

The Mucus Binding Factor Is Not Necessary for Lacticaseibacillus rhamnosus CRL1505 to Exert Its Immunomodulatory Activities in Local and Distal Mucosal Sites
 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Results
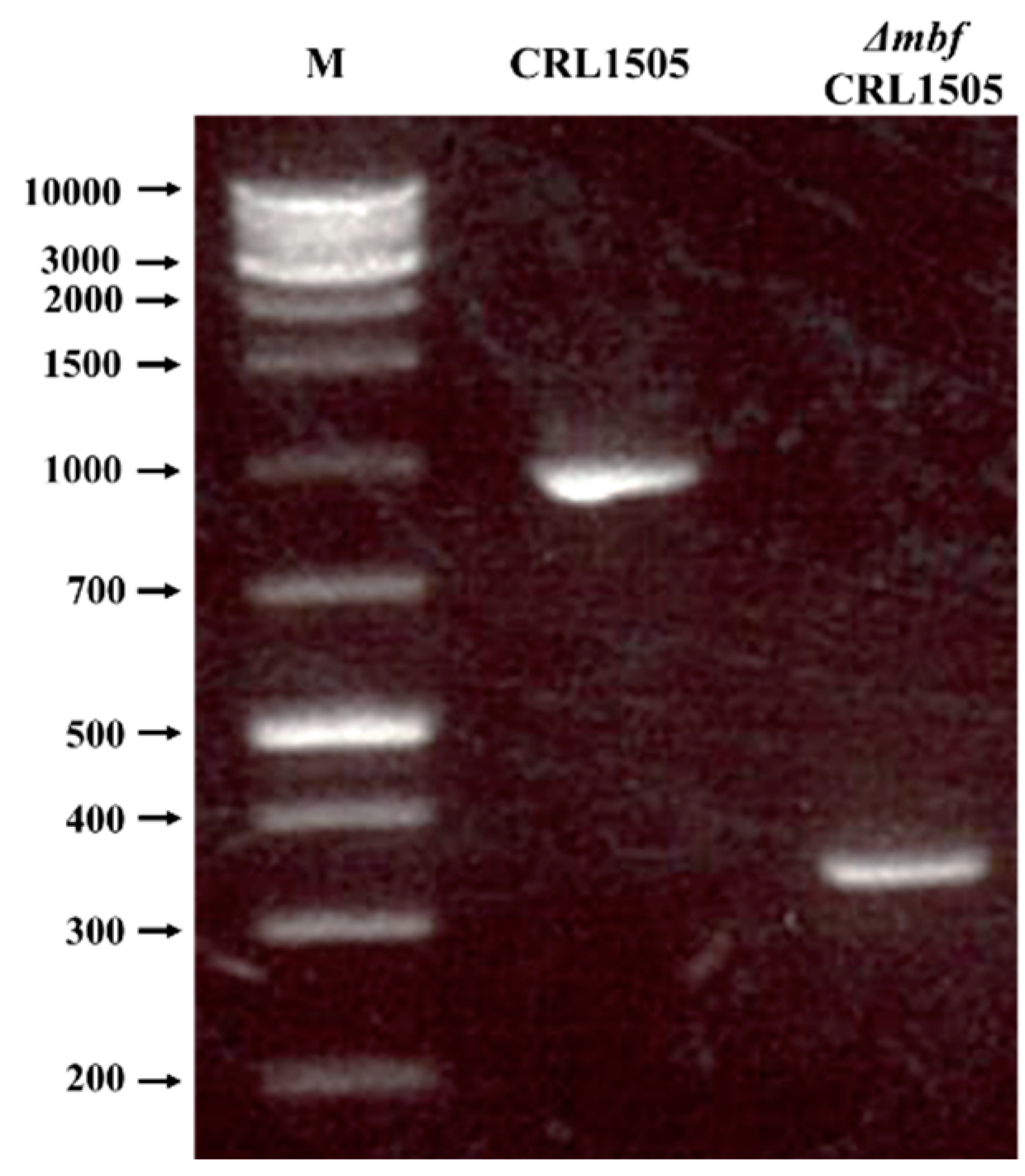
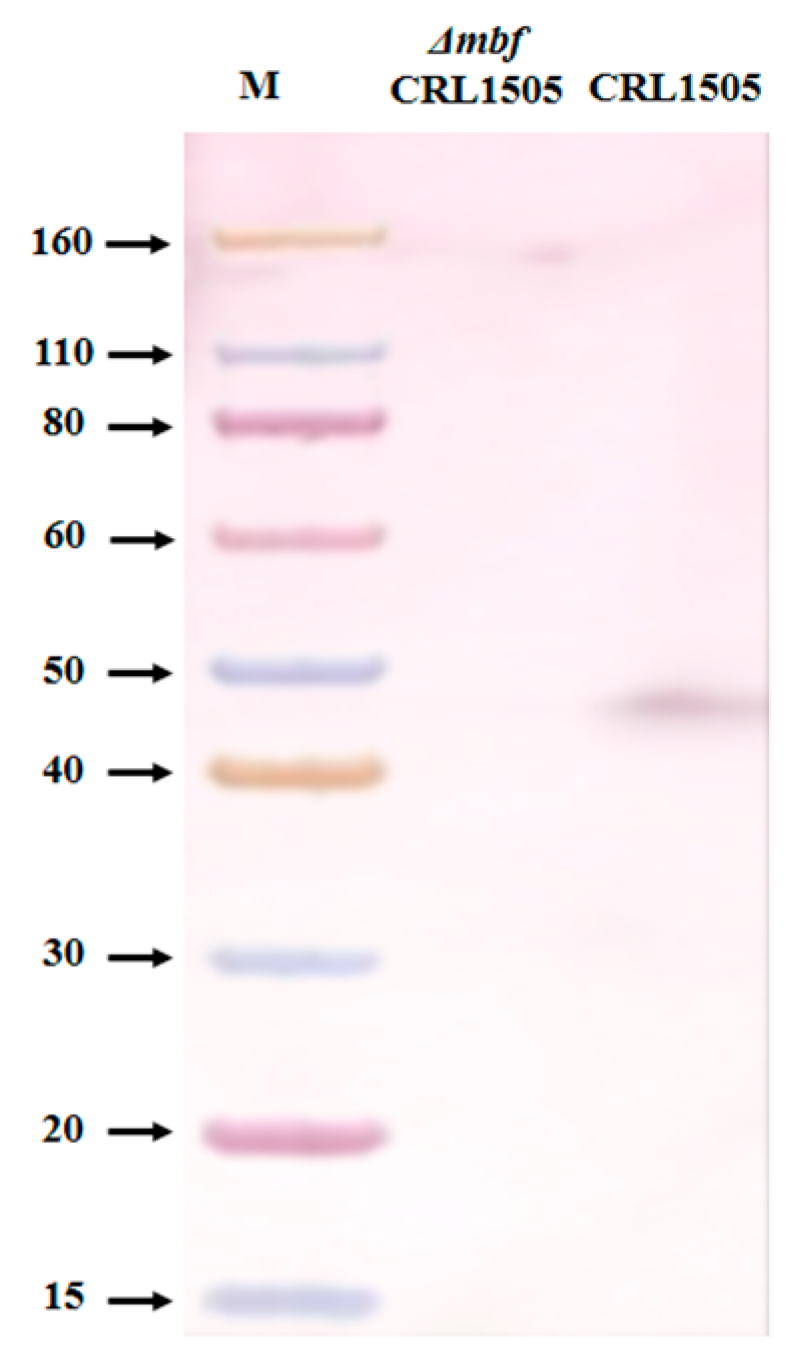
2.1. Construction of L. rhamnosus ΔmbfCRL1505
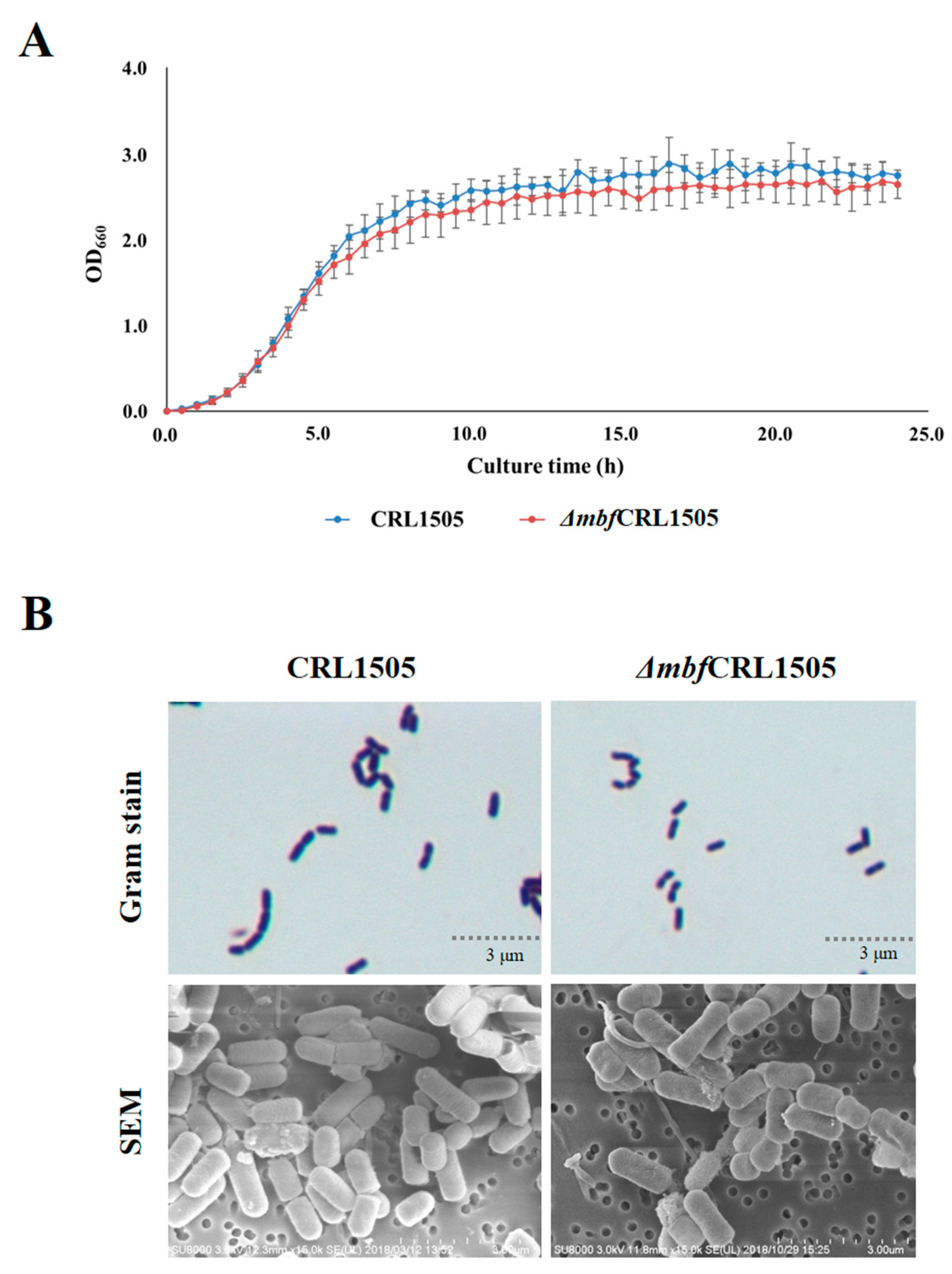
2.2. L. rhamnosus ΔmbfCRL1505 Phenotypic Evaluation
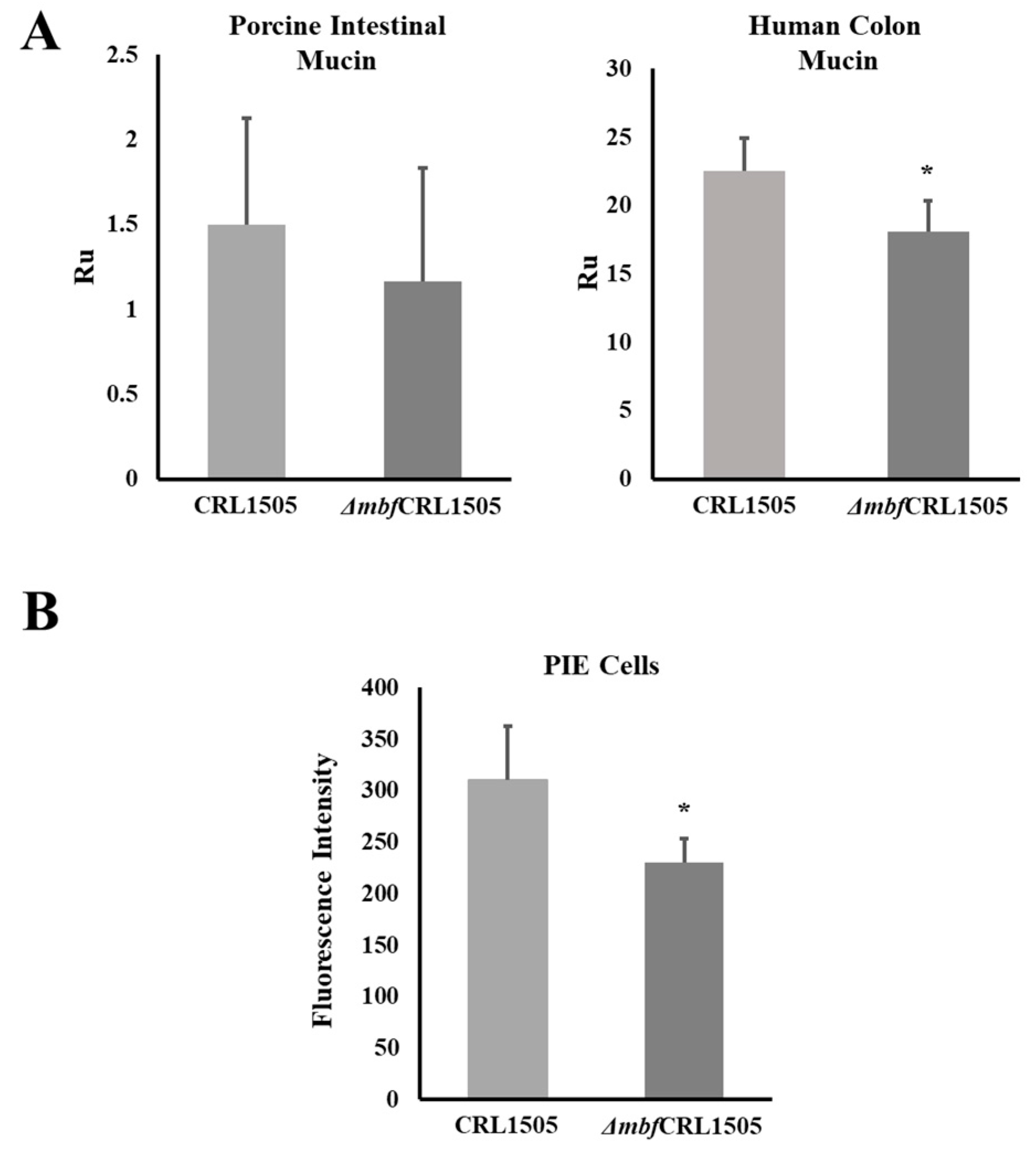
2.3. L. rhamnosus ΔmbfCRL1505 Adhesion to Mucins and PIE Cells
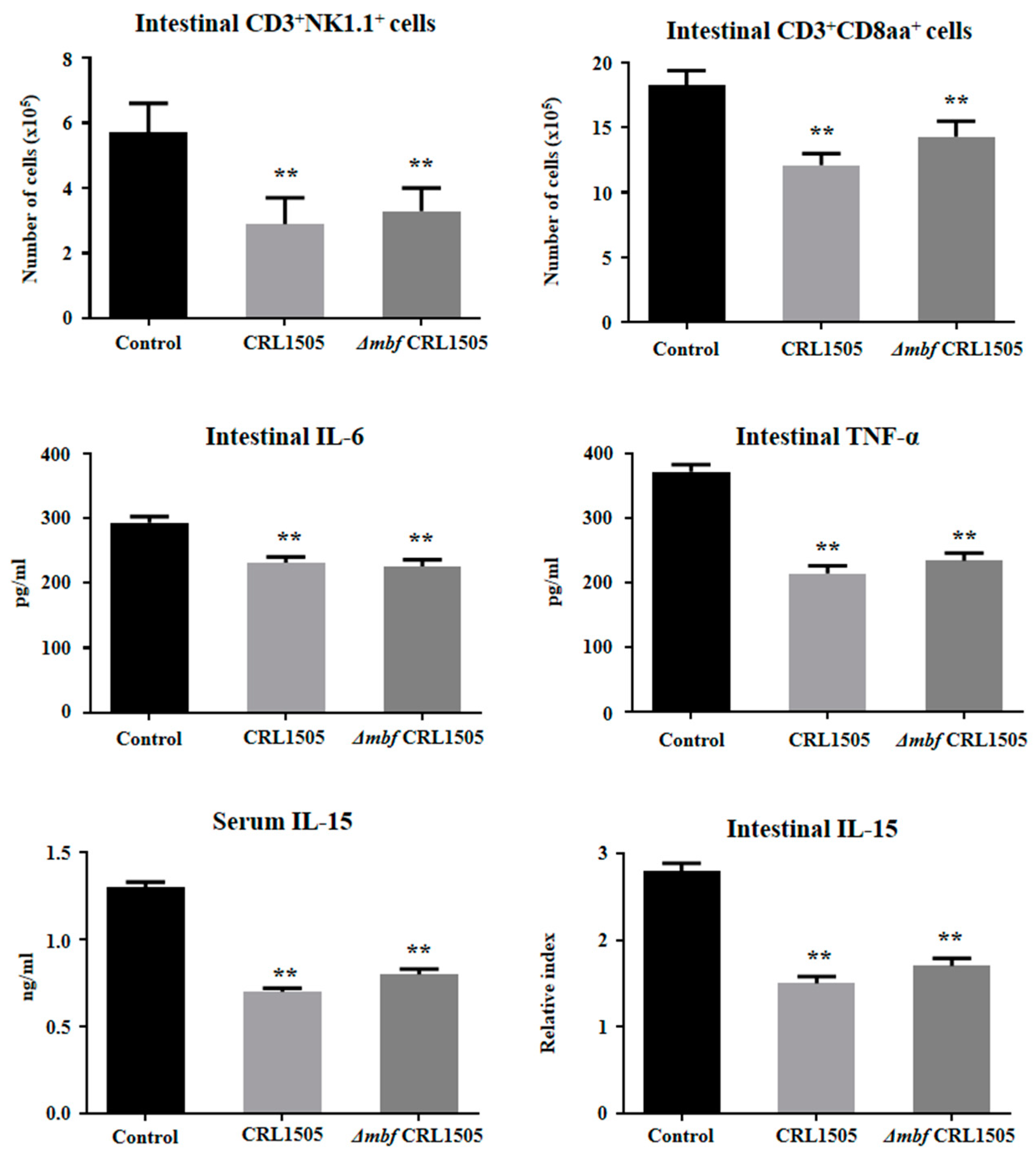
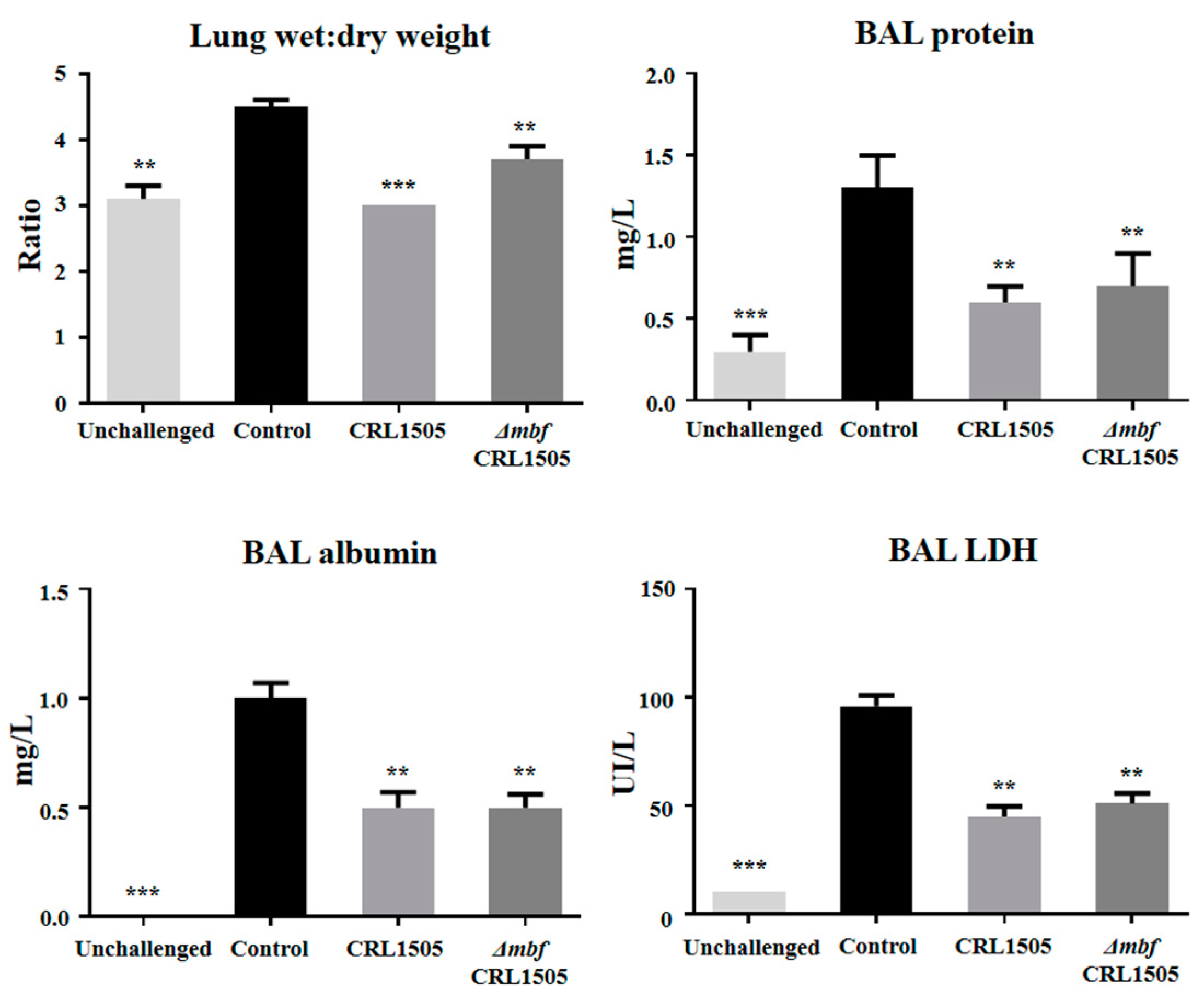
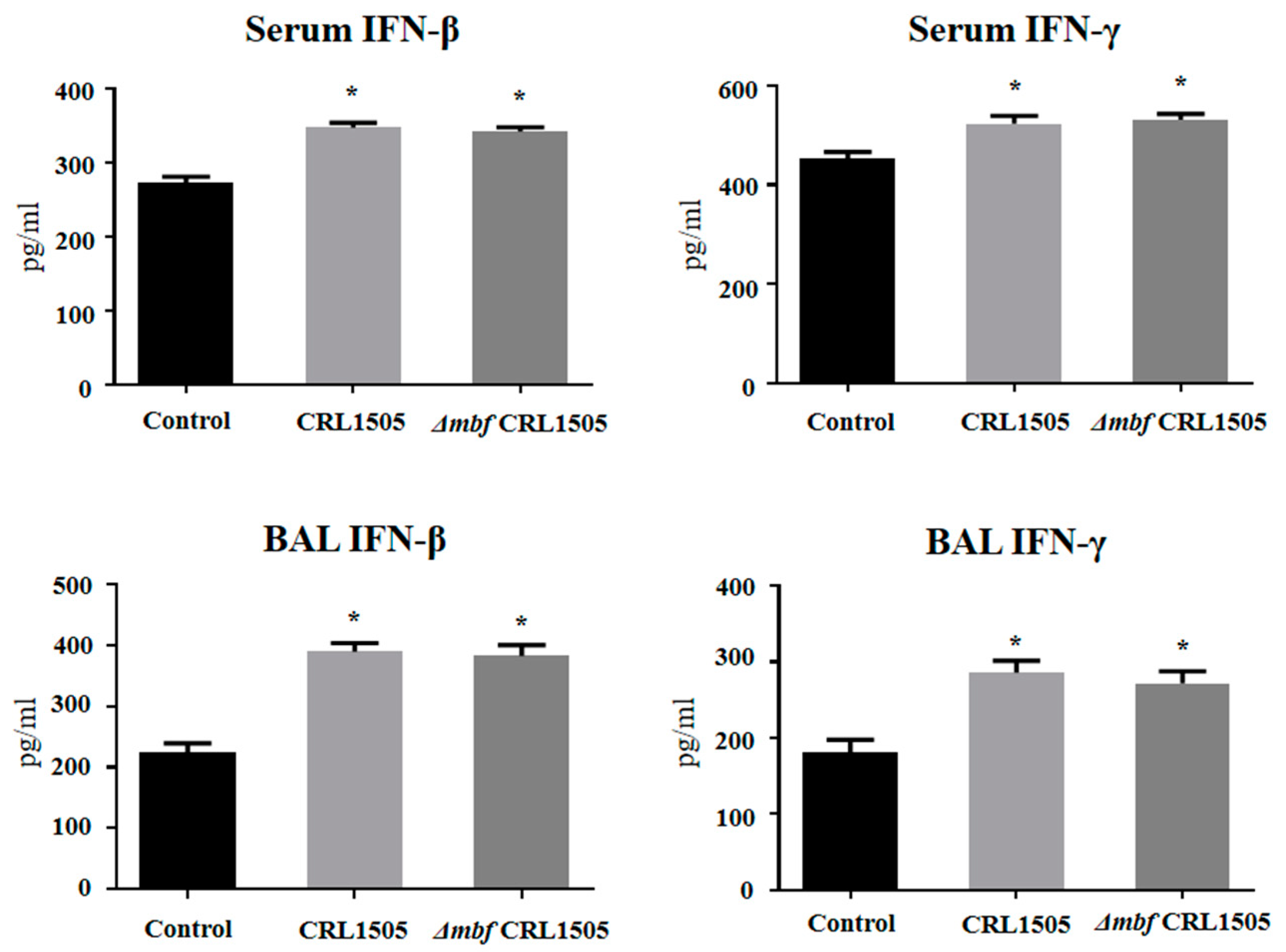
2.4. Immunomodulatory Activity of L. rhamnosus ΔmbfCRL1505 In Vivo
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Growth Conditions
4.2. Construction of mbf Knockout Gene by PCR
4.3. Recombinant Cloning of the mbf Knockout Gene and Construction of the Knockout Strain
4.4. Protein Detection by Western Blotting
4.5. Effect of the Knockout mbf Gene on the Viability and Phenotype of L. rhamnosus CRL1505
4.6. Binding Ability Assays
4.7. Animals, Feeding Procedures and Administration of Poly(I:C)
4.8. Serum, Bronchoalveolar Lavage (BAL) and Intestinal Fluid Sampling
4.9. Cytokines and Chemokines Analysis
4.10. Determination of Blood Cell Populations
4.11. Biochemical Assay of BAL Samples
4.12. Lung Wet:Dry Weight Ratio
4.13. Study of Intestinal Intraepithelial Lymphocytes
4.14. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Capurso, L. Thirty years of Lactobacillus rhamnosus GG: A Review. J. Clin. Gastroenterol. 2019, 53 (Suppl. S1), S1–S41. [Google Scholar] [CrossRef] [PubMed]
