

Transcriptomic Analysis Revealed Antimicrobial Mechanisms of Lactobacillus rhamnosus SCB0119 against Escherichia coli and Staphylococcus aureus
Abstract
:1. Introduction
2. Results
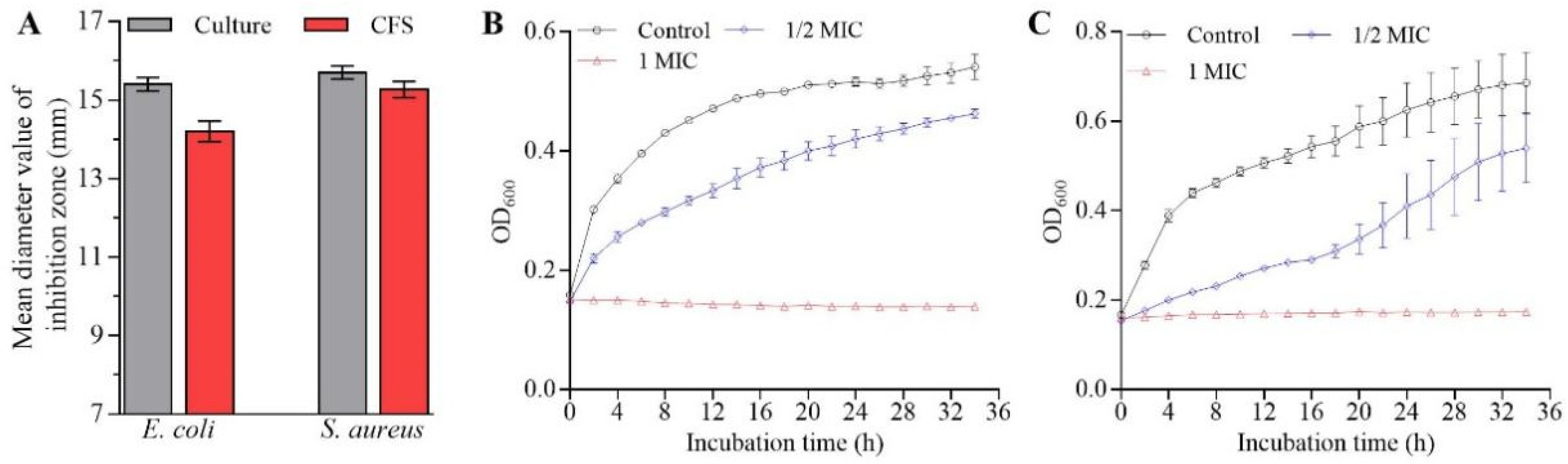
2.1. Reduction in Bacterial Growth of E. coli and S. aureus Caused by L. rhamnosus SCB0119
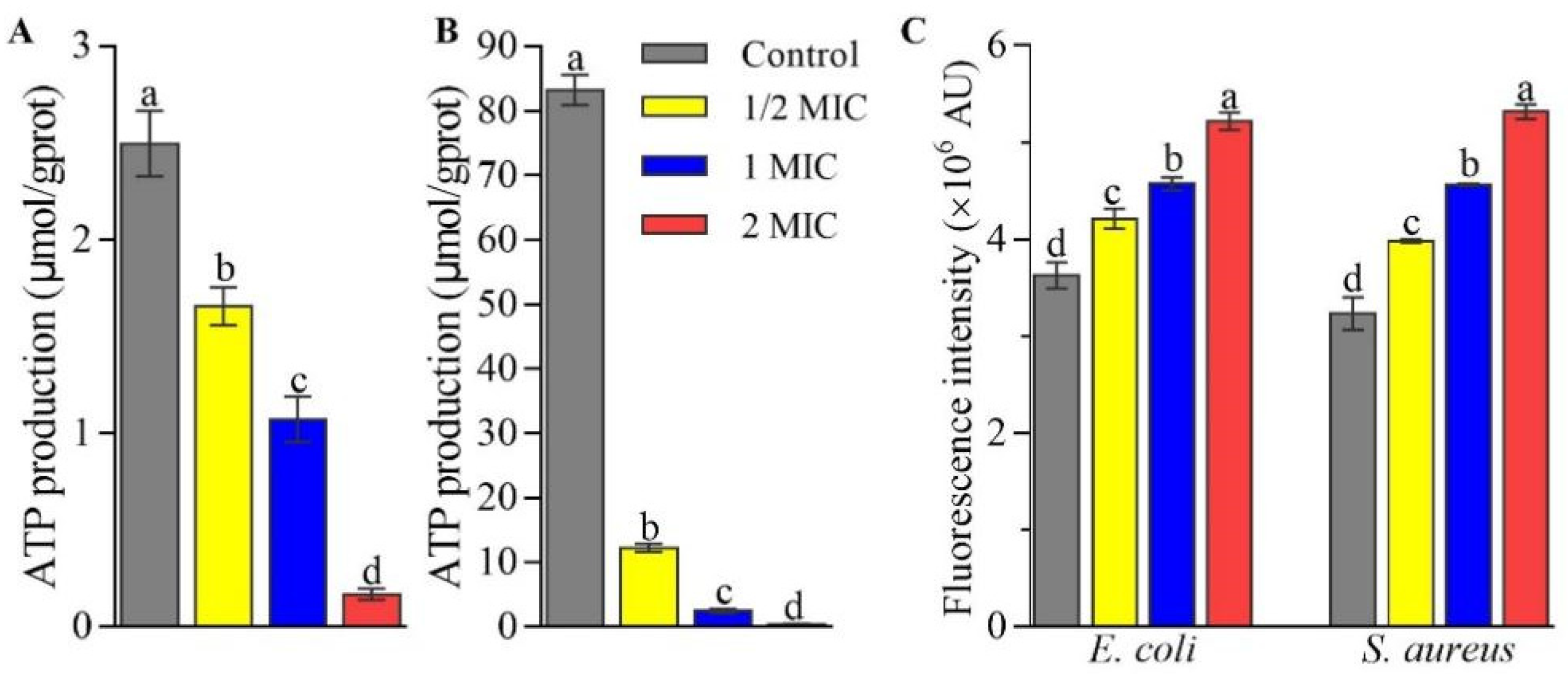
2.2. Effects of CFS on the Production of Intracellular ATP and Reactive Oxygen Species in E. coli and S. aureus
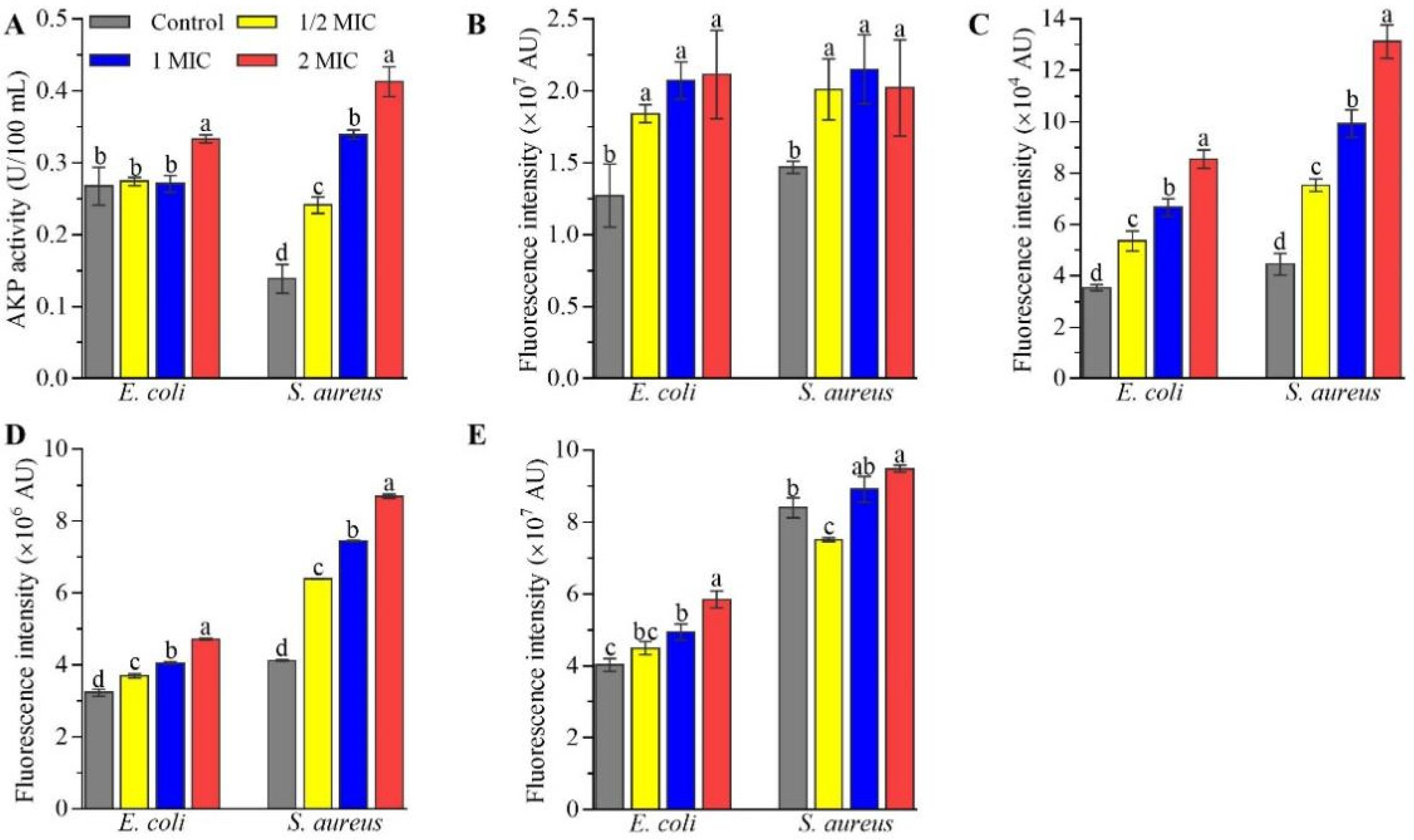
2.3. Changes in Cell Morphology and Structure of E. coli and S. aureus Caused by CFS
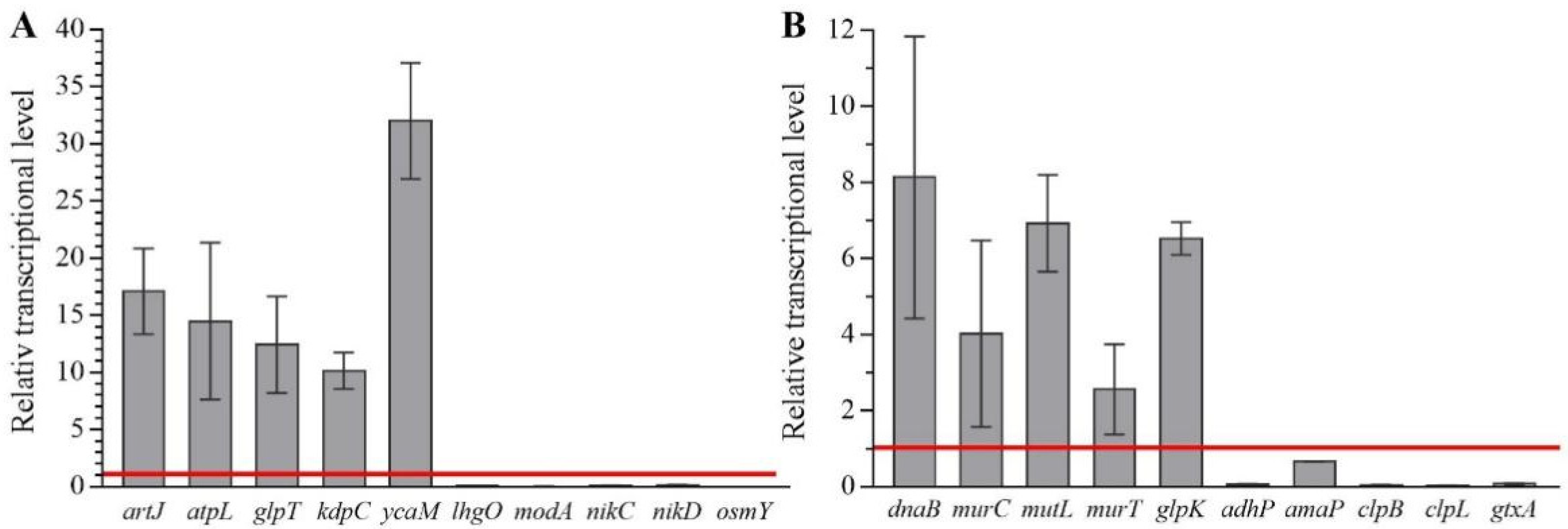
2.4. Transcriptomic Analysis of E. coli and S. aureus Cells in Response to CFS
3. Discussion
4. Material and Methods
4.1. Bacterial Strains and Culture Condition
4.2. Preparation of Cell-Free Culture Supernatants and Cells of L. rhamnosus SCB0119
4.3. Determination of the Antibacterial Activity of L. rhamnosus SCB0119
4.4. Minimum Inhibitory Concentrations of CFS against E. coli and S. aureus
4.5. Growth Curve of E. coli and S. aureus after Treatment with CFS
4.6. Scanning Electron Microscopy and Transmission Electron Microscopy
4.7. Concentrations of Alkaline Phosphatase and Intracellular ATP
4.8. Assay for Outer Membrane Permeability and Inner Membrane Integrity
4.9. Assays for Membrane Potential, pH Gradient, and Reactive Oxygen Species
4.10. Transcriptome Sequencing and Analysis
4.11. qRT-PCR
4.12. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| CFS | cell-free culture supernatant |
| LB | Luria Bertani |
| MICs | minimum inhibitory concentrations |
| SEM | scanning electron microscopy |
| TEM | transmission electron microscopy |
| AKP | alkaline phosphatase |
| NPN | 1-N-phenylnaphthylamine |
| PI | propidium iodide |
| ΔΨ | membrane potential |
| ΔpH | pH gradient |
| ROS | reactive oxygen species |
| DEGs | differentially expressed genes |
| GO | gene ontology |
| KEGG | Kyoto encyclopedia of genes and genomes |
| MRS | De Man Rogosa Sharpe |
| PBS | phosphate buffer saline |
| FPKM | fragments per kilobase per million fragments mapped |
| ATCC | American Type Culture Collection |
| SD | standard deviation |
References
- Kirk, M.D.; Pires, S.M.; Black, R.E.; Caipo, M.; Crump, J.A.; Devleesschauwer, B.; Dopfer, D.; Fazil, A.; Fischer-Walker, C.L.; Hald, T.; et al. World health organization estimates of the global and regional disease burden of 22 foodborne bacterial, protozoal, and viral diseases, 2010: A data synthesis. PLos Med. 2015, 12, e1001921. [Google Scholar] [CrossRef] [Green Version]
- Mangal, M.; Bansal, S.; Sharma, S.K.; Gupta, R.K. Molecular detection of foodborne pathogens: A rapid and accurate answer to food safety. Crit. Rev. Food Sci. Nutr. 2016, 56, 1568–1584. [Google Scholar] [CrossRef] [PubMed]
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne illness acquired in the United States-major pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [CrossRef]
- Johnson, J.R. Foodborne illness acquired in the United States. Emerg. Infect. Dis. 2011, 17, 1338–1339. [Google Scholar] [CrossRef] [Green Version]
