
Biochemical Targets and Molecular Mechanism of Ginsenoside Compound K in Treating Osteoporosis Based on Network Pharmacology

Abstract
:1. Introduction
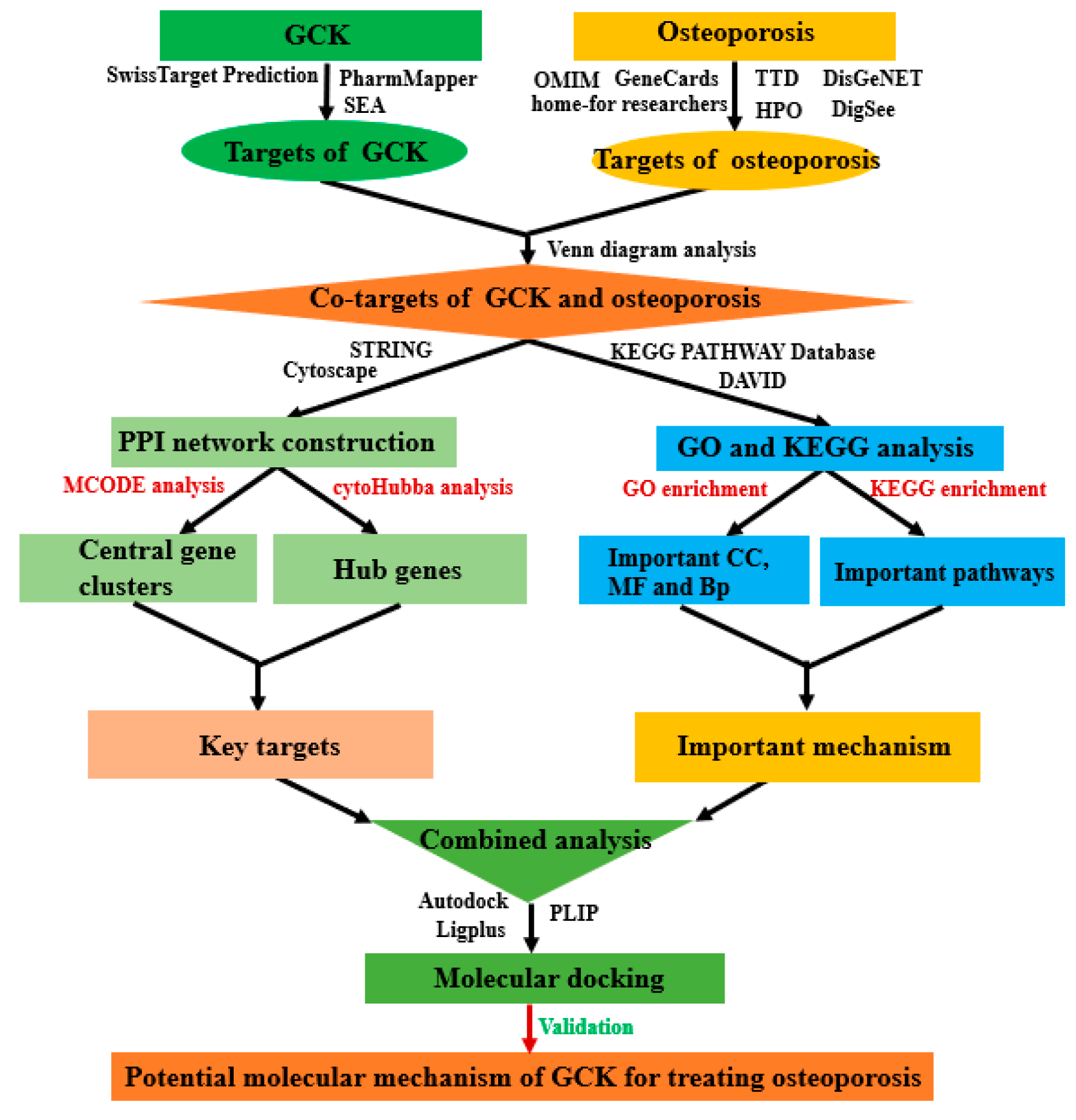
2. Results
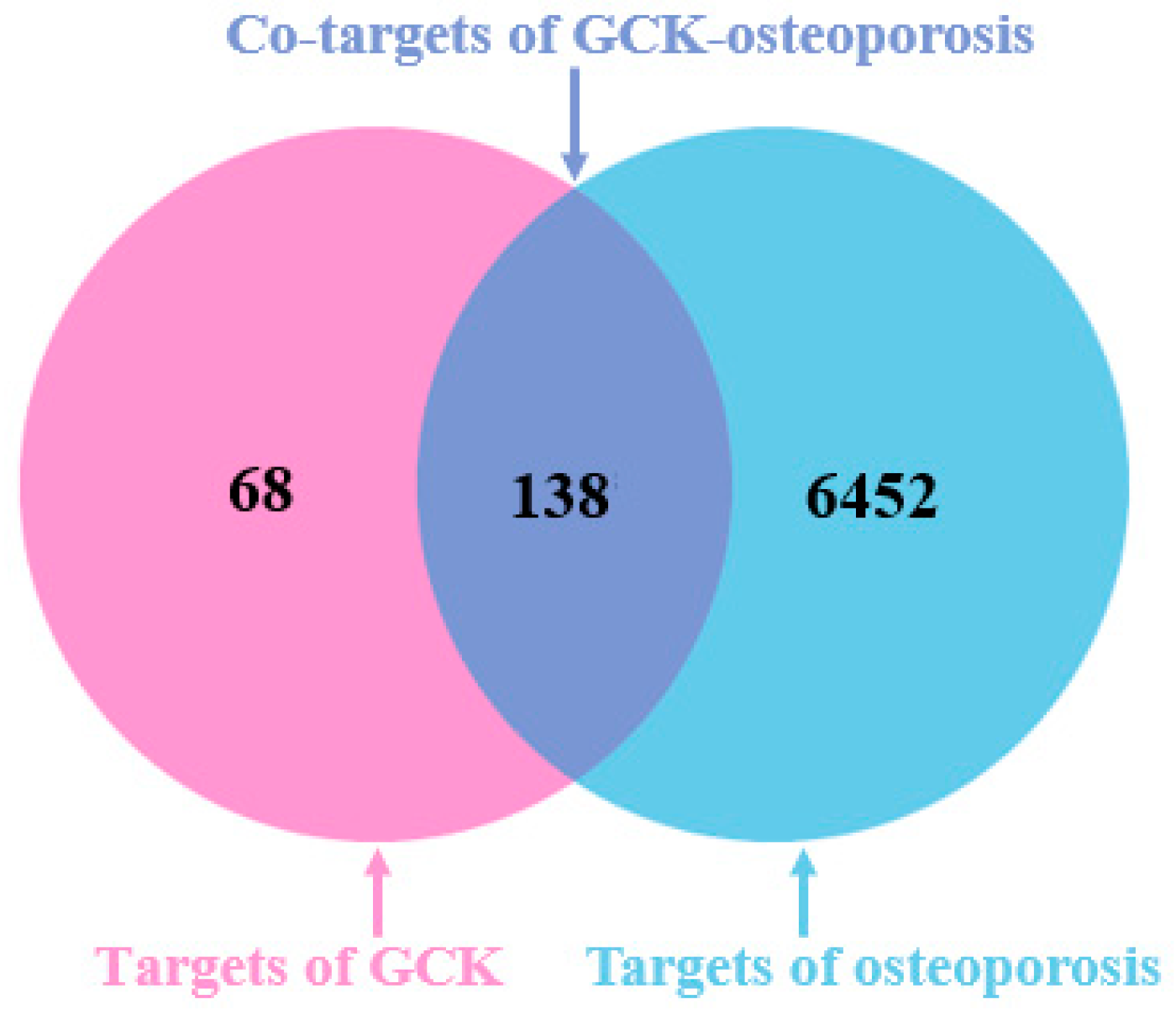
2.1. Molecular Targets of GCK and Osteoporosis
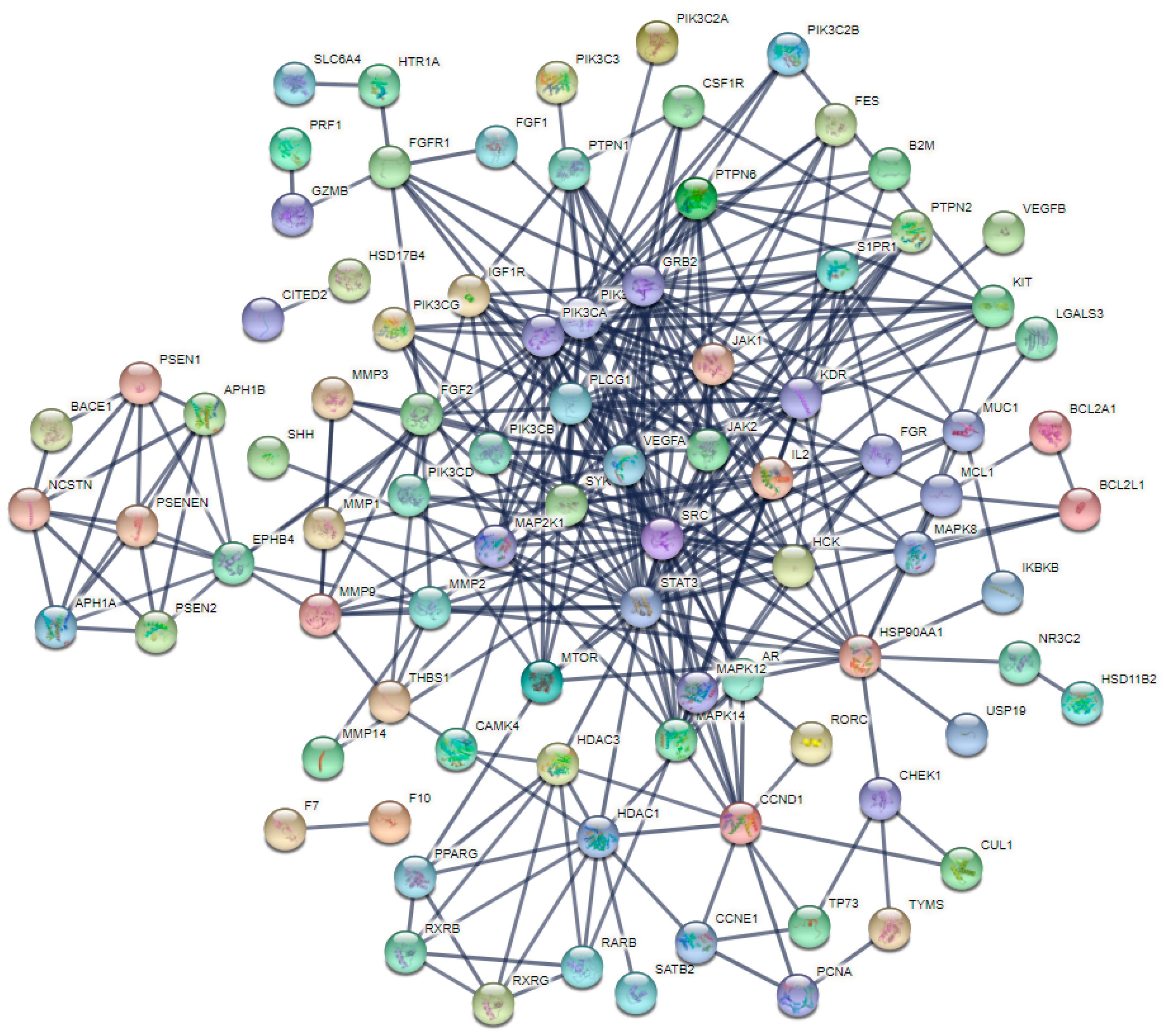
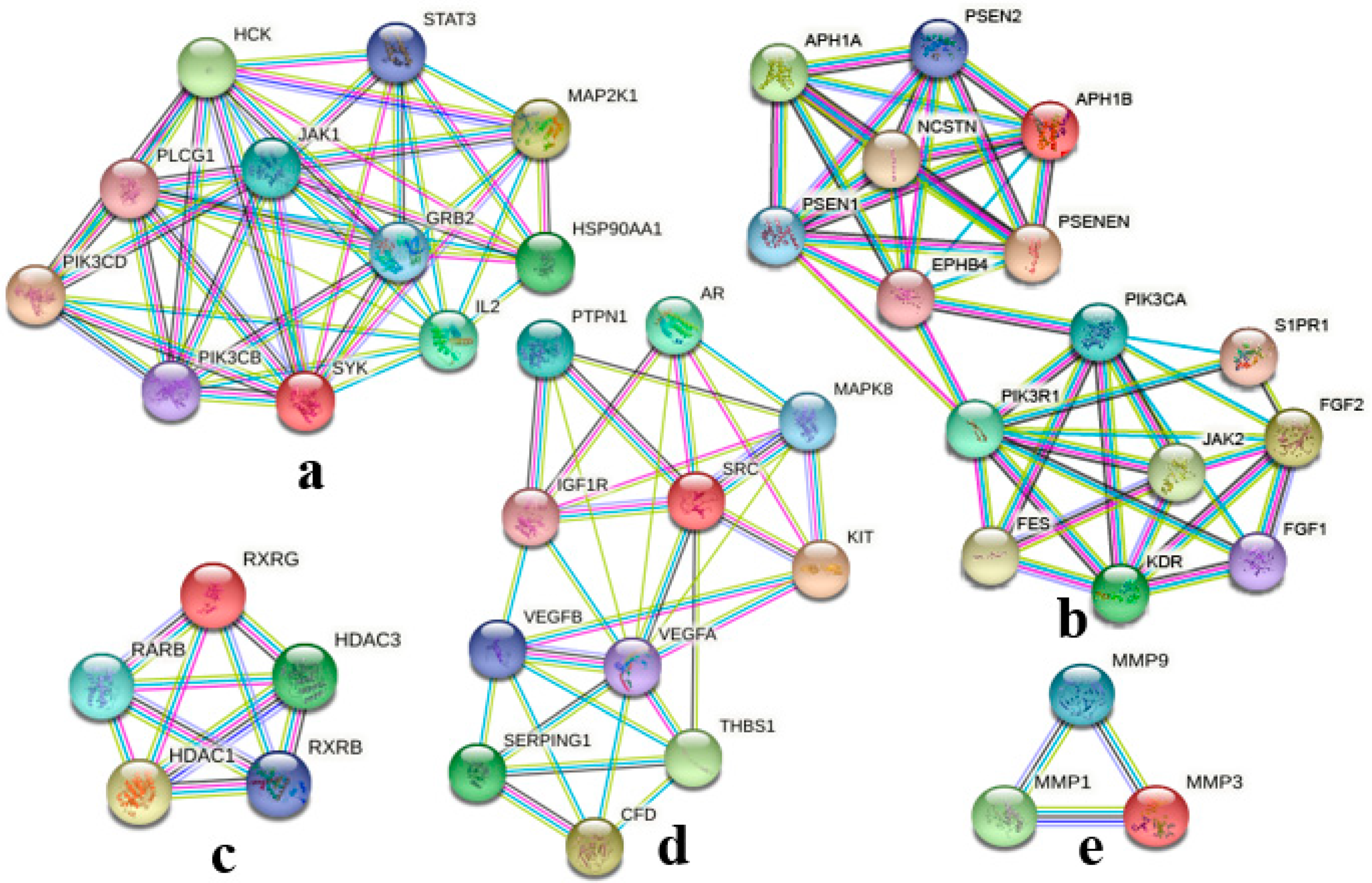
2.2. PPI Network Analysis of Co-Targets of GCK and Osteoporosis
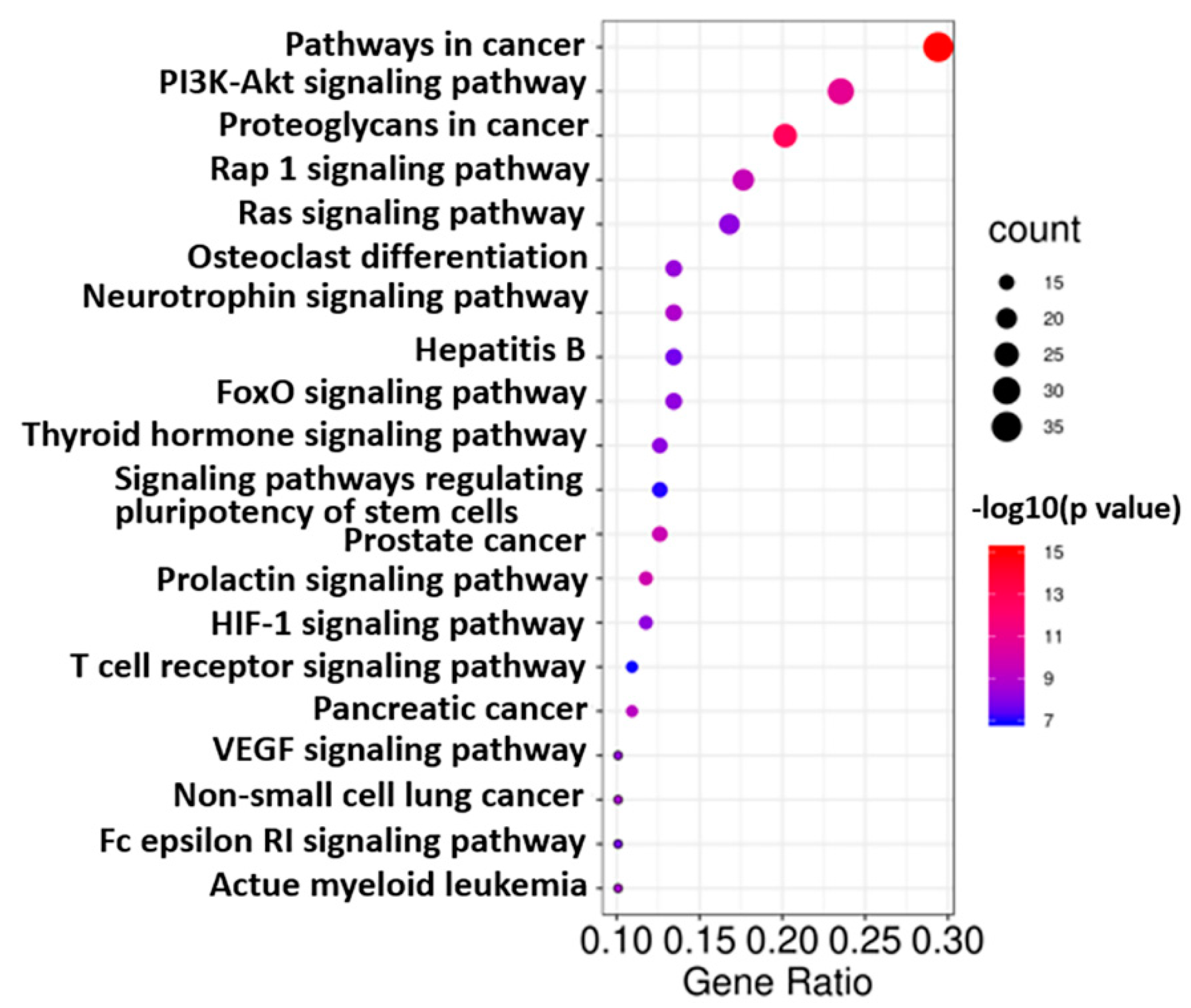
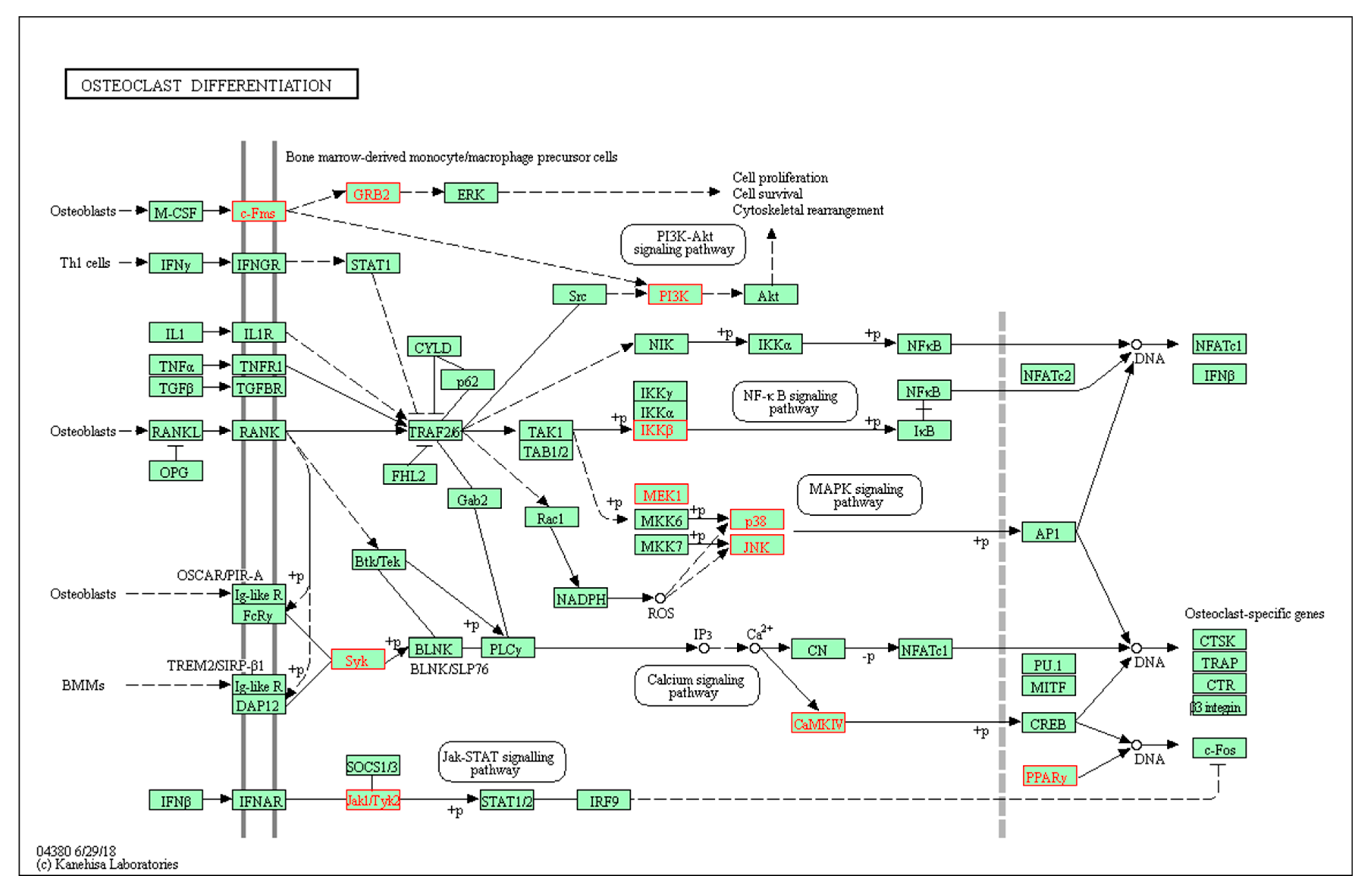
2.3. GO and KEGG Analysis of Co-Targets of GCK and Osteoporosis
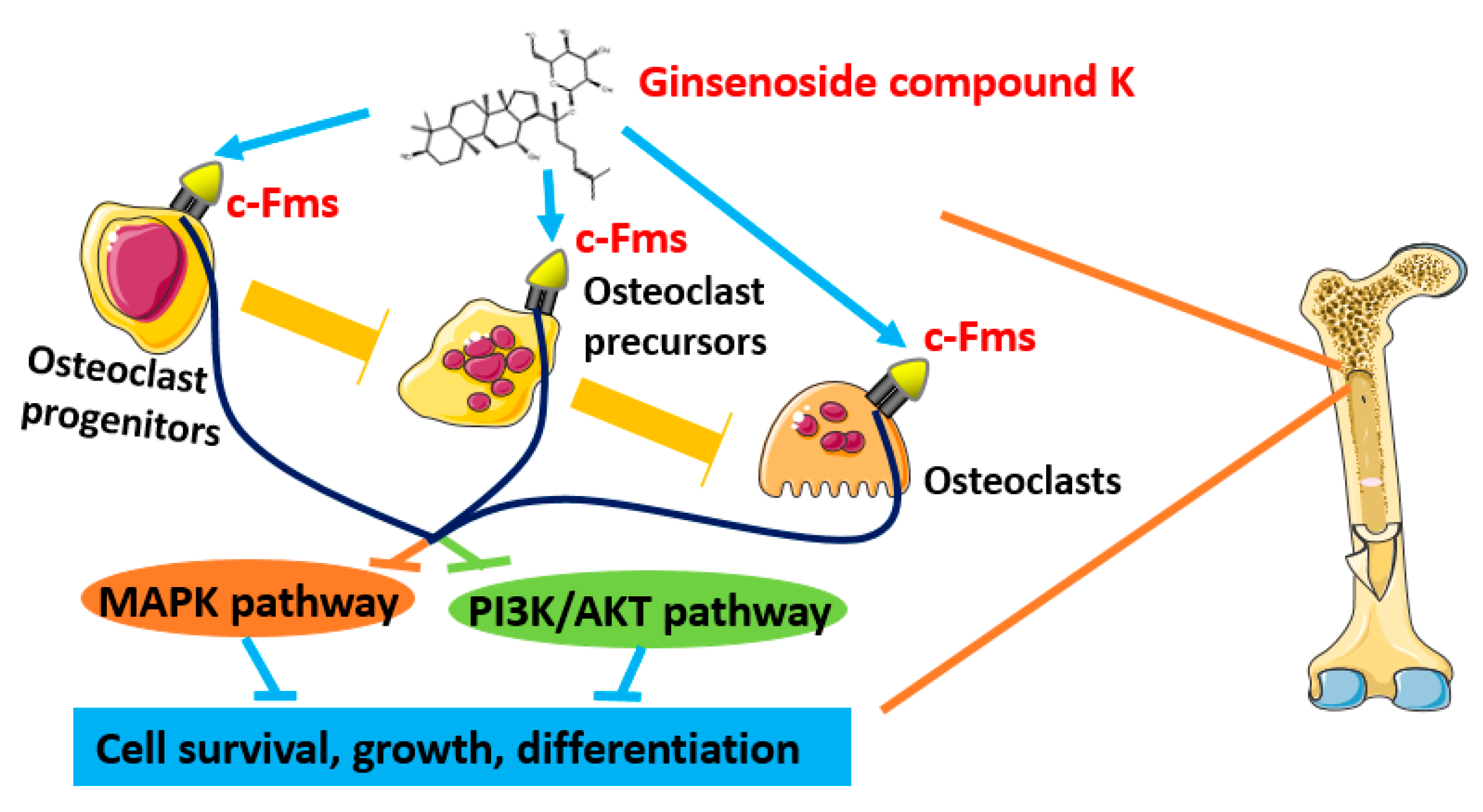
2.4. Molecular Docking of GCK- c-Fms Interaction
3. Discussion
4. Materials and Methods
4.1. Targets of GCK
4.2. Targets of Osteoporosis
4.3. Putative Targets of GCK-Treated Osteoporosis
4.4. Protein–Protein Interactions (PPIs), Network Construction and Analysis
4.5. Enrichment Analysis
4.6. Molecular Docking of Compound–Target Interaction
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Qin, C.; Hu, J.; Wan, Y.; Cai, M.; Wang, Z.; Peng, Z.; Liao, Y.; Li, D.; Yao, P.; Liu, L.; et al. Narrative review on potential role of gut microbiota in certain substance addiction. Prog. Neuropsychopharmacol. Biol. Psychiatry 2021, 106, 110093. [Google Scholar] [CrossRef] [PubMed]
- Eastell, R.; O’Neill, T.W.; Hofbauer, L.C.; Langdahl, B.; Reid, I.R.; Gold, D.T.; Cummings, S.R. Postmenopausal osteoporosis. Nat. Rev. Dis. Prim. 2016, 2, 16069. [Google Scholar] [CrossRef] [PubMed]
- Kanis, J.A.; McCloskey, E.V.; Johansson, H.; Oden, A. Approaches to the targeting of treatment for osteoporosis. Nat. Rev. Rheumatol. 2009, 5, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.; Jia, L.; Tang, Z.; Zheng, Y. Long non-coding RNA MIR22HG promotes osteogenic differentiation of bone marrow mesenchymal stem cells via PTEN/ AKT pathway. Cell Death Dis. 2020, 11, 601. [Google Scholar] [CrossRef]
