
Rational Design of a Thermostable 2′-Deoxyribosyltransferase for Nelarabine Production by Prediction of Disulfide Bond Engineering Sites

 ,
,
Abstract
:
1. Introduction
2. Results and Discussion
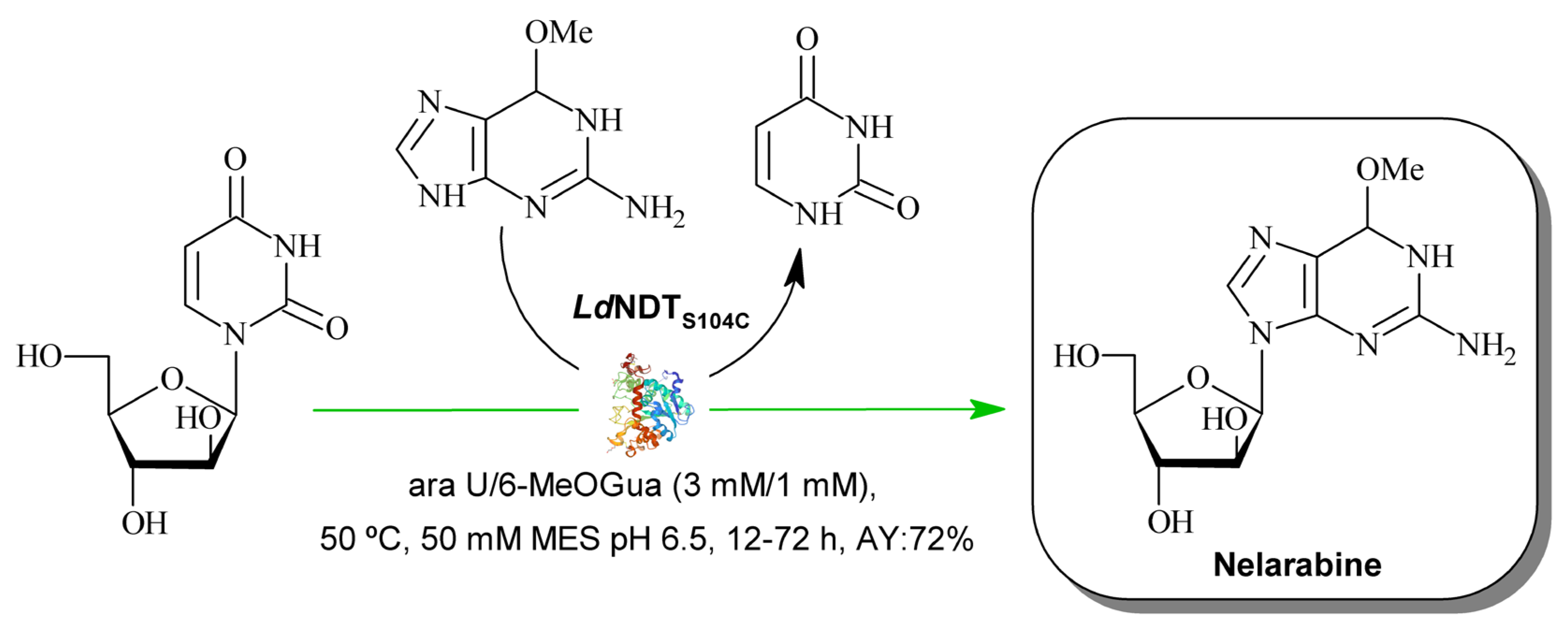
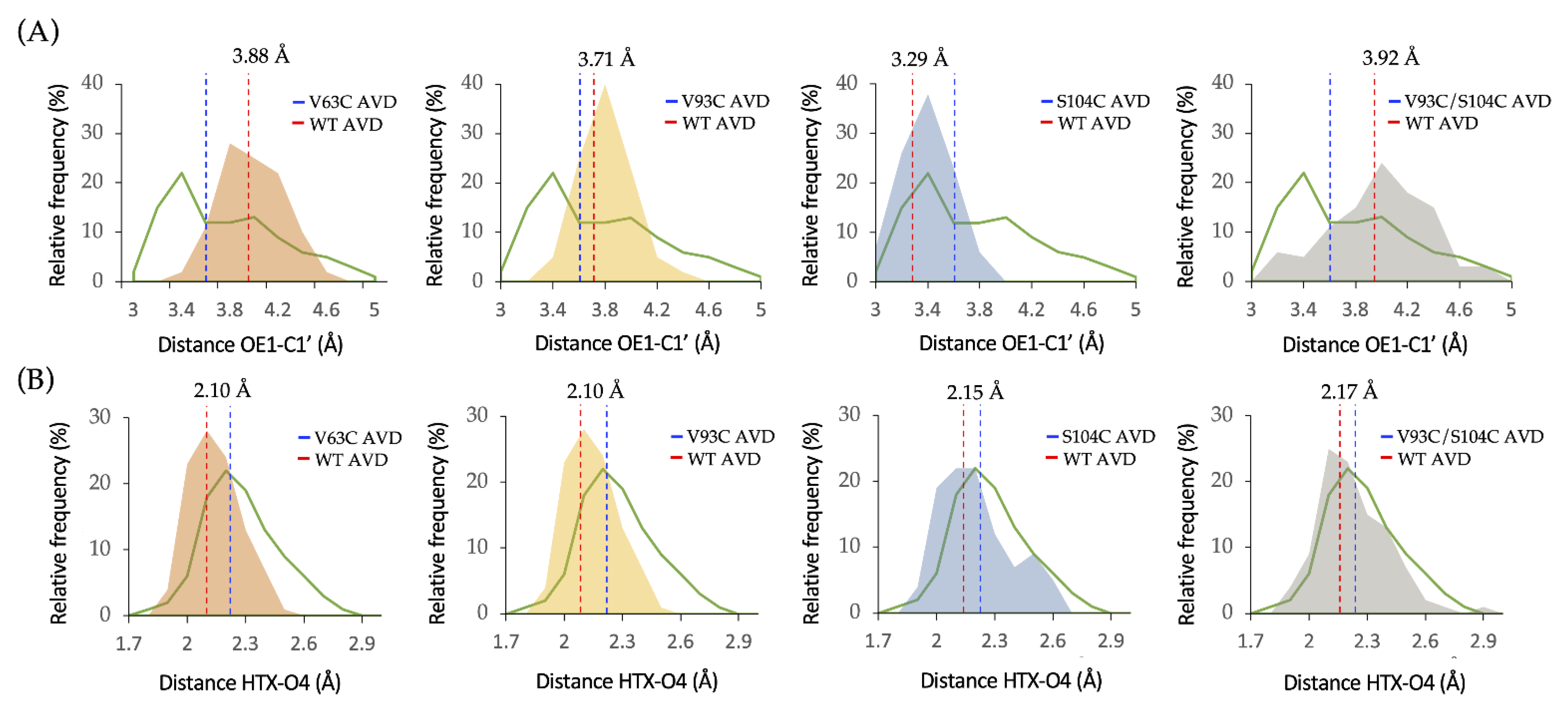
2.1. Computational Design of LdNDT Variants
2.2. Production of LdNDT Variants
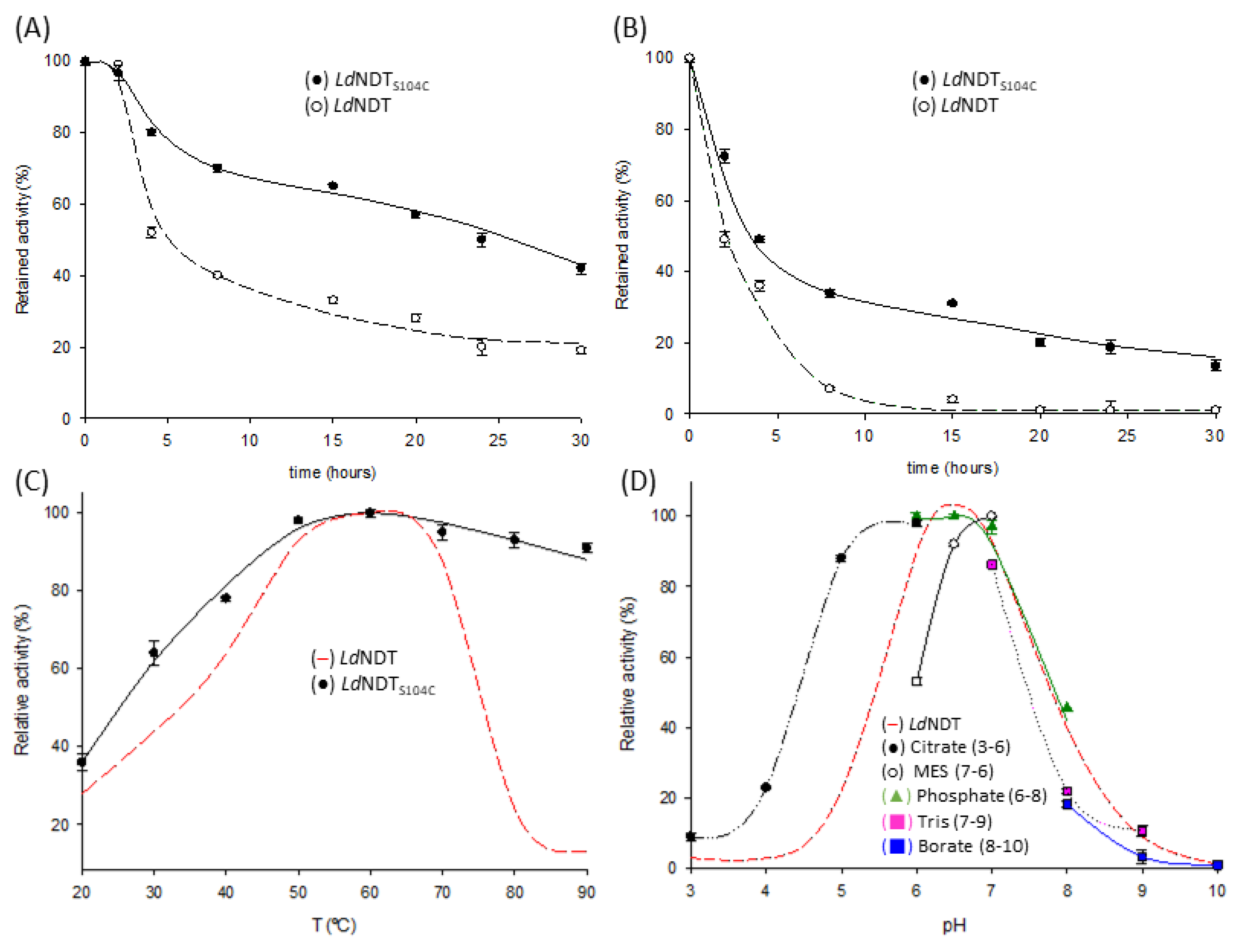
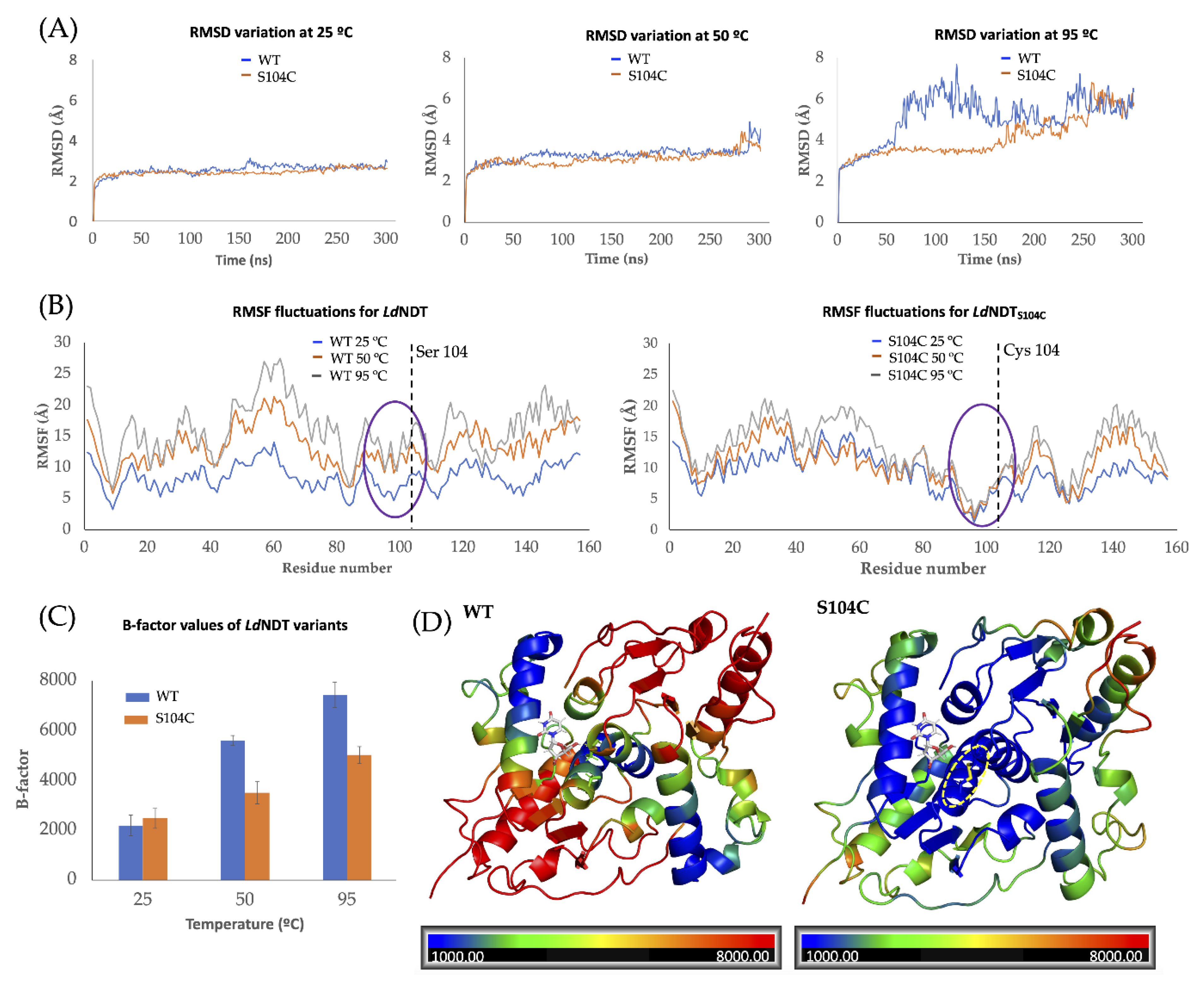
2.3. Thermal Stability
2.4. Biochemical Characterization
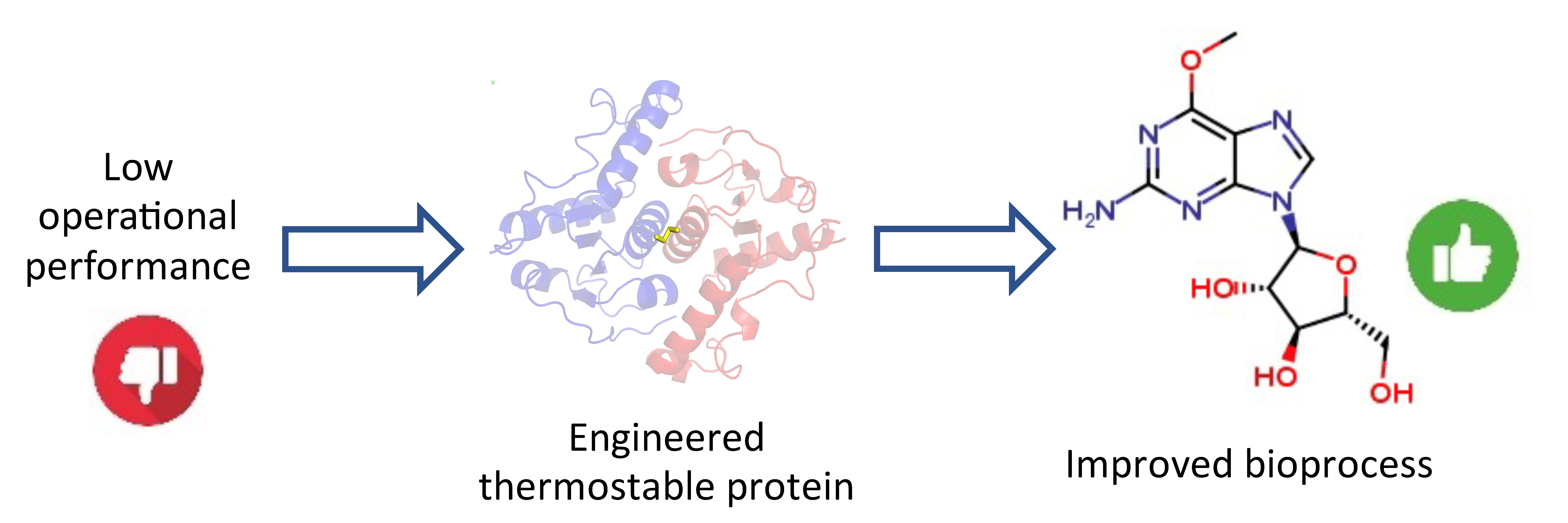
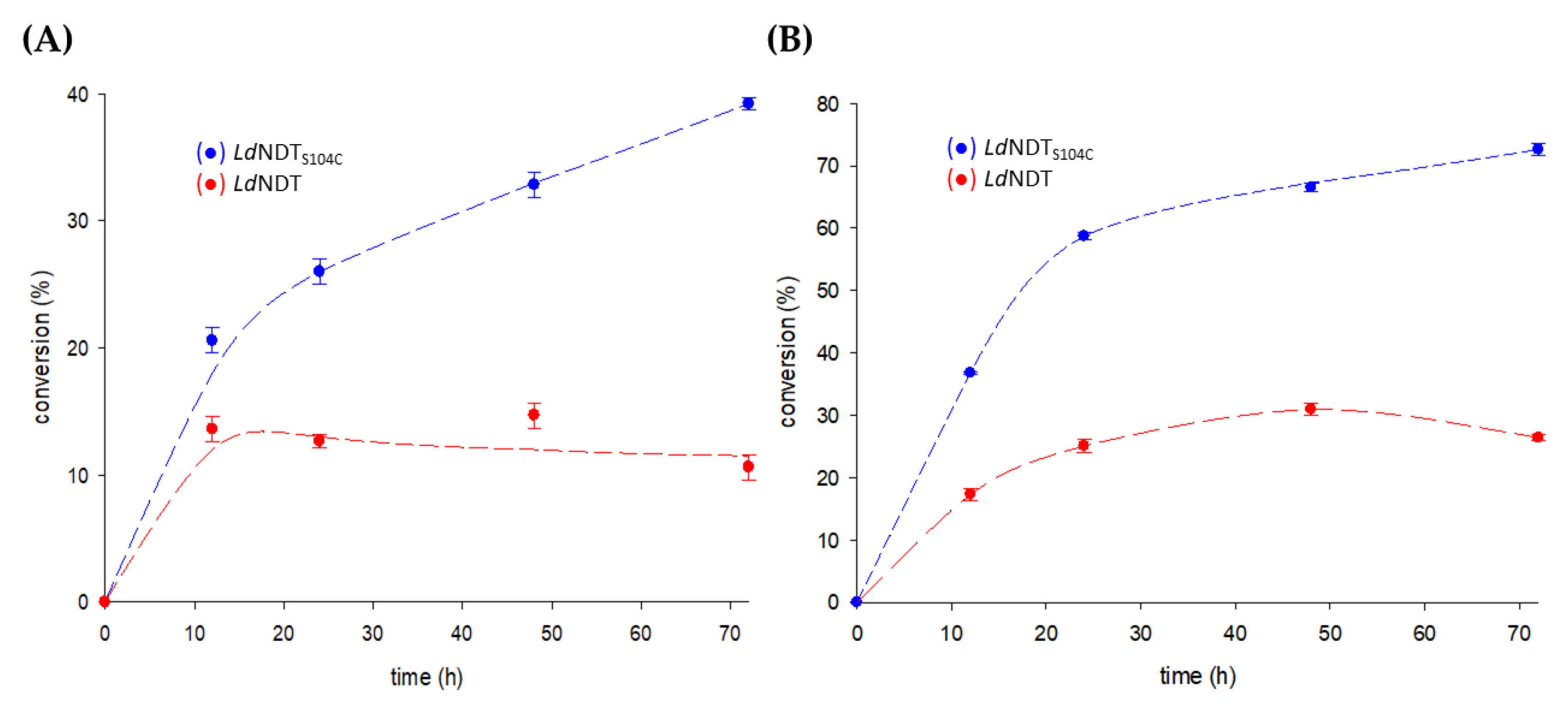
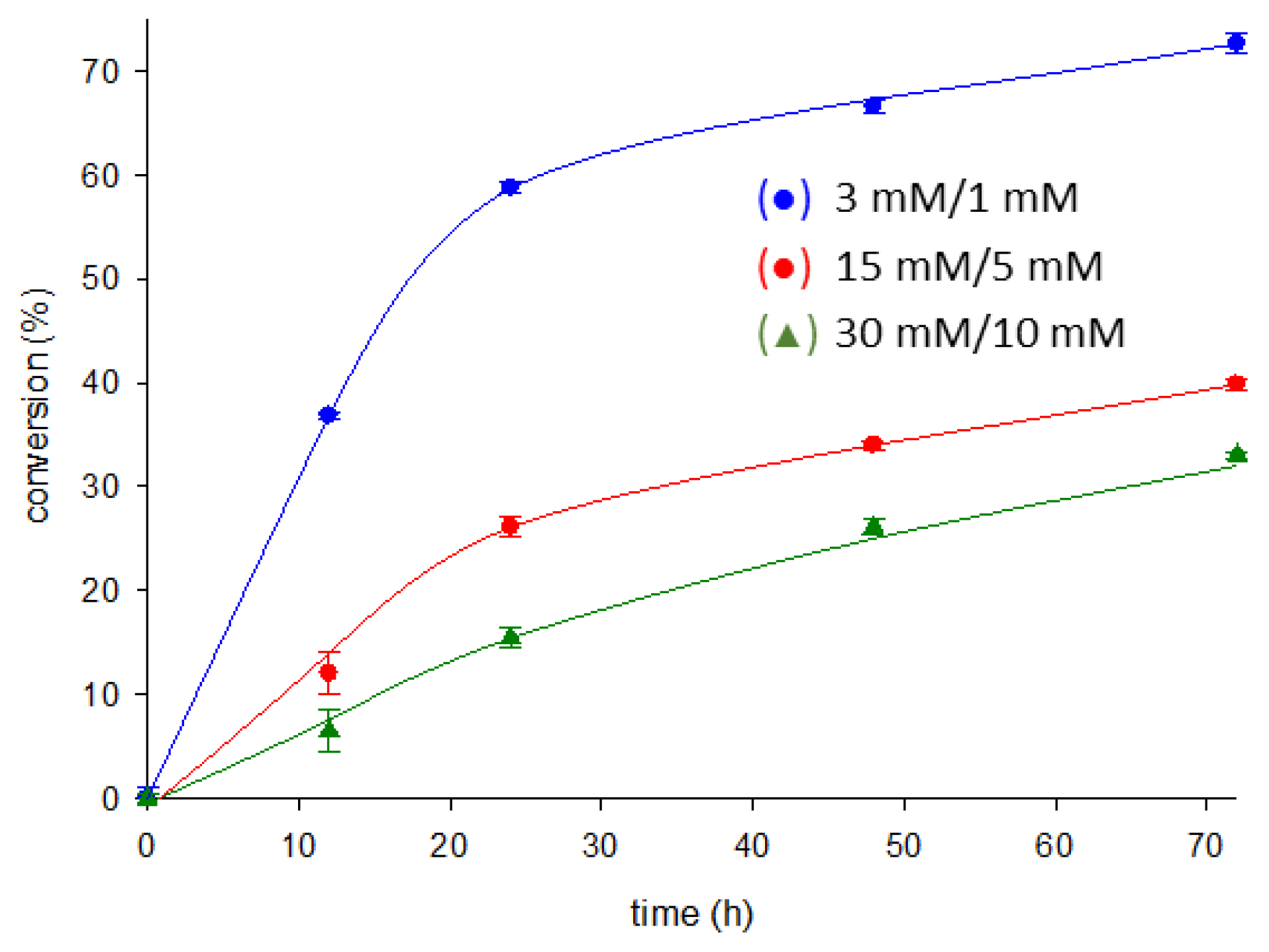
2.5. Enzymatic Synthesis of Nelarabine
3. Materials and Methods
3.1. Materials
3.2. Gene Expression and Protein Purification
3.3. N-2′-Deoxyribosyltransferase Activity Assay
3.4. Thermal Inactivation
3.5. Biochemical Characterization
3.6. Enzymatic Synthesis of Nelarabine
3.7. Molecular Modeling
3.8. QM/MM MD Simulations
3.9. MD Simulations for Thermal Stability Predictions
3.10. Analytical Methods
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jordheim, L.P.; Durantel, D.; Zoulim, F.; Dumontet, C. Advances in the development of nucleoside and nucleotide analogues for cancer and viral diseases. Nat. Rev. Drug Discov. 2013, 12, 447–464. [Google Scholar] [CrossRef] [PubMed]
- Parker, W.B. Enzymology of purine and pyrimidine antimetabolites used in the treatment of cancer. Chem. Rev. 2009, 109, 2880–2893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomson, J.M.; Lamont, I.L. Nucleoside analogues as antibacterial agents. Front. Microbiol. 2019, 10, 952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Lucas, J.; Camarasa, M.J. Enzymatic and Chemical Synthesis of Nucleic acid Derivatives; John Wiley & Sons: Hoboken, NJ, USA, 2019. [Google Scholar]
- Simić, S.; Zukić, E.; Schmermund, L.; Faber, K.; Winkler, C.K.; Kroutil, W. Shortening synthetic routes to small molecule active pharmaceutical ingredients employing biocatalytic methods. Chem. Rev. 2021, 122, 1052–1126. [Google Scholar] [CrossRef]
- Lapponi, M.J.; Rivero, C.W.; Zinni, M.A.; Britos, C.N.; Trelles, J.A. New developments in nucleoside analogues biosynthesis: A review. J. Mol. Catal. B Enzym. 2016, 133, 218–233. [Google Scholar] [CrossRef]
