
From Cocoa to Chocolate: Effect of Processing on Flavanols and Methylxanthines and Their Mechanisms of Action


Abstract
:1. Introduction
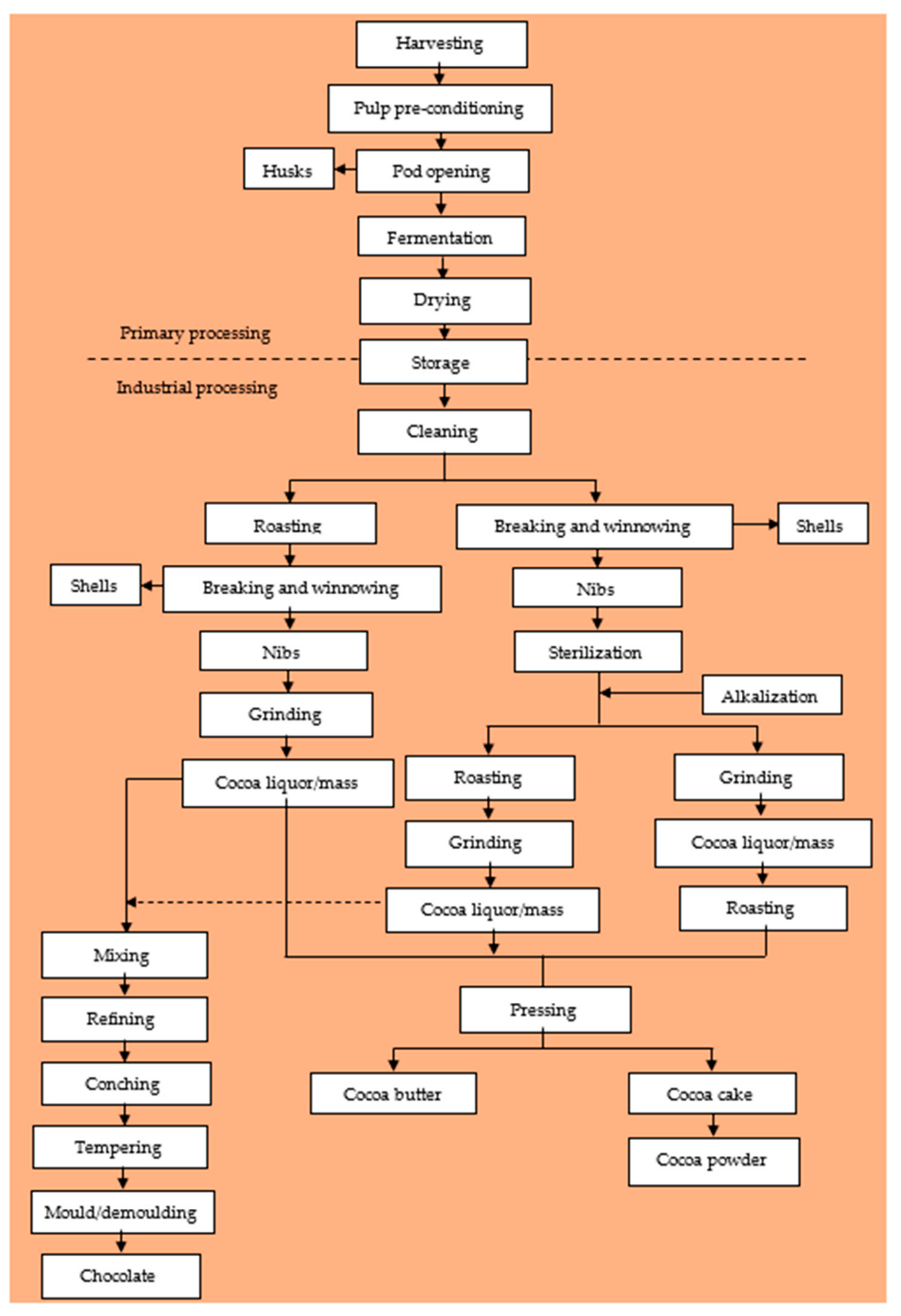
2. Cocoa Bean Processing
2.1. Primary Processing of Cocoa Beans
2.1.1. Harvesting
2.1.2. Pod Storage
2.1.3. Pod Breaking
2.1.4. Cocoa Bean Fermentation
2.1.5. Drying of Fermented Cocoa Beans
2.2. Industrial Processing of Cocoa Beans
2.2.1. Cleaning
2.2.2. Breaking and Winnowing
2.2.3. Sterilization
2.2.4. Alkalization (Dutching)
2.2.5. Cocoa Bean Roasting
2.2.6. Cocoa Nib Grinding and Liquor Processing
2.3. Chocolate Production
3. Mechanisms of Action of Cocoa Flavanols
3.1. Recent Advances in Cocoa Flavanols on Signaling Pathways
3.2. Recent Advances in Cocoa Flavanols on Nrf2 Pathway/Antioxidant Defenses and Inflammatory Process
3.3. Recent Advances in Cocoa Flavanols on Cognitive Function
3.4. Recent Advances in Cocoa Flavanols in Cardiovascular Function
4. Mechanisms of Action of Cocoa Methylxanthines
4.1. Recent Advances in Mechanisms of Action of Cocoa Theobromine
4.2. Recent Advances in Theobromine on Signaling Pathways
4.3. Recent Advances in Cocoa Theobromine on Cardiovascular Function
4.4. Recent Advances in Cocoa Theobromine on Cognitive Function and Other Effects
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Perez, M.; Lopez-Yerena, A.; Vallverdú-Queralt, A. Traceability, authenticity and sustainability of cocoa and chocolate products: A challenge for the chocolate industry. Crit. Rev. Food Sci. Nutr. 2022, 62, 475–489. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Escudero, F.; Casimiro-Gonzales, S.; Fernández-Prior, Á.; Cancino Chávez, K.; Gómez-Mendoza, J.; de la Fuente-Carmelino, L.; Muñoz, A.M. Colour, fatty acids, bioactive compounds, and total antioxidant capacity in commercial cocoa beans (Theobroma cacao L.). LWT Food Sci. Technol. 2021, 147, 111629. [Google Scholar] [CrossRef]
- Melo, T.S.; Pires, T.C.; Engelmann, J.V.P.; Monteiro, A.L.O.; Maciel, L.F.; Bispo, E.D.S. Evaluation of the content of bioactive compounds in cocoa beans during the fermentation process. J. Food Sci. Technol. 2021, 58, 1947–1957. [Google Scholar] [CrossRef] [PubMed]
- Samanta, S.; Sarkar, T.; Chakraborty, R.; Rebezov, M.; Shariati, M.A.; Thiruvengadam, M.; Rengasamy, K.R.R. Dark chocolate: An overview of its biological activity, processing, and fortification approaches. Curr. Res. Food Sci. 2022, 5, 1916–1943. [Google Scholar] [CrossRef] [PubMed]
- Rimbach, G.; Egert, S.; de Pascual-Teresa, S. Chocolate: (un)Healthy source of polyphenols? Genes Nutr. 2011, 6, 1–3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kongor, J.E.; Hinneh, M.; Van de Walle, D.; Afoakwa, E.O.; Boeckx, P.; Dewettinck, K. Factors influencing quality variation in cocoa (Theobroma cacao) bean flavour profile—A review. Food Res. Int. 2016, 82, 44–52. [Google Scholar] [CrossRef]
- Quarmine, W.; Haagsma, R.; Sakyi-Dawson, O.; Asante, F.; van Huis, A.; Obeng-Ofori, D. Incentives for cocoa bean production in Ghana: Does quality matter? NJAS Wagening J. Life Sci. 2012, 60–63, 7–14. [Google Scholar] [CrossRef] [Green Version]
- Babar, A.; Bujold, E.; Leblanc, V.; Lavoie-Lebel, E.; Paquette, J.; Bazinet, L. Changes in endothelial function, arterial stiffness and blood pressure in pregnant women after consumption of high-flavanol and high-theobromine chocolate: A double blind randomized clinical trial. Hypertens. Pregnancy 2018, 37, 68–80. [Google Scholar] [CrossRef]
- Ahmad Bhat, J.; Gupta, S.; Kumar, M. Neuroprotective effects of theobromine in transient global cerebral ischemia-reperfusion rat model. Biochem. Biophys. Res. Commun. 2021, 571, 74–80. [Google Scholar] [CrossRef]
- Ahmad Bhat, J.; Kumar, M. Neuroprotective effects of theobromine in permanent bilateral common carotid artery occlusion rat model of cerebral hypoperfusion. Metab. Brain Dis. 2022, 37, 1787–1801. [Google Scholar] [CrossRef]
- Gao, L.; Ge, W.; Peng, C.; Guo, J.; Chen, N.; He, L. Association between dietary theobromine and cognitive function in a representative American population: A cross-sectional study. J. Prev. Alzheimer Dis. 2022, 3, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Cichon, N.; Saluk-Bijak, J.; Gorniak, L.; Przyslo, L.; Bijak, M. Flavonoids as a natural enhancer of neuroplasticity. An overview of the mechanism of neurorestorative action. Antioxidants 2020, 9, 1035. [Google Scholar] [CrossRef] [PubMed]
- Goya, L.; San Román, R.; de Pascual-Teresa, S. Polyphenols’ effect on cerebrovascular health. Curr. Med. Chem. 2021, 29, 1029–1044. [Google Scholar] [CrossRef] [PubMed]
- Gasmi, A.; Mujawdiya, P.K.; Noor, S.; Lysiuk, R.; Darmohray, R.; Piscopo, S.; Lenchyk, L.; Antonyak, H.; Dehtiarova, K.; Shanaida, M.; et al. Polyphenols in Metabolic Diseases. Molecules 2022, 27, 6280. [Google Scholar] [CrossRef] [PubMed]
- Afoakwa, E.O. Chocolate Science and Technology, 2nd ed.; Wiley-Blackwell: Oxford, UK, 2016; pp. 21–550. [Google Scholar]
- Di Mattia, C.D.; Sacchetti, G.; Mastrocola, D.; Serafini, M. From cocoa to chocolate: The impact of processing on in vitro antioxidant activity and the effects of chocolate on antioxidant markers in vivo. Front. Immunol. 2017, 8, 1207. [Google Scholar] [CrossRef] [Green Version]
- Żyżelewicz, D.; Budryn, G.; Oracz, J.; Antolak, H.; Kręgiel, D.; Kaczmarska, M. The effect on bioactive components and characteristics of chocolate by functionalization with raw cocoa beans. Food Res. Int. 2018, 113, 234–244. [Google Scholar] [CrossRef]
- Muñoz, M.S.; Cortina, J.R.; Vaillant, F.E.; Parra, S.E. An overview of the physical and biochemical transformation of cocoa seeds to beans and to chocolate: Flavor formation. Crit. Rev. Food Sci. Nutr. 2020, 60, 1593–1613. [Google Scholar] [CrossRef]
- Hinneh, M.; Van de Walle, D.; Tzompa-Sosa, D.A.; De Winne, A.; Termote, S.; Messens, K.; Van Durme, J.; Afoakwa, E.O.; De Cooman, L.; Dewettinck, K. Tuning the aroma profiles of FORASTERO cocoa liquors by varying pod storage and bean roasting temperature. Food Res. Int. 2019, 125, 108550. [Google Scholar] [CrossRef]
- Sharif, S. Flavor development during cocoa roasting. In Drying and Roasting of Cocoa and Coffee, 1st ed.; Hii, C.L., Borém, F.M., Eds.; CRC Press—Taylor & Francis Group: Boca Raton, FL, USA, 2020; pp. 63–87. [Google Scholar]
- Febrianto, N.A.; Wang, S.; Zhu, F. Chemical and biological properties of cocoa beans affected by processing: A review. Crit. Rev. Food Sci. Nutr. 2021, 62, 1–32. [Google Scholar] [CrossRef]
- Febrianto, N.A.; Zhu, F. Changes in the composition of methylxanthines, polyphenols, and volatiles and sensory profiles of cocoa beans from the Sul 1 genotype affected by fermentation. J. Agric. Food Chem. 2020, 68, 8658–8675. [Google Scholar] [CrossRef]
- Júnior, P.C.G.; dos Santos, V.B.; Lopes, A.S.; de Souza, J.P.I.; Pina, J.R.S.; Júnior, G.C.A.C.; Marinho, P.S.B. Determination of theobromine and caffeine in fermented and unfermented Amazonian cocoa (Theobroma cacao L.) beans using square wave voltammetry after chromatographic separation. Food Control 2020, 108, 106887. [Google Scholar] [CrossRef]
- Andres-Lacueva, C.; Monagas, M.; Khan, N.; Izquierdo-Pulido, M.; Urpi-Sarda, M.; Permanyer, J.; Lamuela-Raventós, R.M. Flavanol and flavonol contents of cocoa powder products: Influence of the manufacturing process. J. Agric. Food Chem. 2008, 56, 3111–3117. [Google Scholar] [CrossRef] [PubMed]
- Eker, M.E.; Aaby, K.; Budic-Leto, I.; Rimac Brnčić, S.; El, S.N.; Karakaya, S.; Simsek, S.; Manach, C.; Wiczkowski, W.; de Pascual-Teresa, S. A review of factors affecting anthocyanin bioavailability: Possible implications for the inter-individual variability. Foods 2020, 9, 2. [Google Scholar] [CrossRef] [Green Version]
- Afoakwa, E. Cocoa Production and Processing Technology, 1st ed.; CRC Press: Boca Raton, FL, USA, 2014; pp. 1–374. [Google Scholar]