- Salminen, S.; Nybom, S.; Meriluoto, J.; Collado, M.C.; Vesterlund, S.; El-Nezami, H. Interaction of probiotics and pathogens—Benefits to human health? Curr. Opin. Biotechnol. 2010, 21, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Dicks, L.M.; Botes, M. Probiotic lactic acid bacteria in the gastro-intestinal tract: Health benefits, safety and mode of action. Benef. Microbes 2010, 1, 11–29. [Google Scholar] [CrossRef] [PubMed]
- Von Ossowski, I.; Satokari, R.; Reunanen, J.; Lebeer, S.; De Keersmaecker, S.C.; Vanderleyden, J.; de Vos, W.M.; Palva, A. Functional characterization of a mucus-specific LPXTG surface adhesin from probiotic Lactobacillus rhamnosus GG. Appl. Environ. Microbiol. 2011, 77, 4465–4472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, F.; Liu, L.; Cao, H.; Moore, D.J.; Washington, M.K.; Wang, B.; Peek, R.M.; Acra, S.A.; Polk, D.B. Neonatal colonization of mice with LGG promotes intestinal development and decreases susceptibility to colitis in adulthood. Mucosal. Immunol. 2017, 10, 117–127. [Google Scholar] [CrossRef] [Green Version]
- Velez, M.P.; Petrova, M.I.; Lebeer, S.; Verhoeven, T.L.; Claes, I.; Lambrichts, I.; Tynkkynen, S.; Vanderleyden, J.; De Keersmaecker, S.C. Characterization of MabA, a modulator of Lactobacillus rhamnosus GG adhesion and biofilm formation. FEMS Immunol. Med. Microbiol. 2010, 59, 386–398. [Google Scholar] [CrossRef] [Green Version]
- He, F.; Morita, H.; Kubota, A.; Ouwehand, A.C.; Hosoda, M.; Hiramatsu, M.; Kurisaki, J. Effect of orally administered non-viable Lactobacillus cells on murine humoral immune responses. Microbiol. Immunol. 2005, 49, 993–997. [Google Scholar] [CrossRef]