- Rinaldi, F.; Fernández-Lucas, J.; de la Fuente, D.; Zheng, C.; Bavaro, T.; Peters, B.; Massolini, G.; Annunziata, F.; Conti, P.; De la Mata, I.; et al. Immobilized enzyme reactors based on nucleoside phosphorylases and 2′-deoxyribosyltransferase for the in-flow synthesis of pharmaceutically relevant nucleoside analogues. Bioresour. Technol. 2020, 307, 123258. [Google Scholar] [CrossRef]
- Kamel, S.; Weiß, M.; Klare, H.F.; Mikhailopulo, I.A.; Neubauer, P.; Wagner, A. Chemo-enzymatic synthesis of α-D-pentofuranose-1-phosphates using thermostable pyrimidine nucleoside phosphorylases. Mol. Catal. 2018, 458, 52–59. [Google Scholar] [CrossRef]
- Del Arco, J.; Acosta, J.; Fernández-Lucas, J. New trends in the biocatalytic production of nucleosidic active pharmaceutical ingredients using 2′-deoxyribosyltransferases. Biotechnol. Adv. 2021, 51, 107701. [Google Scholar] [CrossRef]
- Fresco-Taboada, A.; De La Mata, I.; Arroyo, M.; Fernández-Lucas, J. New insights on nucleoside 2′-deoxyribosyltransferases: A versatile biocatalyst for one-pot one-step synthesis of nucleoside analogs. Appl. Microbiol. Biotechnol. 2013, 97, 3773–3785. [Google Scholar] [CrossRef]
- Acosta, J.; Del Arco, J.; Martinez-Pascual, S.; Clemente-Suárez, V.J.; Fernández-Lucas, J. One-pot multi-enzymatic production of purine derivatives with application in pharmaceutical and food industry. Catalysts 2018, 8, 9. [Google Scholar] [CrossRef]
- Del Arco, J.; Pérez, E.; Naitow, H.; Kunishima, N.; Fernández-Lucas, J. Structural and functional characterization of thermostable biocatalysts for the synthesis of 6-aminopurine nucleoside-5’-monophospate analogues. Bioresour. Technol. 2019, 276, 244–252. [Google Scholar] [CrossRef]
- Mikhailopulo, I.A.; Miroshnikov, A.I. Biologically important nucleosides: Modern trends in biotechnology and application. Mendeleev Commun. 2011, 21, 57–69. [Google Scholar] [CrossRef]
- Frisch, J.; Maršić, T.; Loderer, C. A novel one-pot enzyme cascade for the biosynthesis of cladribine triphosphate. Biomolecules 2021, 11, 346. [Google Scholar] [CrossRef]
- Ding, Y.; Ou, L.; Ding, Q. Enzymatic synthesis of nucleoside triphosphates and deoxynucleoside triphosphates by surface-displayed kinases. Appl. Biochem. Biotechnol. 2020, 190, 1271–1288. [Google Scholar] [CrossRef]
- Li, J.; Yu, L.; Li, J.; Xie, L.; Zhang, R.; Wang, H. Establishment of a high throughput-screening system for nucleoside deoxyribosyltransferase II mutant enzymes with altered substrate specificity. J. Biosci. Bioeng. 2019, 128, 22–27. [Google Scholar] [CrossRef]