- Krähmer, A.; Engel, A.; Kadow, D.; Ali, N.; Umaharan, P.; Kroh, L.W.; Schulz, H. Fast and neat—Determination of biochemical quality parameters in cocoa using near-infrared spectroscopy. Food Chem. 2015, 181, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Dand, R. The International Cocoa Trade, 3rd ed.; Woodhead Publishing: Sawston, UK, 2010; pp. 55–59. [Google Scholar]
- Amoa-Awua, W.K. Methods of cocoa fermentation and drying. In Cocoa and Coffee Fermentations, 1st ed.; Schwan, R.F., Fleet, G.H., Eds.; CRC Press—Taylor & Francis Group: New York, NY, USA, 2015; pp. 71–116. [Google Scholar]
- Pereira-Caro, G.; Borges, G.; Nagai, C.; Jackson, M.C.; Yokota, T.; Crozier, A.; Ashihara, H. Profiles of phenolic compounds and purine alkaloids during the development of seeds of Theobroma cacao cv. Trinitario. J. Agric. Food Chem. 2012, 61, 427–434. [Google Scholar] [CrossRef]
- Febrianto, N.A.; Zhu, F. Diversity in composition of bioactive compounds among 26 cocoa genotypes. J. Agric. Food Chem. 2019, 67, 9501–9509. [Google Scholar] [CrossRef] [PubMed]
- Dang, Y.K.T.; Nguyen, H.V.H. Effects of maturity at harvest and fermentation conditions on bioactive compounds of cocoa beans. Plant Foods Hum. Nutr. 2019, 74, 54–60. [Google Scholar] [CrossRef]
- Zheng, X.Q.; Koyama, Y.; Nagai, C.; Ashihara, H. Biosynthesis, accumulation and degradation of theobromine in developing Theobroma cacao fruits. J. Plant Physiol. 2004, 161, 363–369. [Google Scholar] [CrossRef]
- Nazaruddin, R.; Seng, L.; Hassan, O.; Said, M. Effect of pulp pre-conditioning on the content of polyphenols in cocoa beans (Theobroma cacao) during fermentation. Ind. Crops Prod. 2006, 24, 87–94. [Google Scholar] [CrossRef]
- Afoakwa, E.O.; Kongor, J.E.; Takrama, J.; Budu, A. Changes in nib acidification and biochemical composition during fermentation of pulp pre-conditioned cocoa (Theobroma cacao) beans. Int. Food Res. J. 2013, 20, 1843–1853. [Google Scholar]
- Sulaiman, K.B.; Yang, T.A.; Ariffin, F. Colour and antioxidant properties of cocoa beans from pods storage and fermentation using shallow box. Int. J. Sci. Technol. 2017, 3, 455–465. [Google Scholar] [CrossRef]
- Aikpokpodion, P.E.; Dongo, L.N. Effects of fermentation intensity on polyphenols and antioxidant capacity of cocoa beans. Int. J. Sustain. Crop Prod. 2010, 5, 66–70. [Google Scholar]
- Suazo, Y.; Davidov-Pardo, G.; Arozarena, I. Effect of fermentation and roasting on the phenolic concentration and antioxidant activity of cocoa from Nicaragua. J. Food Qual. 2014, 37, 50–56. [Google Scholar] [CrossRef]
- Albertini, B.; Schoubben, A.; Guarnaccia, D.; Pinelli, F.; Della Vecchia, M.; Ricci, M.; Di Renzo, G.C.; Blasi, P. Effect of fermentation and drying on cocoa polyphenols. J. Agric. Food Chem. 2015, 63, 9948–9953. [Google Scholar] [CrossRef] [PubMed]
- Oracz, J.; Żyżelewicz, D.; Nebesny, E. The content of polyphenolic compounds in cocoa beans (Theobroma cacao L.), depending on variety, growing region, and processing operations: A review. Crit. Rev. Food Sci. Nutr. 2015, 55, 1176–1192. [Google Scholar] [CrossRef]
- Hernández-Hernández, C.; Viera-Alcaide, I.; Morales-Sillero, A.M.; Fernández-Bolaños, J.; Rodríguez-Gutiérrez, G. Bioactive compounds in Mexican genotypes of cocoa cotyledon and husk. Food Chem. 2018, 240, 831–839. [Google Scholar] [CrossRef] [Green Version]
- Urbańska, B.; Derewiaka, D.; Lenart, A.; Kowalska, J. Changes in the composition and content of polyphenols in chocolate resulting from pre-treatment method of cocoa beans and technological process. Eur. Food Res. Technol. 2019, 245, 2101–2112. [Google Scholar] [CrossRef] [Green Version]
- Camu, N.; De Winter, T.; Addo, K.S.; Takrama, J.S.; Bernaert, H.; De Vuyst, L. Fermentation of cocoa beans: Influence of microbial activities and polyphenol concentrations on the flavour of chocolate. J. Sci. Food Agric. 2008, 88, 2288–2297. [Google Scholar] [CrossRef]
- Peláez, P.; Bardón, I.; Camasca, P. Methylxanthine and catechin content of fresh and fermented cocoa beans, dried cocoa beans, and cocoa liquor. Sci. Agropecu. 2016, 7, 355–365. [Google Scholar] [CrossRef] [Green Version]
- Gil, M.; Uribe, D.; Gallego, V.; Bedoya, C.; Arango-Varela, S. Traceability of polyphenols in cocoa during the postharvest and industrialization processes and their biological antioxidant potential. Heliyon 2021, 7, e07738. [Google Scholar] [CrossRef]
- Hurst, W.J.; Krake, S.H.; Bergmeier, S.C.; Payne, M.J.; Miller, K.B.; Stuart, D.A. Impact of fermentation, drying, roasting and Dutch processing on flavan-3-ol stereochemistry in cacao beans and cocoa ingredients. Chem. Cent. J. 2011, 5, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giacometti, J.; Jolić, S.M.; Josić, D. Cocoa processing and impact on composition. In Processing and Impact on Active Components in Food; Preedy, V., Ed.; Academic Press: Cambridge, MA, USA, 2015; pp. 605–612. [Google Scholar] [CrossRef]
- Aprotosoaie, A.C.; Luca, S.V.; Miron, A. Flavor chemistry of cocoa and cocoa products—An overview. Compr. Rev. Food Sci. Food Saf. 2016, 15, 73–91. [Google Scholar] [CrossRef] [PubMed]
- Żyżelewicz, D.; Krysiak, W.; Oracz, J.; Sosnowska, D.; Budryn, G.; Nebesny, E. The influence of the roasting process conditions on the polyphenol content in cocoa beans, nibs and chocolates. Food Res. Int. 2016, 89, 918–929. [Google Scholar] [CrossRef]
- Stanley, T.H.; Van Buiten, C.B.; Baker, S.A.; Elias, R.J.; Anantheswaran, R.C.; Lambert, J.D. Impact of roasting on the flavan-3-ol composition, sensory-related chemistry, and in vitro pancreatic lipase inhibitory activity of cocoa beans. Food Chem. 2018, 255, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Oracz, J.; Nebesny, E. Effect of roasting parameters on the physicochemical characteristics of high-molecular-weight Maillard reaction products isolated from cocoa beans of different Theobroma cacao L. groups. Eur. Food Res. Technol. 2019, 245, 111–128. [Google Scholar] [CrossRef]
- Barišić, V.; Kopjar, M.; Jozinović, A.; Flanjak, I.; Ačkar, Đ.; Miličević, B.; Šubarić, D.; Jokić, S.; Babić, J. The chemistry behind chocolate production. Molecules 2019, 24, 3163. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Feng, Y.; Zhu, S.; Luo, C.; Ma, J.; Zhong, F. The effect of alkalization on the bioactive and flavor related components in commercial cocoa powder. J. Food Compos. Anal. 2012, 25, 17–23. [Google Scholar] [CrossRef]
- Valverde García, D.; Pérez Esteve, É.; Barat Baviera, J.M. Changes in cocoa properties induced by the alkalization process: A review. Compr. Rev. Food Sci. Food Saf. 2020, 19, 2200–2221. [Google Scholar] [CrossRef]
- Sulistyowati, M.; Misnawi, J. Effects of alkali concentration and conching temperature on antioxidant activity and physical properties of chocolate. Int. Food Res. J. 2008, 15, 297–304. [Google Scholar]
- Afoakwa, E.O.; Paterson, A.; Fowler, M.; Ryan, A. Flavor formation and character in cocoa and chocolate: A critical review. Crit. Rev. Food Sci. Nutr. 2008, 48, 840–857. [Google Scholar] [CrossRef]
- Di Mattia, C.D.; Martuscelli, M.; Sacchetti, G.; Beheydt, B.; Mastrocola, D.; Pittia, P. Effect of different conching processes on procyanidins content and antioxidant properties of chocolate. Food Res. Int. 2014, 63, 367–372. [Google Scholar] [CrossRef]
- Bordin Schumacher, A.B.; Brandelli, A.; Schumacher, E.W.; Macedo, F.C.; Pieta, L.; Klug, T.V. Development and evaluation of a laboratory-scale conch for chocolate production. Int. J. Food Sci. Technol. 2009, 44, 606–622. [Google Scholar] [CrossRef]
- Albak, F.; Tekin, A.R. Variation of total aroma and polyphenol content of dark chocolate during three phase of conching. J. Food Sci. Technol. 2016, 53, 848–855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gültekin-Ozgüven, M.; Berktas, I.; Ozçelik, B. Influence of processing conditions on procyanidin profiles and antioxidant capacity of chocolates: Optimization of dark chocolate manufacturing by response surface methodology. LWT Food Sci. Technol. 2016, 66, 252–259. [Google Scholar] [CrossRef]
