
Silicone Oil-Induced Glaucomatous Neurodegeneration in Rhesus Macaques
 ,
,
Abstract
:1. Introduction
2. Results
2.1. Intracameral Injection of SO in Rhesus Macaque Monkey Causes RNFL Thinning and Decreases PhNR
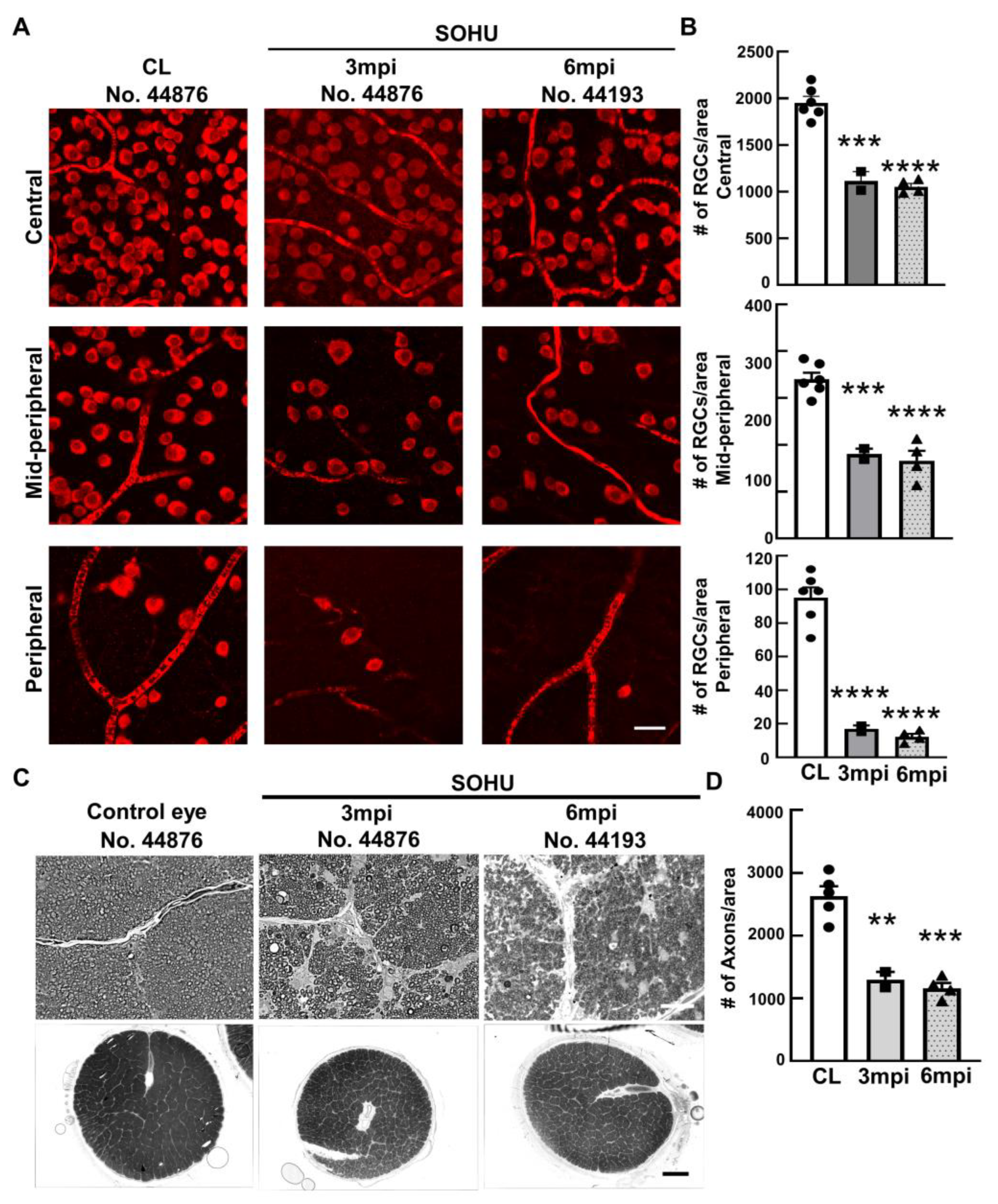
2.2. Significant RGC and ON Degeneration of the SOHU Eyes at 3 mpi and 6 mpi in All Tested Animals
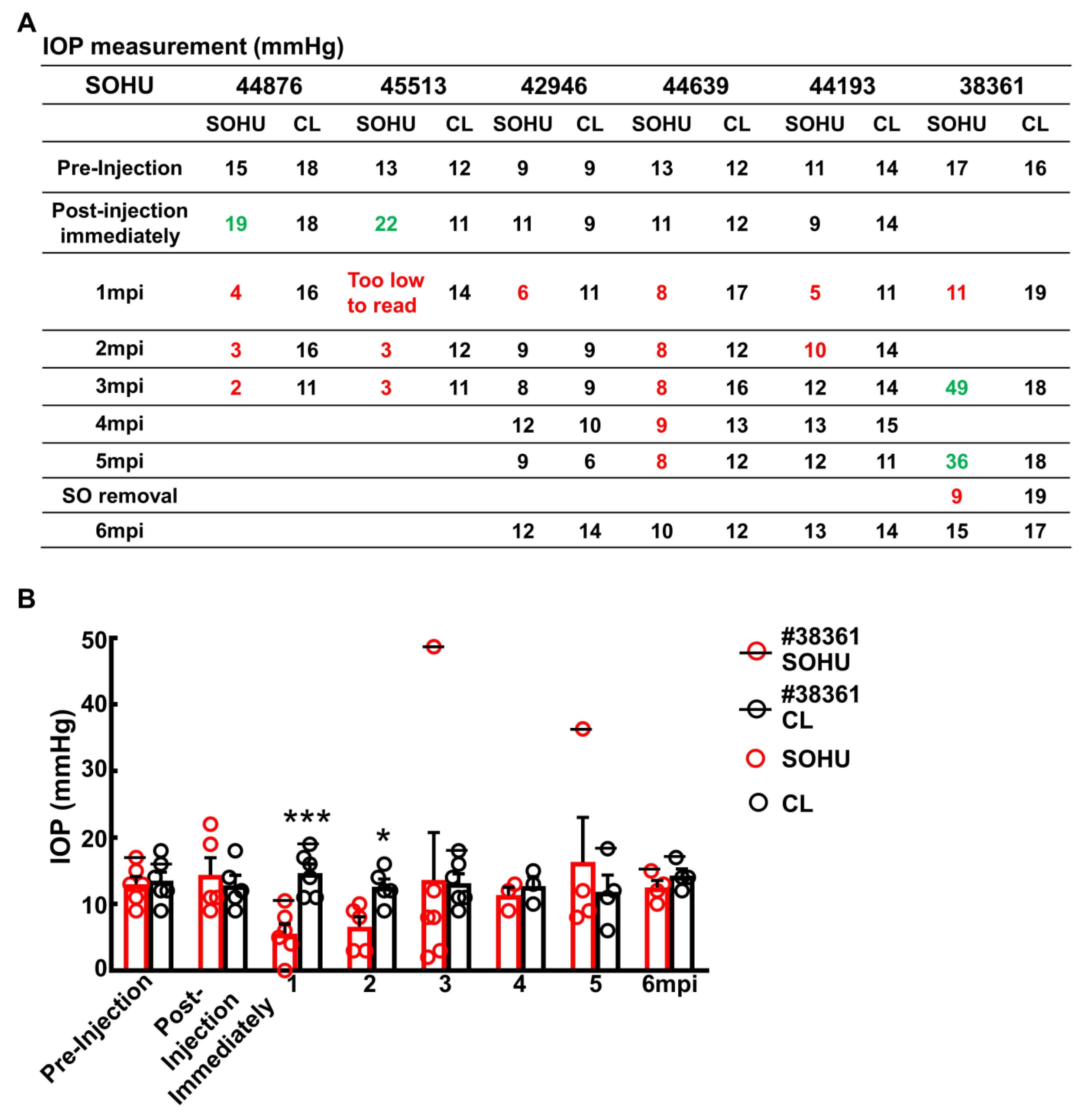
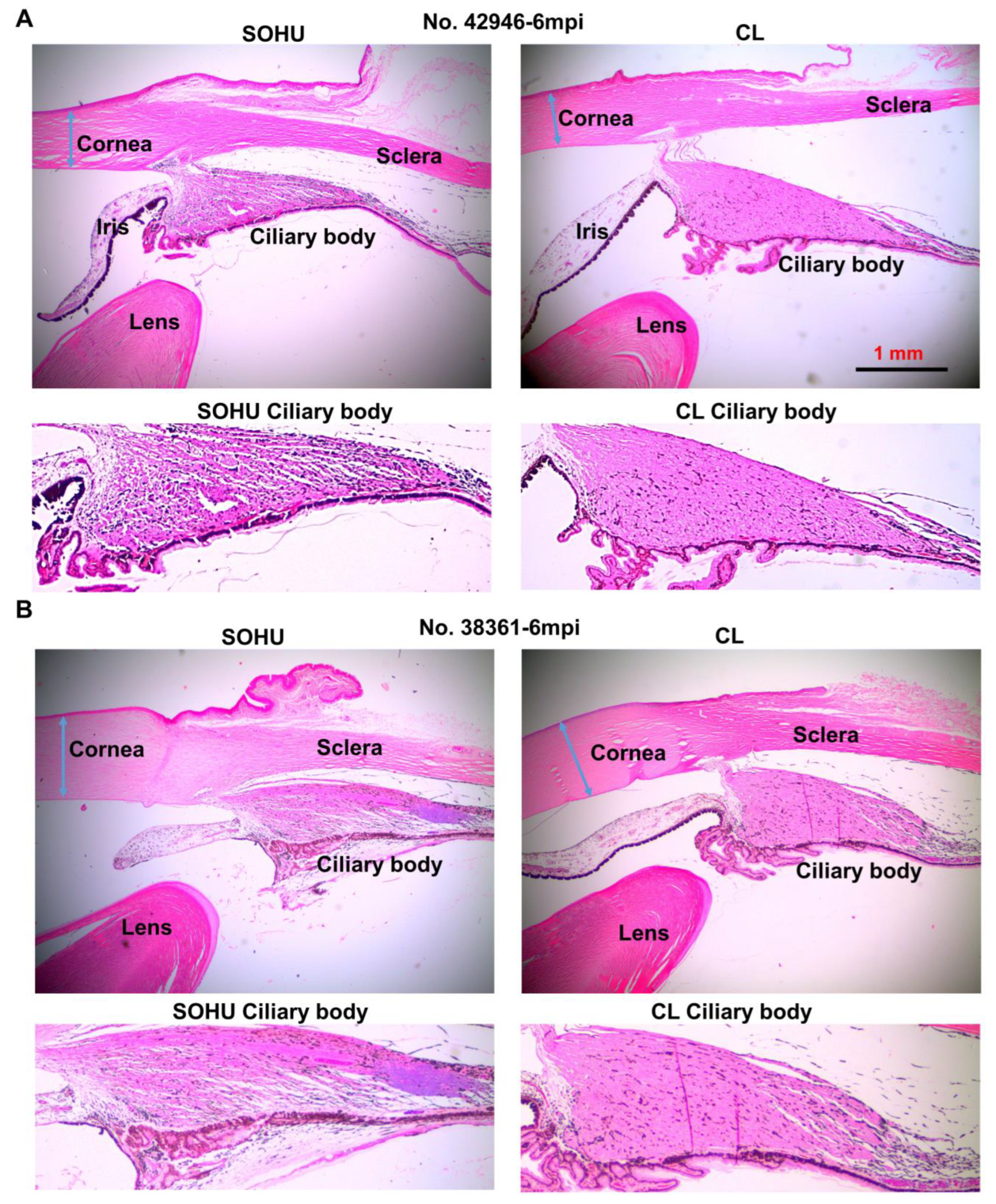
2.3. Dynamic IOP Changes in the SOHU Macaque Eyes Associated with Ciliary Body Atrophy
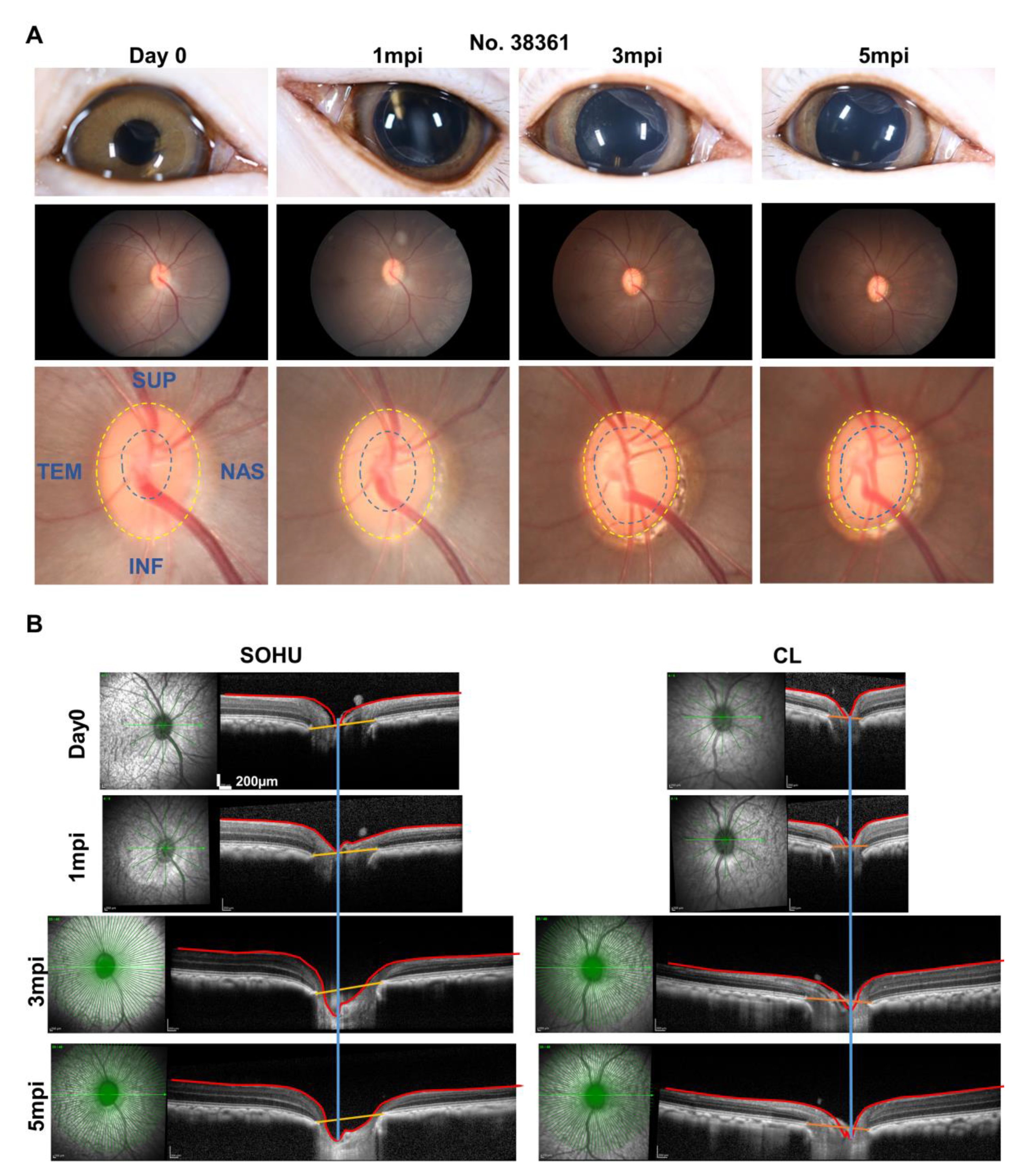
2.4. ON Head “Cupping” Is Present in the SOHU Eye with Persistent IOP Elevation
3. Discussion
4. Materials and Methods
5. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Howell, G.R.; Libby, R.; Jakobs, T.; Smith, R.S.; Phalan, F.C.; Barter, J.W.; Barbay, J.M.; Marchant, J.K.; Mahesh, N.; Porciatti, V.; et al. Axons of retinal ganglion cells are insulted in the optic nerve early in DBA/2J glaucoma. J. Cell Biol. 2007, 179, 1523–1537. [Google Scholar] [CrossRef] [Green Version]
- Tham, Y.C.; Li, X.; Wong, T.Y.; Quigley, H.A.; Aung, T.; Cheng, C.Y. Global prevalence of glaucoma and projections of glaucoma burden through 2040: A systematic review and meta-analysis. Ophthalmology 2014, 121, 2081–2090. [Google Scholar] [CrossRef] [PubMed]
- Weinreb, R.N.; Leung, C.K.; Crowston, J.G.; Medeiros, F.A.; Friedman, D.S.; Wiggs, J.L.; Martin, K.R. Primary open-angle glaucoma. Nat. Rev. Dis. Prim. 2016, 2, 16067. [Google Scholar] [CrossRef] [PubMed]
