Simal-Gandara, J.; Barba, F.J.; Carrillo, C.; Rajoka, M.S.R. Current perspectives in cell-based approaches towards the definition of the antioxidant activity in food. Trends Food Sci. Technol. 2021, 116, 232–243. [Google Scholar] [CrossRef]
- Xiao, F.; Xu, T.; Lu, B.; Liu, R. Guidelines for antioxidant assays for food components. Food Front. 2020, 1, 60–69. [Google Scholar] [CrossRef] [Green Version]
- Saeed, I.; Guo, X.; Azeem, M.; Elshikh, M.S.; Zainab, B.; Ayaz, Z.; You, L.; Alwahibi, M.S.; Abbasi, A.M. Comparative assessment of polyphenolics’ content, free radicals’ scavenging and cellular antioxidant potential in apricot fruit. J. King Saud Univ. 2021, 33, 101459. [Google Scholar] [CrossRef]
- Mannino, G.; Gentile, C.; Maffei, M.E. Chemical partitioning and DNA fingerprinting of some pistachio (Pistacia vera L.) varieties of different geographical origin. Phytochemistry 2019, 160, 40–47. [Google Scholar] [CrossRef]
- Prior, R.L.; Fan, E.; Ji, H.; Howell, A.; Nio, C.; Paynef, M.J.; Reed, J. Multi-laboratory validation of a standard method for quantifying proanthocyanidins in cranberry powders. J. Sci. Food Agric. 2010, 90, 1473–1478. [Google Scholar] [CrossRef]
- Granato, D.; Shahidi, F.; Wrolstad, R.; Kilmartin, P.; Melton, L.D.; Hidalgo, F.J.; Miyashita, K.; van Camp, J.; Alasalvar, C.; Ismail, A.B. Antioxidant activity, total phenolics and flavonoids contents: Should we ban in vitro screening methods? Food Chem. 2018, 264, 471–475. [Google Scholar] [CrossRef]
- Gentile, C.; Mannino, G.; Palazzolo, E.; Gianguzzi, G.; Perrone, A.; Serio, G.; Farina, V. Pomological, Sensorial, Nutritional and Nutraceutical Profile of Seven Cultivars of Cherimoya (Annona cherimola Mill). Foods 2021, 10, 35. [Google Scholar] [CrossRef]
- Thaipong, K.; Boonprakob, U.; Crosby, K.; Cisneros-Zevallos, L.; Byrne, D.H. Comparison of ABTS, DPPH, FRAP, and ORAC assays for estimating antioxidant activity from guava fruit extracts. J. Food Compos. Anal. 2006, 19, 669–675. [Google Scholar] [CrossRef]
- Mannino, G.; Maffei, M.E. Metabolomics-Based Profiling, Antioxidant Power, and Uropathogenic Bacterial Anti-Adhesion Activity of SP4TM, a Formulation with a High Content of Type-A Proanthocyanidins. Antioxidants 2022, 11, 1234. [Google Scholar] [CrossRef]