- Racedo, S.; Villena, J.; Medina, M.; Aguero, G.; Rodriguez, V.; Alvarez, S. Lactobacillus casei administration reduces lung injuries in a Streptococcus pneumoniae infection in mice. Microbes Infect. 2006, 8, 2359–2366. [Google Scholar] [CrossRef]
- Villena, J.; Oliveira, M.L.; Ferreira, P.C.; Salva, S.; Alvarez, S. Lactic acid bacteria in the prevention of pneumococcal respiratory infection: Future opportunities and challenges. Int. Immunopharmacol. 2011, 11, 1633–1645. [Google Scholar] [CrossRef]
- Villena, J.; Chiba, E.; Tomosada, Y.; Salva, S.; Marranzino, G.; Kitazawa, H.; Alvarez, S. Orally administered Lactobacillus rhamnosus modulates the respiratory immune response triggered by the viral pathogen-associated molecular pattern poly(I:C). BMC Immunol. 2012, 13, 53. [Google Scholar] [CrossRef]
- Villena, J.; Barbieri, N.; Salva, S.; Herrera, M.; Alvarez, S. Enhanced immune response to pneumococcal infection in malnourished mice nasally treated with heat-killed Lactobacillus casei. Microbiol. Immunol. 2009, 53, 636–646. [Google Scholar] [CrossRef] [PubMed]
- Raya Tonetti, F.; Arce, L.; Salva, S.; Alvarez, S.; Takahashi, H.; Kitazawa, H.; Vizoso-Pinto, M.G.; Villena, J. Immunomodulatory properties of bacterium-like particles obtained from immunobiotic lactobacilli: Prospects for their use as mucosal adjuvants. Front. Immunol. 2020, 11, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishiyama, K.; Nakamata, K.; Ueno, S.; Terao, A.; Aryantini, N.P.; Sujaya, I.N.; Fukuda, K.; Urashima, T.; Yamamoto, Y.; Mukai, T. Adhesion properties of Lactobacillus rhamnosus mucus-binding factor to mucin and extracellular matrix proteins. Biosci. Biotechnol. Biochem. 