- De Vuyst, L.; Weckx, S. The cocoa bean fermentation process: From ecosystem analysis to starter culture development. J. Appl. Microbiol. 2016, 121, 5–17. [Google Scholar] [CrossRef] [Green Version]
- Wollgast, J.; Anklam, E. Review on polyphenols in Theobroma cacao: Changes in composition during the manufacture of chocolate and methodology for identification and quantification. Food Res. Int. 2000, 33, 423–447. [Google Scholar] [CrossRef]
- Afoakwa, E.O. Chocolate Science and Technology, 1st ed.; Wiley-Blackwell Publishers: Oxford, UK, 2010; pp. 3–82. [Google Scholar]
- Thompson, S.S.; Miller, K.B.; Lopez, A.; Camu, N. Cocoa and coffee. In Food Microbiology: Fundamentals and Frontiers, 4th ed.; Doyle, M.P., Beuchat, R.L., Eds.; ASM Press: Washington, DC, USA, 2013; pp. 881–889. [Google Scholar]
- Prabhakaran Nair, K.P. Cocoa (Theobroma cacao L.). In The Agronomy and Economy of Important Tree Crops of the Developing World; Prabhakaran Nair, K.P., Ed.; Elsevier: Amsterdam, The Netherlands, 2010; Chapter 5; pp. 131–180. [Google Scholar] [CrossRef]
- Dzelagha, B.F.; Ngwa, N.M.; Nde Bup, D. A review of cocoa drying technologies and the effect on bean quality parameters. Int. J. Food Sci. 2020, 2020, 8830127. [Google Scholar] [CrossRef]
- Ackah, E.; Dompey, E. Effects of fermentation and drying durations on the quality of cocoa (Theobroma cacao L.) beans during the rainy season in the Juaboso District of the Western-North Region, Ghana. Bull. Natl. Res. Cent. 2021, 45, 175. [Google Scholar] [CrossRef]
- Zahouli, G.I.B.; Guehi, S.T.; Fae, A.M.; Nemlin, J.G. Effect of drying methods on the chemical quality traits of cocoa raw material. Adv. J. Food Sci. Technol. 2010, 2, 184–190. [Google Scholar]
- Saltini, R.; Akkerman, R.; Frosc, S. Optimizing chocolate production through traceability: A review of the influence of farming practices on cocoa bean quality. Food Control 2013, 29, 167–187. [Google Scholar] [CrossRef] [Green Version]
- Bharath, S.; Bowen-O’Connor, C. Assessing Drying Rates of Cacao Beans Using Small Samples; U.W.I. Annual Report; Cocoa Research Unit: St. Augustine, Trinidad and Tobago, 2008; pp. 52–58. Available online: https://www.worldcocoafoundation.org/wp-content/uploads/files_mf/bharath2008.pdf (accessed on 20 July 2022).
- Kamphuis, H.J. Production and quality standards of cocoa mass, cocoa butter and cocoa powder. In Industrial Chocolate Manufacture and Use, 4th ed.; Beckett, S.T., Ed.; John Wiley & Sons: Chichester, UK, 2009; pp. 121–139. [Google Scholar] [CrossRef]
- Moser, A. Alkalizing cocoa and chocolate. Manuf. Confect. 2015, 95, 31–38. [Google Scholar]
- Sommer, J. Developments in cocoa bean processing. Manuf. Confect. 2009, 89, 99. [Google Scholar]
- Sioriki, E.; Tuenter, E.; Van de Walle, D.; Lemarcq, V.; Cazin, C.S.; Nolan, S.P.; Pieters, L.; Dewettinck, K. The effect of cocoa alkalization on the non-volatile and volatile mood-enhancing compounds. Food Chem. 2022, 381, 132082. [Google Scholar] [CrossRef]
- Quelal-Vásconez, M.A.; Lerma-García, M.J.; Pérez-Esteve, É.; Arnau-Bonachera, A.; Barat, J.M.; Talens, P. Changes in methylxanthines and flavanols during cocoa powder processing and their quantification by near-infrared spectroscopy. LWT Food Sci. Technol. 2020, 117, 108598. [Google Scholar] [CrossRef]
- Dyer, B. Alkalized cocoa powders. In Proceedings of the 57th PMCA Production Conference, Hershey, PA, USA, 28–30 April 2003. [Google Scholar]
- Ziegleder, G. Flavour development in cocoa and chocolate. In Industrial Chocolate Manufacture and Use, 5th ed.; Beckett, S.T., Fowler, M.S., Ziegler, G.R., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2017; pp. 185–215. [Google Scholar]
- Misnawi, J.; Febrianto, N.; Ariza, B.T.S. Roasting equipment for cocoa processing. In Drying and Roasting of Cocoa and Coffee; Hii, C.L., Borém, F.M., Eds.; CRC Press—Taylor & Francis Group: Boca Raton, FL, USA, 2020; pp. 47–62. [Google Scholar]
- Krysiak, W.; Adamski, R.; Żyżelewicz, D. Factors affecting the colour of roasted cocoa bean. J. Food Qual. 2013, 36, 21–31. [Google Scholar] [CrossRef] [Green Version]
- Beckett, S.T. Traditional chocolate making. In Industrial Chocolate Manufacture and Use, 5th ed.; Beckett, S.T., Fowler, M.S., Ziegler, G.R., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2017; pp. 1–9. [Google Scholar]
- Kothe, L.; Zimmermann, B.F.; Galensa, R. Temperature influences epimerization and composition of flavanol monomers, dimers and trimers during cocoa bean roasting. Food Chem. 2013, 141, 3656–3663. [Google Scholar] [CrossRef]
- Ioannone, F.; Di Mattia, C.; De Gregorio, M.; Sergi, M.; Serafini, M.; Sacchetti, G. Flavanols, proanthocyanidins and antioxidant activity changes during cocoa (Theobroma cacao L.) roasting as affected by temperature and time of processing. Food Chem. 2015, 174, 256–262. [Google Scholar] [CrossRef]
- Stadler, R.H.; Hughes, G.; Guillaume-Gentil, O. Safety of food and beverages: Coffee, tea and herbals, cocoa and derived products. In Encyclopedia of Food Safety; Motarjemi, Y., Moy, G.G., Todd, E.C.D., Eds.; Elsevier: Amsterdam, The Netherlands, 2014; Volume 3, pp. 371–383. [Google Scholar] [CrossRef]
- Biehl, B.; Ziegleder, G. Cocoa | Production, products, and use. In Encyclopedia of Food Sciences and Nutrition, 2nd ed.; Caballero, B., Ed.; Academic Press: Cambridge, MA, USA, 2003; pp. 1448–1463. [Google Scholar]
- Velthuis, T. The nib-grinding process. Manuf. Confect. 2009, 89, 95. [Google Scholar]
- Anoraga, S.B.; Wijanarti, S.; Sabarisman, I. Effect of extraction time and pressing temperature on characteristic of cocoa powder quality. IOP Conf. Ser. Earth Environ. Sci. 2019, 355, 012050. [Google Scholar] [CrossRef]
- Gutiérrez, T.J. State-of-the-art chocolate manufacture: A review. Compr. Rev. Food Sci. Food Saf. 2017, 16, 1313–1344. [Google Scholar] [CrossRef] [Green Version]
- Owusu, M.; Petersen, M.A.; Heimdal, H. Effect of fermentation method roasting and conching conditions on the aroma volatiles of dark chocolate. J. Food Process. Preserv. 2012, 36, 446–456. [Google Scholar] [CrossRef]
- Toker, O.S.; Palabiyik, I.; Konar, N. Chocolate quality and conching. Trends Food Sci. Technol. 2019, 91, 446–453. [Google Scholar] [CrossRef]
- Urbańska, B.; Kowalska, H.; Szulc, K.; Ziarno, M.; Pochitskaya, I.; Kowalska, J. Comparison of the effects of conching parameters on the contents of three dominant flavan3-ols, rheological properties and sensory quality in chocolate milk mass based on liquor from unroasted cocoa beans. Molecules 2021, 26, 2502. [Google Scholar] [CrossRef]
- Konar, N. Influence of conching temperature and some bulk sweeteners on physical and rheological properties of prebiotic milk chocolate including containing inulin. Eur. Food Res. Technol. 2013, 23, 135–143. [Google Scholar] [CrossRef]
- Owusu, M.; Petersen, M.A.; Heimdal, H. Relationship of sensory and instrumental aroma measurements of dark chocolate as influenced by fermentation method, roasting and conching conditions. J. Food Sci. Technol. 2013, 50, 909–917. [Google Scholar] [CrossRef] [Green Version]
- Engeseth, N.J.; Pangan, M.F.A. Current context on chocolate flavor development—A review. Curr. Opin. Food Sci. 2018, 21, 84–91. [Google Scholar] [CrossRef]
- Tiefenbacher, K.F. Adjuncts—Filling creams, inclusions, cacao and chocolate. In Wafer and Waffle; Tiefenbacher, K.F., Ed.; Academic Press: Cambridge, MA, USA, 2017; Chapter 5; pp. 313–404. [Google Scholar]
- Caligiani, A.; Marseglia, A.; Palla, G. Cocoa: Production, chemistry, and use. In The Encyclopedia of Food and Health; Caballero, B., Finglas, P.M., Toldrá, F., Eds.; Academic Press: Cambridge, MA, USA, 2016; Volume 5, pp. 185–190. [Google Scholar] [CrossRef]