- Fernandes, R.P.P.; Trindade, M.A.; Tonin, F.G.; Lima, C.G.; Pugine, S.M.P.; Munekata, P.E.S.; Lorenzo, J.M.; de Melo, M.P. Evaluation of antioxidant capacity of 13 plant extracts by three different methods: Cluster analyses applied for selection of the natural extracts with higher antioxidant capacity to replace synthetic antioxidant in lamb burgers. J. Food Sci. Technol. 2016, 53, 451–460. [Google Scholar] [CrossRef] [Green Version]
- Wolfe, K.L.; Kang, X.; He, X.; Dong, M.; Zhang, Q.; Liu, R.H. Cellular antioxidant activity of common fruits. J. Agric. Food Chem. 2008, 56, 8418–8426. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, K.L.; Liu, R.H. Cellular antioxidant activity (CAA) assay for assessing antioxidants, foods, and dietary supplements. J. Agric. Food Chem. 2007, 55, 8896–8907. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | RT | m/z | MS/MS | Chemical Formula | CAS-ID | Chemical Name |
|---|---|---|---|---|---|---|
| 1 | 12.6 | 451 | 319 | C21H22O12 | n.a. | Dihydromyricetin-3-O-glucoside |
| 2 | 13.3 | 289 | 261; 245 | C15H14O6 | 7295-85-4 | Catechin |
| 3 | 13.5 | 595 | 510.3; 426.4; 342.3; 271.6 | C27H32O14 | 14259-46-2 | Naringenin-7-O-rutinoside |
| 4 | 14.2 | 575 | 557; 439; 289 | C30H24O12 | 41743-41-3 | PAC-A type dimer |
| 5 | 14.3 | 289 | 261; 245 | C15H14O6 | 490-46-0 | Epicatechin |
| 6 | 15.8 | 431 | 395; 329; 293 | C21H20O10 | n.a. | Luteolin-7-O-rhamnoside |
| 7 | 15.8 | 447 | 400.8; 285 | C21H20O11 | 480-10-4 | Kaempferol-3-O-glucoside |
| 8 | 15.8 | 461 | 414.7; 291.7 | C21H18O12 | 22688-78-4 | Kaempferol-3-O-glucuronide |
| 9 | 16.9 | 447 | 400.8; 285 | C21H20O11 | 23627-87-4 | Kaempferol-3-O-galactoside |
| 10 | 17.4 | 461 | 414.7; 340.7; 298.7 | C21H18O12 | 29741-10-4 | Luteolin-7-O-glucuronide |
| 11 | 18.2 | 431 | 384.8; 285 | C21H20O10 | 482-39-3 | Kaempferol-3-O-rhamnoside |
| 12 | 20.2 | 577 | 531; 289 | C30H26O12 | 20315-25-7 | PAC-B type dimer |
| 13 | 22.7 | 403 | 356.7; 294.7271 | C21H22O10 | 529-55-5 | Naringenin-7-O-glucoside |
| 14 | 23.6 | 447 | 404.7; 319; 243.2 | C21H20O11 | 1237479-07-0 | Naringenin-7-O-glucuronide |
| 15 | 24.3 | 613 | 571; 553; 289 | C27H34O16 | 105330-54-9 | Catechin 3′,5-diglucoside |