- Croxen, M.A.; Law, R.J.; Scholz, R.; Keeney, K.M.; Wlodarska, M.; Finlay, B.B. Recent advances in understanding enteric pathogenic Escherichia coli. Clin. Microbiol. Rev. 2013, 26, 822–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Croxen, M.A.; Finlay, B.B. Molecular mechanisms of Escherichia coli pathogenicity. Nat. Rev. Microbiol. 2010, 8, 26–38. [Google Scholar] [CrossRef]
- Muller, A.; Stephan, R.; Nuesch-Inderbinen, M. Distribution of virulence factors in ESBL-producing Escherichia coli isolated from the environment, livestock, food and humans. Sci. Total Environ. 2016, 541, 667–672. [Google Scholar] [CrossRef] [Green Version]
- Majowicz, S.E.; Scallan, E.; Jones-Bitton, A.; Sargeant, J.M.; Stapleton, J.; Angulo, F.J.; Yeung, D.H.; Kirk, M.D. Global incidence of human Shiga toxin-producing Escherichia coli infections and deaths: A systematic review and knowledge synthesis. Foodborne Pathog. Dis. 2014, 11, 447–455. [Google Scholar] [CrossRef] [Green Version]
- Argudin, M.A.; Mendoza, M.C.; Rodicio, M.R. Food poisoning and Staphylococcus aureus enterotoxins. Toxins 2010, 2, 1342–1751. [Google Scholar] [CrossRef]
- Mourenza, A.; Gil, J.A.; Mateos, L.M.; Letek, M. Novel treatments and preventative strategies against food-poisoning caused by staphylococcal species. Pathogens 2021, 10, 91. [Google Scholar] [CrossRef]
- Kadariya, J.; Smith, T.C.; Thapaliya, D. Staphylococcus aureus and staphylococcal food-borne disease: An ongoing challenge in public health. Biomed Res. Int. 2014, 2014, 827965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, S.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G. Staphylococcus aureus infections: Epidemiology, pathophysiology, clinical manifestations, and management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mostafa, A.A.; Al-Askar, A.A.; Almaary, K.S.; Dawoud, T.M.; Sholkamy, E.N.; Bakri, M.M. Antimicrobial activity of some plant extracts against bacterial strains causing food poisoning diseases. Saudi J. Biol. Sci. 2018, 25, 361–366. [Google Scholar] [CrossRef] [PubMed]
- Davies, J.; Davies, D. Origins and evolution of antibiotic resistance. Microbiol. Mol. Biol. Rev. 2010, 74, 417. [Google Scholar] [CrossRef] [Green Version]
- Reis, J.A.; Paula, A.T.; Casarotti, S.N.; Penna, A. Lactic acid bacteria antimicrobial compounds: Characteristics and applications. Food Eng. Rev. 2012, 4, 124–140. [Google Scholar] [CrossRef]
- Vieco-Saiz, N.; Belguesmia, Y.; Raspoet, R.; Auclair, E.; Gancel, F.; Kempf, I.; Drider, D. Benefits and inputs from lactic acid bacteria and their bacteriocins as alternatives to antibiotic growth promoters during food-animal production. Front. Microbiol. 2019, 10, 57. [Google Scholar] [CrossRef] [Green Version]
- Zamberlin, S.; Spehar, I.D.; Kelava, N.; Samarzija, D. Probiotic bacterium Lactobacillus rhamnosus: Beneficial and a dverse effects on human health. Milchwissenschaft-Milk Sci. Int. 2012, 67, 30–33. [Google Scholar]
- Ozogul, F.; Hamed, I. The importance of lactic acid bacteria for the prevention of bacterial growth and their biogenic amines formation: A review. Crit. Rev. Food Sci. 2018, 10, 1660–1670. [Google Scholar] [CrossRef]
- Zhang, Y.C.; Zhang, L.W.; Du, M.; Yi, H.X.; Guo, C.F.; Tuo, Y.F.; Han, X.; Li, J.Y.; Zhang, L.L.; Yang, L. Antimicrobial activity against Shigella sonnei and probiotic properties of wild Lactobacilli from fermented food. Microbiol. Res. 2011, 167, 27–31. [Google Scholar] [CrossRef]