2015, 79, 271–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tada, A.; Zelaya, H.; Clua, P.; Salva, S.; Alvarez, S.; Kitazawa, H.; Villena, J. Immunobiotic Lactobacillus strains reduce small intestinal injury induced by intraepithelial lymphocytes after Toll-like receptor 3 activation. Inflamm. Res. 2016, 65, 771–783. [Google Scholar] [CrossRef]
- Tonetti, F.R.; Islam, M.A.; Vizoso-Pinto, M.G.; Takahashi, H.; Kitazawa, H.; Villena, J. Nasal priming with immunobiotic lactobacilli improves the adaptive immune response against influenza virus. Int. Immunopharmacol. 2020, 78, 106115. [Google Scholar] [CrossRef]
- Zelaya, H.; Alvarez, S.; Kitazawa, H.; Villena, J. Respiratory antiviral immunity and immunobiotics: Beneficial effects on inflammation-coagulation interaction during influenza virus infection. Front. Immunol. 2016, 7, 633. [Google Scholar] [CrossRef] [Green Version]
- Villena, J.; Kitazawa, H. Editorial: Immunobiotics-interactions of beneficial microbes with the immune system. Front. Immunol. 2017, 8, 1580. [Google Scholar] [CrossRef] [Green Version]
- Villena, J.; Li, C.; Vizoso-Pinto, M.G.; Sacur, J.; Ren, L.; Kitazawa, H. Lactiplantibacillus plantarum as a potential adjuvant and delivery system for the development of SARS-CoV-2 oral vaccines. Microorganisms 2021, 9, 683. [Google Scholar] [CrossRef]
- Villena, J.; Kitazawa, H. The modulation of mucosal antiviral immunity by immunobiotics: Could they offer any benefit in the SARS-CoV-2 pandemic? Front. Physiol. 2020, 11, 699. [Google Scholar] [CrossRef]
- Garcia-Castillo, V.; Albarracin, L.; Kitazawa, H.; Villena, J. Screening and characterization of immunobiotic lactic acid bacteria with porcine immunoassay systems. Methods Mol. Biol. 2019, 1887, 131–144. [Google Scholar] [CrossRef]