| 16 | 25.0 | 300 | 239 | C16H14O6 | 520-33-2 | Hesperetin |
| 17 | 26.8 | 595 | 534.8; 287 | C27H32O15 | 13463-28-0 | Eriodictyol 7-O-rutinoside |
| 18 | 27.5 | 593 | 344.7; 285.2; 272.6 | C27H30O15 | 20633-84-5 | Luteolin 7-O-Rutinoside |
| 19 | 27.7 | 609 | 562.8; 462.7; 300.7 | C27H30O16 | 250249-75-3 | Quercetin-3-O-rutinoside |
| 20 | 27.9 | 595 | 463; 449; 287 | C27H32O15 | 13241-32-2 | Eriodictyol 7-O-neohesperidoside |
| 21 | 28.3 | 463 | 301.8 | C21H20O12 | 482-36-0 | Quercetin-3-O-galactoside |
| 22 | 28.8 | 463 | 301.7 | C21H20O12 | 482-35-9 | Quercetin-3-O-glucoside |
| 23 | 28.8 | 465 | 418.8; 396.6; 302.7 | C21H22O12 | n.a. | Dihydroquercetin-3-O-galactoside |
| 24 | 29.4 | 303 | 285; 257 | C15H12O8 | 27200-12-0 | Dihydromyricetin |
| 25 | 30.4 | 465 | 418.8; 396.6; 302.7 | C21H22O12 | 27297-45-6 | Dihydroquercetin-3-O-glucoside |
| 26 | 34.8 | 609 | 562.9; 301.4 | C28H34O15 | 520-26-3 | Hesperetin 7-O-rutinoside |
| 27 | 35.0 | 301 | 273; 273; 257 | C15H10O7 | 6151-25-3 | Quercetin |
| 28 | 37.6 | 285 | 241 | C15H10O6 | 491-70-3 | Luteolin |
| 29 | 37.8 | 285 | 269; 257 | C15H10O6 | 520-18-3 | Kaempferol |
| 30 | 37.9 | 287 | 269; 231 | C15H12O6 | 104486-98-8 | Dihydrokaempferol |
| 31 | 38.4 | 315 | 299; 271 | C16H12O7 | 480-19-3 | 3′-O-Methylquercetin |
| 32 | 39.0 | 449 | 417; 387; 319: 287.2 | C21H22O11 | 38965-51-4 | Eriodictyol 7-O-glucoside |
| 33 | 39.4 | 287 | 269; 241 | C15H12O6 | 552-58-9 | Eriodictyol |
| 34 | 45.4 | 271 | 243; 221 | C15H12O5 | 480-41-1 | Naringenin |
| 35 | 46.7 | 623 | 554.9; 528.9; 315.4 | C28H32O16 | 604-80-8 | Isorhamnetin-3-O-rutinoside |
| 36 | 54.2 | 865 | 575; 557; 439; 289 | C45H38O18 | 37064-30-5 | PAC-A type trimer |
| 37 | 56.3 | 865 | 577; 863; 531; 289 | C45H38O18 | 37064-31-6 | PAC-B type tetramer |
| No. | Peel | Pulp | ||||||
|---|---|---|---|---|---|---|---|---|
| CTRL | 5.0 L/ha | 4.0 L/ha | 2.5 L/ha | CTRL | 5.0 L/ha | 4.0 L/ha | 2.5 L/ha | |
| 1 | 3.17 ± 0.19 | 12.68 ± 0.68 | 9.49 ± 0.23 | 12.26 ± 0.28 | 8.18 ± 0.34 | 7.05 ± 0.32 | 8.65 ± 0.47 | 7.61 ± 0.39 |
| 2 | 1.48 ± 0.07 | 3.09 ± 0.07 | 3.25 ± 0.25 | 3.86 ± 0.08 | 0.37 ± 0.01 | 5.98 ± 0.17 | 2.14 ± 0.07 | 1.68 ± 0.04 |