- Harvey, N.; Dennison, E.; Cooper, C. Osteoporosis: Impact on health and economics. Nat. Rev. Rheumatol. 2010, 6, 99–105. [Google Scholar] [CrossRef]
- Silva, A.M.; Moura, S.R.; Teixeira, J.H.; Barbosa, M.A.; Santos, S.G.; Almeida, M.I. Long noncoding RNAs: A missing link in osteoporosis. Bone Res. 2019, 7, 10. [Google Scholar] [CrossRef] [Green Version]
- Tella, S.H.; Gallagher, J.C. Prevention and treatment of postmenopausal osteoporosis. J. Steroid. Biochem. Mol. Biol. 2014, 142, 155–170. [Google Scholar] [CrossRef] [Green Version]
- Eriksen, E.F.; Diez-Perez, A.; Boonen, S. Update on long-term treatment with bisphosphonates for postmenopausal osteoporosis: A systematic review. Bone 2014, 58, 126–135. [Google Scholar] [CrossRef]
- Niu, Y.B.; Yang, Y.Y.; Xiao, X.; Sun, Y.; Zhou, Y.M.; Zhang, Y.H.; Dong, D.; Li, C.R.; Wu, X.L.; Li, Y.H.; et al. Quercetin prevents bone loss in hindlimb suspension mice via stanniocalcin 1-mediated inhibition of osteoclastogenesis. Acta Pharm. Sin. 2020, 41, 1476–1486. [Google Scholar] [CrossRef]
- Gong, Y.S.; Chen, J.; Zhang, Q.Z.; Zhang, J.T. Effect of 17beta-oestradiol and ginsenoside on osteoporosis in ovariectomised rats. J. Asian Nat. Prod. Res. 2006, 8, 649–656. [Google Scholar] [CrossRef]
- Yang, N.; Liu, D.; Zhang, X.; Li, J.; Wang, M.; Xu, T.; Liu, Z. Effects of ginsenosides on bone remodelling for novel drug applications: A review. Chin. Med. 2020, 15, 42. [Google Scholar] [CrossRef]