- Martín, M.A.; Goya, L.; Ramos, S. Anti-diabetic actions of cocoa flavonoids. Mol. Nutr. Food Res. 2016, 60, 1756–1769. [Google Scholar] [CrossRef]
- Granado-Serrano, A.B.; Martín, M.A.; Izquierdo-Pulido, M.; Goya, L.; Bravo, L.; Ramos, S. Molecular mechanisms of (−)-epicatechin and chlorogenic acid on the regulation of the apoptotic and survival/proliferation pathways in a human hepatoma cell line (HepG2). J. Agric. Food Chem. 2007, 55, 2020–2027. [Google Scholar] [CrossRef]
- Granado-Serrano, A.B.; Martín, M.A.; Goya, L.; Bravo, L.; Ramos, S. Time-course regulation of survival pathways by epicathechin on HepG2. J. Nutr. Biochem. 2009, 20, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Martín, M.A.; Granado-Serrano, A.B.; Ramos, S.; Izquierdo-Pulido, M.; Bravo, L.; Goya, L. Cocoa flavonoids up-regulate antioxidant enzymes activity via ERK1/2 pathway to protect against oxidative stress-induced apoptosis in HepG2 cells. J. Nutr. Biochem. 2010, 21, 196–205. [Google Scholar] [CrossRef]
- Granado-Serrano, A.B.; Martín, M.A.; Haegeman, G.; Goya, L.; Bravo, L.; Ramos, S. Epicatechin induces NF-κB, activator protein-1 (AP-1) and nuclear transcription factor erythroid 2p45-related (Nrf2) via phosphatidylinositol-3-kiinase/protein kinase B (PI3K/AKT) and extracellular regulated kinase (ERK) signalling in HepG2 cells. Brit. J. Nutr. 2010, 103, 168–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Ramiro, I.; Ramos, S.; Bravo, L.; Goya, L.; Martín, M.A. Procyanidin B2 and a cocoa polyphenolic extract inhibit acrylamide-induced apoptosis in human Caco-2 cells by preventing oxidative stress and activation of JNK pathway. J. Nutr. Biochem. 2011, 22, 1186–1194. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Ramiro, I.; Ramos, S.; Bravo, L.; Goya, L.; Martín, M.A. Procyanidin B2 induces Nrf2 translocation and glutathione-S-transferase P1 expression and via ERKs and p38-MAPK pathways and protect human colonic cells against oxidative stress. Eur. J. Nutr. 2012, 51, 881–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cordero-Herrera, I.; Martín, M.A.; Bravo, L.; Goya, L.; Ramos, S. Cocoa flavonoids improve insulin signalling and repress glucose production via AKT and AMPK in HepG2 cells. Mol. Nutr. Food Res. 2013, 57, 974–985. [Google Scholar] [CrossRef] [Green Version]
- Cordero-Herrera, I.; Martín, M.A.; Goya, L.; Ramos, S. Cocoa flavonoids protect hepatic cells against high glucose-induced oxidative stress: Relevance of MAPKs. Mol. Nutr. Food Res. 2015, 59, 597–609. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Millán, E.; Ramos, S.; Alvarez, C.; Bravo, L.; Goya, L.; Martín, M.A. Microbial phenolic metabolites improve glucose-stimulated insulin secretion and protect pancreatic beta cells against oxidative stress via ERKs and PKC pathways. Food Chem. Toxicol. 2014, 66, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Cilleros, D.; Martín, M.A.; Goya, L.; Ramos, S. (−)-Epicatechin and the colonic metabolite 3,4-dihydroxyphenylacetic acid protect renal proximal tubular cell against high glucose-induced oxidative stress by modulating NOX-4/SIRT-1 signalling. J. Funct. Foods 2018, 46, 19–28. [Google Scholar] [CrossRef] [Green Version]
- Zhihao, Q.; Ailing, L.; Penghui, L.; Changwei, L.; Wenjun, X.; Jianan, H.; Zhonghua, L.; Sheng, Z. Advances in physiological functions and mechanisms of (−)-epicatechin. Crit. Rev. Food Sci. Nutr. 2021, 61, 211–233. [Google Scholar] [CrossRef]
- Gao, M.; Peng, X.; Tang, J.; Deng, J.; Wang, F.; Zhang, Y.; Zhao, P.; Kan, H.; Liu, Y. Anti-Inflammatory Effects of Camellia fascicularis Polyphenols via Attenuation of NF-κB and MAPK Pathways in LPS-Induced THP-1 Macrophages. J. Inflamm. Res. 2022, 15, 851–864. [Google Scholar] [CrossRef]
- Kim, J.K.; Yang, H.J.; Go, Y. Quercus acuta Thunb. Suppresses LPS-Induced Neuroinflammation in BV2 Microglial Cells via Regulating MAPK/NF-κB and Nrf2/HO-1 Pathway. Antioxidants 2022, 11, 1851. [Google Scholar] [CrossRef]
- Huang, P.; Hong, J.; Mi, J.; Sun, B.; Zhang, J.; Li, C.; Yang, W. Polyphenols extracted from Enteromorpha clathrata alleviates inflammation in lipopolysaccharide-induced RAW 264.7 cells by inhibiting the MAPKs/NF-κB signaling pathways. J. Ethnopharmacol. 2022, 286, 114897. [Google Scholar] [CrossRef]
- Khan, A.; Khan, S.U.; Khan, A.; Shal, B.; Rehman, S.U.; Rehman, S.U.; Htar, T.T.; Khan, S.; Anwar, S.; Alafnan, A.; et al. Anti-inflammatory and anti-rheumatic potential of selective plant compounds by targeting TLR-4/AP-1 signaling: A comprehensive molecular docking and simulation approaches. Molecules 2022, 27, 4319. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Lu, J.; Wang, Z.; Wang, L.; Wu, G.; Guo, Y.; Dong, Z. Small ubiquitin-related modifier (SUMO)ylation of SIRT1 mediates (−)-epicatechin inhibited- differentiation of cardiac fibroblasts into myofibroblasts. Pharm. Biol. 2022, 60, 1762–1770. [Google Scholar] [CrossRef] [PubMed]
- Behl, T.; Upadhyay, T.; Singh, S.; Chigurupati, S.; Alsubayiel, A.M.; Mani, V.; Vargas-De-La-Cruz, C.; Uivarosan, D.; Bustea, C.; Sava, C.; et al. Polyphenols targeting MAPK mediated oxidative stress and inflammation in rheumatoid arthritis. Molecules 2021, 26, 6570. [Google Scholar] [CrossRef] [PubMed]
- Cháirez-Ramírez, M.H.; de la Cruz-López, K.G.; García-Carrancá, A. Polyphenols as antitumor agents targeting key players in cancer-driving signaling pathways. Front. Pharmacol. 2021, 12, 710304. [Google Scholar] [CrossRef] [PubMed]
- Anjum, J.; Mitra, S.; Das, R.; Alam, R.; Mojumder, A.; Emran, T.B.; Islam, F.; Rauf, A.; Hossain, J.; Aljohani, A.S.M.; et al. A renewed concept on the MAPK signaling pathway in cancers: Polyphenols as a choice of therapeutics. Pharmacol. Res. 2022, 184, 106398. [Google Scholar] [CrossRef] [PubMed]
- Behl, T.; Mehta, K.; Sehgal, A.; Singh, S.; Sharma, N.; Ahmadi, A.; Arora, S.; Bungau, S. Exploring the role of polyphenols in rheumatoid arthritis. Crit. Rev. Food Sci. Nutr. 2022, 62, 5372–5393. [Google Scholar] [CrossRef]
- Hazafa, A.; Iqbal, M.O.; Javaid, U.; Tareen, M.B.K.; Amna, D.; Ramzan, A.; Piracha, S.; Naeem, M. Inhibitory effect of polyphenols (phenolic acids, lignans and stilbenes) on cancer by regulating signal transduction pathways: A review. Clin. Transl. Oncol. 2022, 24, 432–445. [Google Scholar] [CrossRef]
- Zheng, Z.; Zhang, L.; Hou, X. Potential roles and molecular mechanisms of phytochemicals against cancer. Food Funct. 2022, 13, 9208–9225. [Google Scholar] [CrossRef]
- Bernatoniene, J.; Kopustinskiene, D.M. The role of catechins in cellular responses to oxidative stress. Molecules 2018, 23, 965. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Iglesias, P.; Massot-Cladera, M.; Rodríguez-Lagunas, M.J.; Franch, A.; Camps-Bossacoma, M.; Pérez-Cano, F.J.; Castell, M. Protective effect of a cocoa-enriched diet on oxidative stress induced by intensive acute exercise in rats. Antioxidants 2022, 11, 753. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.E. The effects of dietary supplements that overactivate the Nrf2/ARE system. Curr. Med. Chem. 2020, 27, 2077–2094. [Google Scholar] [CrossRef] [PubMed]
- Bahia, P.; Rattray, M.; Williams, R. Dietary flavonoid (−) epicatechin stimulates phosphatidylinositol 3-kinase-dependent anti-oxidant response element activity and up-regulates glutathione in cortical astrocytes. J. Neurochem. 2008, 106, 2194–2204. [Google Scholar] [CrossRef]
- Cheng, Y.T.; Wu, C.H.; Ho, C.Y.; Yen, G.C. Catechin protects against ketoprofen-induced oxidative damage of the gastric mucosa by up-regulating Nrf2 in vitro and in vivo. J. Nutr. Biochem. 2013, 24, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Nichols, M.; Zhang, J.; Polster, B.M.; Elustondo, P.A.; Thirumaran, A.; Pavlov, E.V.; Robertson, G.S. Synergistic neuroprotection by epicatechin and quercetin: Activation of convergent mitochondrial signaling pathways. Neuroscience 2015, 308, 75–94. [Google Scholar] [CrossRef]