- Shi, S.Q.; Gong, L.; Yu, H.; He, G.Y.; Zhang, J.J.; Han, Y.; Liu, Y.N.; Hu, J.; Dong, J.S.; Liu, J.; et al. Antagonistic activity and mechanism of Lactobacillus rhamnosus SQ511 against Salmonella enteritidis. 3 Biotech 2022, 12, 126. [Google Scholar] [CrossRef]
- Wei, Y.H.; Wang, J.Z.; Liu, Z.; Pei, J.J.; Brennan, C.; Abd El-Aty, A.M. Isolation and characterization of bacteriocin-producing Lacticaseibacillus rhamnosus XN2 from yak yoghurt and its bacteriocin. Molecules 2022, 27, 2066. [Google Scholar] [CrossRef]
- Zorova, L.D.; Popkov, V.A.; Plotnikov, E.Y.; Silachev, D.N.; Pevzner, I.B.; Jankauskas, S.S.; Babenko, V.A.; Zorov, S.D.; Balakireva, A.V.; Juhaszova, M.; et al. Mitochondrial membrane potential. Anal. Biochem. 2018, 552, 50–59. [Google Scholar] [CrossRef]
- Kamal, R.M.; Alnakip, M.E.; Abd El Aal, S.F.; Bayoumi, M.A. Bio-controlling capability of probiotic strain Lactobacillus rhamnosus against some common foodborne pathogens in yoghurt. Int. Dairy J. 2018, 85, 1–7. [Google Scholar] [CrossRef]
- Khan, I.; Bahuguna, A.; Kumar, P.; Bajpai, V.K.; Kang, S.C. Antimicrobial potential of carvacrol against uropathogenic Escherichia coli via membrane disruption, depolarization, and reactive oxygen species generation. Front. Microbiol. 2017, 8, 2421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ezraty, B.; Gennaris, A.; Barras, F.; Collet, J.F. Oxidative stress, protein damage and repair in bacteria. Nat. Rev. Microbiol. 2017, 15, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Warnes, S.L.; Caves, V.; Keevil, C.W. Mechanism of copper surface toxicity in Escherichia coli O157:H7 and Salmonella involves immediate membrane depolarization followed by slower rate of DNA destruction which differs from that observed for Gram-positive bacteria. Environ. Microbiol. 2012, 14, 1730–1743. [Google Scholar] [CrossRef]
- Sun, D.D.; Zhang, W.W.; Mou, Z.P.; Chen, Y.; Guo, F.; Yang, E.D.; Wang, W.Y. Transcriptome analysis reveals silver nanoparticle-decorated quercetin antibacterial molecular mechanism. ACS Appl. Mater. Interfaces 2017, 9, 10047–10060. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Chen, W.X.; Dou, Z.M.; Chen, R.H.; Hu, Y.Y.; Chen, W.J.; Chen, H.M. Antimicrobial effect of black pepper petroleum ether extract for the morphology of Listeria monocytogenes and Salmonella typhimurium. J. Food Sci. Tech. Mys. 2017, 54, 2067–2076. [Google Scholar] [CrossRef]
- Diao, M.M.; Qi, D.P.; Xu, M.M.; Lu, Z.X.; Lv, F.X.; Bie, X.M.; Zhang, C.; Zhao, H.Z. Antibacterial activity and mechanism of monolauroyl-galactosylglycerol against Bacillus cereus. Food Control 2018, 85, 339–344. [Google Scholar] [CrossRef]
- Ibrahim, H.R.; Sugimoto, Y.; Aoki, T. Ovotransferrin antimicrobial peptide (OTAP-92) kills bacteria through a membrane damage mechanism. Biochim. Biophys. Acta 2000, 1523, 196–205. [Google Scholar] [CrossRef]
- Cotter, P.D.; Ross, R.P.; Hill, C. Bacteriocins—A viable alternative to antibiotics? Nat. Rev. Microbiol. 2013, 11, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Fu, Y.Y.; Liu, F.; Liu, Z.J.; Ma, J.G.; Jiang, R.; Song, C.N.; Jiang, Z.M.; Hou, J.C. Purification and antimicrobial mechanism of a novel bacteriocin produced by Lactobacillus rhamnosus 1.0320. LWT-Food Sci. Technol. 2021, 137, 110338. [Google Scholar] [CrossRef]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods—A review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef]