- Zhang, X.; Huang, F.; Chen, X.; Wu, X.; Zhu, J. Ginsenoside Rg3 attenuates ovariectomy-induced osteoporosis via AMPK/mTOR signaling pathway. Drug Dev. Res. 2020, 81, 875–884. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Lee, J.; Jang, J.H.; Lee, S.H.; Nan, M.H.; Oh, B.C.; Lee, S.G.; Kim, H.H.; Soung, N.K.; Ahn, J.S.; et al. Ginsenoside Rh2 inhibits osteoclastogenesis through down-regulation of NF-kappaB, NFATc1 and c-Fos. Bone 2012, 50, 1207–1213. [Google Scholar] [CrossRef] [PubMed]
- Su, K.; Qiu, Y.-C.; Wang, C.-M.; Yuan, Z. Effects of ginsenoside Rb2 on osteoclast regulation in vitro. Chin. J. Bone Jt. 2017, 6, 874–877. [Google Scholar]
- Han, J.; Wang, Y.; Cai, E.; Zhang, L.; Zhao, Y.; Sun, N.; Zheng, X.; Wang, S. Study of the Effects and Mechanisms of Ginsenoside Compound K on Myelosuppression. J. Agric. Food Chem. 2019, 67, 1402–1408. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Lee, H.-J. Ginsenoside Compound K: Insights into Recent Studies on Pharmacokinetics and Health-Promoting Activities. Biomolecules 2020, 10, 1028. [Google Scholar] [CrossRef]
- He, D.; Huang, J.H.; Zhang, Z.Y.; Du, Q.; Peng, W.J.; Yu, R.; Zhang, S.F.; Zhang, S.H.; Qin, Y.H. A Network Pharmacology-Based Strategy For Predicting Active Ingredients And Potential Targets Of LiuWei DiHuang Pill In Treating Type 2 Diabetes Mellitus. Drug Des. Dev. Ther. 2019, 13, 3989–4005. [Google Scholar] [CrossRef] [Green Version]
- Bai, C.; Lei, X. New trends in population aging and challenges for China’s sustainable development. China Econ. J. 2019, 13, 3–23. [Google Scholar] [CrossRef]
- Cheng, B.; Li, J.; Du, J.; Lv, X.; Weng, L.; Ling, C. Ginsenoside Rb1 inhibits osteoclastogenesis by modulating NF-kappaB and MAPKs pathways. Food Chem. Toxicol. 2012, 50, 1610–1615. [Google Scholar] [CrossRef]