- Slagman, S.; Fessner, W.D. Biocatalytic routes to anti-viral agents and their synthetic intermediates. Chem. Soc. Rev. 2021, 50, 1968–2009. [Google Scholar] [CrossRef]
- Lewkowicz, E.S.; Iribarren, A.M. Whole cell biocatalysts for the preparation of nucleosides and their derivatives. Curr. Pharm. Des. 2017, 23, 6851–6878. [Google Scholar] [CrossRef]
- Gupta, M.; Nair, V. Adenosine deaminase in nucleoside synthesis. A review. Collect. Czechoslov. Chem. Commun. 2006, 71, 769–787. [Google Scholar] [CrossRef]
- Woodley, J.M. Accelerating the implementation of biocatalysis in industry. Appl. Microbiol. Biotechnol. 2019, 103, 4733–4739. [Google Scholar] [CrossRef]
- Wu, S.; Snajdrova, R.; Moore, J.C.; Baldenius, K.; Bornscheuer, U.T. Biocatalysis: Enzymatic synthesis for industrial applications. Angew. Chem. Int. Ed. 2021, 60, 88–119. [Google Scholar] [CrossRef]
- Acosta, J.; Nguyen, K.; Spitale, R.C.; Fernández-Lucas, J. Taylor-made production of pyrimidine nucleoside-5′-monophosphate analogues by highly stabilized mutant uracil phosphoribosyltransferase from Toxoplasma gondii. Bioresour. Technol. 2021, 339, 125649. [Google Scholar] [CrossRef]
- Del Arco, J.; Alcántara, A.R.; Fernández-Lafuente, R.; Fernández-Lucas, J. Magnetic micro-macro biocatalysts applied to industrial bioprocesses. Bioresour. Technol. 2021, 322, 124547. [Google Scholar] [CrossRef]
- Acosta, J.; Del Arco, J.; Pisabarro, V.; Gago, F.; Fernández-Lucas, J. N-ribosyltransferase from Archaeoglobus veneficus: A novel halotolerant and thermostable biocatalyst for the synthesis of purine ribonucleoside analogs. Front. Bioeng. Biotechnol. 2020, 8, 593. [Google Scholar] [CrossRef]
- Del Arco, J.; Fernández-Lucas, J. Purine and pyrimidine salvage pathway in thermophiles: A valuable source of biocatalysts for the industrial production of nucleic acid derivatives. Appl. Microbiol. Biotechnol. 2018, 102, 7805–7820. [Google Scholar] [CrossRef]
- Bosch, S.; Sanchez-Freire, E.; Del Pozo, M.L.; Česnik, M.; Quesada, J.; Mate, D.M.; Hernández, K.; Qi, Y.; Clapés, P.; Vasić-Rački, D.; et al. Thermostability engineering of a class II pyruvate aldolase from Escherichia coli by in vivo folding interference. ACS Sustain. Chem. Eng. 2021, 9, 5430–5436. [Google Scholar] [CrossRef]