- Gong, H.S.; Meng, X.C.; Wang, H. Mode of action of plantaricin MG, a bacteriocin active against Salmonella typhimurium. J. Basic Microbiol. 2010, 50, S37–S45. [Google Scholar] [CrossRef]
- Lou, Z.X.; Wang, H.X.; Zhu, S.; Ma, C.Y.; Wang, Z.P. Antibacterial activity and mechanism of action of chlorogenic acid. J. Food Sci. 2011, 76, M398–M403. [Google Scholar] [CrossRef]
- Liu, R.; Zhu, F.; Lu, L.; Fu, A.; Lu, J.; Deng, Z.; Liu, T. Metabolic engineering of fatty acyl-ACP reductase-dependent pathway to improve fatty alcohol production in Escherichia coli. Metab. Eng. 2014, 22, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Xiao, W.; Zhang, J.; Xie, Z.; Ding, M.; Yuan, Y. Heterologous biosynthesis and manipulation of alkanes in Escherichia coli. Metab. Eng. 2016, 38, 19–28. [Google Scholar] [CrossRef]
- Li, Y.; Zamble, D.B. Nickel homeostasis and nickel regulation: An overview. Chem. Rev. 2009, 109, 4617–4643. [Google Scholar] [CrossRef]
- Dixon, S.J.; Stockwell, B.R. The role of iron and reactive oxygen species in cell death. Nat. Chem. Biol. 2014, 10, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Kohanski, M.A.; Dwyer, D.J.; Wierzbowski, J.; Cottarel, G.; Collins, J.J. Mistranslation of membrane proteins and two-component system activation trigger antibiotic-mediated cell death. Cell 2008, 135, 679–690. [Google Scholar] [CrossRef] [Green Version]
- Chenault, S.S.; Earhart, C.F. Organization of genes encoding membrane proteins of the Escherichia coli ferrienterobactin permease. Mol. Microbiol. 1991, 5, 1405–1413. [Google Scholar] [CrossRef] [PubMed]
- Mousavi, F.; Bojko, B.; Bessonneau, V.; Pawliszyn, J. Cinnamaldehyde characterization as an antibacterial agent toward E. coli metabolic profile using 96-blade solid-phase microextraction coupled to liquid chromatography–mass spectrometry. J. Proteome Res. 2016, 15, 963–975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyanoiri, Y.; Ishida, Y.; Takeda, M.; Terauchi, T.; Inouye, M.; Kainosho, M. Highly efficient residue-selective labeling with isotope-labeled Ile, Leu, and Val using a new auxotrophic E. coli strain. J. Biomol. NMR 2016, 65, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Lennon, C.W.; Thamsen, M.; Friman, E.T.; Cacciaglia, A.; Sachsenhauser, V.; Sorgenfrei, F.A.; Wasik, M.A.; Bardwell, J.C.A. Folding optimization in vivo uncovers new chaperones. J. Mol. Biol. 2015, 427, 2983–2994. [Google Scholar] [CrossRef] [Green Version]
- Yan, Z.; Hussain, S.; Wang, X.; Bernstein, H.D.; Bardwell, J.C.A. Chaperone OsmY facilitates the biogenesis of a major family of autotransporters. Mol. Microbiol. 2019, 112, 1373–1387. [Google Scholar] [CrossRef]
- McCoy, L.S.; Xie, Y.; Tor, Y. Antibiotics that target protein synthesis. WIREs RNA 2011, 2, 209–232. [Google Scholar] [CrossRef]
- Bai, J.; Wu, Y.; Bu, Q.; Zhong, K.; Gao, H. Comparative study on antibacterial mechanism of shikimic acid and quinic acid against Staphylococcus aureus through transcriptomic and metabolomic approaches. LWT-Food Sci. Technol. 2022, 153, 112441. [Google Scholar] [CrossRef]
