
Using Computational Drug-Gene Analysis to Identify Novel Therapeutic Candidates for Retinal Neuroprotection
 , , and
, , and
Abstract
:1. Introduction
2. Results
3. Discussion
Limitations
4. Materials and Methods
4.1. Literature Search and Data Extraction
4.2. Discovering Potential Neuroprotection Therapeutic Targets via Enrichment Analysis
4.3. Narrowing down Drugs/Chemicals Useful in Neuroprotection
4.4. Visualization of Networks
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Weber, A.J.; Harman, C.D.; Viswanathan, S. Effects of optic nerve injury, glaucoma, and neuroprotection on the survival, structure, and function of ganglion cells in the mammalian retina. J. Physiol. 2008, 586, 4393–4400. [Google Scholar] [CrossRef] [PubMed]
- Pardue, M.T.; Allen, R.S. Neuroprotective strategies for retinal disease. Prog. Retin. Eye Res. 2018, 65, 50–76. [Google Scholar] [CrossRef] [PubMed]
- Yonekawa, Y.; Miller, J.W.; Kim, I.K. Age-Related Macular Degeneration: Advances in Management and Diagnosis. J. Clin. Med. 2015, 4, 343–359. [Google Scholar] [CrossRef] [Green Version]
- Bernstein, P.S.; Li, B.; Vachali, P.P.; Gorusupudi, A.; Shyam, R.; Henriksen, B.S.; Nolan, J.M. Lutein, zeaxanthin, and meso-zeaxanthin: The basic and clinical science underlying carotenoid-based nutritional interventions against ocular disease. Prog. Retin. Eye Res. 2015, 50, 34–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrone, L.A.; Rombolà, L.; Corasaniti, M.T.; Bagetta, G.; Nucci, C.; Russo, R. Natural compounds and retinal ganglion cell neuroprotection. Prog. Brain Res. 2015, 220, 257–281. [Google Scholar] [CrossRef] [PubMed]
- Handa, J.T.; Rickman, C.B.; Dick, A.D.; Gorin, M.B.; Miller, J.W.; Toth, C.A.; Ueffing, M.; Zarbin, M.; Farrer, L.A. A systems biology approach towards understanding and treating non-neovascular age-related macular degeneration. Nat. Commun. 2019, 10, 3347. [Google Scholar] [CrossRef] [Green Version]
- Pool, F.M.; Kiel, C.; Serrano, L.; Luthert, P.J. Repository of proposed pathways and protein–protein interaction networks in age-related macular degeneration. NPJ Aging Mech. Dis. 2020, 6, 2. [Google Scholar] [CrossRef] [Green Version]
- Oulas, A.; Minadakis, G.; Zachariou, M.; Sokratous, K.; Bourdakou, M.M.; Spyrou, G.M. Systems Bioinformatics: Increasing precision of computational diagnostics and therapeutics through network-based approaches. Brief. Bioinform. 2019, 20, 806–824. [Google Scholar]
- Chu, H.; Sun, P.; Yin, J.; Liu, G.; Wang, Y.; Zhao, P.; Zhu, Y.; Yang, X.; Zheng, T.; Zhou, X.; et al. Integrated network analysis reveals potentially novel molecular mechanisms and therapeutic targets of refractory epilepsies. PLoS ONE 2017, 12, e0174964. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Zang, W.; Xue, F.; Shen, Z.; Zhang, Q. Bioinformatics analysis reveals potential candidate drugs for different subtypes of glioma. Neurol. Sci. 2012, 34, 1139–1143. [Google Scholar] [CrossRef]
- Hurgobin, B.; de Jong, E.; Bosco, A. Insights into respiratory disease through bioinformatics. Respirology 2018, 23, 1117–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Wang, Z.; Shen, X.; Cui, X.; Guo, Y. Identification of novel biomarkers and small molecule drugs in human colorectal cancer by microarray and bioinformatics analysis. Mol. Genet. Genom. Med. 2019, 7, e00713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siavelis, J.C.; Bourdakou, M.M.; Athanasiadis, E.I.; Spyrou, G.M.; Nikita, K.S. Bioinformatics methods in drug repurposing for Alzheimer’s disease. Brief. Bioinform. 2016, 17, 322–335. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Yuan, M.; Xin, J.; Liu, X.; Wang, J. Screening novel drug candidates for Alzheimer’s disease by an integrated network and transcriptome analysis. Bioinformatics 2020, 36, 4626–4632. [Google Scholar] [CrossRef]
- Nadeem, U.; Xie, B.; Xie, E.F.; D’Souza, M.; Dao, D.; Sulakhe, D.; Skondra, D. Using Advanced Bioinformatics Tools to Identify Novel Therapeutic Candidates for Age-Related Macular Degeneration. Transl. Vis. Sci. Technol. 2022, 11, 10. [Google Scholar] [CrossRef]
- Platania, C.B.M.; Leggio, G.M.; Drago, F.; Salomone, S.; Bucolo, C. Computational systems biology approach to identify novel pharmacological targets for diabetic retinopathy. Biochem. Pharmacol. 2018, 158, 13–26. [Google Scholar] [CrossRef]