- Calkins, D.J. Adaptive responses to neurodegenerative stress in glaucoma. Prog. Retin. Eye Res. 2021, 84, 100953. [Google Scholar] [CrossRef] [PubMed]
- Iyer, J.; Vianna, J.R.; Chauhan, B.C.; Quigley, H.A. Toward a new definition of glaucomatous optic neuropathy for clinical research. Curr. Opin. Ophthalmol. 2020, 31, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Burgoyne, C.F. A biomechanical paradigm for axonal insult within the optic nerve head in aging and glaucoma. Exp. Eye Res. 2011, 93, 120–132. [Google Scholar] [CrossRef] [Green Version]
- Calkins, D.J. Critical pathogenic events underlying progression of neurodegeneration in glaucoma. Prog. Retin. Eye Res. 2012, 31, 702–719. [Google Scholar] [CrossRef] [Green Version]
- Heijl, A.; Leske, M.C.; Bengtsson, B.; Hyman, L.; Bengtsson, B.; Hussein, M.; Early Manifest Glaucoma Trial Group. Reduction of intraocular pressure and glaucoma progression: Results from the Early Manifest Glaucoma Trial. Arch. Ophthalmol. 2002, 120, 1268–1279. [Google Scholar] [CrossRef]
- Garway-Heath, D.F.; Crabb, D.P.; Bunce, C.; Lascaratos, G.; Amalfitano, F.; Anand, N.; Azuara-Blanco, A.; Bourne, R.R.; Broadway, D.C.; Cunliffe, I.A.; et al. Latanoprost for open-angle glaucoma (UKGTS): A randomised, multicentre, placebo-controlled trial. Lancet 2015, 385, 1295–1304. [Google Scholar] [CrossRef] [Green Version]
- Anderson, D.R. Collaborative normal tension glaucoma study. Curr. Opin. Ophthalmol. 2003, 14, 86–90. [Google Scholar] [CrossRef]