| 3 | 7.18 ± 0.08 | 9.43 ± 0.34 | 1.32 ± 0.05 | 2.96 ± 0.15 | 1.32 ± 0.05 | 5.61 ± 0.27 | 4.25 ± 0.29 | 0.75 ± 0.04 |
| 4 | 4.84 ± 0.29 | 7.24 ± 0.41 | 2.87 ± 0.05 | 4.67 ± 0.22 | 2.58 ± 0.11 | 6.16 ± 0.21 | 2.91 ± 0.18 | 3.34 ± 0.05 |
| 5 | 2.78 ± 0.14 | 10.32 ± 0.32 | 5.1 ± 0.28 | 1.48 ± 0.05 | 1.69 ± 0.11 | 1.91 ± 0.11 | 1.73 ± 0.05 | 2.02 ± 0.04 |
| 7 | 58.45 ± 2.76 | 78.89 ± 3.43 | 47.01 ± 1.94 | 20.91 ± 0.58 | n.d. | n.d. | n.d. | n.d. |
| 8 | 3.76 ± 0.23 | 21.32 ± 0.93 | 16.29 ± 0.43 | 12.86 ± 0.81 | 8.18 ± 0.52 | 9.44 ± 0.23 | 8.31 ± 0.25 | 8.44 ± 0.15 |
| 9 | 41.09 ± 2.63 | 211.08 ± 12.14 | 14.73 ± 0.69 | 16.34 ± 0.89 | 50.8 ± 3.16 | 67.14 ± 1.49 | 43.12 ± 2.85 | 34.74 ± 1.38 |
| 10 | 14.55 ± 0.54 | 156.89 ± 8.99 | 129.21 ± 4.69 | 8.45 ± 0.29 | 9.55 ± 0.19 | 51.55 ± 1.71 | 60.69 ± 3.52 | 49.82 ± 1.26 |
| 11 | 80.55 ± 2.39 | 129.32 ± 8.98 | 107.14 ± 5.86 | 65.64 ± 4.56 | 8.83 ± 0.23 | 54.37 ± 2.63 | 56.96 ± 2.62 | 50.36 ± 2.91 |
| 12 | 224.89 ± 12.28 | 488.57 ± 13.32 | 119.13 ± 5.68 | 1762.13 ± 53.3 | 48.14 7 1.73 | 151.86 ± 6.33 | 91.23 ± 5.52 | 110.54 ± 7.62 |
| 13 | 3.2 ± 0.21 | 29.89 ± 1.83 | 32.46 ± 1.34 | 16.86 ± 0.91 | 2.75 ± 0.1 | 54.33 ± 0.94 | 26.02 ± 1.16 | 36.89 ± 0.82 |
| 14 | 3.79 ± 0.17 | 9.09 ± 0.11 | 13.01 ± 0.37 | 9.44 ± 0.21 | 2.42 ± 0.16 | 2.01 ± 0.03 | 3.46 ± 0.21 | 1.94 ± 0.12 |
| 15 | 4.21 ± 0.15 | 2.25 ± 0.06 | 4.32 ± 0.28 | 3.73 ± 0.25 | 0.79 ± 0.01 | 1.77 ± 0.11 | 1.24 ± 0.04 | 1.27 ± 0.06 |
| 16 | 5.63 ± 0.36 | 3.43 ± 0.22 | 3.89 ± 0.19 | 13.75 ± 0.56 | 2.84 ± 0.02 | 9.53 ± 0.32 | 2.55 ± 0.17 | 1.39 ± 0.08 |
| 17 | 29.83 ± 0.64 | 6.49 ± 0.39 | 5.2 ± 0.31 | 6.15 ± 0.35 | 8.87 ± 0.09 | 12.38 ± 0.19 | 10.37 ± 0.44 | 4.11 ± 0.04 |
| 18 | 2.71 ± 0.16 | 20.3 ± 0.39 | 14.01 ± 0.68 | 0.64 ± 0.01 | 0.12 ± 0.01 | 0.89 ± 0.04 | 0.22 ± 0.01 | 0.22 ± 0.01 |
| 19 | 28.57 ± 1.94 | 50.61 ± 2.69 | 24.79 ± 1.22 | 22.55 ± 1.37 | 2.42 ± 0.15 | 5.08 ± 0.06 | 4.66 ± 0.32 | 2.45 ± 0.14 |
| 20 | 0.33 ± 0.01 | 8.37 ± 0.29 | 16.95 ± 0.51 | 2.08 ± 0.02 | 0.41 ± 0.02 | 1.02 ± 0.03 | 0.77 ± 0.01 | 0.71 ± 0.04 |
| 21 | 58.29 ± 32.48 | 79.11 ± 30.37 | 55.51 ± 36.01 | 38.06 ± 7.83 | 5.98 ± 0.27 | 5.87 ± 0.36 | 13.91 ± 0.79 | 7.23 ± 0.39 |