- Peacock, J.R.; Walvoord, R.R.; Chang, A.Y.; Kozlowski, M.C.; Gamper, H.; Hou, Y. Amino acid-dependent stability of the acyl linkage in aminoacyl-tRNA. RNA 2014, 20, 758–764. [Google Scholar] [CrossRef] [Green Version]
- Lin, E.; Huang, C. Crystal structure of the single-stranded DNA-binding protein SsbB in complex with the anticancer drug 5-fluorouracil: Extension of the 5-fluorouracil interactome to include the oligonucleotide/oligosaccharide-binding fold protein. Biochem. Biophys. Res. Commun. 2021, 534, 41–46. [Google Scholar] [CrossRef]
- Grosser, M.R.; Paluscio, E.; Thurlow, L.R.; Dillon, M.M.; Cooper, V.S.; Kawula, T.H.; Richardson, A.R. Genetic requirements for Staphylococcus aureus nitric oxide resistance and virulence. PLoS Pathog. 2018, 14, e1006907. [Google Scholar] [CrossRef] [Green Version]
- Arrioja-Breton, D.; Mani-Lopez, E.; Palou, E.; Lopez-Malo, A. Antimicrobial activity and storage stability of cell-free supernatants from lactic acid bacteria and their applications with fresh beef. Food Control 2020, 115, 107286. [Google Scholar] [CrossRef]
- De Alcntara, A.; Bruzaroski, S.R.; Luiz, L.L.; de Souza, C.; Poli-Frederico, R.C.; Fagnani, R.; de Santana, E. Antimicrobial activity of Lactobacillus rhamnosus against Pseudomonas fluorescens and Pseudomonas putida from raw milk. J. Food Process Preserv. 2019, 43, e14082. [Google Scholar] [CrossRef]
- Benavides, A.B.; Ulcuango, M.; Yepez, L.; Tenea, G.N. Assessment of the in vitro bioactive properties of lactic acid bacteria isolated from native ecological niches of Ecuador. Rev. Argent. Microbiol. 2016, 48, 236–244. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.X.; Li, J.H.; Qu, G.R.; Guo, D.; Yang, Y.P.; Ma, X.Y.; Wang, M.X.; Xu, Y.F.; Wang, Y.T.; Xia, X.D.; et al. Antibacterial activity and mechanism of coenzyme Q (0) against Escherichia coli. Foodborne Pathog. Dis. 2021, 18, 398–404. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Lu, Y.Y.; Li, X.; Yi, Y.L.; Wang, X.; Shan, Y.Y.; Liu, B.F.; Zhou, Y.; Lu, X. Action mode of bacteriocin BM1829 against Escherichia coli and Staphylococcus aureus. Food Biosci. 2021, 39, 100794. [Google Scholar] [CrossRef]
- Ali, Z.I.; Saudi, A.M.; Albrecht, R.; Talaat, A.M. The inhibitory effect of nisin on Mycobacterium avium ssp. paratuberculosis and its effect on mycobacterial cell wall. J. Dairy Sci. 2019, 102, 4935–4944. [Google Scholar] [CrossRef]
- Modiri, S.; Kermanshahi, R.K.; Soudi, M.R.; Arab, S.S.; Khammari, A.; Cousineau, B.; Vali, H.; Zahiri, H.S.; Noghabi, K.A. Multifunctional acidocin 4356 combats Pseudomonas aeruginosa through membrane perturbation and virulence attenuation: Experimental results confirm molecular dynamics simulation. Appl. Environ. Microbiol. 2020, 86, e00367-20. [Google Scholar] [CrossRef]
- Yuroff, A.S.; Sabat, G.; Hickey, W.J. Transporter-mediated uptake of 2-chloro- and 2-hydroxybenzoate by Pseudomonas huttiensis strain D1. Appl. Environ. Microbiol. 2003, 69, 7401–7408. [Google Scholar] [CrossRef] [Green Version]
- Ma, B.; Fang, C.; Lu, L.S.; Wang, M.Z.; Xue, X.Y.; Zhou, Y.; Li, M.K.; Hu, Y.; Luo, X.X.; Hou, Z. The antimicrobial peptide thanatin disrupts the bacterial outer membrane and inactivates the NDM-1 metallo-beta-lactamase. Nat. Commun. 2019, 10, 3517. [Google Scholar] [CrossRef] [Green Version]