- Pushpakom, S.; Iorio, F.; Eyers, P.A.; Escott, K.J.; Hopper, S.; Wells, A.; Doig, A.; Guilliams, T.; Latimer, J.; McNamee, C.; et al. Drug repurposing: Progress, challenges and recommendations. Nat. Rev. Drug Discov. 2019, 18, 41–58. [Google Scholar] [CrossRef]
- Gesualdo, C.; Balta, C.; Platania, C.B.M.; Trotta, M.C.; Herman, H.; Gharbia, S.; Rosu, M.; Petrillo, F.; Giunta, S.; Della Corte, A.; et al. Fingolimod and Diabetic Retinopathy: A Drug Repurposing Study. Front. Pharmacol. 2021, 12, 718902. [Google Scholar] [CrossRef]
- Platania, C.B.M.; Ronchetti, S.; Riccardi, C.; Migliorati, G.; Marchetti, M.C.; Di Paola, L.; Lazzara, F.; Drago, F.; Salomone, S.; Bucolo, C. Effects of protein-protein interface disruptors at the ligand of the glucocorticoid-induced tumor necrosis factor receptor-related gene (GITR). Biochem. Pharmacol. 2020, 178, 114110. [Google Scholar] [CrossRef]
- Nosengo, N. Can you teach old drugs new tricks? Nature 2016, 534, 314–316. [Google Scholar] [CrossRef] [Green Version]
- López-Malo, D.; Villarón-Casares, C.A.; Alarcón-Jiménez, J.; Miranda, M.; Díaz-Llopis, M.; Romero, F.J.; Villar, V.M. Curcumin as a Therapeutic Option in Retinal Diseases. Antioxidants 2020, 9, 48. [Google Scholar] [CrossRef] [PubMed]
- Akinleye, A.; Furqan, M.; Mukhi, N.; Ravella, P.; Liu, D. MEK and the inhibitors: From bench to bedside. J. Hematol. Oncol. 2013, 6, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, J.Y. Neuroprotection signaling of nuclear akt in neuronal cells. Exp. Neurobiol. 2014, 23, 200–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beatty, S.; Koh, H.; Phil, M.; Henson, D.; Boulton, M. The role of oxidative stress in the pathogenesis of age-related macular degeneration. Surv. Ophthalmol. 2000, 45, 115–134. [Google Scholar] [CrossRef] [Green Version]
- Levkovitch-Verbin, H. Retinal ganglion cell apoptotic pathway in glaucoma: Initiating and downstream mechanisms. Prog. Brain Res. 2015, 220, 37–57. [Google Scholar] [CrossRef]
- Frank, R.N. Diabetic retinopathy. N. Engl. J. Med. 2004, 350, 48–58. [Google Scholar] [CrossRef]
- Datta, S.; Cano, M.; Ebrahimi, K.; Wang, L.; Handa, J.T. The impact of oxidative stress and inflammation on RPE degeneration in non-neovascular AMD. Prog. Retin. Eye Res. 2017, 60, 201–218. [Google Scholar] [CrossRef]
- Kang, Q.; Yang, C. Oxidative stress and diabetic retinopathy: Molecular mechanisms, pathogenetic role and therapeutic implications. Redox Biol. 2020, 37, 101799. [Google Scholar] [CrossRef]
- Agus, D.B.; Gambhir, S.S.; Pardridge, W.M.; Spielholz, C.; Baselga, J.; Vera, J.C.; Golde, D.W. Vitamin C crosses the blood-brain barrier in the oxidized form through the glucose transporters. J. Clin. Investig. 1997, 100, 2842–2848. [Google Scholar] [CrossRef] [Green Version]
- Sano, H.; Namekata, K.; Kimura, A.; Shitara, H.; Guo, X.; Harada, C.; Mitamura, Y.; Harada, T. Differential effects of N-acetylcysteine on retinal degeneration in two mouse models of normal tension glaucoma. Cell Death Dis. 2019, 10, 75. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Zhang, X.L.; Zhu, B.F.; Ding, Y.N. Effect of antioxidant N-acetylcysteine on diabetic retinopathy and expression of VEGF and ICAM-1 from retinal blood vessels of diabetic rats. Mol. Biol. Rep. 2011, 39, 3727–3735. [Google Scholar] [CrossRef] [PubMed]
- Ajith, T.A. Alpha-lipoic acid: A possible pharmacological agent for treating dry eye disease and retinopathy in diabetes. Clin. Exp. Pharmacol. Physiol. 2020, 47, 1883–1890. [Google Scholar] [CrossRef] [PubMed]
- Monti, D.A.; Zabrecky, G.; Kremens, D.; Liang, T.W.; Wintering, N.A.; Bazzan, A.J.; Zhong, L.; Bowens, B.K.; Chervoneva, I.; Intenzo, C.; et al. N-Acetyl Cysteine Is Associated With Dopaminergic Improvement in Parkinson’s Disease. Clin. Pharmacol. Ther. 2019, 106, 884–890. [Google Scholar] [CrossRef] [PubMed]
- Voloboueva, L.A.; Liu, J.; Suh, J.H.; Ames, B.N.; Miller, S.S. (R)-alpha-lipoic acid protects retinal pigment epithelial cells from oxidative damage. Investig. Opthalmol. Vis. Sci. 2005, 46, 4302–4310. [Google Scholar] [CrossRef] [Green Version]
- Kowluru, R.A.; Odenbach, S. Role of interleukin-1beta in the development of retinopathy in rats: Effect of antioxidants. Investig. Opthalmol. Vis. Sci. 2004, 45, 4161–4166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.F.; Zou, H.D. PEDF in diabetic retinopathy: A protective effect of oxidative stress. J. Biomed. Biotechnol. 2012, 2012, 580687. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.Y.; Usui, S.; Zafar, A.B.; Oveson, B.C.; Jo, Y.J.; Lu, L.; Masoudi, S.; Campochiaro, P.A. N-Acetylcysteine promotes long-term survival of cones in a model of retinitis pigmentosa. J. Cell. Physiol. 2011, 226, 1843–1849. [Google Scholar] [CrossRef]