- Barberi, C.; Campana, S.; De Pasquale, C.; Rabbani Khorasgani, M.; Ferlazzo, G.; Bonaccorsi, I. T cell polarizing properties of probiotic bacteria. Immunol. Lett. 2015, 168, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Monteagudo-Mera, A.; Rastall, R.A.; Gibson, G.R.; Charalampopoulos, D.; Chatzifragkou, A. Adhesion mechanisms mediated by probiotics and prebiotics and their potential impact on human health. Appl. Microbiol. Biotechnol. 2019, 103, 6463–6472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bierne, H.; Cossart, P. Listeria monocytogenes surface proteins: From genome predictions to function. Microbiol. Mol. Biol. Rev. 2007, 71, 377–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desvaux, M.; Dumas, E.; Chafsey, I.; Hebraud, M. Protein cell surface display in Gram-positive bacteria: From single protein to macromolecular protein structure. FEMS Microbiol. Lett. 2006, 256, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Villena, J.; Vizoso-Pinto, M.G.; Kitazawa, H. Intestinal innate antiviral immunity and immunobiotics: Beneficial effects against rotavirus infection. Front. Immunol. 2016, 7, 563. [Google Scholar] [CrossRef] [Green Version]
- Albarracin, L.; Kobayashi, H.; Iida, H.; Sato, N.; Nochi, T.; Aso, H.; Salva, S.; Alvarez, S.; Kitazawa, H.; Villena, J. Transcriptomic analysis of the innate antiviral immune response in porcine intestinal epithelial cells: Influence of immunobiotic lactobacilli. Front. Immunol. 2017, 8, 57. [Google Scholar] [CrossRef] [Green Version]