- Mateljak, I.; Alcalde, M. Engineering a highly thermostable high-redox potential laccase. ACS Sustain. Chem. Eng. 2021, 9, 9632–9637. [Google Scholar] [CrossRef]
- Planas-Iglesias, J.; Marques, S.M.; Pinto, G.P.; Musil, M.; Stourac, J.; Damborsky, J.; Bednar, D. Computational design of enzymes for biotechnological applications. Biotechnol. Adv. 2021, 47, 107696. [Google Scholar] [CrossRef] [PubMed]
- Koudelakova, T.; Chaloupkova, R.; Brezovsky, J.; Prokop, Z.; Sebestova, E.; Hesseler, M.; Khabiri, M.; Plevaka, M.; Kulik, D.; Smatanova, I.K.; et al. Engineering enzyme stability and resistance to an organic cosolvent by modification of residues in the access tunnel. Angew. Chem. Int. Ed. Engl. 2013, 125, 2013–2017. [Google Scholar] [CrossRef]
- Sumbalova, L.; Stourac, J.; Martinek, T.; Bednar, D.; Damborsky, J. HotSpot Wizard 3.0: Web server for automated design of mutations and smart libraries based on sequence input information. Nucleic Acids Res. 2018, 46, W356–W362. [Google Scholar] [CrossRef] [PubMed]
- Suplatov, D.; Timonina, D.; Sharapova, Y.; Švedas, V. Yosshi: A web-server for disulfide engineering by bioinformatic analysis of diverse protein families. Nucleic Acids Res. 2019, 47, W308–W314. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.H.; Chan, C.H.; Chen, B.J.; Kao, C.Y.; Liu, H.L.; Hsu, J.P. Bioinformatics approaches for disulfide connectivity prediction. Curr. Protein Pept. Sci. 2007, 8, 243–260. [Google Scholar] [CrossRef]
- Zeiske, T.; Stafford, K.A.; Palmer, A.G., III. Thermostability of enzymes from molecular dynamics simulations. J. Chem. Theory Comput. 2016, 12, 2489–9237. [Google Scholar] [CrossRef] [Green Version]
- Reetz, M.T.; Carballeira, J.D.; Vogel, A. Iterative saturation mutagenesis on the basis of B factors as a strategy for increasing protein thermostability. Angew. Chem. 2006, 45, 7745–7775. [Google Scholar] [CrossRef]
- Krüger, D.M.; Rathi, P.C.; Pfleger, C.; Gohlke, H. CNA web server: Rigidity theory-based thermal unfolding simulations of proteins for linking structure, (thermo-)stability, and function. Nucleic Acids Res. 2013, 41, W340–W348. [Google Scholar] [CrossRef]
- Buß, O.; Muller, D.; Jager, S.; Rudat, J.; Rabe, K.S. Improvement in the thermostability of a β-amino acid converting ω-transaminase by using FoldX. ChemBioChem 2018, 19, 379–387. [Google Scholar] [CrossRef]