- Campochiaro, P.A.; Iftikhar, M.; Hafiz, G.; Akhlaq, A.; Tsai, G.; Wehling, D.; Lu, L.; Wall, G.M.; Singh, M.S.; Kong, X. Oral N-acetylcysteine improves cone function in retinitis pigmentosa patients in phase I trial. J. Clin. Investig. 2020, 130, 1527–1541. [Google Scholar] [CrossRef] [Green Version]
- Pringsheim, T.; Davenport, W.; Mackie, G.; Worthington, I.; Aubé, M.; Christie, S.N.; Gladstone, J.; Becker, W.J. Canadian Headache Society Prophylactic Guidelines Development Group. Canadian Headache Society guideline for migraine prophylaxis. Can. J. Neurol. Sci. 2012, 39 (Suppl. S2), S1–S59. [Google Scholar]
- Dinte, E.; Vostinaru, O.; Samoila, O.; Sevastre, B.; Bodoki, E. Ophthalmic Nanosystems with Antioxidants for the Prevention and Treatment of Eye Diseases. Coatings 2020, 10, 36. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Medina, J.J.; Rubio-Velazquez, E.; Lopez-Bernal, M.D.; Cobo-Martinez, A.; Zanon-Moreno, V.; Pinazo-Duran, M.D.; Del-Rio-Vellosillo, M. Glaucoma and Antioxidants: Review and Update. Antioxidants 2020, 9, 1031. [Google Scholar] [CrossRef] [PubMed]
- Haritoglou, C.; Gerss, J.; Hammes, H.P.; Kampik, A.; Ulbig, M.W. RETIPON Study Group. Alpha-lipoic acid for the prevention of diabetic macular edema. Ophthalmologica 2011, 226, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Nebbioso, M.; Federici, M.; Rusciano, D.; Evangelista, M.; Pescosolido, N. Oxidative stress in preretinopathic diabetes subjects and antioxidants. Diabetes Technol. Ther. 2012, 14, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Medina, J.J.; Pinazo-Duran, M.D.; Garcia-Medina, M.; Zanon-Moreno, V.; Pons-Vazquez, S. A 5-year follow-up of antioxidant supplementation in type 2 diabetic retinopathy. Eur. J. Ophthalmol. 2011, 21, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Venditti, A.; Sharifi-Rad, M.; Kręgiel, D.; Sharifi-Rad, J.; Durazzo, A.; Lucarini, M.; Santini, A.; Souto, E.B.; Novellino, E.; et al. The Therapeutic Potential of Apigenin. Int. J. Mol. Sci. 2019, 20, 1305. [Google Scholar] [CrossRef] [Green Version]
- McCormack, D.; McFadden, D. A review of pterostilbene antioxidant activity and disease modification. Oxidative Med. Cell. Longev. 2013, 2013, 575482. [Google Scholar] [CrossRef] [Green Version]
- Al-Dosari, D.I.; Ahmed, M.M.; Al-Rejaie, S.S.; Alhomida, A.S.; Ola, M.S. Flavonoid Naringenin Attenuates Oxidative Stress, Apoptosis and Improves Neurotrophic Effects in the Diabetic Rat Retina. Nutrients 2017, 9, 1161. [Google Scholar] [CrossRef] [Green Version]
- Mandal, M.N.; Patlolla, J.M.; Zheng, L.; Agbaga, M.P.; Tran, J.T.; Wicker, L.; Kasus-Jacobi, A.; Elliott, M.H.; Rao, C.V.; Anderson, R.E. Curcumin protects retinal cells from light-and oxidant stress-induced cell death. Free Radic. Biol. Med. 2009, 46, 672–679. [Google Scholar] [CrossRef] [Green Version]
- Kowluru, R.A.; Kanwar, M. Effects of curcumin on retinal oxidative stress and inflammation in diabetes. Nutr. Metab. 2007, 4, 8. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Zuo, Z.; Lu, S.; Liu, A.; Liu, X. Naringin attenuates diabetic retinopathy by inhibiting inflammation, oxidative stress and NF-κB activation in vivo and in vitro. Iran J. Basic Med. Sci. 2017, 20, 813–821. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, Y.; Yu, H.; Li, M.; Hang, L.; Xu, X. Apigenin Protects Mouse Retina against Oxidative Damage by Regulating the Nrf2 Pathway and Autophagy. Oxid. Med. Cell. Longev. 2020, 2020, 9420704. [Google Scholar] [CrossRef] [PubMed]
- Muangnoi, C.; Ratnatilaka Na Bhuket, P.; Jithavech, P.; Supasena, W.; Paraoan, L.; Patumraj, S.; Rojsitthisak, P. Curcumin diethyl disuccinate, a prodrug of curcumin, enhances anti-proliferative effect of curcumin against HepG2 cells via apoptosis induction. Sci. Rep. 2019, 9, 11718. [Google Scholar] [CrossRef] [PubMed]
- Mazzolani, F.; Togni, S. Oral administration of a curcumin-phospholipid delivery system for the treatment of central serous chorioretinopathy: A 12-month follow-up study. Clin. Ophthalmol. 2013, 7, 939–945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehta, R. Effect of Oral Curcumin Supplementation in Dry Age-Related Macular Degeneration (AMD) Patient. Available online: https://clinicaltrials.gov/ct2/show/NCT04590196 (accessed on 22 August 2022).