| 22 | n.d. | n.d. | n.d. | n.d. | 7.66 ± 0.39 | 6.47 ± 0.16 | 10.45 ± 0.68 | 7.97 ± 0.22 |
| 23 | 85.28 ± 4.47 | 160.55 ± 6.98 | 42.31 ± 2.54 | 23.09 ± 0.3 | 6.42 ± 0.28 | 9.51 ± 0.22 | 6.83 ± 0.42 | 4.31 ± 0.1 |
| 24 | 0.31 ± 0.01 | 0.57 ± 0.01 | 1.45 ± 0.08 | 0.65 ± 0.01 | n.d. | n.d. | n.d. | n.d. |
| 25 | 82.23 ± 5.28 | 130.8 ± 2.75 | 42.19 ± 2.77 | 38.96 ± 2.58 | 54.59 ± 1.9 | 42.93 ± 0.91 | 44.92 ± 1.31 | 41.59 ± 2.59 |
| 26 | 30.37 ± 0.69 | 55.79 ± 1.56 | 2.63 ± 0.04 | 4.25 ± 0.14 | 1.22 ± 0.03 | 3.37 ± 0.07 | 4.14 ± 0.26 | 0.96 ± 0.02 |
| 27 | 7.91 ± 0.39 | 12.81 ± 0.77 | 5.34 ± 0.23 | 6.11 ± 0.12 | 0.66 ± 0.01 | 0.55 ± 0.03 | 0.61 ± | 0.94 ± 0.05 |
| 28 | 2.34 ± 0.04 | 2.97 ± 0.19 | 1.72 ± 0.09 | 0.05 ± 0.01 | n.d. | n.d. | n.d. | n.d. |
| 29 | 0.18 ± 0.01 | 1.54 ± 0.08 | 2.04 ± 0.05 | 2.98 ± 0.06 | 8.94 ± 0.22 | 55.61 ± 2.88 | 58.92 ± 1.29 | 50.52 ± 1.79 |
| 30 | 0.28 ± 0.01 | 0.29 ± 0.01 | 0.39 ± 0.01 | 0.45 ± 0.01 | n.d. | n.d. | n.d. | n.d. |
| 31 | 7.81 ± 0.36 | 10.24 ± 0.55 | 6.98 ± 0.34 | 12.58 ± 0.48 | 1.53 ± 0.08 | 2.24 ± 0.13 | 2.21 ± 0.08 | 1.52 ± 0.01 |
| 32 | 0.45 ± 0.02 | 1.62 ± 0.08 | 1.97 ± 0.05 | 0.48 ± 0.01 | 0.51 ± 0.01 | 0.93 ± 0.04 | 0.37 ± 0.02 | 1.02 ± 0.05 |
| 33 | 0.36 ± 0.02 | 0.51 ± 0.01 | 0.43 ± 0.01 | 0.43 ± 0.02 | n.d. | n.d. | n.d. | n.d. |
| 34 | 1.3 ± 0.04 | 0.56 ± 0.02 | 2.13 ± 0.12 | 1.22 ± 0.02 | 0.16 ± 0.01 | 8.78 ± 0.42 | 2.49 ± 0.11 | 0.71 ± 0.01 |
| 35 | 5.07 ± 0.07 | 22.66 ± 1.02 | 10.61 ± 0.51 | 4.61 ± 0.24 | 0.39 ± 0.01 | 2.82 ± 0.15 | 2.48 ± 0.04 | 0.39 ± 0.01 |
| 36 | 0.12 ± 0.01 | 0.29 ± 0.02 | 0.07 ± 0.01 | 0.12 ± 0.01 | 0.04 ± 0.01 | 0.16 ± 0.01 | 0.12 ± 0.01 | 0.04 ± 0.01 |
| 37 | 0.44 ± 0.01 | 2.72 ± 0.17 | 3.07 ± 0.14 | 3.53 ± 0.15 | n.d. | n.d. | n.d. | n.d. |
| # | Peel | Pulp | ||||||
|---|---|---|---|---|---|---|---|---|
| CTRL | 5.0 L/ha | 4.0 L/ha | 2.5 L/ha | CTRL | 5.0 L/ha | 4.0 L/ha | 2.5 L/ha | |
| 1 | 32.14 ± 1.97 | 35.44 ± 1.16 | 38.16 ± 2.36 | 31.01 ± 0.54 | 2.41 ± 0.06 | 4.84 ± 0.12 | 2.12 ± 0.06 | 2.26 ± 0.08 |
| 2 | 8.56 ± 0.48 | 10.12 ± 0.22 | 1.16 ± 0.06 | 0.82 ± 0.05 | 1.08 ± 0.03 | 1.23 ± 0.05 | 1.22 ± 0.07 | 1.84 ± 0.04 |
| 3 | 0.81 ± 0.01 | 1.25 ± 0.07 | 1.09 ± 0.06 | 1.73 ± 0.07 | n.d. | n.d. | n.d. | n.d. |