- Kaminski, P.A.; Dacher, P.; Dugué, L.; Pochet, S. In vivo reshaping the catalytic site of nucleoside 2′- deoxyribosyltransferase for dideoxy- and didehydronucleosides via a single amino acid substitution. J. Biol. Chem. 2008, 283, 20053–20059. [Google Scholar] [CrossRef] [Green Version]
- Fresco-Taboada, A.; Serra, I.; Arroyo, M.; Fernández-Lucas, J.; de la Mata, I.; Terreni, M. Development of an immobilized biocatalyst based on Bacillus psychrosaccharolyticus NDT for the preparative synthesis of trifluridine and decytabine. Catal. Today 2016, 259, 197–204. [Google Scholar] [CrossRef]
- Pérez, E.; Sánchez-Murcia, P.A.; Jordaan, J.; Blanco, M.D.; Mancheño, J.M.; Gago, F.; Fernández-Lucas, J. Enzymatic synthesis of therapeutic nucleosides using a highly versatile purine nucleoside 2′-deoxyribosyltransferase from Trypanosoma brucei. ChemCatChem 2018, 10, 4406–4416. [Google Scholar] [CrossRef]
- Del Arco, J.; Mills, A.; Gago, F.; Fernández-Lucas, J. Structure-guided tuning of a selectivity switch towards ribonucleosides in Trypanosoma brucei purine nucleoside 2′-deoxyribosyltransferase. Chembiochem 2019, 20, 2996–3000. [Google Scholar] [CrossRef]
- Fernández-Lucas, J.; Acebrón, I.; Wu, R.Y.; Alfaro, Y.; Acosta, J.; Kaminski, P.A.; Arroyo, M.; Joachimiak, A.; Nocek, B.P.; De la Mata, I.; et al. Biochemical and structural studies of two tetrameric nucleoside 2′-deoxyribosyltransferases from psychrophilic and mesophilic bacteria: Insights into cold-adaptation. Int. J. Biol. Macromol. 2021, 192, 138–150. [Google Scholar] [CrossRef]
- Del Arco, J.; Jordaan, J.; Moral-Dardé, V.; Fernández-Lucas, J. Sustainable production of nucleoside analogues by a high-efficient purine 2′-deoxyribosyltransferase immobilized onto Ni2+ chelate magnetic microparticles. Bioresour. Technol. 2019, 289, 121772. [Google Scholar] [CrossRef]
- Rivero, C.W.; García, N.S.; Fernández-Lucas, J.; Betancor, L.; Romanelli, G.P.; Trelles, J. Green production of cladribine by using immobilized 2′-deoxyribosyltransferase from Lactobacillus delbrueckii stabilized through a double covalent/entrapment technology. Biomolecules 2021, 11, 657. [Google Scholar] [CrossRef] [PubMed]