- Villena, J.; Chiba, E.; Vizoso-Pinto, M.G.; Tomosada, Y.; Takahashi, T.; Ishizuka, T.; Aso, H.; Salva, S.; Alvarez, S.; Kitazawa, H. Immunobiotic Lactobacillus rhamnosus strains differentially modulate antiviral immune response in porcine intestinal epithelial and antigen presenting cells. BMC Microbiol. 2014, 14, 126. [Google Scholar] [CrossRef] [Green Version]
- Hosoya, S.; Villena, J.; Shimazu, T.; Tohno, M.; Fujie, H.; Chiba, E.; Shimosato, T.; Aso, H.; Suda, Y.; Kawai, Y.; et al. Immunobiotic lactic acid bacteria beneficially regulate immune response triggered by poly(I:C) in porcine intestinal epithelial cells. Vet. Res. 2011, 42, 111. [Google Scholar] [CrossRef] [Green Version]
- Zhou, R.; Wei, H.; Sun, R.; Tian, Z. Recognition of double-stranded RNA by TLR3 induces severe small intestinal injury in mice. J. Immunol. 2007, 178, 4548–4556. [Google Scholar] [CrossRef] [Green Version]
- Chiba, E.; Tomosada, Y.; Vizoso-Pinto, M.G.; Salva, S.; Takahashi, T.; Tsukida, K.; Kitazawa, H.; Alvarez, S.; Villena, J. Immunobiotic Lactobacillus rhamnosus improves resistance of infant mice against respiratory syncytial virus infection. Int. Immunopharmacol. 2013, 17, 373–382. [Google Scholar] [CrossRef]
- Zelaya, H.; Tsukida, K.; Chiba, E.; Marranzino, G.; Alvarez, S.; Kitazawa, H.; Aguero, G.; Villena, J. Immunobiotic lactobacilli reduce viral-associated pulmonary damage through the modulation of inflammation-coagulation interactions. Int. Immunopharmacol. 2014, 19, 161–173. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Castillo, V.; Tomokiyo, M.; Raya Tonetti, F.; Islam, M.A.; Takahashi, H.; Kitazawa, H.; Villena, J. Alveolar macrophages are key players in the modulation of the respiratory antiviral immunity induced by orally administered Lacticaseibacillus rhamnosus CRL1505. Front. Immunol. 2020, 11, 568636. [Google Scholar] [CrossRef] [PubMed]