| 4 | 3.89 ± 0.11 | 4.56 ± 0.16 | 4.15 ± 0.16 | 3.95 ± 0.19 | 5.51 ± 0.35 | 7.13 ± 0.44 | 3.98 ± 0.06 | 3.81 ± 0.07 |
| 5 | 1.15 ± 0.06 | 1.45 ± 0.03 | 10.16 ± 0.63 | 9.23 ± 0.46 | 8.84 ± 0.32 | 9.91 ± 0.63 | 9.67 ± 0.32 | 9.69 ± 0.63 |
| 6 | 74.88 ± 3.17 | 15.31 ± 0.52 | 64.44 ± 2.55 | 71.56 ± 4.68 | 36.46 ± 1.03 | 107.02 ± 0.21 | 104.96 ± 4.67 | 134.49 ± 8.74 |
| 7 | 7.14 ± 0.31 | 12.34 ± 0.32 | 13.15 ± 0.68 | 10.77 ± 0.74 | n.d. | n.d. | n.d. | n.d. |
| 8 | 69.4 ± 2.72 | 90.35 ± 1.09 | 72.91 ± 2.71 | 73.18 ± 2.35 | 5.71 ± 0.32 | 6.89 ± 0.41 | 8.12 ± 0.22 | 8.59 ± 0.54 |
| 9 | 19.26 ± 0.85 | 81.67 ± 1.96 | 61.05 ± 3.47 | 60.79 ± 1.94 | 77.76 ± 3.67 | 95.12 ± 4.96 | 115.96 ± 1.91 | 113.25 ± 1.17 |
| 10 | 67.62 ± 2.59 | 81.36 ± 1.37 | 69.26 ± 3.57 | 66.58 ± 4.21 | 14.41 ± 0.92 | 21.82 ± 1.27 | 19.89 ± 0.58 | 20.97 ± 0.67 |
| 11 | 15.56 ± 0.95 | 21.92 ± 1.36 | 15.04 ± 0.51 | 15.77 ± 1.09 | 36.46 ± 2.05 | 32.26 ± 1.02 | 34.12 ± 0.72 | 44.64 ± 1.81 |
| 12 | 172.5 ± 97.08 | 197.55 ± 48.53 | 177.27 ± 44.56 | 182.5 ± 35.87 | 51.17 ± 2.38 | 109.17 ± 1.47 | 63.28 ± 2.01 | 27.48 ± 0.61 |
| 13 | 2.28 ± 0.09 | 3.89 ± 0.24 | 3.75 ± 0.11 | 4.38 ± 0.23 | 3.18 ± 0.05 | 6.75 ± 0.43 | 6.99 ± 0.47 | 8.16 ± 0.52 |
| 14 | n.d. | n.d. | n.d. | n.d. | 1.74 ± 0.02 | 1.54 ± 0.02 | 1.79 ± 0.07 | 2.44 ± 0.05 |
| 15 | n.d. | n.d. | n.d. | n.d. | 0.84 ± 0.05 | 1.44 ± 0.07 | 0.46 ± 0.01 | 0.79 ± 0.03 |
| 16 | 2.77 ± 0.18 | 3.32 ± 0.25 | 4.56 ± 0.27 | 3.46 ± 0.21 | 2.61 ± 0.07 | 5.23 ± 0.08 | 3.82 ± 0.09 | 0.99 ± 0.04 |
| 17 | 2.46 ± 0.03 | 7.74 ± 0.12 | 1.98 ± 0.06 | 7.83 ± 0.45 | 0.63 ± 0.02 | 2.41 ± 0.08 | 2.3 ± 0.14 | 1.94 ± 0.11 |
| 19 | 3.65 ± 0.08 | 6.53 ± 0.07 | 4.28 ± 0.04 | 3.78 ± 0.23 | 1.05 ± 0.05 | 1.56 ± 0.13 | 1.33 ± 0.06 | 1.56 ± 0.02 |
| 21 | 9.26 ± 0.22 | 9.86 ± 0.15 | 8.73 ± 0.12 | 7.22 ± 0.19 | 2.26 ± 0.07 | 2.85 ± 0.16 | 2.66 ± 0.04 | 2.19 ± 0.02 |
| 22 | 61.12 ± 1.07 | 42.35 ± 0.43 | 51.33 ± 0.61 | 35.38 ± 0.86 | n.d. | n.d. | n.d. | n.d. |
| 23 | 3.61 ± 0.08 | 10.13 ± 0.29 | 4.33 ± 0.28 | 3.96 ± 0.07 | 4.61 ± 0.27 | 5.52 ± 0.32 | 3.48 ± 0.08 | 5.43 ± 0.06 |
| 25 | 4.86 ± 0.25 | 13.39 ± 0.78 | 9.83 ± 0.47 | 10.36 ± 0.37 | 9.04 ± 0.32 | 9.25 ± 0.53 | 7.93 ± 0.18 | 8.14 ± 0.09 |