- Kaminski, P.A.; Labesse, G. Phosphodeoxyribosyltransferases, designed enzymes for deoxyribonucleotides synthesis. J. Biol. Chem. 2013, 288, 6534–6541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vichier-Guerre, S.; Dugué, L.; Bonhomme, F.; Pochet, S. Expedient and generic synthesis of imidazole nucleosides by enzymatic transglycosylation. Org. Biomol. Chem. 2016, 14, 3638–3653. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, S.R.; Cook, W.J.; Short, S.A.; Ealick, S.E. Crystal Structures of Nucleoside 2-deoxyribosyltransferase in native and ligand-bound forms reveal architecture of the active site. Structure 1996, 4, 97–107. [Google Scholar] [CrossRef] [Green Version]
- Acosta, J.; Pérez, E.; Sánchez-Murcia, P.A.; Fillat, C.; Fernández-Lucas, J. Molecular basis of NDT-mediated activation of nucleoside-based prodrugs and application in suicide gene therapy. Biomolecules 2021, 11, 120. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Dong, X.; Li, X.; Liu, Z.; Liu, H. Prediction of disulfide bond engineering sites using a machine learning method. Sci. Rep. 2020, 10, 10330. [Google Scholar] [CrossRef]
- Liu, T.; Wang, Y.; Luo, X.; Li, J.; Reed, S.A.; Xiao, H.; Young, T.S.; Schultz, P.G. Enhancing protein stability with extended disulfide bonds. PNAS 2016, 113, 5910–5915. [Google Scholar] [CrossRef] [Green Version]
- Qin, M.; Wang, W.; Thirumalai, D. Protein folding guides disulfide bond formation. PNAS 2015, 112, 11241–11246. [Google Scholar] [CrossRef] [Green Version]
- Fresco-Taboada, A.; Fernández-Lucas, J.; Acebal, C.; Arroyo, M.; Ramón, F.; De la Mata, I.; Mancheño, J.M. 2′-Deoxyribosyltransferase from Bacillus psychrosaccharolyticus: A mesophilic-like biocatalyst for the synthesis of modified nucleosides from a psychrotolerant bacterium. Catalysts 2018, 8, 8. [Google Scholar] [CrossRef]
- Okafor, C.D.; Pathak, M.C.; Fagan, C.E.; Bauer, N.C.; Cole, M.F.; Gaucher, E.A.; Ortlund, E.A. Structural and dynamics comparison of thermostability in ancient, modern, and consensus elongation factor Tus. Structure 2018, 26, 118–129. [Google Scholar] [CrossRef] [Green Version]
- Acosta, J.; Del Arco, J.; Del Pozo, M.L.; Herrera-Tapias, B.; Clemente-Suárez, V.J.; Berenguer, J.; Hidalgo, A.; Fernández-Lucas, J. Hypoxanthine-guanine phosphoribosyltransferase/adenylate kinase from Zobellia galactanivorans: A bifunctional catalyst for the synthesis of nucleoside-5′-mono-, di-and triphosphates. Front. Bioeng. Biotechnol. 2020, 24, 677. [Google Scholar] [CrossRef]