- Lebeer, S.; Vanderleyden, J.; De Keersmaecker, S.C. Host interactions of probiotic bacterial surface molecules: Comparison with commensals and pathogens. Nat. Rev. Microbiol. 2010, 8, 171–184. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, K.; Ochiai, A.; Tsubokawa, D.; Ishihara, K.; Yamamoto, Y.; Mukai, T. Identification and characterization of sulfated carbohydrate-binding protein from Lactobacillus reuteri. PLoS ONE 2013, 8, e83703. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Albarracin, L.; Indo, Y.; Arce, L.; Masumizu, Y.; Tomokiyo, M.; Islam, M.A.; Garcia-Castillo, V.; Ikeda-Ohtsubo, W.; Nochi, T.; et al. Selection of immunobiotic Ligilactobacillus salivarius strains from the intestinal tract of wakame-fed pigs: Functional and genomic studies. Microorganisms 2020, 8, 1659. [Google Scholar] [CrossRef] [PubMed]
- Salva, S.; Villena, J.; Alvarez, S. Immunomodulatory activity of Lactobacillus rhamnosus strains isolated from goat milk: Impact on intestinal and respiratory infections. Int. J. Food Microbiol. 2010, 141, 82–89. [Google Scholar] [CrossRef]
- Villena, J.; Racedo, S.; Aguero, G.; Bru, E.; Medina, M.; Alvarez, S. Lactobacillus casei improves resistance to pneumococcal respiratory infection in malnourished mice. J. Nutr. 2005, 135, 1462–1469. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′—3′) | Restriction Site | Annealing Temperature |
|---|---|---|---|
| Lr13 | GTCAACCATCTAGAAGCATGAA | 55 °C | |
| Lr14 | TACTGCGATTCATCGCTAGG | 55 °C | |