- Wormald, R.; Virgili, G.; Azuara-Blanco, A. Systematic reviews and randomised controlled trials on open angle glaucoma. Eye 2019, 34, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Wareham, L.K.; Risner, M.L.; Calkins, D.J. Protect, Repair, and Regenerate: Towards Restoring Vision in Glaucoma. Curr. Ophthalmol. Rep. 2020, 8, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Beykin, G.; Norcia, A.M.; Srinivasan, V.J.; Dubra, A.; Goldberg, J.L. Discovery and clinical translation of novel glaucoma biomarkers. Prog. Retin. Eye Res. 2020, 80, 100875. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, L.; Huang, H.; Fang, F.; Webber, H.C.; Zhuang, P.; Dalal, R.; Tang, P.H.; Mahajan, V.; Sun, Y.; et al. Silicone oil-induced ocular hypertension and glaucomatous neurodegeneration in mouse. eLife 2019, 8, e45881. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Fang, F.; Li, L.; Huang, H.; Webber, H.C.; Sun, Y.; Mahajan, V.; Hu, Y. A Reversible Silicon Oil-Induced Ocular Hypertension Model in Mice. J. Vis. Exp. 2019, 153, e60409. [Google Scholar] [CrossRef]
- Fang, F.; Zhang, J.; Zhuang, P.; Liu, P.; Li, L.; Huang, H.; Webber, H.C.; Xu, Y.; Dalal, R.; Sun, Y.; et al. Chronic mild and acute severe glaucomatous neurodegeneration derived from silicone oil-induced ocular hypertension. Sci. Rep. 2021, 11, 9052. [Google Scholar] [CrossRef]
- Ichhpujani, P.; Jindal, A.; Jay Katz, L. Silicone oil induced glaucoma: A review. Graefe’s Arch. Clin. Exp. Ophthalmol. 2009, 247, 1585–1593. [Google Scholar] [CrossRef]