| 26 | 10.98 ± 0.23 | 16.21 ± 0.25 | 14.06 ± 0.21 | 11.68 ± 0.28 | 0.72 ± 0.04 | 1.11 ± 0.06 | 1.17 ± 0.02 | 1.01 ± 0.06 |
| 27 | 1.24 ± 0.05 | 1.21 ± 0.04 | 0.71 ± 0.04 | 2.68 ± 0.06 | 4.42 ± 0.19 | 10.05 ± 0.36 | 8.23 ± 0.12 | 0.82 ± 0.05 |
| 28 | 0.78 ± 0.01 | 1.39 ± 0.08 | 0.65 ± 0.04 | 5.13 ± 0.26 | 2.98 ± 0.15 | 2.72 ± 0.15 | 2.89 ± 0.16 | 2.15 ± 0.05 |
| 29 | 0.13 ± 0.01 | 0.58 ± 0.03 | 0.43 ± 0.02 | 0.67 ± 0.04 | 0.76 ± 0.03 | 0.56 ± 0.03 | 0.61 ± 0.02 | 0.51 ± 0.02 |
| 30 | 0.54 ± 0.02 | 0.68 ± 0.01 | 0.33 ± 0.01 | 0.41 ± 0.01 | n.d. | n.d. | n.d. | n.d. |
| 31 | n.d. | n.d. | n.d. | n.d. | 12.77 ± 0.82 | 15.4 ± 0.19 | 15.32 ± 0.67 | 8.92 ± 0.51 |
| 32 | n.d. | n.d. | n.d. | n.d. | 1.81 ± 0.05 | 1.96 ± 0.02 | 3.56 ± 0.25 | 1.44 ± 0.09 |
| 34 | n.d. | n.d. | n.d. | n.d. | 0.33 ± 0.01 | 1.69 ± 0.11 | 1.39 ± 0.04 | 0.72 ± 0.04 |
| 35 | n.d. | n.d. | n.d. | n.d. | 0.48 ± 0.02 | 2.12 ± 0.05 | 2.06 ± 0.12 | 0.86 ± 0.02 |
| 36 | n.d. | n.d. | n.d. | n.d. | 0.04 ± 0.01 | 0.08 ± 0.01 | 0.13 ± 0.01 | 0.11 ± 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mannino, G.; Ricciardi, M.; Gatti, N.; Serio, G.; Vigliante, I.; Contartese, V.; Gentile, C.; Bertea, C.M. Changes in the Phytochemical Profile and Antioxidant Properties of Prunus persica Fruits after the Application of a Commercial Biostimulant Based on Seaweed and Yeast Extract. Int. J. Mol. Sci. 2022, 23, 15911. https://doi.org/10.3390/ijms232415911
Mannino G, Ricciardi M, Gatti N, Serio G, Vigliante I, Contartese V, Gentile C, Bertea CM. Changes in the Phytochemical Profile and Antioxidant Properties of Prunus persica Fruits after the Application of a Commercial Biostimulant Based on Seaweed and Yeast Extract. International Journal of Molecular Sciences. 2022; 23(24):15911. https://doi.org/10.3390/ijms232415911
Chicago/Turabian StyleMannino, Giuseppe, Maddalena Ricciardi, Noemi Gatti, Graziella Serio, Ivano Vigliante, Valeria Contartese, Carla Gentile, and Cinzia M. Bertea. 2022. "Changes in the Phytochemical Profile and Antioxidant Properties of Prunus persica Fruits after the Application of a Commercial Biostimulant Based on Seaweed and Yeast Extract" International Journal of Molecular Sciences 23, no. 24: 15911. https://doi.org/10.3390/ijms232415911