- Ma, Y.; Ran, D.; Zhao, H.; Song, R.; Zou, H.; Gu, J.; Yuan, Y.; Bian, J.; Zhu, J.; Liu, Z. Cadmium exposure triggers osteoporosis in duck via P2X7/PI3K/AKT-mediated osteoblast and osteoclast differentiation. Sci. Total Environ. 2021, 750, 141638. [Google Scholar] [CrossRef]
- Sanpaolo, E.R.; Rotondo, C.; Cici, D.; Corrado, A.; Cantatore, F.P. JAK/STAT pathway and molecular mechanism in bone remodeling. Mol. Biol. Rep. 2020, 47, 9087–9096. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.Y.; Putney, J.W., Jr. Calcium signaling in osteoclasts. Biochim. Biophys. Acta 2011, 1813, 979–983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, E.O.; Cha, K.H.; Lee, E.H.; Kim, S.M.; Choi, S.W.; Pan, C.H.; Um, B.H. Bioavailability of ginsenosides from white and red ginsengs in the simulated digestion model. J. Agric. Food Chem. 2014, 62, 10055–10063. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Zhu, X.M.; Hu, J.N.; Ye, H.; Luo, T.; Liu, X.R.; Li, H.Y.; Li, W.; Zheng, Y.N.; Deng, Z.Y. Absorption mechanism of ginsenoside compound K and its butyl and octyl ester prodrugs in Caco-2 cells. J. Agric. Food Chem. 2012, 60, 10278–10284. [Google Scholar] [CrossRef] [PubMed]
- Schubert, C.; Schalk-Hihi, C.; Struble, G.T.; Ma, H.C.; Petrounia, I.P.; Brandt, B.; Deckman, I.C.; Patch, R.J.; Player, M.R.; Spurlino, J.C.; et al. Crystal structure of the tyrosine kinase domain of colony-stimulating factor-1 receptor (cFMS) in complex with two inhibitors. J. Biol. Chem. 2007, 282, 4094–4101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldberg, F.W.; Finlay, M.R.V.; Ting, A.K.T.; Beattie, D.; Lamont, G.M.; Fallan, C.; Wrigley, G.L.; Schimpl, M.; Howard, M.R.; Williamson, B.; et al. The Discovery of 7-Methyl-2-[(7-methyl[1,2,4]triazolo[1,5-a]pyridin-6-yl)amino]-9-(tetrahydro-2H-p yran-4-yl)-7,9-dihydro-8H-purin-8-one (AZD7648), a Potent and Selective DNA-Dependent Protein Kinase (DNA-PK) Inhibitor. J. Med. Chem. 2020, 63, 3461–3471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.F.; Wang, Y.J.; Li, T.Y.; Guo, J.X.; Lv, F.; Li, C.L.; Ge, X.T. Colony-stimulating factor 1 receptor inhibition prevents against lipopolysaccharide -induced osteoporosis by inhibiting osteoclast formation. Biomed. Pharm. 2019, 115, 108916. [Google Scholar] [CrossRef]
- Mun, S.H.; Park, P.S.U.; Park-Min, K.H. The M-CSF receptor in osteoclasts and beyond. Exp. Mol. Med. 2020, 52, 1239–1254. [Google Scholar] [CrossRef]
- Zhou, R.P.; Lin, S.J.; Wan, W.B.; Zuo, H.L.; Yao, F.F.; Ruan, H.B.; Xu, J.; Song, W.; Zhou, Y.C.; Wen, S.Y.; et al. Chlorogenic Acid Prevents Osteoporosis by Shp2/PI3K/Akt Pathway in Ovariectomized Rats. PLoS ONE 2016, 11, e0166751. [Google Scholar] [CrossRef] [Green Version]
- Xi, J.C.; Zang, H.Y.; Guo, L.X.; Xue, H.B.; Liu, X.D.; Bai, Y.B.; Ma, Y.Z. The PI3K/AKT cell signaling pathway is involved in regulation of osteoporosis. J. Recept. Signal Transduct. 2015, 35, 640–645. [Google Scholar] [CrossRef]
- Fu, L.; Wu, W.; Sun, X.; Zhang, P. Glucocorticoids Enhanced Osteoclast Autophagy Through the PI3K/Akt/mTOR Signaling Pathway. Calcif. Tissue Int. 2020, 107, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Niu, X.; Bao, X.; Wang, Q.; Zhang, J.; Lu, S.; Wang, Y.; Xu, L.; Wang, M.; Zhang, J. The PI3K inhibitor buparlisib suppresses osteoclast formation and tumour cell growth in bone metastasis of lung cancer, as evidenced by multimodality molecular imaging. Oncol. Rep. 2019, 41, 2636–2646. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Fan, J.; Zhang, L.; Ning, Z.; Zeng, J.; Zhou, J.; Li, L.; Chen, Y.; Zhang, T.; Wang, X.; et al. PI3K/Akt to GSK3beta/beta-catenin signaling cascade coordinates cell colonization for bladder cancer bone metastasis through regulating ZEB1 transcription. Cell Signal 2012, 24, 2273–2282. [Google Scholar] [CrossRef] [PubMed]
