
A Proteomic Platform Unveils the Brain Glycogen Phosphorylase as a Potential Therapeutic Target for Glioblastoma Multiforme

 , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
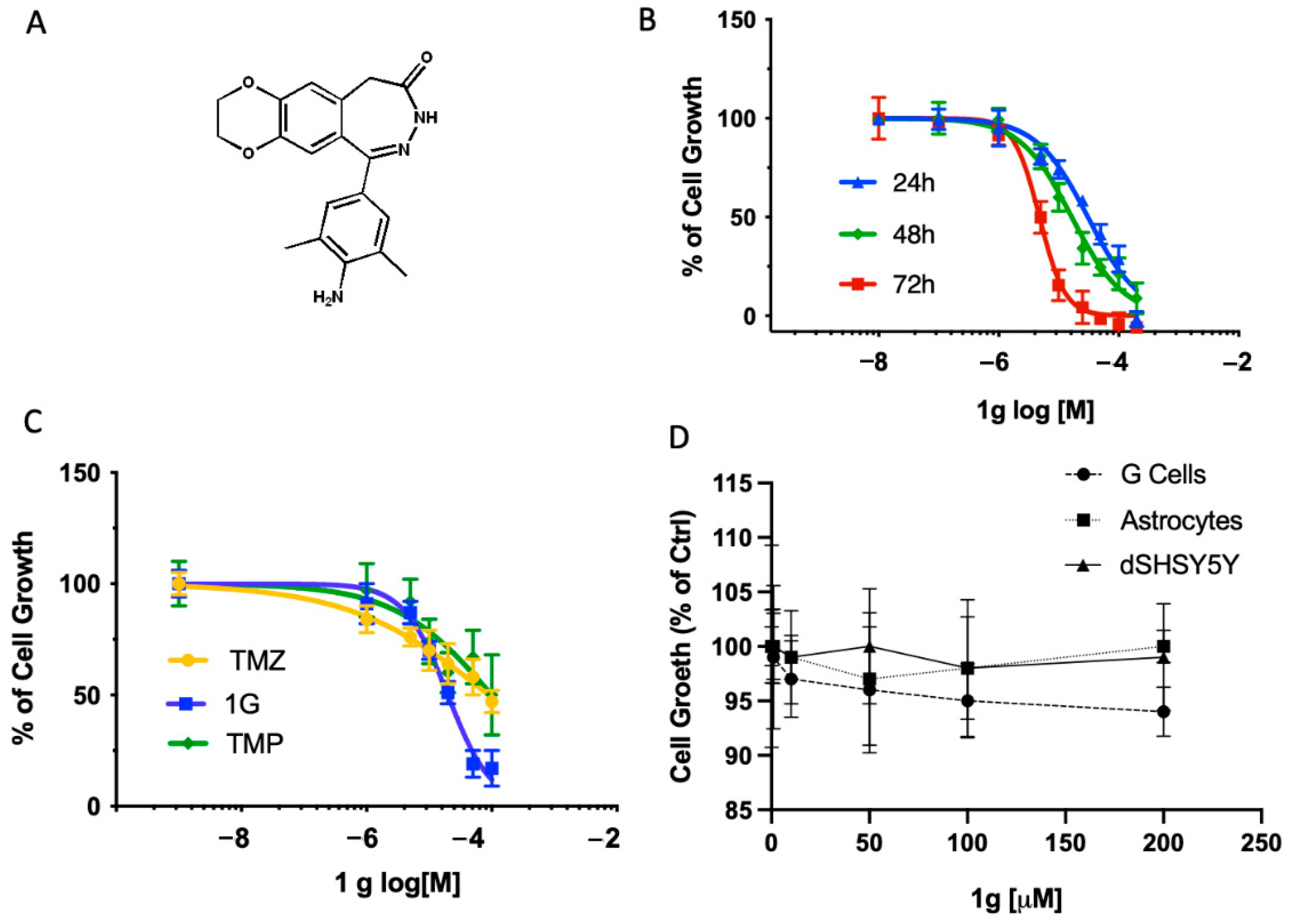
2.1. 1g Permeation by PAMPA Assays
2.2. Effects of 1g on Cell Growth in U87MG, Neuronal Differentiated SHSY5Y and