- Han, Y.; Xie, H.; Liu, Y.; Gao, P.; Yang, X.; Shen, Z. Effect of metformin on all-cause and cardiovascular mortality in patients with coronary artery diseases: A systematic review and an updated meta-analysis. Cardiovasc Diabetol. 2019, 18, 96. [Google Scholar] [CrossRef] [Green Version]
- Kasznicki, J.; Sliwinska, A.; Drzewoski, J. Metformin in cancer prevention and therapy. Ann. Transl. Med. 2014, 2, 57. [Google Scholar] [CrossRef]
- Samaras, K.; Makkar, S.; Crawford, J.D.; Kochan, N.A.; Wen, W.; Draper, B.; Trollor, J.N.; Brodaty, H.; Sachdev, P.S. Metformin Use Is Associated With Slowed Cognitive Decline and Reduced Incident Dementia in Older Adults With Type 2 Diabetes: The Sydney Memory and Ageing Study. Diabetes Care 2020, 43, 2691–2701. [Google Scholar] [CrossRef]
- Han, J.; Li, Y.; Liu, X.; Zhou, T.; Sun, H.; Edwards, P.; Gao, H.; Yu, F.S.; Qiao, X. Metformin suppresses retinal angiogenesis and inflammation in vitro and in vivo. PLoS ONE 2018, 13, e0193031. [Google Scholar] [CrossRef] [Green Version]
- Mori, A.; Ishikawa, E.; Amano, T.; Sakamoto, K.; Nakahara, T. Anti-diabetic drug metformin dilates retinal blood vessels through activation of AMP-activated protein kinase in rats. Eur. J. Pharmacol. 2017, 798, 66–71. [Google Scholar] [CrossRef]
- Altmann, C.; Schmidt, M.H. The Role of Microglia in Diabetic Retinopathy: Inflammation, Microvasculature Defects and Neurodegeneration. Int. J. Mol. Sci. 2018, 19, 110. [Google Scholar] [CrossRef] [Green Version]
- Zeng, X.X.; Ng, Y.K.; Ling, E.A. Neuronal and microglial response in the retina of streptozotocin-induced diabetic rats. Vis. Neurosci. 2000, 17, 463–471. [Google Scholar] [CrossRef]
- Luodan, A.; Zou, T.; He, J.; Chen, X.; Sun, D.; Fan, X.; Xu, H. Rescue of Retinal Degeneration in rd1 Mice by Intravitreally Injected Metformin. Front. Mol. Neurosci. 2019, 12, 102. [Google Scholar] [CrossRef]
- Fan, Y.P.; Wu, C.T.; Lin, J.L.; Hsiung, C.A.; Liu, H.Y.; Lai, J.N.; Yang, C.C. Metformin Treatment Is Associated with a Decreased Risk of Nonproliferative Diabetic Retinopathy in Patients with Type 2 Diabetes Mellitus: A Population-Based Cohort Study. J. Diabetes Res. 2020, 2020, 9161039. [Google Scholar] [CrossRef] [PubMed]
- Blitzer, A.L.; Ham, S.A.; Colby, K.A.; Skondra, D. Association of Metformin Use With Age-Related Macular Degeneration: A Case-Control Study. JAMA Ophthalmol. 2021, 139, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Evangelho, K.; Mogilevskaya, M.; Losada-Barragan, M.; Vargas-Sanchez, J.K. Pathophysiology of primary open-angle glaucoma from a neuroinflammatory and neurotoxicity perspective: A review of the literature. Int. Ophthalmol. 2019, 39, 259–271. [Google Scholar] [CrossRef]