- Ramírez-Sánchez, I.; Rodríguez, A.; Ulloa, A.M.; Ceballos, G.; Villarreal, F. (−)-Epicatechin-induced recovery of mitochondria from simulated diabetes: Potential role of endothelial nitric oxide synthase. Diabetes Vasc. Dis. Res. 2016, 13, 201–210. [Google Scholar] [CrossRef] [Green Version]
- Rowley, T.J., IV; Bitner, B.F.; Ray, J.D.; Lathen, D.R.; Smithson, A.T.; Dallon, B.W.; Plowman, C.J.; Bikman, B.T.; Hansen, J.M.; Dorenkott, M.R.; et al. Monomeric cocoa catechins enhance β-cell function by increasing mitochondrial respiration. J. Nutr. Biochem. 2017, 49, 30–41. [Google Scholar] [CrossRef]
- Sapian, S.; Taib, I.S.; Latip, J.; Katas, H.; Chin, K.Y.; Nor, N.A.M.; Jubaidi, F.F.; Budin, S.B. Therapeutic Approach of Flavonoid in Ameliorating Diabetic Cardiomyopathy by Targeting Mitochondrial-Induced Oxidative Stress. Int. J. Mol. Sci. 2021, 22, 11616. [Google Scholar] [CrossRef]
- Hamsalakshmi, A.M.A.; Mahalakshmi, A.M.; Joghee, S.; Chidambaram, S.B. Therapeutic benefits of flavonoids against neuroinflammation: A systematic review. Inflammopharmacology 2022, 30, 111–136. [Google Scholar] [CrossRef]
- Clifford, T.; Acton, J.P.; Cocksedge, S.P.; Bowden Davies, K.A.; Bailey, S.J. The effect of dietary phytochemicals on nuclear factor erythroid 2-related factor 2 (Nrf2) activation: A systematic review of human intervention trials. Mol. Biol. Rep. 2021, 48, 1745–1761. [Google Scholar] [CrossRef]
- Bustamante-Pozo, M.; Haro, M.; Garcia, R.; Carson, N.; Ceballos, G.; Villarreal, F. Epicatechin ameliorates cardiac fibrosis in a female rat model of pre-heart failure with preserved ejection fraction. J. Med. Food 2022, 25, 836–844. [Google Scholar] [CrossRef] [PubMed]
- Daussin, F.N.; Cuillerier, A.; Touron, J.; Bensaid, S.; Melo, B.; Rewashdy, A.A.; Vasam, G.; Menzies, K.J.; Harper, M.E.; Heyman, E.; et al. Dietary cocoa flavanols enhance mitochondrial function in skeletal muscle and modify whole-body metabolism in healthy mice. Nutrients 2021, 13, 3466. [Google Scholar] [CrossRef] [PubMed]
- García-Díez, E.; López-Oliva, M.E.; Caro-Vadillo, A.; Pérez-Vizcaíno, F.; Pérez-Jiménez, J.; Ramos, S.; Martín, M.A. Supplementation with a cocoa–carob blend, alone or in combination with metformin, attenuates diabetic cardiomyopathy, cardiac oxidative stress and inflammation in zucker diabetic rats. Antioxidants 2022, 11, 432. [Google Scholar] [CrossRef] [PubMed]
- García-Merino, J.A.; de Lucas, B.; Herrera-Rocha, K.; Moreno-Pérez, D.; Montalvo-Lominchar, M.G.; Fernández-Romero, A.; Santiago, C.; Pérez-Ruiz, M.; Larrosa, M. Flavanol-rich cocoa supplementation inhibits mitochondrial biogenesis triggered by exercise. Antioxidants 2022, 11, 1522. [Google Scholar] [CrossRef]
- Muralidhara, K.G. Dietary Supplements as Cognitive Enhancers: The Role of Flavonoid-Rich Foods and their Relevance in Age-Related Neurodegeneration. In Bioactive Nutraceuticals and Dietary Supplements in Neurological and Brain Disease; Watson, R.R., Preedy, V.R., Eds.; Academic Press: Cambridge, MA, USA, 2015; pp. 281–290. [Google Scholar]
- Vauzour, D.; Camprubi-Robles, M.; Miquel-Kergoat, S.; Andres-Lacueva, C.; Bánáti, D.; Barberger-Gateau, P.; Bowman, G.L.; Caberlotto, L.; Clarke, R.; Hogervorst, E.; et al. Nutrition for the ageing brain: Towards evidence for an optimal diet. Ageing Res. Rev. 2017, 35, 222–240. [Google Scholar] [CrossRef] [Green Version]
- Dominguez, L.J.; Barbagallo, M. Nutritional prevention of cognitive decline and dementia. Acta Biomed. 2018, 89, 276–290. [Google Scholar] [CrossRef]
- Socci, V.; Tempesta, D.; Desideri, G.; De Gennaro, L.; Ferrara, M. Enhancing human cognition with cocoa flavonoids. Front. Nutr. 2017, 4, 19. [Google Scholar] [CrossRef] [Green Version]
- Haskell-Ramsay, C.F.; Schmitt, J.; Actis-Goretta, L. The impact of epicatechin on human cognition: The role of cerebral blood flow. Nutrients 2018, 10, 986. [Google Scholar] [CrossRef] [Green Version]
- Abd El Mohsen, M.M.; Kuhnle, G.; Rechner, A.R.; Schroeter, H.; Rose, S.; Jenner, P.; Rice-Evans, C.A. Uptake and metabolism of epicatechin and its access to the brain after oral ingestion. Free Radic. Biol. Med. 2002, 33, 1693–1702. [Google Scholar] [CrossRef]
- Francis, S.T.; Head, K.; Morris, P.G.; Macdonald, I.A. The effect of flavanol-rich cocoa on the fMRI response to a cognitive task in healthy young people. J. Cardiovasc. Pharmacol. 2006, 47 (Suppl. S2), S215–S220. [Google Scholar] [CrossRef]
- Kelleher, R.J., III; Govindarajan, A.; Jung, H.Y.; Kang, H.; Tonegawa, S. Translational control by MAPK signaling in long-term synaptic plasticity and memory. Cell 2004, 116, 467–479. [Google Scholar] [CrossRef] [Green Version]
- Schroeter, H.; Bahia, P.; Spencer, J.P.; Sheppard, O.; Rattray, M.; Cadenas, E.; Rice-Evans, C.; Williams, R.J. (−)Epicatechin stimulates ERK-dependent cyclic AMP response element activity and upregulates GluR2 in cortical neurons. J. Neurochem. 2007, 101, 1596–1606. [Google Scholar] [CrossRef] [PubMed]
- Desideri, G.; Kwik-Uribe, C.; Grassi, D.; Necozione, S.; Ghiadoni, L.; Mastroiacovo, D.; Ferri, C. Benefits in cognitive function, blood pressure, and insulin resistance through cocoa flavanol consumption in elderly subjects with mild cognitive impairment: The Cocoa, Cognition, and Aging (CoCoA) study. Hypertension 2012, 60, 794–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pase, M.P.; Scholey, A.B.; Pipingas, A.; Kras, M.; Nolidin, K.; Gibbs, A.; Stough, C. Cocoa polyphenols enhance positive mood states but not cognitive performance: A randomized, placebo-controlled trial. J. Psychopharmacol. 2013, 27, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Lamport, D.J.; Pal, D.; Moutsiana, C.; Field, D.T.; Williams, C.M.; Spencer, J.P.; Butler, L.T. The effect of flavanol-rich cocoa on cerebral perfusion in healthy older adults during conscious resting state: A placebo controlled, crossover, acute trial. J. Psychopharmacol. 2015, 232, 3227–3234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ammar, A.; Trabelsi, K.; Müller, P.; Bouaziz, B.; Boukhris, O.; Glenn, J.M.; Bott, N.T.; Driss, T.; Chtourou, H.; Müller, N.; et al. The effect of (poly)phenol-rich interventions on cognitive functions and neuroprotective measures in healthy aging adults: A systematic review and meta-analysis. J. Clin. Med. 2020, 9, 835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrera-Reyes, P.K.; De Lara, J.C.-F.; González-Soto, M.; Tejero, M. Effects of cocoa-derived polyphenols on cognitive function in humans. Systematic review and analysis of methodological aspects. Plant Foods Hum. Nutr. 2020, 75, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Di Meo, F.; Valentino, A.; Petillo, O.; Peluso, G.; Filosa, S.; Crispi, S. Bioactive polyphenols and neuromodulation: Molecular mechanisms in neurodegeneration. Int. J. Mol. Sci. 2020, 21, 2564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flanagan, E.; Lamport, D.; Brennan, L.; Burnet, P.; Calabrese, V.; Cunnane, S.C.; de Wilde, M.C.; Dye, L.; Farrimond, J.A.; Emerson Lombardo, N.; et al. Nutrition and the ageing brain: Moving towards clinical applications. Ageing Res. Rev. 2020, 62, 101079. [Google Scholar] [CrossRef] [PubMed]
- Martín, M.A.; Goya, L.; de Pascual-Teresa, S. Effect of cocoa and cocoa products on cognitive performance in young adults. Nutrients 2020, 12, 3691. [Google Scholar] [CrossRef]
- Vinciguerra, F.; Graziano, M.; Hagnäs, M.; Frittitta, L.; Tumminia, A. Influence of the Mediterranean and ketogenic diets on cognitive status and decline: A narrative review. Nutrients 2020, 12, 1019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, J.; Wang, Z.; Oteiza, P.I. (−)-Epicatechin mitigates high fat diet-induced neuroinflammation and altered behavior in mice. Food Funct. 2020, 11, 5065–5076. [Google Scholar] [CrossRef] [PubMed]