- Yarlagadda, V.; Akkapeddi, P.; Manjunath, G.B.; Haldar, J. Membrane active vancomycin analogues: A strategy to combat bacterial resistance. J. Med. Chem. 2014, 57, 4558–4568. [Google Scholar] [CrossRef]
- Fei, P.; Ali, M.A.; Gong, S.Y.; Sun, Q.; Bi, X.; Liu, S.F.; Guo, L. Antimicrobial activity and mechanism of action of olive oil polyphenols extract against Cronobacter sakazakii. Food Control 2018, 94, 289–294. [Google Scholar] [CrossRef]
- Molenaar, D.; Abee, T.; Konings, W.N. Continuous measurement of the cytoplasmic pH in Lactococcus lactis with a fluorescent pH indicator. Biochim. Biophys. Acta 1991, 1115, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
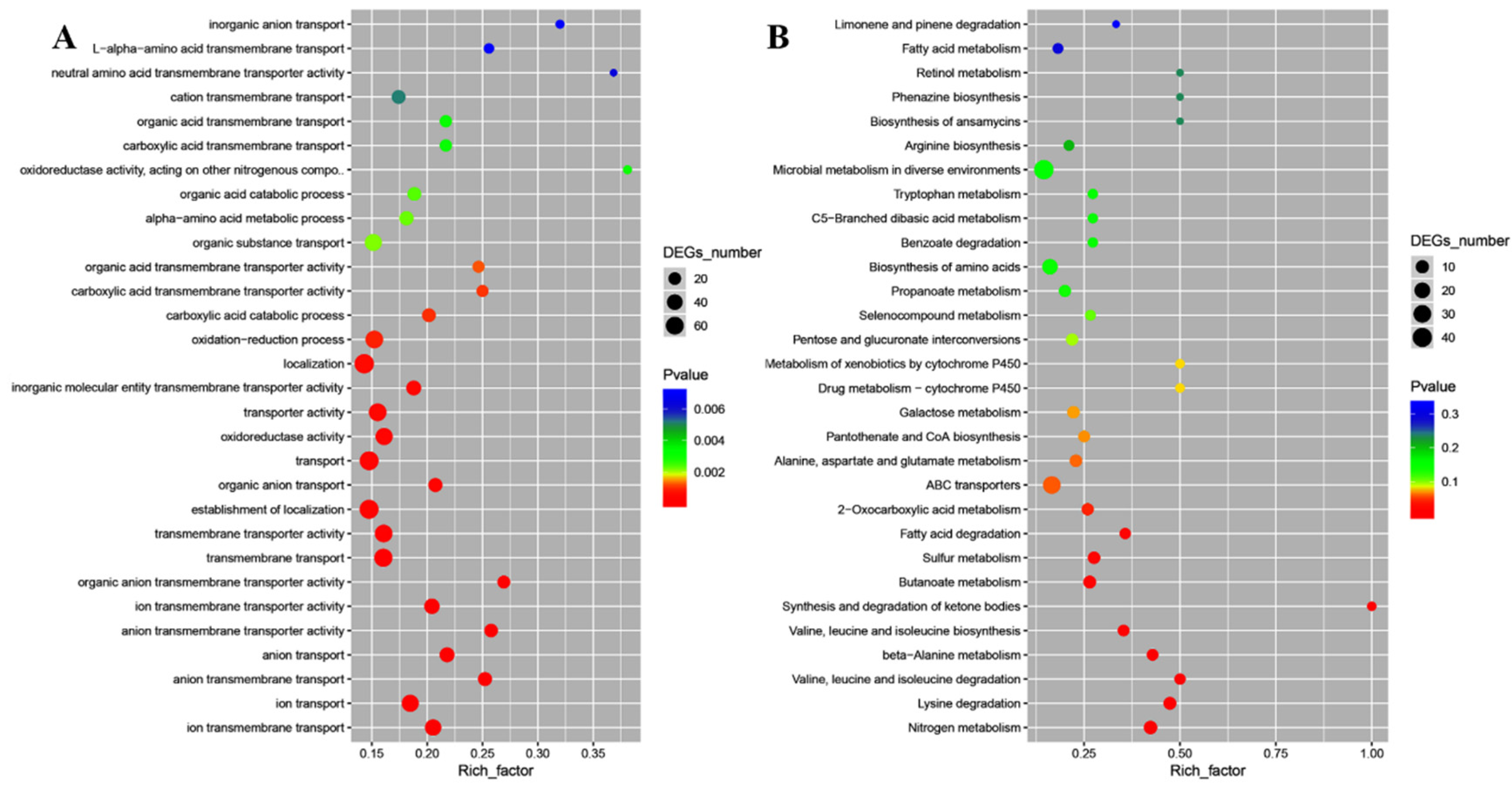{kind=link}
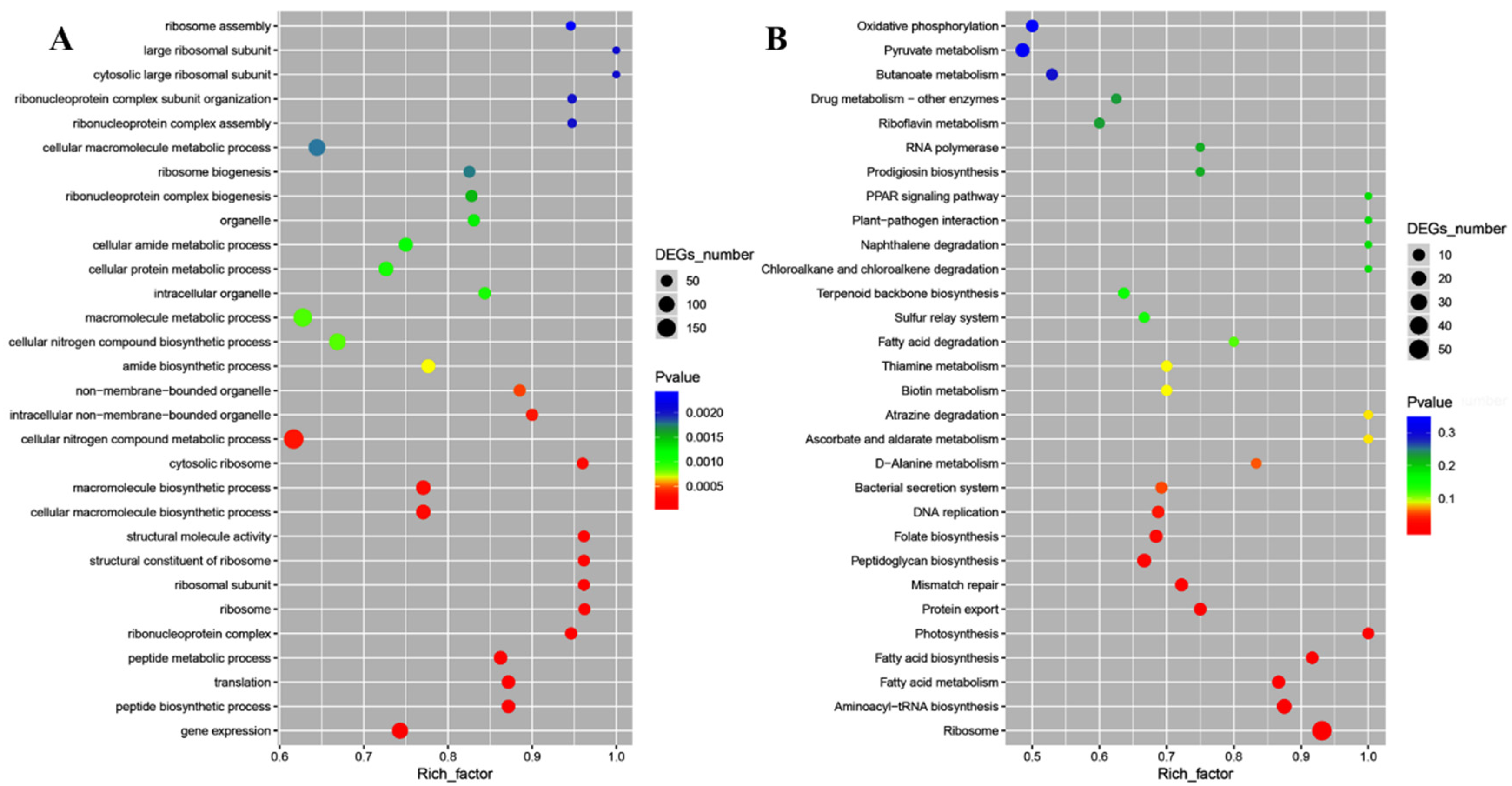{kind=link}
| Gene ID | RPKM Control | RPKM CFS | log2 Ratio | Gene Annotation |
|---|---|---|---|---|
| Fatty acid degradation | ||||
| C7A06_RS17240 | 9.40 ± 0.16 | 2.62 ± 0.05 | −1.83961 | adhP; alcohol dehydrogenase AdhP |
| Biosynthesis of amino acids | ||||
| C7A06_RS27640 | 4.88 ± 1.53 | 21.23 ± 2.32 | 2.119947 | ilvN; acetolactate synthase small subunit |
| C7A06_RS27645 | 5.18 ± 0.68 | 24.79 ± 0.65 | 2.259582 | ilvI; acetolactate synthase 3 large subunit |
| C7A06_RS27660 | 17.62 ± 0.97 | 58.19 ± 0.51 | 1.723499 | leuA; 2-isopropylmalate synthase |
| C7A06_RS27665 | 15.63 ± 2.17 | 48.10 ± 7.45 | 1.621654 | leuB; 3-isopropylmalate dehydrogenase |
| C7A06_RS30700 | 15.96 ± 0.02 | 155.56 ± 1.24 | 3.284926 | lysC; lysine-sensitive aspartokinase 3 |
| ABC transporters | ||||
| C7A06_RS01665 | 14.27 ± 0.12 | 6.44 ± 0.43 | −1.14856 | nikE; nickel import ATP-binding protein NikE |
| C7A06_RS01670 | 39.57 ± 0.39 | 15.70 ± 1.38 | −1.33379 | nikD; nickel import ATP-binding protein NikD |
| C7A06_RS01675 | 43.38 ± 0.05 | 11.55 ± 1.74 | −1.90906 | nikC; nickel ABC transporter permease subunit NikC |
| C7A06_RS01680 | 39.74 ± 2.21 | 10.25 ± 2.12 | −1.95437 | nikB; nickel ABC transporter permease subunit NikB |
| C7A06_RS01685 | 10.05 ± 1.12 | 1.86 ± 0.26 | −2.43055 | nikA; nickel ABC transporter substrate-binding protein |
| C7A06_RS01780 | 0.15 ± 0.08 | 1.01 ± 0.42 | 2.788886 | livH; ABC transporter permease LivH |
| C7A06_RS01790 | 4.29 ± 0.27 | 14.90 ± 1.90 | 1.794947 | livG;ABC transporter ATP-binding protein LivG |
| C7A06_RS24375 | 3.74 ± 0.98 | 1.53 ± 0.65 | −1.29111 | fepD; Fe3+ -siderophore ABC transporter permease |
| Gene ID | RPKM Control | RPKM CFS | log2 Ratio | Gene Annotation |
|---|---|---|---|---|
| Fatty acid degradation | ||||
| EKM74_RS11915 | 1832.07 ± 490.15 | 697.14 ± 121.09 | −1.39396 | adhP; alcohol dehydrogenase |
| DNA replication | ||||
| EKM74_RS04795 | 85.2 ± 44.94 | 39.85 ± 1.91 | −1.09611 | single-stranded DNA-binding protein |
| Ribosome | ||||
| EKM74_RS01000 | 77.06 ± 19.54 | 474.46 ± 30.77 | 2.622215 | accC; acetyl-CoA carboxylase biotin carboxylase subunit |
| EKM74_RS01125 | 36.7 ± 12.69 | 642.43 ± 184.29 | 4.129847 | rpmG; 50S ribosomal protein L33 |
| EKM74_RS01310 | 21.83 ± 0.96 | 168.95 ± 53.18 | 2.951911 | rpsT; 30S ribosomal protein S20 |
| EKM74_RS01660 | 458.35 ± 97.45 | 2756.92 ± 263.1 | 2.588527 | rpmA; 50S ribosomal protein L27 |
| EKM74_RS01670 | 102.05 ± 29.64 | 4057 ± 488.97 | 5.313122 | rplU; 50S ribosomal protein L21 |