- Rossino, M.G.; Dal Monte, M.; Casini, G. Relationships Between Neurodegeneration and Vascular Damage in Diabetic Retinopathy. Front. Neurosci. 2019, 13, 1172. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, K.; Asano, D.; Ushikubo, H.; Morita, A.; Mori, A.; Sakamoto, K.; Ishii, K.; Nakahara, T. Metformin Protects against NMDA-Induced Retinal Injury through the MEK/ERK Signaling Pathway in Rats. Int. J. Mol. Sci. 2021, 22, 4439. [Google Scholar] [CrossRef]
- Hasanvand, A. The role of AMPK-dependent pathways in cellular and molecular mechanisms of metformin: A new perspective for treatment and prevention of diseases. Inflammopharmacology 2022, 30, 775–788. [Google Scholar] [CrossRef]
- Li, Q.; Zhuang, Q.K.; Yang, J.N.; Zhang, Y.Y. Statins excert neuroprotection on cerebral ischemia independent of their lipid-lowering action: The potential molecular mechanisms. Eur. Rev. Med. Pharmacol. Sci. 2014, 18, 1113–1126. [Google Scholar]
- Trapani, L.; Segatto, M.; Pallottini, V. New compounds able to control hepatic cholesterol metabolism: Is it possible to avoid statin treatment in aged people? World J. Hepatol. 2013, 5, 676–684. [Google Scholar] [CrossRef]
- Pfrieger, F.W. Outsourcing in the brain: Do neurons depend on cholesterol delivery by astrocytes? Bioessays 2003, 25, 72–78. [Google Scholar] [CrossRef]
- Mysore, Y.; del Amo, E.M.; Loukovaara, S.; Hagström, M.; Urtti, A.; Kauppinen, A. Author Correction: Statins for the prevention of proliferative vitreoretinopathy: Cellular responses in cultured cells and clinical statin concentrations in the vitreous. Sci. Rep. 2021, 11, 980. [Google Scholar] [CrossRef] [PubMed]
- Vavvas, D.G.; Daniels, A.B.; Kapsala, Z.G.; Goldfarb, J.W.; Ganotakis, E.; Loewenstein, J.I.; Young, L.H.; Gragoudas, E.S.; Eliott, D.; Kim, I.K.; et al. Regression of Some High-risk Features of Age-related Macular Degeneration (AMD) in Patients Receiving Intensive Statin Treatment. eBioMedicine 2016, 5, 198–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loukovaara, S.; Sahanne, S.; Takala, A.; Haukka, J. Statin use and vitreoretinal surgery: Findings from a Finnish population-based cohort study. Acta Ophthalmol. 2018, 96, 442–451. [Google Scholar] [CrossRef] [PubMed]
- Tuuminen, R.; Yegutkin, G.G.; Jalkanen, S.; Loukovaara, S. Simvastatin use associated with low intraocular ADP levels in patients with sight-threatening diabetic retinopathy. Graefes Arch. Clin. Exp. Ophthalmol. 2016, 254, 1643–1644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuuminen, R.; Sahanne, S.; Loukovaara, S. Low intravitreal angiopoietin-2 and VEGF levels in vitrectomized diabetic patients with simvastatin treatment. Acta Ophthalmol. 2014, 92, 675–681. [Google Scholar] [CrossRef]
- Kang, E.Y.; Chen, T.H.; Garg, S.J.; Sun, C.C.; Kang, J.H.; Wu, W.C.; Hung, M.J.; Lai, C.C.; Cherng, W.J.; Hwang, Y.S. Association of Statin Therapy With Prevention of Vision-Threatening Diabetic Retinopathy. JAMA Ophthalmol. 2019, 137, 363–371. [Google Scholar] [CrossRef]