- Gratton, G.; Weaver, S.R.; Burley, C.V.; Low, K.A.; Maclin, E.L.; Johns, P.W.; Pham, Q.S.; Lucas, S.J.E.; Fabiani, M.; Rendeiro, C. Dietary flavanols improve cerebral cortical oxygenation and cognition in healthy adults. Sci. Rep. 2020, 10, 19409. [Google Scholar] [CrossRef] [PubMed]
- Baynham, R.; Veldhuijzen van Zanten, J.J.C.S.; Johns, P.W.; Pham, Q.S.; Rendeiro, C. Cocoa flavanols improve vascular responses to acute mental stress in young healthy adults. Nutrients 2021, 13, 1103. [Google Scholar] [CrossRef]
- Navarrete-Yañez, V.; Garate-Carrillo, A.; Ayala, M.; Rodriguez-Castañeda, A.; Mendoza-Lorenzo, P.; Ceballos, G.; Ordoñez-Razo, R.; Dugar, S.; Schreiner, G.; Villarreal, F.; et al. Stimulatory effects of (−)-epicatechin and its enantiomer (+)-epicatechin on mouse frontal cortex neurogenesis markers and short-term memory: Proof of concept. Food Funct. 2021, 12, 3504. [Google Scholar] [CrossRef]
- García-Cordero, J.; Pino, A.; Cuevas, C.; Puertas-Martín, V.; San Román, R.; de Pascual-Teresa, S. Neurocognitive effects of cocoa and red-berries consumption in healthy adults. Nutrients 2022, 14, 1. [Google Scholar] [CrossRef]
- Caruso, G.; Torrisi, S.A.; Mogavero, M.P.; Currenti, W.; Castellano, S.; Godos, J.; Ferri, R.; Galvano, F.; Leggio, G.M.; Grosso, G.; et al. Polyphenols and neuroprotection: Therapeutic implications for cognitive decline. Pharmacol. Ther. 2022, 232, 108013. [Google Scholar] [CrossRef]
- Lalonde, R.; Strazielle, C. Cocoa flavanols and the aging brain. Curr. Aging Sci. 2022, 15. [Google Scholar] [CrossRef]
- Suominen, M.H.; Laaksonen, M.M.L.; Salmenius-Suominena, H.; Kautiainen, H.; Hongisto, S.M.; Tuukkanen, K.; Jyväkorpi, S.K.; Pitkälä, K.H. The short-term effect of dark chocolate flavanols on cognition in older adults: A randomized controlled trial (FlaSeCo). Exp. Gerontol. 2020, 136, 110933. [Google Scholar] [CrossRef]
- Baker, L.D.; Manson, J.E.; Rapp, S.R.; Sesso, H.D.; Gaussoin, S.A.; Shumaker, S.A.; Espeland, M.A. Effects of cocoa extract and a multivitamin on cognitive function: A randomized clinical trial. Alzheimer Dement. 2022, 1–12. [Google Scholar] [CrossRef]
- Shishtar, E.; Rogers, G.T.; Blumberg, J.B.; Au, R.; Jacques, P.F. Long-term dietary flavonoid intake and change in cognitive function in the Framingham Offspring Cohort. Public Health Nutr. 2020, 23, 1576–1588. [Google Scholar] [CrossRef] [PubMed]
- Gardener, S.L.; Rainey-Smith, S.R.; Weinborn, M.; Bondonno, C.P.; Martins, R.N. Intake of products containing anthocyanins, flavanols, and flavanones, and cognitive function: A narrative review. Front. Aging Neurosci. 2021, 13, 640381. [Google Scholar] [CrossRef] [PubMed]
- Lamport, D.J.; Williams, C.M. Polyphenols and cognition in humans: An overview of current evidence from recent systematic reviews and meta-analyses. Brain Plast. 2021, 6, 139–153. [Google Scholar] [CrossRef] [PubMed]
- Rodrigo-Gonzalo, M.J.; González-Manzano, S.; Mendez-Sánchez, R.; Santos-Buelga, C.; Recio-Rodríguez, J.I. Effect of polyphenolic complements on cognitive function in the elderly: A systematic review. Antioxidants 2022, 11, 1549. [Google Scholar] [CrossRef]
- Zeli, C.; Lombardo, M.; Storz, M.A.; Ottaviani, M.; Rizzo, G. Chocolate and cocoa-derived biomolecules for brain cognition during ageing. Antioxidants 2022, 11, 1353. [Google Scholar] [CrossRef]
- European Food Safety Authority. Scientific Opinion on the substantiation of health claims related to cocoa flavanols and protection of lipids from oxidative damage and maintenance of normal blood pressure. EFSA J. 2010, 8, 1792. [Google Scholar]
- European Food Safety Authority. Scientific opinion on the substantiation of a health claim related to cocoa flavanols and maintenance of normal endothelium-dependent vasodilation. EFSA J. 2012, 10, 2809. [Google Scholar]
- Sun, Y.; Zimmermann, D.; De Castro, C.A.; Actis-Goretta, L. Dose-response relationship between cocoa flavanols and human endothelial function: A systematic review and meta-analysis of randomized trials. Food Funct. 2019, 10, 6322. [Google Scholar] [CrossRef] [Green Version]
- Kris-Etherton, P.M.; Keen, C.L. Evidence that the antioxidant flavonoids in tea and cocoa are beneficial for cardiovascular health. Curr. Opin. Lipidol. 2002, 13, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Heiss, C.; Dejam, A.; Kleinbongard, P.; Schewe, T.; Sies, H.; Kelm, M. Vascular effects of cocoa rich in flavan-3-ols. J. Am. Med. Assoc. 2003, 290, 1030–1031. [Google Scholar] [CrossRef]
- Fisher, N.D.; Hughes, M.; Gerhard-Herman, M.; Hollenberg, N.K. Flavanol rich cocoa induces nitric oxide-dependent vasodilation in healthy humans. J. Hypertens. 2003, 1, 2281–2286. [Google Scholar] [CrossRef] [PubMed]
- Hollenberg, N.K. Vascular action of cocoa flavanols in humans: The roots of the story. J. Cardiovasc. Pharmacol. 2006, 47, S99–S102. [Google Scholar] [CrossRef] [PubMed]
- Schroeter, H.; Heiss, C.; Balzer, J.; Kleinbongard, P.; Keen, C.L.; Hollenberg, N.K.; Sies, H.; Kwik-Uribe, C.; Schmitz, H.H.; Kelm, M. (−)-Epicatechin mediates beneficial effects of flavanol-rich cocoa on vascular function in humans. Proc. Natl. Acad. Sci. USA 2006, 3, 1024–1029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steffen, Y.; Schewe, T.; Sies, H. (−)-Epicatechin elevates nitric oxide in endothelial cells via inhibition of NADPH oxidase. Biochem. Biophys. Res. Commun. 2007, 359, 828–833. [Google Scholar] [CrossRef] [PubMed]
- Taubert, D.; Roesen, R.; Lehmann, C.; Jung, N.; Schömig, E. Effects of low habitual cocoa intake on blood pressure and bioactive nitric oxide. J. Am. Med. Assoc. 2007, 298, 49–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balzer, J.; Rassaf, T.; Heiss, C.; Kleinbongard, P.; Lauer, T.; Merx, M.; Heussen, N.; Gross, H.B.; Keen, C.L.; Schroeter, H.; et al. Sustained benefits in vascular function through flavanol-containing cocoa in medicated diabetic patients. a double-masked, randomized, controlled trial. J. Am. Coll. Cardiol. 2008, 51, 2141–2149. [Google Scholar] [CrossRef] [Green Version]
- Faridi, Z.; Njike, V.Y.; Dutta, S.; Ali, A.; Katz, D.L. Acute dark chocolate and cocoa ingestion and endothelial function: A randomized controlled crossover trial. Am. J. Clin. Nutr. 2008, 8, 58–63. [Google Scholar] [CrossRef] [Green Version]
- Grassi, D.; Desideri, G.; Necozione, S.; Lippi, C.; Casale, R.; Properzi, G.; Blumberg, J.B.; Ferri, C. Blood pressure is reduced and insulin sensitivity increased in glucose-intolerant, hypertensive subjects after 15 days of consuming high polyphenol dark chocolate. J. Nutr. 2008, 38, 1671–1676. [Google Scholar] [CrossRef] [Green Version]
- Hooper, L.; Kroon, P.A.; Rimm, E.B.; Cohn, J.S.; Harvey, I.; Le Cornu, K.A.; Ryder, J.J.; Hall, W.L.; Cassidy, A. Flavonoids, flavonoid-rich foods, and cardiovascular risk: A meta-analysis of randomized controlled trials. Am. J. Clin. Nutr. 2008, 88, 38–50. [Google Scholar] [CrossRef] [Green Version]
- Ried, K.; Sullivan, T.R.; Fakler, P.; Frank, O.; Stocks, N. Does chocolate reduce blood pressure? A meta-analysis. BMC Med. 2010, 8, 39. [Google Scholar] [CrossRef] [Green Version]
- Ried, K.; Sullivan, T.R.; Fakler, P.; Frank, O.; Stocks, N.P. Effect of cocoa on blood pressure. Cochrane Database Syst. Rev. 2012, 15, CD008893. [Google Scholar] [CrossRef]
- de Pascual-Teresa, S.; Clifford, M.N. Advances in polyphenol research: A journal of agricultural and food chemistry virtual issue. J. Agric. Food Chem. 2017, 65, 8093–8095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludovici, V.; Barthelmes, J.; Nägele, M.P.; Enseleit, F.; Ferri, C.; Flammer, A.J.; Ruschitzka, F.; Sudano, I. Cocoa, blood pressure, and vascular function. Front. Nutr. 2017, 4, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dugo, L.; Tripodo, G.; Santi, L.; Fanali, C. Cocoa polyphenols: Chemistry, bioavailability and effects on cardiovascular performance. Curr. Med. Chem. 2018, 25, 4903–4917. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D.; Wu, J.H.Y. Flavonoids, dairy foods, and cardiovascular and metabolic health. A review of emerging biologic pathways. Circ Res. 2018, 122, 369–384. [Google Scholar] [CrossRef] [PubMed]