- Kornmann, H.L.; Gedde, S.J. Glaucoma management after vitreoretinal surgeries. Curr. Opin. Ophthalmol. 2016, 27, 125–131. [Google Scholar] [CrossRef]
- Harwerth, R.S.; Smith, E.L., 3rd. Rhesus monkey as a model for normal vision of humans. Am. J. Optom. Vis. Sci. 1985, 62, 633–641. [Google Scholar] [CrossRef]
- Chen, L.; Zhao, Y.; Zhang, H. Comparative Anatomy of the Trabecular Meshwork, the Optic Nerve Head and the Inner Retina in Rodent and Primate Models Used for Glaucoma Research. Vision 2016, 1, 4. [Google Scholar] [CrossRef]
- Burgoyne, C. The morphological difference between glaucoma and other optic neuropathies. J. Neuro-Ophthalmol. Off. J. North Am. Neuro-Ophthalmol. Soc. 2015, 35 (Suppl S1), S8–S21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgoyne, C.F. The non-human primate experimental glaucoma model. Exp. Eye Res. 2015, 141, 57–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balcer, L.J.; Miller, D.H.; Reingold, S.C.; Cohen, J.A. Vision and vision-related outcome measures in multiple sclerosis. Brain 2014, 138, 11–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aktas, O.; Albrecht, P.; Hartung, H.P. Optic neuritis as a phase 2 paradigm for neuroprotection therapies of multiple sclerosis: Update on current trials and perspectives. Curr. Opin. Neurol. 2016, 29, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Costello, F.; Coupland, S.; Hodge, W.; Lorello, G.; Koroluk, J.; Pan, Y.I.; Freedman, M.S.; Zackon, D.H.; Kardon, R.H. Quantifying axonal loss after optic neuritis with optical coherence tomography. Ann. Neurol. 2006, 59, 963–969. [Google Scholar] [CrossRef]