- Yuan, Y.; Wang, W.; Shang, X.; Xiong, R.; Ha, J.; Zhang, L.; Zhu, Z.; He, M. Association between statin use and the risks of glaucoma in Australia: A 10-year cohort study. Br. J. Ophthalmol. 2021; ahead of print. [Google Scholar] [CrossRef]
- Sang, N.; Stiehl, D.P.; Bohensky, J.; Leshchinsky, I.; Srinivas, V.; Caro, J. MAPK signaling up-regulates the activity of hypoxia-inducible factors by its effects on p300. J. Biol. Chem. 2003, 278, 14013–14019. [Google Scholar] [CrossRef] [Green Version]
- Ruan, Y.; Jiang, S.; Gericke, A. Age-Related Macular Degeneration: Role of Oxidative Stress and Blood Vessels. Int. J. Mol. Sci. 2021, 22, 1296. [Google Scholar] [CrossRef]
- Baynes, J.W. Role of oxidative stress in development of complications in diabetes. Diabetes. 1991, 40, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Kowluru, R.A.; Chan, P.S. Oxidative stress and diabetic retinopathy. Exp. Diabetes Res. 2007, 2007, 43603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruschi, M.; Bartolucci, M.; Petretto, A.; Calzia, D.; Caicci, F.; Manni, L.; Traverso, C.E.; Candiano, G.; Panfoli, I. Differential expression of the five redox complexes in the retinal mitochondria or rod outer segment disks is consistent with their different functionality. FASEB Bioadv. 2020, 2, 315–324. [Google Scholar] [CrossRef]
- Calzia, D.; Barabino, S.; Bianchini, P.; Garbarino, G.; Oneto, M.; Caicci, F.; Diaspro, A.; Tacchetti, C.; Manni, L.; Candiani, S.; et al. New findings in ATP supply in rod outer segments: Insights for retinopathies. Biol. Cell. 2013, 105, 345–358. [Google Scholar] [CrossRef] [PubMed]
- Calzia, D.; Oneto, M.; Caicci, F.; Bianchini, P.; Ravera, S.; Bartolucci, M.; Diaspro, A.; Degan, P.; Manni, L.; Traverso, C.E.; et al. Effect of polyphenolic phytochemicals on ectopic oxidative phosphorylation in rod outer segments of bovine retina. Br. J. Pharmacol. 2015, 172, 3890–3903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravera, S.; Caicci, F.; Degan, P.; Maggi, D.; Manni, L.; Puddu, A.; Nicolò, M.; Traverso, C.E.; Panfoli, I. Inhibitory Action of Antidiabetic Drugs on the Free Radical Production by the Rod Outer Segment Ectopic Aerobic Metabolism. Antioxidants 2020, 9, 1133. [Google Scholar] [CrossRef]
- Gu, H.; Huang, Z.; Chen, G.; Zhou, K.; Zhang, Y.; Chen, J.; Xu, J.; Yin, X. Network and pathway-based analyses of genes associated with osteoporosis. Medicine 2020, 99, e19120. [Google Scholar] [CrossRef]
- Hu, Y.; Pan, Z.; Hu, Y.; Zhang, L.; Wang, J. Network and Pathway-Based Analyses of Genes Associated with Parkinson’s Disease. Mol. Neurobiol. 2017, 54, 4452–4465. [Google Scholar] [CrossRef]
- Tao, C.; Sun, J.; Zheng, W.J.; Chen, J.; Xu, H. Colorectal cancer drug target prediction using ontology-based inference and network analysis. Database 2015, 2015, bav015. [Google Scholar] [CrossRef] [Green Version]