- Ciumarnean, L.; Milaciu, M.V.; Runcan, O.; Vesa, S.C.; Rachis, A.L.; Negrean, V.; Perné, M.-G.; Donca, V.I.; Alexescu, T.-G.; Para, I. The effects of flavonoids in cardiovascular diseases. Molecules 2020, 25, 4320. [Google Scholar] [CrossRef] [PubMed]
- Ebaditabar, M.; Djafarian, K.; Saeidifard, N.; Shab-Bidar, S. Effect of dark chocolate on flow-mediated dilatation: Systematic review, meta-analysis, and dose–response analysis of randomized controlled trials. Clin. Nutr. ESPEN 2020, 36, 17–27. [Google Scholar] [CrossRef]
- Martin, M.A.; Ramos, S. Impact of cocoa flavanols on human health. Food Chem. Toxicol. 2021, 151, 112121. [Google Scholar] [CrossRef]
- Azad, B.J.; Daneshzad, E.; Meysamie, A.P.; Koohdani, F. Chronic and acute effects of cocoa products intake on arterial stiffness and platelet count and function: A systematic review and dose-response Meta-analysis of randomized clinical trials. Crit. Rev. Food Sci. Nutr. 2021, 61, 357–379. [Google Scholar] [CrossRef]
- Tanghe, A.; Heyman, E.; Lespagnol, E.; Stautemas, J.; Celie, B.; Op ‘t Roodt, J.; Rietzschel, E.; Dias Soares, D.; Hermans, N.; Tuenter, E.; et al. Acute effects of cocoa flavanols on blood pressure and peripheral vascular reactivity in type 2 diabetes mellitus and essential hypertension. Nutrients 2022, 14, 2692. [Google Scholar] [CrossRef]
- Alañon, M.E.; Castle, S.M.; Serra, G.; Leveques, A.; Poquet, L.; Actis-Goretta, L.; Spencer, J.P.E. Acute study of dose-dependent effects of (−)-epicatechin on vascular function in healthy male volunteers: A randomized controlled trial. Clin. Nutr. 2020, 39, 746–754. [Google Scholar] [CrossRef]
- Gröne, M.; Sansone, R.; Höffken, P.; Horn, P.; Rodriguez-Mateos, A.; Schroeter, H.; Kelm, M.; Heiss, C. Cocoa flavanols improve endothelial functional integrity in healthy young and elderly subjects. J. Agric. Food Chem. 2020, 68, 1871–1876. [Google Scholar] [CrossRef] [PubMed]
- Ottaviani, J.I.; Britten, A.; Lucarelli, D.; Luben, R.; Mulligan, A.A.; Lentjes, M.A.; Fong, R.; Gray, N.; Grace, P.B.; Mawson, D.H.; et al. Biomarker-estimated flavan-3-ol intake is associated with lower blood pressure in cross-sectional analysis in EPIC Norfolk. Sci. Rep. 2020, 10, 17964. [Google Scholar] [CrossRef] [PubMed]
- Milenkovic, D.; Rodriguez-Mateos, A.; Lucosz, M.; Istas, G.; Declerck, K.; Sansone, R.; Deenen, R.; Köhrer, K.; Corral-Jara, K.F.; Altschmied, J.; et al. Flavanol consumption in healthy men preserves integrity of immunological-endothelial barrier cell functions: Nutri(epi)genomic analysis. Mol. Nutr. Food Res. 2022, 66, 2100991. [Google Scholar] [CrossRef] [PubMed]
- Sesso, H.D.; Manson, J.E.; Aragaki, A.K.; Rist, P.M.; Johnson, L.G.; Friedenberg, G.; Copeland, T.; Clar, A.; Mora, S.; Vinayaga Moorthy, M.; et al. Effect of cocoa flavanol supplementation for the prevention of cardiovascular disease events: The COcoa Supplement and Multivitamin Outcomes Study (COSMOS) randomized clinical trial. Am. J. Clin. Nutr. 2022, 115, 1490–1500. [Google Scholar] [CrossRef] [PubMed]
- Jafarnejad, S.; Salek, M.; Clark, C.C.T. Cocoa consumption and blood pressure in middle-aged and elderly subjects: A meta-analysis. Curr. Hypertens. Rep. 2020, 22, 1. [Google Scholar] [CrossRef]
- Rashid, M.; Verhoeven, A.J.M.; Mulder, M.T.; Timman, R.; Ozcan, B.; van Beek-Nieuwland, Y.; Chow, L.M.; van de Laar, R.J.J.M.; Dik, W.A.; Sijbrands, E.J.G.; et al. The effect of monomeric and oligomeric FLAVAnols in patients with type 2 diabetes and microalbuminuria (FLAVA-trial): A double blind randomized controlled trial. Clin. Nutr. 2021, 40, 5587–5594. [Google Scholar] [CrossRef]
- Cadoná, F.C.; Ferreira Dantas, R.; Haas de Mello, G.; Paes Silva, F., Jr. Natural products targeting into cancer hallmarks: An update on caffeine, theobromine, and (+) catechin. Crit. Rev. Food Sci. Nutr. 2022, 62, 7222–7241. [Google Scholar] [CrossRef]
- Monteiro, J.; Alves, M.G.; Oliveira, P.F.; Silva, B.M. Pharmacological potential of methylxanthines: Retrospective analysis and future expectations. Crit. Rev. Food Sci. Nutr. 2019, 59, 2597–2625. [Google Scholar] [CrossRef] [Green Version]
- Camps-Bossacoma, M.; Garcia-Aloy, M.; Saldaña-Ruiz, S.; Cambras, T.; Gonzalez-Domínguez, R.; Franch, A.; Perez-Cano, F.J.; Andres-Lacueva, C.; Castell, M. Role of theobromine in cocoa’s metabolic properties in healthy rats. J. Agric. Food Chem. 2019, 67, 3605–3614. [Google Scholar] [CrossRef]
- Leehey, D.J. Targeting inflammation in diabetic kidney disease: Is there a role for pentoxifylline? Kidney 360 2020, 1, 292–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aronsen, L.; Orvoll, E.; Lysaa, R.; Ravna, A.W.; Sager, G. Modulation of high affinity ATP-dependent cyclic nucleotide transporters by specific and non-specific cyclic nucleotide phosphodiesterase inhibitors. Eur. J. Pharmacol. 2014, 745, 249–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gil, M.; Skopinska-Rozewska, E.; Radomska, D.; Demkow, U.; Skurzak, H.; Rochowska, M.; Beuth, J.; Roszkowski, K. Effect of purinergic receptor antagonists suramin and theobromine on tumor-induced angiogenesis in BALB/c mice. Folia Biol. 1993, 39, 63–68. [Google Scholar]
- Barcz, E.; Sommer, E.; Sokolnicka, I.; Gawrychowski, K.; Roszkowska-Purska, K.; Janik, P.; Skopinska-Rozewska, E. The influence of theobromine on angiogenic activity and proangiogenic cytokines production of human ovarian cancer cells. Oncol. Rep. 1998, 5, 517–537. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, N.; Miwa, S.; Hitomi, Y.; Nakamura, H.; Tsuchiya, H.; Yachie, A. Theobromine, the primary methylxanthine found in theobroma cacao, prevents malignant glioblastoma proliferation by negatively regulating phosphodiesterase-4, extracellular signal-regulated kinase, Akt/mammalian target of rapamycin kinase, and nuclear factor-κB. Nutr. Cancer 2014, 66, 419–423. [Google Scholar] [CrossRef] [PubMed]
- Jang, Y.J.; Koo, H.J.; Sohn, E.H.; Kang, S.C.; Rhee, D.K.; Pyo, S. Theobromine inhibits differentiation of 3T3-L1 cells during the early stage of adipogenesis via AMPK and MAPK signaling pathways. Food Funct. 2015, 6, 2365–2374. [Google Scholar] [CrossRef]
- Papadimitriou, A.; Silva, K.C.; Peixoto, E.B.M.I.; Borges, C.M.; Lopes de Faria, J.M.; Lopes de Faria, J.B. Theobromine increases NAD+/Sirt-1 activity and protects the kidney under diabetic conditions. Am. J. Physiol. Renal Physiol. 2015, 308, F209–F225. [Google Scholar] [CrossRef]
- Cadonna, F.C.; Machado, A.K.; Azzolin, V.F.; Barbisan, F.; Dornelles, E.B.; Glanzner, W.; Goncalves, P.B.D.; Assmann, C.E.; Ribeiro, E.E.; da Cruz, I.B.M. Guarana a caffeine-rich food increases oxaliplatin sensitivity of colorectal HT-29 cells by apoptosis pathway modulation. Anti-Cancer Agents Med. Chem. 2016, 16, 1055–1065. [Google Scholar] [CrossRef]
- Yoneda, M.; Sugimoto, N.; Katakura, M.; Matsuzaki, K.; Tanigami, H.; Yachi, A.; Ohno-Shosaku, T.; Shido, O. Theobromine up-regulates cerebral brain-derived neurotrophic factor and facilitates motor learning in mice. J. Nutr. Biochem. 2017, 39, 110–116. [Google Scholar] [CrossRef] [Green Version]
- Mitani, T.; Watanabe, S.; Yoshioka, Y.; Katayama, S.; Nakamura, S.; Ashida, H. Theobromine suppresses adipogenesis through enhancement of CCAAT enhancer-binding protein β degradation by adenosine receptor A1. BBA Mol. Cell Res. 2017, 1864, 2438–2448. [Google Scholar] [CrossRef]