- Viswanathan, S.; Frishman, L.J.; Robson, J.G.; Harwerth, R.S.; Smith, E.L. The photopic negative response of the macaque electroretinogram: Reduction by experimental glaucoma. Investig. Ophthalmol. Vis. Sci. 1999, 40, 1124–1136. [Google Scholar]
- Wilsey, L.; Gowrisankaran, S.; Cull, G.; Hardin, C.; Burgoyne, C.F.; Fortune, B. Comparing three different modes of electroretinography in experimental glaucoma: Diagnostic performance and correlation to structure. Doc. Ophthalmol. 2017, 134, 111–128. [Google Scholar] [CrossRef] [Green Version]
- Coleman, D.J. Evaluation of ciliary body detachment in hypotony. Retina 1995, 15, 312–318. [Google Scholar] [CrossRef]
- Flores-Sanchez, B.C.; Tatham, A.J. Acute angle closure glaucoma. Br. J. Hosp. Med. (Lond) 2019, 80, C174–C179. [Google Scholar] [CrossRef]
- Lockwood, H.; Reynaud, J.; Gardiner, S.; Grimm, J.; Libertiaux, V.; Downs, J.C.; Yang, H.; Burgoyne, C.F. Lamina Cribrosa Microarchitecture in Normal Monkey Eyes Part 1: Methods and Initial Results. Investig. Opthalmology Vis. Sci. 2015, 56, 1618–1637. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Reynaud, J.; Lockwood, H.; Williams, G.; Hardin, C.; Reyes, L.; Stowell, C.; Gardiner, S.K.; Burgoyne, C.F. The connective tissue phenotype of glaucomatous cupping in the monkey eye—Clinical and research implications. Prog. Retin. Eye Res. 2017, 59, 1–52. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Yang, H.; Gardiner, S.K.; Williams, G.; Hardin, C.; Strouthidis, N.G.; Fortune, B.; Burgoyne, C.F. Longitudinal Detection of Optic Nerve Head Changes by Spectral Domain Optical Coherence Tomography in Early Experimental Glaucoma. Investig. Opthalmology Vis. Sci. 2014, 55, 574–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Downs, J.C.; Girkin, C.A. Lamina cribrosa in glaucoma. Curr. Opin. Ophthalmol. 2017, 28, 113–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hopkins, A.; Murphy, R.; Irnaten, M.; Wallace, D.M.; Quill, B.; O’Brien, C. The role of lamina cribrosa tissue stiffness and fibrosis as fundamental biomechanical drivers of pathological glaucoma cupping. Am. J. Physiol. Physiol. 2020, 319, C611–C623. [Google Scholar] [CrossRef]
- Kim, J.-A.; Kim, T.-W.; Weinreb, R.N.; Lee, E.J.; Girard, M.J.A.; Mari, J.M. Lamina Cribrosa Morphology Predicts Progressive Retinal Nerve Fiber Layer Loss In Eyes with Suspected Glaucoma. Sci. Rep. 2018, 8, 738. [Google Scholar] [CrossRef] [Green Version]
- Carelli, V.; La Morgia, C.; Ross-Cisneros, F.N.; Sadun, A.A. Optic neuropathies: The tip of the neurodegeneration iceberg. Hum. Mol. Genet. 2017, 26, R139–R150. [Google Scholar] [CrossRef] [Green Version]
- Toosy, A.T.; Mason, D.F.; Miller, D.H. Optic neuritis. Lancet Neurol. 2014, 13, 83–99. [Google Scholar] [CrossRef]
- Talman, L.S.; Bisker, E.R.; Sackel, D.J.; Long, D.A., Jr.; Galetta, K.M.; Ratchford, J.N.; Lile, D.J.; Farrell, S.K.; Loguidice, M.J.; Remington, G.; et al. Longitudinal study of vision and retinal nerve fiber layer thickness in multiple sclerosis. Ann. Neurol. 2010, 67, 749–760. [Google Scholar]