- Boyle, E.; Shuai Weng, J.G.; Sherlock, G. GO: TermFinder–open Source Software for Accessing Gene Ontology Information and Finding Significantly Enriched Gene Ontology Terms Associated with a List of Genes. Bioinformatics 2004, 20, 3710–3715. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Bardes, E.E.; Aronow, B.J.; Jegga, A.G. ToppGene Suite for gene list enrichment analysis and candidate gene prioritization. Nucleic Acids Res. 2009, 37, W305–W311. [Google Scholar] [CrossRef] [Green Version]
- Davis, A.P.; Grondin, C.J.; Johnson, R.J.; Sciaky, D.; Wiegers, J.; Wiegers, T.C.; Mattingly, C.J. The Comparative Toxicogenomics Database: Update 2021. Nucleic Acids Res. 2021, 49, D1138–D1143. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, M.; von Mering, C.; Campillos, M.; Jensen, L.J.; Bork, P. STITCH: Interaction networks of chemicals and proteins. Nucleic Acids Res. 2007, 36, D684–D688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Sulakhe, D.; Balasubramanian, S.; Xie, B.; Feng, B.; Taylor, A.; Wang, S.; Berrocal, E.; Dave, U.; Xu, J.; Börnigen, D.; et al. Lynx: A database and knowledge extraction engine for integrative medicine. Nucleic Acids Res. 2013, 42, D1007–D1012. [Google Scholar] [CrossRef]



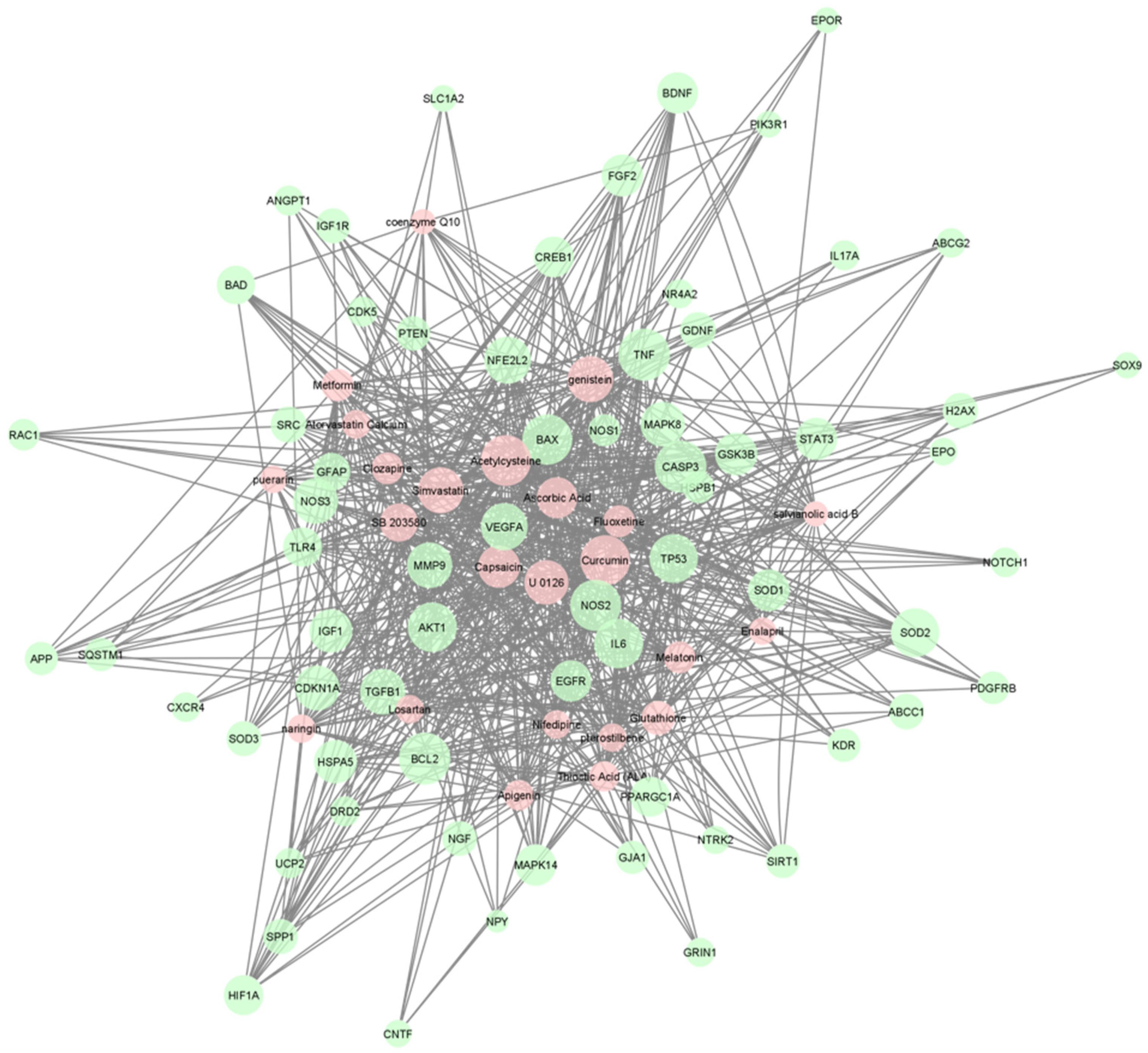{kind=link}
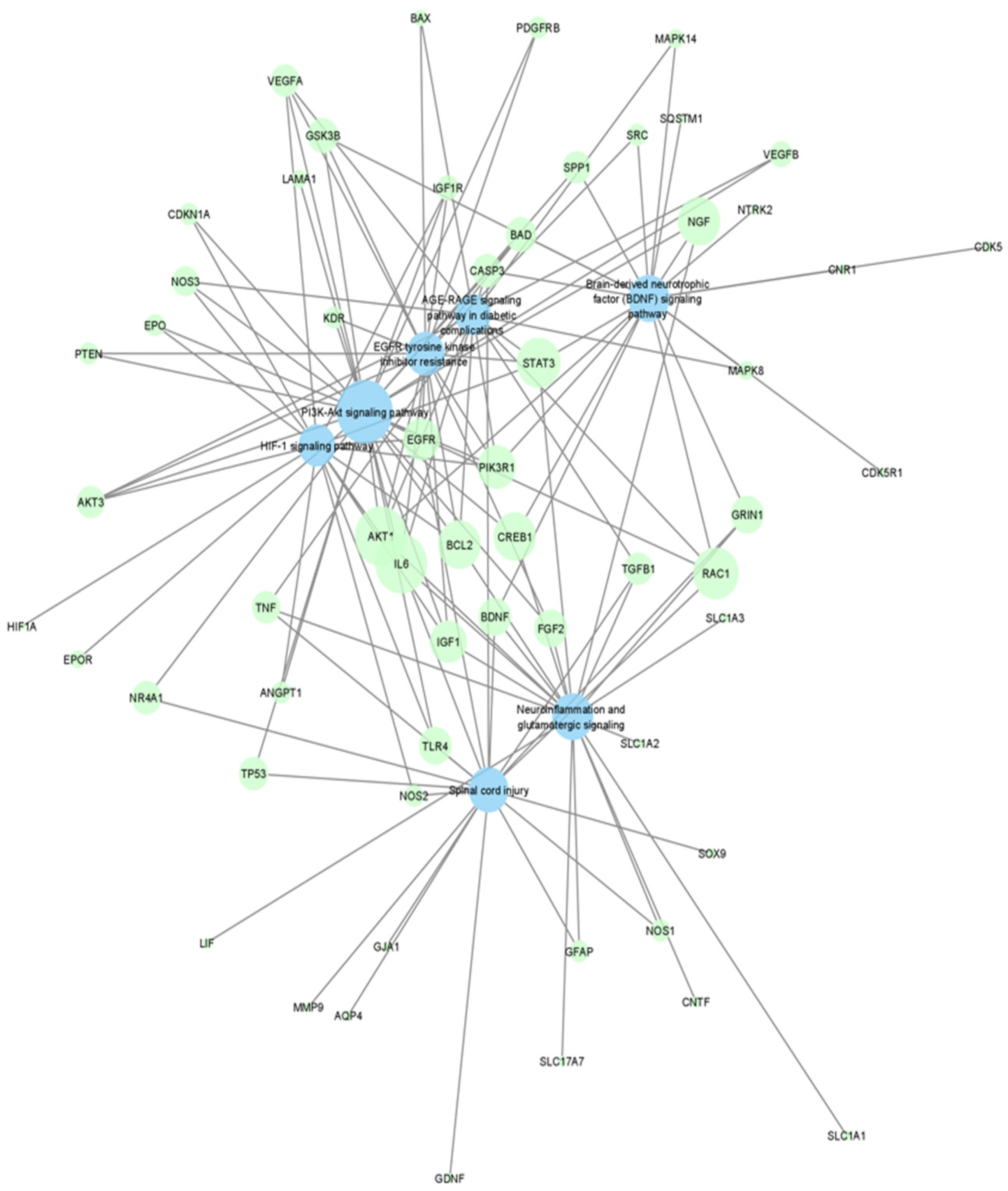{kind=link}
| FILTERED POSITION | UNFILTERED POSITION | NAME | SOURCE | P-VALUE | Q-VALUE FDR B&H | HIT COUNT IN QUERY LIST | HIT COUNT IN GENOME |
|---|---|---|---|---|---|---|---|
| 1 | 2 | U 0126 | CTD | 2.51 × 10−52 | 3.64 × 10−52 | 51 | 444 |
| 2 | 3 | Acetylcysteine | CTD | 3.67 × 10−55 | 3.55 × 10−51 | 59 | 781 |
| 3 | 5 | Simvastatin | CTD | 2.72 × 10−53 | 1.58 × 10−49 | 53 | 581 |
| 4 | 7 | Curcumin | CTD | 1.66 × 10−51 | 6.86 × 10−48 | 58 | 851 |
| 5 | 9 | Capsaicin | CTD | 1.97 × 10−49 | 6.36 × 10−46 | 48 | 488 |
| 6 | 18 | SB 203580 | CTD | 1.18 × 10−44 | 1.90 × 10−41 | 42 | 388 |
| 7 | 20 | Ascorbic Acid | CTD | 2.56 × 10−41 | 3.72 × 10−38 | 46 | 627 |
| 8 | 29 | Genistein | Stitch | 2.20 × 10−38 | 2.21 × 10−35 | 53 | 1117 |
| 9 | 43 | Glutathione | CTD | 3.34 × 10−36 | 2.25 × 10−33 | 35 | 339 |
| 10 | 44 | Thioctic Acid | CTD | 2.51 × 10−35 | 1.65 × 10−32 | 28 | 163 |
| 11 | 45 | Melatonin | CTD | 6.47 × 10−35 | 4.17 × 10−32 | 31 | 243 |
| 12 | 55 | Nifedipine | CTD | 8.36 × 10−33 | 4.41 × 10−30 | 24 | 112 |
| 13 | 61 | Apigenin | CTD | 2.96 × 10−32 | 1.41 × 10−29 | 28 | 207 |
| 14 | 64 | Deferoxamine | CTD | 5.7 × 10−32 | 2.59×10−29 | 27 | 186 |
| 15 | 72 | Pterostilbene | CTD | 4.13 × 10−31 | 1.66 × 10−28 | 24 | 130 |
| 16 | 84 | Salvianolic acid | CTD | 1.24 × 10−31 | 4.28 × 10−28 | 17 | 35 |
| 17 | 83 | Fluoxetine | CTD | 1.18 × 10−30 | 4.12 × 10−28 | 31 | 331 |
| 18 | 86 | Puerarin | CTD | 1.34 × 10−30 | 4.54 × 10−28 | 22 | 98 |
| 19 | 98 | Naringin | CTD | 8.18 × 10−30 | 2.42 × 10−27 | 24 | 146 |