| EKM74_RS01855 | 19.42 ± 10.05 | 621.08 ± 94.23 | 4.999194 | rplT; 50S ribosomal protein L20 |
| EKM74_RS01860 | 51.94 ± 21.24 | 5478.49 ± 45.03 | 6.720704 | rpmI; 50S ribosomal protein L35 |
| EKM74_RS02080 | 80.66 ± 17.27 | 3451.94 ± 642.32 | 5.419314 | rpsD; 30S ribosomal protein S4 |
| EKM74_RS05465 | 190.13 ± 11.18 | 3317.01 ± 500.37 | 4.124828 | rpsI; 30S ribosomal protein S9 |
| EKM74_RS05470 | 150.17 ± 31.96 | 5880.76 ± 17.12 | 5.291357 | rplM; 50S ribosomal protein L13 |
| EKM74_RS05495 | 122.74 ± 4.32 | 3341.82 ± 309.69 | 4.766903 | rplQ; 50S ribosomal protein L17 |
| EKM74_RS05505 | 287.56 ± 78.6 | 2321.16 ± 704.2 | 3.01292 | rpsK; 30S ribosomal protein S11 |
| EKM74_RS05510 | 582.25 ± 5.82 | 10,572.69 ± 1079.55 | 4.182553 | rpsM; 30S ribosomal protein S13 |
| EKM74_RS05515 | 259.55 ± 26.92 | 8724.69 ± 798.78 | 5.071017 | rpmJ; 50S ribosomal protein L36 |
| EKM74_RS05535 | 328.44 ± 51.85 | 2770.35 ± 239.31 | 3.076378 | rplO; 50S ribosomal protein L15 |
| EKM74_RS05540 | 370.08 ± 105.31 | 13,564.79 ± 1089.74 | 5.195877 | rpmD; 50S ribosomal protein L30 |
| EKM74_RS05545 | 183.9 ± 22.05 | 4254.61 ± 754.45 | 4.532035 | rpsE; 30S ribosomal protein S5 |
| EKM74_RS05550 | 205.01 ± 26.43 | 9035.82 ± 328.78 | 5.461917 | rplR; 50S ribosomal protein L18 |
| EKM74_RS05555 | 81.16 ± 4.22 | 1809.37 ± 428.25 | 4.478643 | rplF; 50S ribosomal protein L6 |
| EKM74_RS05560 | 98.44 ± 3.27 | 3014.88 ± 247.03 | 4.93666 | rpsH; 30S ribosomal protein S8 |
| EKM74_RS05570 | 126.29 ± 2.95 | 4279.22 ± 391.93 | 5.082565 | rplE; 50S ribosomal protein L5 |
| EKM74_RS05575 | 138.6 ± 11.64 | 4600.35 ± 70.63 | 5.052719 | rplX; 50S ribosomal protein L24 |
| EKM74_RS05580 | 181.27 ± 11.93 | 4136.6 ± 837.23 | 4.512211 | rplN; 50S ribosomal protein L14 |
| EKM74_RS05585 | 140.46 ± 16.11 | 4981.9 ± 113.47 | 5.148408 | rpsQ; 30S ribosomal protein S17 |
| EKM74_RS05590 | 154.79 ± 34.38 | 4723.29 ± 418.72 | 4.931395 | rpmC; 50S ribosomal protein L29 |
| EKM74_RS05595 | 194.99 ± 8.39 | 3442.61 ± 618.09 | 4.142009 | rplP; 50S ribosomal protein L16 |
| EKM74_RS05600 | 333.68 ± 100.38 | 6017.46 ± 493.68 | 4.172617 | rpsC; 30S ribosomal protein S3 |
| EKM74_RS05605 | 267.84 ± 59.12 | 9325.05 ± 0.95 | 5.121682 | rplV; 50S ribosomal protein L22 |
| EKM74_RS05610 | 210.47 ± 4.59 | 7349.55 ± 191.55 | 5.125963 | rpsS; 30S ribosomal protein S19 |
| EKM74_RS05615 | 231.04 ± 32.27 | 4313.16 ± 576.59 | 4.222508 | rplB; 50S ribosomal protein L2 |
| EKM74_RS05620 | 87.68 ± 7.24 | 1918.56 ± 543.34 | 4.451662 | rplW; 50S ribosomal protein L23 |
| EKM74_RS05625 | 145.16 ± 12.67 | 3427.73 ± 563.02 | 4.561522 | rplD; 50S ribosomal protein L4 |
| EKM74_RS05630 | 188.98 ± 15.87 | 7819.97 ± 221.75 | 5.37082 | rplC; 50S ribosomal protein L3 |
| EKM74_RS05635 | 294.61 ± 86.56 | 9259.91 ± 978.26 | 4.97412 | rpsJ; 30S ribosomal protein S10 |
| EKM74_RS07510 | 151.39 ± 9.3 | 705.28 ± 120.89 | 2.219912 | rplI; 50S ribosomal protein L9 |
| EKM74_RS07590 | 114.58 ± 17.68 | 1144.54 ± 138.16 | 3.320336 | rpmH; 50S ribosomal protein L34 |
| EKM74_RS10450 | 236.72 ± 37.3 | 7542.84 ± 237.11 | 4.993827 | rpsF; 30S ribosomal protein S6 |
| EKM74_RS10460 | 55.84 ± 4.69 | 531.48 ± 99.45 | 3.250636 | rpsR; 30S ribosomal protein S18 |
| EKM74_RS11530 | 51.86 ± 16.74 | 493.86 ± 62.28 | 3.251456 | rpmG; 50S ribosomal protein L33 |