- Zhang, D.; Lazim, R. Application of conventional molecular dynamics simulation in evaluating the stability of apomyoglobin in urea solution. Sci. Rep. 2017, 7, 44651. [Google Scholar] [CrossRef] [Green Version]
- Shen, C.; Liu, J.; Ouyang, W.; Ding, H.; Bai, J.; Xiao, Q. Practical synthesis of fludarabine and nelarabine. Synthesis 2020, 52, 417–423. [Google Scholar] [CrossRef]
- Konstantinova, I.D.; Antonov, K.V.; Fateev, I.V.; Miroshnikov, A.I.; Stepchenko, V.A.; Baranovsky, A.V.; Mikhailopulo, I.A. A chemo-enzymatic synthesis of β-D-arabinofuranosyl purine nucleosides. Synthesis 2011, 10, 1555–1560. [Google Scholar]
- Fernández-Lucas, J.; Acebal, C.; Sinisterra, J.V.; Arroyo, M.; de la Mata, I. Lactobacillus reuteri 2′-deoxyribosyltransferase, a novel biocatalyst for tailoring of nucleosides. Appl. Environm. Microbiol. 2010, 76, 1462–1470. [Google Scholar] [CrossRef] [Green Version]
- Crespo, N.; Sánchez-Murcia, P.A.; Gago, F.; Cejudo-Sanches, J.; Galmes, M.A.; Fernández-Lucas, J.; Mancheño, J.M. 2′-Deoxyribosyltransferase from Leishmania mexicana, an efficient biocatalyst for one-pot, one-step synthesis of nucleosides from poorly soluble purine bases. Appl. Microbiol. Biotechnol. 2017, 101, 7187–7200. [Google Scholar] [CrossRef]
- Del Arco, J.; Sánchez-Murcia, P.A.; Mancheño, J.M.; Gago, F.; Fernández-Lucas, J. Characterization of an atypical, thermostable, organic solvent-and acid-tolerant 2′-deoxyribosyltransferase from Chroococcidiopsis thermalis. Appl. Microbiol. Biotechnol. 2018, 102, 6947–6957. [Google Scholar] [CrossRef]
- Fernández-Lucas, J.; Harris, R.; Mata-Casar, I.; Heras, A.; de la Mata, I.; Arroyo, M. Magnetic chitosan beads for covalent immobilization of nucleoside 2′-deoxyribosyltransferase: Application in nucleoside analogues synthesis. J. Ind. Microbiol. Biotechnol. 2013, 40, 955–966. [Google Scholar] [CrossRef]
- Anandakrishnan, R.; Aguilar, B.; Onufriev, A.V. H++ 3.0: Automating pK prediction and the preparation of biomolecular structures for atomistic molecular modeling and simulations. Nucleic Acids Res. 2012, 40, 537–541. [Google Scholar] [CrossRef] [Green Version]
- Case, D.A.; Betz, R.M.; Cerutti, D.S.; Cheatham, T.E.; Darden, T.A.; Duke, R.E.; Giese, T.J.; Gohlke, H.; Goetz, A.W.; Homeyer, N.; et al. AMBER 2016; University of California: San Francisco, CA, USA, 2016. [Google Scholar]