- Agthong, S.; Kaewsema, A.; Tanomsridejchai, N.; Chentanez, V. Activation of MAPK ERK in peripheral nerve after injury. BMC Neurosci. 2006, 7, 45. [Google Scholar] [CrossRef] [Green Version]
- Ross, F.P.; Teitelbaum, S.L. avb3 and macrophage colonystimulating factor: Partners in osteoclast biology. Immunol. Rev. 2005, 208, 88–105. [Google Scholar] [CrossRef]
- Roy, B. Biomolecular basis of the role of diabetes mellitus in osteoporosis and bone fractures. World J. Diabetes 2013, 4, 101–113. [Google Scholar] [CrossRef]
- Siddiqi, M.H.; Siddiqi, M.Z.; Kang, S.; Noh, H.Y.; Ahn, S.; Simu, S.Y.; Aziz, M.A.; Sathishkumar, N.; Pérez, Z.E.J.; Yang, D.C. Inhibition of osteoclast differentiation by ginsenoside Rg3 in RAW264.7 cells via RANKL, JNK and p38 MAPK pathways through a modulation of cathepsin K: An in silico and in vitro study. Phytother. Res. 2015, 29, 1286–1294. [Google Scholar] [CrossRef]
- Chin, C.-H.; Chen, S.-H.; Wu, H.-H.; Ho, C.-W.; Ko, M.-T.; Lin, C.-Y. cytoHubba: Identifying hub objects and sub-networks from complex interactome. BMC Syst. Biol. 2014, 8, S11. [Google Scholar] [CrossRef] [Green Version]
- Morris, G.M.; Goodsell, D.S.; Halliday, R.S.; Huey, R.; Hart, W.E.; Belew, R.K.; Olson, A.J. Automated docking using a Lamarckian genetic algorithm and an empirical binding free energy function. J. Comput. Chem. 1998, 19, 1639–1662. [Google Scholar] [CrossRef]
- Kerwin, S.M. ChemBioOffice Ultra 2010 Suite. J. Am. Chem. Soc. 2010, 132, 2466. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cluster | Nodes | Edges | Score | Genes |
|---|---|---|---|---|