- Shojaei-Zarghani, S.; Khosroushahi, A.Y.; Rafraf, M. Oncopreventive effects of theanine and theobromine on dimethylhydrazine-induced colon cancer model. Biomed. Pharmacother. 2021, 134, 111140. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, E.; Mitani, T.; Nakashima, M.; Yonemoto, E.; Fujii, H.; Ashida, H. Theobromine enhances the conversion of white adipocytes into beige adipocytes in a PPAR γactivation-dependent manner. J. Nutr. Biochem. 2022, 100, 108898. [Google Scholar] [CrossRef]
- Wei, D.; Wu, S.; Liu, J.; Zhang, X.; Guan, X.; Gao, L.; Xu, Z. Theobromine ameliorates nonalcoholic fatty liver disease by regulating hepatic lipid metabolism via mTOR signaling pathway in vivo and in vitro. Can. J. Physiol. Pharmacol. 2021, 99, 775–785. [Google Scholar] [CrossRef] [PubMed]
- Janitschke, D.; Lauer, A.A.; Bachmann, C.M.; Winkler, J.; Griebsch, L.V.; Pilz, S.M.; Theiss, E.L.; Grimm, H.S.; Hartmann, T.; Grimm, M.O.W. Methylxanthines induce a change in the AD/neurodegeneration- linked lipid profile in neuroblastoma cells. Int. J. Mol. Sci. 2022, 23, 2295. [Google Scholar] [CrossRef] [PubMed]
- Rebollo-Hernanz, M.; Aguilera, Y.; Martin-Cabrejas, M.A.; Gonzalez de Mejia, E. Phytochemicals from the cocoa shell modulate mitochondrial function, lipid and glucose metabolism in hepatocytes via activation of FGF21/ERK, AKT, and mTOR pathways. Antioxidants 2022, 11, 136. [Google Scholar] [CrossRef]
- Fernández-Fernández, L.; Esteban, G.; Giralt, M.; Valente, T.; Bolea, I.; Solé, M.; Sun, P.; Benítez, S.; Morelló, J.R.; Reguant, J.; et al. Catecholaminergic and cholinergic systems of mouse brain are modulated by LMN diet, rich in theobromine polyphenols and polyunsaturated fatty acids. Food Funct. 2015, 6, 1251–1260. [Google Scholar] [CrossRef]
- Gu, R.; Shi, Y.; Huang, W.; Lao, C.; Zou, Z.; Pan, S.; Huang, Z. Theobromine mitigates IL-1β-induced oxidative stress, inflammatory response, and degradation of type II collagen in human chondrocytes. Int. Immunopharmacol. 2020, 82, 106226. [Google Scholar] [CrossRef]
- Sarriá, B.; Gomez-Juaristi, M.; Martínez López, S.; García Cordero, J.; Bravo, L.; Mateos Briz, M.R. Cocoa colonic phenolic metabolites are related to HDL-cholesterol raising effects and methylxanthine metabolites and insoluble dietary fibre to anti-inflammatory and hypoglycemic effects in humans. PeerJ 2020, 8, e9953. [Google Scholar] [CrossRef]
- Rodríguez-Rodríguez, P.; Ragusky, K.; Phuthong, S.; Ruvira, S.; Ramiro-Cortijo, D.; Cañas, S.; Rebollo-Hernanz, M.; Morales, M.D.; López de Pablo, A.L.; Martín-Cabrejas, M.A.; et al. Vasoactive properties of a cocoa shell extract: Mechanism of action and effect on endothelial dysfunction in aged rats. Antioxidants 2022, 11, 429. [Google Scholar] [CrossRef]
- Talbot, C.P.J.; Mensink, R.P.; Smolders, L.; Bakeroot, V.; Plat, J. Theobromine does not affect fasting and postprandial HDL cholesterol efflux capacity, while it decreases fasting miR-92a levels in humans. Mol. Nutr. Food Res. 2018, 62, e1800027. [Google Scholar] [CrossRef]
- Smolders, L.; Mensink, R.P.; van den Driessche, J.J.; Joris, P.J.; Plat, J. Theobromine consumption does not improve fasting and postprandial vascular function in overweight and obese subjects. Eur. J. Nutr. 2019, 58, 981–987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cova, I.; Leta, V.; Mariani, C.; Pantoni, L.; Pomati, S. Exploring cocoa properties: Is theobromine a cognitive modulator? Psychopharmacology 2019, 236, 561–572. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Daly, J.W. Chronic effects of xanthines on levels of central receptors in mice. Cell. Mol. Neurobiol. 1999, 19, 719–732. [Google Scholar] [CrossRef] [PubMed]
- He, R.; Xie, G.; Yao, X.-S.; Kurihara, H. Effect of cocoa tea (Camellia ptilophylla) co-administrated with green tea on ambulatory behaviors. Biosci. Biotechnol. Biochem. 2009, 73, 957–960. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, E.S.; Slettenaar, M.; vd Meer, N.; Transler, C.; Jans, L.; Quadt, F.; Berry, M. Differential contributions of theobromine and caffeine on mood, psychomotor performance and blood pressure. Physiol. Behav. 2011, 104, 816–822. [Google Scholar] [CrossRef]
- Travassos, M.; Santana, I.; Baldeiras, I.; Tsolaki, M.; Gkatzima, O.; Sermin, G.; Yener, G.G.; Simonsen, A.; Hasselbalch, S.G.F.; Kapaki, E.; et al. Does caffeine consumption modify cerebrospinal fluid amyloid-β levels in patients with Alzheimer’s disease? J. Alzheimer Dis. 2015, 47, 1069–1078. [Google Scholar] [CrossRef]
- Mendiola-Precoma, J.; Padilla, K.; Rodríguez-Cruz, A.; Berumen, L.C.; Miledi, R.; García-Alcocer, G. Theobromine-induced changes in A1 purinergic receptor gene expression and distribution in a rat brain Alzheimer’s disease model. J. Alzheimer Dis. 2017, 55, 1273–1283. [Google Scholar] [CrossRef]
- Martın-Pelaez, S.; Camps-Bossacoma, M.; Massot-Cladera, M.; Rigo-Adrover, M.; Franch, A.; Perez-Cano, F.J.; Castell, M. Effect of cocoa’s theobromine on intestinal microbiota of rats. Mol. Nutr. Food Res. 2017, 61, 1700238. [Google Scholar] [CrossRef]
- Rolta, R.; Salaria, D.; Sharma, B.; Awofisayo, O.; Fadare, O.A.; Sharma, S.; Patel, C.N.; Kumar, V.; Sourirajan, A.; Baumler, D.J.; et al. Methylxanthines as potential inhibitor of SARS-CoV-2: An In Silico Approach. Curr. Pharmacol. Rep. 2022, 8, 149–170. [Google Scholar] [CrossRef]


{kind=link}
| Stage | Process | Changes Occurring | Antioxidant Capacity | References |
|---|---|---|---|---|
| Primary processing | Harvesting | Increased proanthocyanidins, CF, and TBR with increasing pod maturation and ripening at harvest. | Increase in the antioxidant capacity with an increase in pod maturation and ripening. | [30,32] |
| Pod storage | Reduction in TP, EC, C, TBR, and CF with increasing duration of pod storage. | Decrease in the antioxidant capacity with increasing duration of pod storage during fermentation. | [18,34,35,36] | |
| Fermentation | Decrease in the concentrations of EC, C, anthocyanins, phenolic acids, and TP with increasing duration of fermentation. Moreover, a decrease in the TBR and CF concentrations. | Decrease in the antioxidant capacity of cocoa beans with increasing duration of fermentation. | [18,22,23,37,38,39,40,41,42] | |
| Drying | Degradation of TP, EC, C, and anthocyanins. Decrease in the alkaloids (TBR and CF) content. | Reduction in the antioxidant capacity during the drying of cocoa beans. | [39,40,42,43,44,45] | |
| Industrial processing | Roasting | Reduction in the amounts of TP, EC, procyanidin B2 and C1, anthocyanins, quercetin glycosides, TBR, and CF. An increase in C due to epimerization of EC. | A general reduction in the antioxidant capacity during cocoa bean roasting. | [22,23,30,40,41,42,45,46,47,48,49,50,51,52] |
| Alkalization | Reduction in TP, EC, C, procyanidin B2 and C1, quercetin, quercetin-3-glucuronide, quercetin-3-arabinoside, isoquercetin, TBR, and CF. | Antioxidant activity is reduced. | [20,21,40,42,47,53,54] | |
| Conching | Decrease in TP during conching. No significant change in the polyphenol content during conching. | Decrease in the antioxidant capacity. | [52,55,56,57,58,59,60] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goya, L.; Kongor, J.E.; de Pascual-Teresa, S. From Cocoa to Chocolate: Effect of Processing on Flavanols and Methylxanthines and Their Mechanisms of Action. Int. J. Mol. Sci. 2022, 23, 14365. https://doi.org/10.3390/ijms232214365
Goya L, Kongor JE, de Pascual-Teresa S. From Cocoa to Chocolate: Effect of Processing on Flavanols and Methylxanthines and Their Mechanisms of Action. International Journal of Molecular Sciences. 2022; 23(22):14365. https://doi.org/10.3390/ijms232214365
Chicago/Turabian StyleGoya, Luis, John Edem Kongor, and Sonia de Pascual-Teresa. 2022. "From Cocoa to Chocolate: Effect of Processing on Flavanols and Methylxanthines and Their Mechanisms of Action" International Journal of Molecular Sciences 23, no. 22: 14365. https://doi.org/10.3390/ijms232214365