- McKinnon, S.J. Glaucoma: Ocular Alzheimer’s disease? Front. Biosci. 2003, 8, s1140–s1156. [Google Scholar] [CrossRef]
- Chiasseu, M.; Vargas, J.L.C.; Destroismaisons, L.; Velde, C.V.; Leclerc, N.; Di Polo, A. Tau Accumulation, Altered Phosphorylation, and Missorting Promote Neurodegeneration in Glaucoma. J. Neurosci. 2016, 36, 5785–5798. [Google Scholar] [CrossRef] [Green Version]
- Minegishi, Y.; Nakayama, M.; Iejima, D.; Kawase, K.; Iwata, T. Significance of optineurin mutations in glaucoma and other diseases. Prog. Retin. Eye Res. 2016, 55, 149–181. [Google Scholar] [CrossRef] [PubMed]
- Wiggs, J.L.; Pasquale, L.R. Genetics of glaucoma. Hum. Mol. Genet. 2017, 26, R21–R27. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.H.; Tran, T.; Kim, S.; Park, S.; Stout, J.T.; Chen, R.; Rogers, J.; Yiu, G.; Thomasy, S.; Moshiri, A. Advanced Retinal Imaging and Ocular Parameters of the Rhesus Macaque Eye. Transl. Vis. Sci. Technol. 2021, 10, 7. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Huang, H.; Fang, F.; Sun, Y.; Hu, Y.; Liu, L. Longitudinal Morphological and Functional Assessment of RGC Neurodegeneration After Optic Nerve Crush in Mouse. Front. Cell. Neurosci. 2020, 14, 109. [Google Scholar] [CrossRef]
- Kwong, J.M.K.; Caprioli, J.; Piri, N. RNA Binding Protein with Multiple Splicing: A New Marker for Retinal Ganglion Cells. Investig. Opthalmology Vis. Sci. 2010, 51, 1052–1058. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, A.R.; Müller, L.P.D.S.; Brecha, N.C. The RNA binding protein RBPMS is a selective marker of ganglion cells in the mammalian retina. J. Comp. Neurol. 2013, 522, 1411–1443. [Google Scholar] [CrossRef]