| 1 | 11 | 36 | 7.200 | SYK; STAT3; PLCG1; PIK3CD; PIK3CB; MAP2K1; JAK1; IL2; HSP90AA1; HCK; GRB2 |
| 2 | 15 | 40 | 5.714 | S1PR1; PSENEN; PSEN2; PSEN1; PIK3R1; PIK3CA; NCSTN; KDR; JAK2; FGF2; FGF1; FES; EPHB4; APH1B; APH1A; |
| 3 | 5 | 10 | 5.000 | RXRG; RXRB; RARB; HDAC3; HDAC1 |
| 4 | 11 | 20 | 4.000 | VEGFB; VEGFA; THBS1; SRC; SERPING1; PTPN1; MAPK8; KIT; IGF1R; CFD; AR |
| 5 | 3 | 3 | 3.000 | MMP9; MMP3; MMP1 |
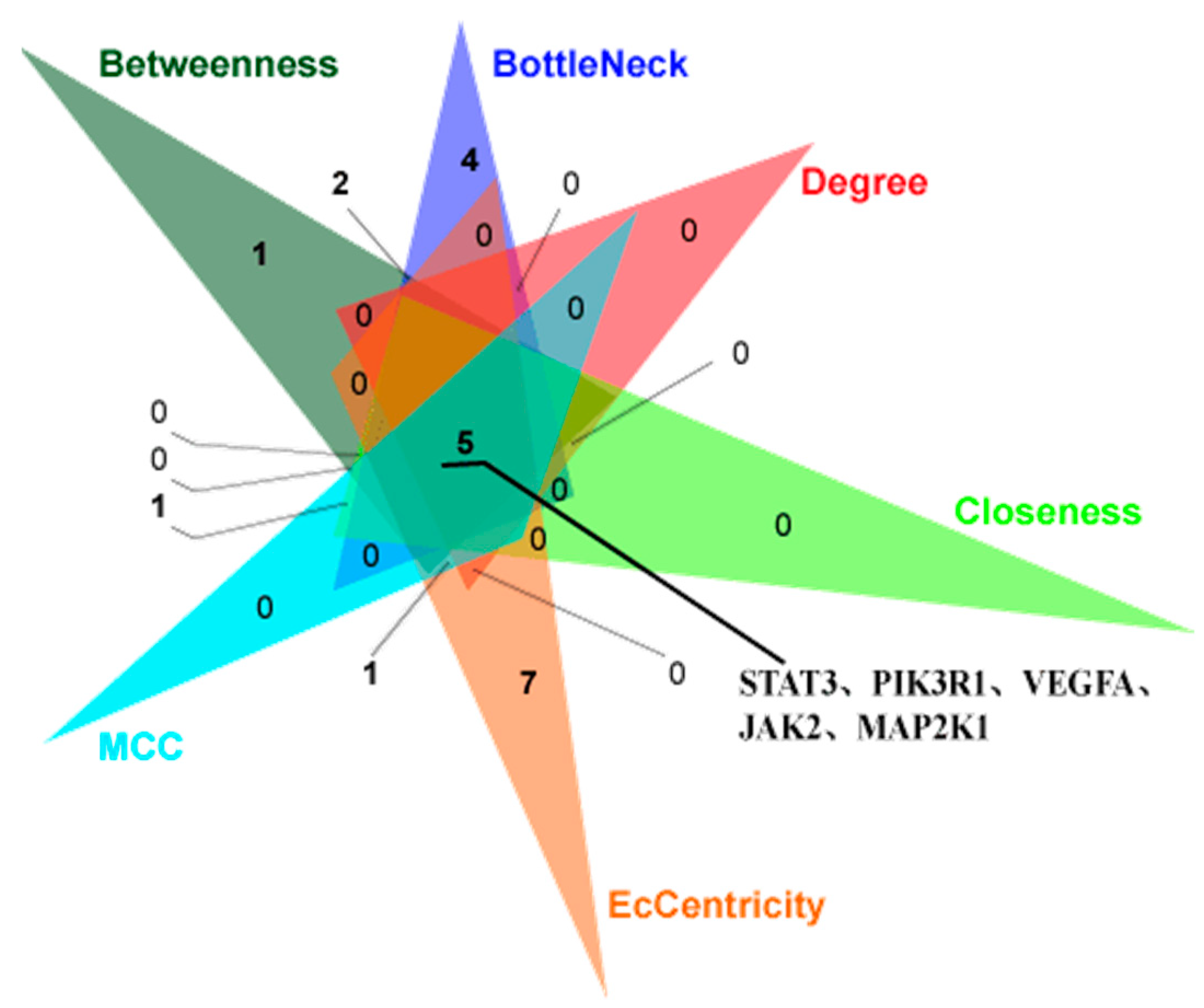
| Methods | Betweenness | BottleNeck | Degree | Closeness | MCC | EcCentricity | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gene | Score | Gene | Score | Gene | Score | Gene | Score | Gene | Score | Gene | Score | |
| 1 | STAT3 | 1858.83 | PIK3R1 | 32.00 | PIK3R1 | 35.00 | PIK3R1 | 60.00 | PIK3R1 | 33,711.00 | PIK3CA | 0.32 |
| 2 | PIK3R1 | 1514.88 | STAT3 | 17.00 | PIK3CA | 34.00 | PIK3CA | 59.33 | PIK3CA | 33,710.00 | PIK3R1 | 0.32 |
| 3 | PIK3CA | 1166.88 | MMP9 | 12.00 | STAT3 | 30.00 | STAT3 | 57.08 | GRB2 | 23,136.00 | MMP9 | 0.24 |
| 4 | EPHB4 | 1159.36 | GRB2 | 9.00 | SRC | 23.00 | SRC | 52.00 | PIK3CB | 22,560.00 | HCK | 0.24 |
| 5 | VEGFA | 568.08 | EPHB4 | 8.00 | GRB2 | 22.00 | GRB2 | 51.75 | PLCG1 | 21,770.00 | SYK | 0.24 |
| 6 | MTOR | 477.35 | VEGFA | 7.00 | PLCG1 | 21.00 | VEGFA | 50.17 | JAK1 | 20,934.00 | PTPN6 | 0.24 |
| 7 | PLCG1 | 449.22 | MAPK14 | 6.00 | VEGFA | 20.00 | JAK2 | 49.67 | SYK | 17,220.00 | IGF1R | 0.24 |
| 8 | CCND1 | 400.60 | MTOR | 4.00 | JAK2 | 19.00 | PLCG1 | 49.08 | HCK | 16,812.00 | PIK3C2B | 0.24 |
| 9 | MAPK14 | 384.62 | KIT | 4.00 | JAK1 | 19.00 | JAK1 | 49.08 | SRC | 11,096.00 | MAP2K1 | 0.24 |
| 10 | JAK2 | 377.93 | S1PR1 | 4.00 | IL2 | 17.00 | IL2 | 48.42 | JAK2 | 10,906.00 | STAT3 | 0.24 |
| 11 | MMP2 | 328.71 | PTPN6 | 3.00 | PIK3CB | 17.00 | HSP90AA1 | 46.42 | IL2 | 8767.00 | MTOR | 0.24 |