- Jorgensen, W.L.; Chandrasekhar, J.; Madura, J.D.; Impey, R.W.; Klein, M.L. Comparison of simple potential functions for simulating liquid water. J. Chem. Phys. 1983, 79, 926–935. [Google Scholar] [CrossRef]
- Berendsen, H.J.C.; Postma, J.P.M.; van Gunsteren, W.F.; DiNola, A.; Haak, J.R. Molecular dynamics with coupling to an external bath. J. Chem. Phys. 1984, 81, 3684–3690. [Google Scholar] [CrossRef] [Green Version]
- Del Arco, J.; Perona, A.; González, L.; Fernández-Lucas, J.; Gago, F.; Sánchez-Murcia, P.A. Reaction mechanism of nucleoside 2′-deoxyribosyltransferases: Free-energy landscape supports an oxocarbenium ion as the reaction intermediate. Org. Biomol. Chem. 2019, 17, 7891–7899. [Google Scholar] [CrossRef] [PubMed]
- DeLano, W.L. The PyMOL Molecular Graphics System. 2002. Available online: http://www.pymol.org (accessed on 2 August 2022).
- Gaus, M.; Cui, Q.; Elstner, M. DFTB3: Extension of the self-consistent-charge density-functional tight-binding method (SCC-DFTB). J. Chem. Theory Comput. 2012, 7, 931–948. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Disulfide Bridge Localization | Specific Activity (IU mg−1enz) | Relative Activity (%) |
|---|---|---|---|
| LdNDT | - | 191.83 ± 1.3 | 96 |
| LdNDTV63C | Interdimer | 117.01 ± 3.0 | 59 |
| LdNDTV93C | Intradimer | 176.37 ± 2.4 | 88 |
| LdNDTS104C | Intradimer | 192.03 ± 4.0 | 100 |
| LdNDTV93C/S104C | Intradimer | 106.88 ± 1.8 | 54 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cruz, G.; Acosta, J.; Mancheño, J.M.; Del Arco, J.; Fernández-Lucas, J. Rational Design of a Thermostable 2′-Deoxyribosyltransferase for Nelarabine Production by Prediction of Disulfide Bond Engineering Sites. Int. J. Mol. Sci. 2022, 23, 11806. https://doi.org/10.3390/ijms231911806
Cruz G, Acosta J, Mancheño JM, Del Arco J, Fernández-Lucas J. Rational Design of a Thermostable 2′-Deoxyribosyltransferase for Nelarabine Production by Prediction of Disulfide Bond Engineering Sites. International Journal of Molecular Sciences. 2022; 23(19):11806. https://doi.org/10.3390/ijms231911806
Chicago/Turabian StyleCruz, Guillermo, Javier Acosta, Jose Miguel Mancheño, Jon Del Arco, and Jesús Fernández-Lucas. 2022. "Rational Design of a Thermostable 2′-Deoxyribosyltransferase for Nelarabine Production by Prediction of Disulfide Bond Engineering Sites" International Journal of Molecular Sciences 23, no. 19: 11806. https://doi.org/10.3390/ijms231911806