| Lr3 | ACACGTCGTCGAACAGTTGTGGTGAAGTGTGTTGAT | SalI | 55 °C |
| Lr4 | CAACCATCACTGGCGGCCAGCCTCATCCAC | 55 °C | |
| Lr5 | GTGGATGAGGCTGGCCGCCAGTGATGGTTG | 55 °C | |
| Lr6 | ACACGAGCTCCCGCCAGTGATTAAACTGGT | SacI | 55 °C |
| p119 | CTTTTACGTTTCCGCCATTC | 50 °C | |
| p120 | ATTTCATCAATGGCCTCAGT | 50 °C | |
| Lr17 | GGTTCGTATACTGCCGTGCCA | 50 °C | |
| Lr18 | GCTTGTTGACCAGGCTGATATTCT | 50 °C |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tomotsune, K.; Raya Tonetti, F.; Mizuno, H.; Elean, M.; Fukuyama, K.; Zhou, B.; Ikeda-Ohtsubo, W.; Nishiyama, K.; Yamamura, A.; Karasawa, H.; et al. The Mucus Binding Factor Is Not Necessary for Lacticaseibacillus rhamnosus CRL1505 to Exert Its Immunomodulatory Activities in Local and Distal Mucosal Sites. Int. J. Mol. Sci. 2022, 23, 14357. https://doi.org/10.3390/ijms232214357
Tomotsune K, Raya Tonetti F, Mizuno H, Elean M, Fukuyama K, Zhou B, Ikeda-Ohtsubo W, Nishiyama K, Yamamura A, Karasawa H, et al. The Mucus Binding Factor Is Not Necessary for Lacticaseibacillus rhamnosus CRL1505 to Exert Its Immunomodulatory Activities in Local and Distal Mucosal Sites. International Journal of Molecular Sciences. 2022; 23(22):14357. https://doi.org/10.3390/ijms232214357
Chicago/Turabian StyleTomotsune, Kae, Fernanda Raya Tonetti, Hiroya Mizuno, Mariano Elean, Kohtaro Fukuyama, Binghui Zhou, Wakako Ikeda-Ohtsubo, Keita Nishiyama, Akihiro Yamamura, Hideaki Karasawa, and et al. 2022. "The Mucus Binding Factor Is Not Necessary for Lacticaseibacillus rhamnosus CRL1505 to Exert Its Immunomodulatory Activities in Local and Distal Mucosal Sites" International Journal of Molecular Sciences 23, no. 22: 14357. https://doi.org/10.3390/ijms232214357