| 20 | 101 | Metformin | CTD | 1.83 × 10−29 | 5.26 × 10−27 | 32 | 400 |
| 21 | 111 | Atorvastatin Calcium | CTD | 9.68 × 10−29 | 2.53 × 10−26 | 25 | 186 |
| 22 | 113 | Losartan | CTD | 1.82 × 10−28 | 4.69 × 10−26 | 24 | 165 |
| 23 | 114 | Clozapine | CTD | 1.92 × 10−28 | 4.89 × 10−26 | 31 | 390 |
| 24 | 127 | Coenzyme Q10 | CTD | 1.10 × 10−27 | 2.51 × 10−25 | 17 | 48 |
| 25 | 128 | Enalapril | CTD | 1.38 × 10−27 | 3.14 × 10−25 | 22 | 131 |
| ID | GO DESCRIPTION | P-VALUE | Q-VALUE FDR B&H | HIT COUNT IN QUERY LIST | HIT COUNT IN GENOME |
|---|---|---|---|---|---|
| GO:1901701 | cellular response to oxygen-containing compound | 6.24 × 10−55 | 3.95 × 10−51 | 78 | 1790 |
| GO:0043067 | regulation of programmed cell death | 1.58 × 10−53 | 4.99 × 10−50 | 79 | 1944 |
| GO:0009628 | response to abiotic stimulus | 1.04 × 10−52 | 1.65 × 10−49 | 77 | 1839 |
| GO:0042981 | regulation of apoptotic process | 1.06 × 10−51 | 1.35 × 10−48 | 77 | 1897 |
| GO:0010243 | response to organonitrogen compound | 4.00 × 10−49 | 4.22 × 10−46 | 71 | 1605 |
| GO:1901214 | regulation of neuron death | 4.75 × 10−47 | 3.76 × 10−44 | 47 | 462 |
| GO:0014070 | response to organic cyclic compound | 7.90 × 10−47 | 5.56 × 10−44 | 69 | 1591 |
| GO:0048666 | neuron development | 2.23 × 10−39 | 6.42 × 10−37 | 64 | 1673 |
| GO:0051094 | positive regulation of developmental process | 4.93 × 10−39 | 1.25 × 10−36 | 64 | 1695 |
| GO:0042327 | positive regulation of phosphorylation | 1.02 × 10−38 | 2.47 × 10−36 | 55 | 1113 |
| GO:0031175 | neuron projection development | 1.18 × 10−37 | 2.67 × 10−35 | 59 | 1424 |
| GO:0070482 | response to oxygen levels | 1.87 × 10−37 | 3.94 × 10−35 | 45 | 647 |
| GO:0031399 | regulation of protein modification process | 3.24 × 10−37 | 6.61 × 10−35 | 66 | 1976 |
| GO:0051247 | positive regulation of protein metabolic process | 4.57 × 10−37 | 9.05 × 10−35 | 64 | 1827 |
| GO:0009611 | response to wounding | 8.65 × 10−37 | 1.66 × 10−34 | 50 | 919 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, E.; Nadeem, U.; Xie, B.; D’Souza, M.; Sulakhe, D.; Skondra, D. Using Computational Drug-Gene Analysis to Identify Novel Therapeutic Candidates for Retinal Neuroprotection. Int. J. Mol. Sci. 2022, 23, 12648. https://doi.org/10.3390/ijms232012648
Xie E, Nadeem U, Xie B, D’Souza M, Sulakhe D, Skondra D. Using Computational Drug-Gene Analysis to Identify Novel Therapeutic Candidates for Retinal Neuroprotection. International Journal of Molecular Sciences. 2022; 23(20):12648. https://doi.org/10.3390/ijms232012648
Chicago/Turabian StyleXie, Edward, Urooba Nadeem, Bingqing Xie, Mark D’Souza, Dinanath Sulakhe, and Dimitra Skondra. 2022. "Using Computational Drug-Gene Analysis to Identify Novel Therapeutic Candidates for Retinal Neuroprotection" International Journal of Molecular Sciences 23, no. 20: 12648. https://doi.org/10.3390/ijms232012648