- Smith, R.S. Systematic Evaluation of the Mouse Eye: Anatomy, Pathology, and Biomethods; CRC Press: Boca Raton, FL, USA, 2002; p. 366. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Sex | Date of Birth | Weight (Kg) |
|---|---|---|---|
| 38361 | F | 10 May 2007 (13 years) | 9.68 |
| 42946 | F | 31 May 2012 (8 years) | 9.91 |
| 44193 | M | 06 April 2014 (7 years) | 9.16 |
| 44639 | M | 28 May 2014 (7 years) | 12.95 |
| 44876 | F | 11 May 2015 (6 years) | 10.38 |
| 45513 | M | 22 June 2015 (6 years) | 8.43 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moshiri, A.; Fang, F.; Zhuang, P.; Huang, H.; Feng, X.; Li, L.; Dalal, R.; Hu, Y. Silicone Oil-Induced Glaucomatous Neurodegeneration in Rhesus Macaques. Int. J. Mol. Sci. 2022, 23, 15896. https://doi.org/10.3390/ijms232415896
Moshiri A, Fang F, Zhuang P, Huang H, Feng X, Li L, Dalal R, Hu Y. Silicone Oil-Induced Glaucomatous Neurodegeneration in Rhesus Macaques. International Journal of Molecular Sciences. 2022; 23(24):15896. https://doi.org/10.3390/ijms232415896
Chicago/Turabian StyleMoshiri, Ala, Fang Fang, Pei Zhuang, Haoliang Huang, Xue Feng, Liang Li, Roopa Dalal, and Yang Hu. 2022. "Silicone Oil-Induced Glaucomatous Neurodegeneration in Rhesus Macaques" International Journal of Molecular Sciences 23, no. 24: 15896. https://doi.org/10.3390/ijms232415896