| 12 | KIT | 317.35 | MAP2K1 | 3.00 | SYK | 15.00 | PIK3CB | 46.00 | STAT3 | 7819.00 | JAK2 | 0.24 |
| 13 | IL2 | 299.91 | JAK2 | 3.00 | KDR | 15.00 | KDR | 46.00 | PIK3CD | 7440.00 | MMP1 | 0.24 |
| 14 | SRC | 296.80 | PPARG | 3.00 | PTPN6 | 14.00 | PTPN6 | 45.42 | MAP2K1 | 5929.00 | PIK3CD | 0.24 |
| 15 | MMP9 | 262.75 | RARB | 3.00 | MAPK14 | 14.00 | HCK | 45.33 | KDR | 2580.00 | MMP3 | 0.24 |
| 16 | HDAC3 | 260.66 | CHEK1 | 3.00 | HCK | 13.00 | KIT | 45.00 | VEGFA | 2392.00 | VEGFA | 0.24 |
| 17 | S1PR1 | 237.33 | MMP2 | 3.00 | HSP90AA1 | 13.00 | SYK | 44.92 | S1PR1 | 1538.00 | KIT | 0.24 |
| 18 | GRB2 | 225.02 | F10 | 3.00 | MAP2K1 | 12.00 | MAPK14 | 44.87 | FGF2 | 1344.00 | PTPN1 | 0.24 |
| 19 | PTPN6 | 212.20 | KDR | 3.00 | S1PR1 | 12.00 | MAP2K1 | 44.75 | HSP90AA1 | 1130.00 | ANXA1 | 0.24 |
| 20 | MAP2K1 | 205.76 | HDAC3 | 3.00 | MMP9 | 11.00 | FGF2 | 44.75 | EPHB4 | 724.00 | GNRHR | 0.24 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, S.; Shen, S.; Ma, P.; Fan, D. Biochemical Targets and Molecular Mechanism of Ginsenoside Compound K in Treating Osteoporosis Based on Network Pharmacology. Int. J. Mol. Sci. 2022, 23, 13921. https://doi.org/10.3390/ijms232213921
Zhang S, Shen S, Ma P, Fan D. Biochemical Targets and Molecular Mechanism of Ginsenoside Compound K in Treating Osteoporosis Based on Network Pharmacology. International Journal of Molecular Sciences. 2022; 23(22):13921. https://doi.org/10.3390/ijms232213921
Chicago/Turabian StyleZhang, Sen, Shihong Shen, Pei Ma, and Daidi Fan. 2022. "Biochemical Targets and Molecular Mechanism of Ginsenoside Compound K in Treating Osteoporosis Based on Network Pharmacology" International Journal of Molecular Sciences 23, no. 22: 13921. https://doi.org/10.3390/ijms232213921