| EKM74_RS11545 | 230.05 ± 33.59 | 6745.81 ± 496 | 4.873994 | rplK; 50S ribosomal protein L11 |
| EKM74_RS11550 | 89.54 ± 9.77 | 1345.42 ± 304.26 | 3.909295 | rplA; 50S ribosomal protein L1 |
| EKM74_RS11560 | 75.58 ± 8 | 6432.15 ± 34.02 | 6.411134 | rplJ; 50S ribosomal protein L10 |
| EKM74_RS11565 | 38.15 ± 13.06 | 2351.81 ± 364.44 | 5.945945 | rplL; 50S ribosomal protein L7/L12 |
| EKM74_RS11590 | 185.09 ± 16.78 | 6759.64 ± 1120.81 | 5.190643 | rpsL; 30S ribosomal protein S12 |
| EKM74_RS11595 | 156.22 ± 21.95 | 3889.22 ± 770.69 | 4.637864 | rpsG; 30S ribosomal protein S7 |
| EKM74_RS14380 | 74.47 ± 6.98 | 956.98 ± 89.75 | 3.683755 | rpmF; 50S ribosomal protein L32 |
| EKM74_RS15340 | 44.89 ± 6.07 | 387.7 ± 59.81 | 3.110434 | rpsP; 30S ribosomal protein S16 |
| EKM74_RS15355 | 56.77 ± 3.55 | 839.03 ± 212.16 | 3.885462 | rplS; 50S ribosomal protein L19 |
| EKM74_RS15435 | 112.72 ± 30.02 | 3718.42 ± 314.52 | 5.043867 | rpsB; 30S ribosomal protein S2 |
| EKM74_RS15530 | 178.88 ± 2.05 | 2219.27 ± 90.62 | 3.633007 | rpsO; 30S ribosomal protein S15 |
| Aminoacyl-tRNA biosynthesis | ||||
| EKM74_RS02305 | 35.37 ± 21.65 | 401.03 ± 43.46 | 3.503254 | leuS; leucine-tRNA ligase |
| EKM74_RS03605 | 63.4 ± 16.82 | 997.64 ± 109.83 | 3.975903 | gatB; Asp-tRNA (Asn)/Glu-tRNA (Gln) amidotransferase subunit |
| EKM74_RS11370 | 90.84 ± 8.58 | 509.92 ± 53.53 | 2.488778 | lysS; lysine-tRNA ligase |
| EKM74_RS11490 | 107.77 ± 37.9 | 605.02 ± 32.1 | 2.489052 | gltX; glutamate-tRNA ligase |
| EKM74_RS14775 | 34.9 ± 14.25 | 239.53 ± 20.28 | 2.77887 | pheT; phenylalanine-tRNA ligase subunit beta |
| EKM74_RS15105 | 30.17 ± 17.78 | 273.57 ± 29.78 | 3.180715 | ileS; isoleucine-tRNA ligase |
| Oxidative phosphorylation | ||||
| EKM74_RS04830 | 170.47 ± 3.15 | 3033.08 ± 240.81 | 4.153207183 | atpD; F0-F1 ATP synthase subunit beta |
| EKM74_RS04840 | 97.18 ± 1.86 | 1251.21 ± 170.89 | 3.686521282 | atpA; F0-F1 ATP synthase subunit alpha |
| EKM74_RS04855 | 241.14 ± 56.65 | 1679.25 ± 147.57 | 2.799862175 | atpE; F0-F1 ATP synthase subunit C |
| EKM74_RS04860 | 239.15 ± 52.69 | 1550.63 ± 98.56 | 2.696892312 | atpB; F0-F1 ATP synthase subunit A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, H.; Zhou, G.; Yang, X.-M.; Chen, G.-J.; Chen, H.-B.; Liao, Z.-L.; Zhong, Q.-P.; Wang, L.; Fang, X.; Wang, J. Transcriptomic Analysis Revealed Antimicrobial Mechanisms of Lactobacillus rhamnosus SCB0119 against Escherichia coli and Staphylococcus aureus. Int. J. Mol. Sci. 2022, 23, 15159. https://doi.org/10.3390/ijms232315159
Peng H, Zhou G, Yang X-M, Chen G-J, Chen H-B, Liao Z-L, Zhong Q-P, Wang L, Fang X, Wang J. Transcriptomic Analysis Revealed Antimicrobial Mechanisms of Lactobacillus rhamnosus SCB0119 against Escherichia coli and Staphylococcus aureus. International Journal of Molecular Sciences. 2022; 23(23):15159. https://doi.org/10.3390/ijms232315159
Chicago/Turabian StylePeng, Huan, Gang Zhou, Xi-Miao Yang, Guo-Jun Chen, Hai-Bin Chen, Zhen-Lin Liao, Qing-Ping Zhong, Li Wang, Xiang Fang, and Jie Wang. 2022. "Transcriptomic Analysis Revealed Antimicrobial Mechanisms of Lactobacillus rhamnosus SCB0119 against Escherichia coli and Staphylococcus aureus" International Journal of Molecular Sciences 23, no. 23: 15159. https://doi.org/10.3390/ijms232315159