Primary Neuronal Culture
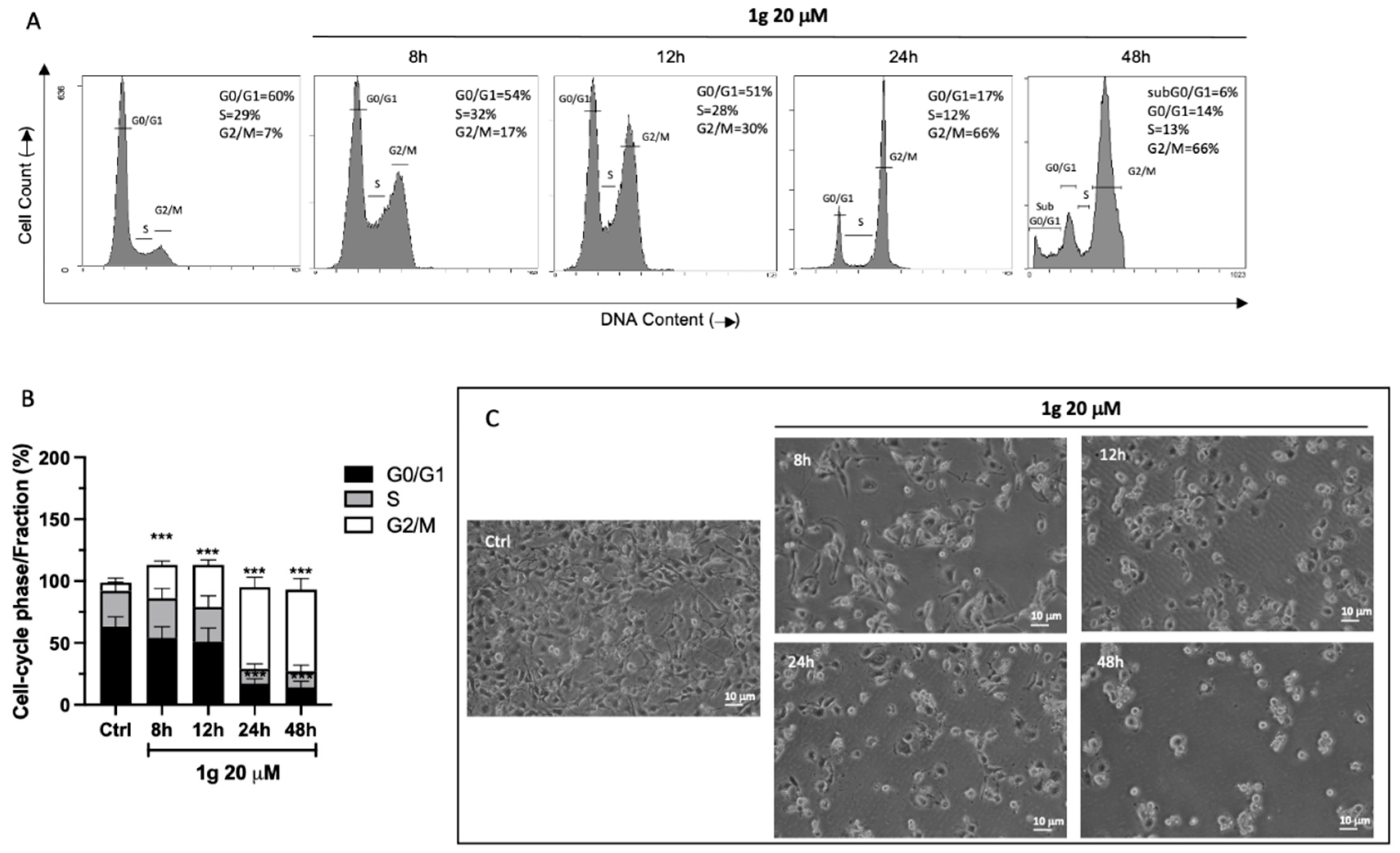
2.3. Cell Cycle Analysis
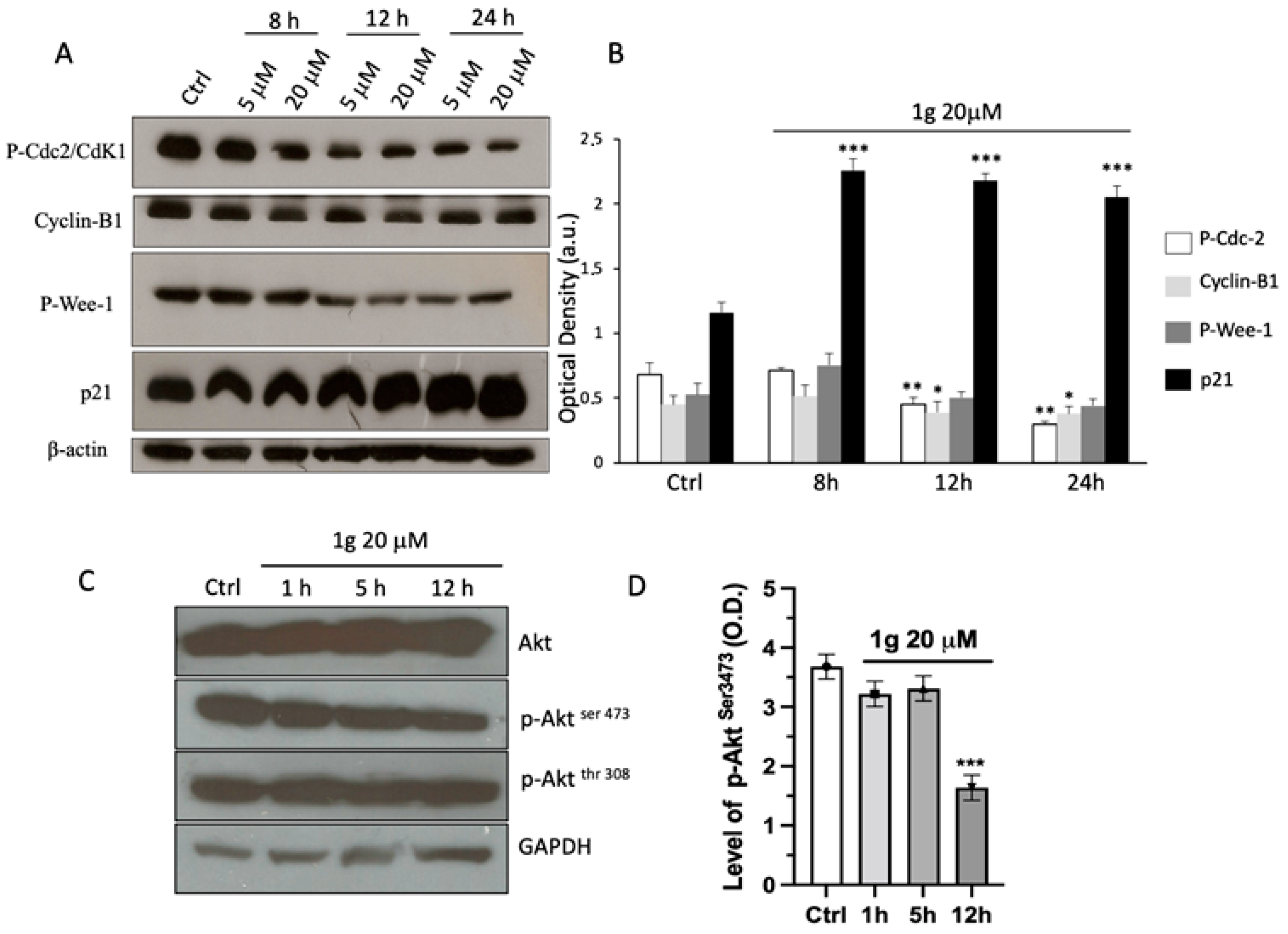
2.4. Protein Expression Evaluation
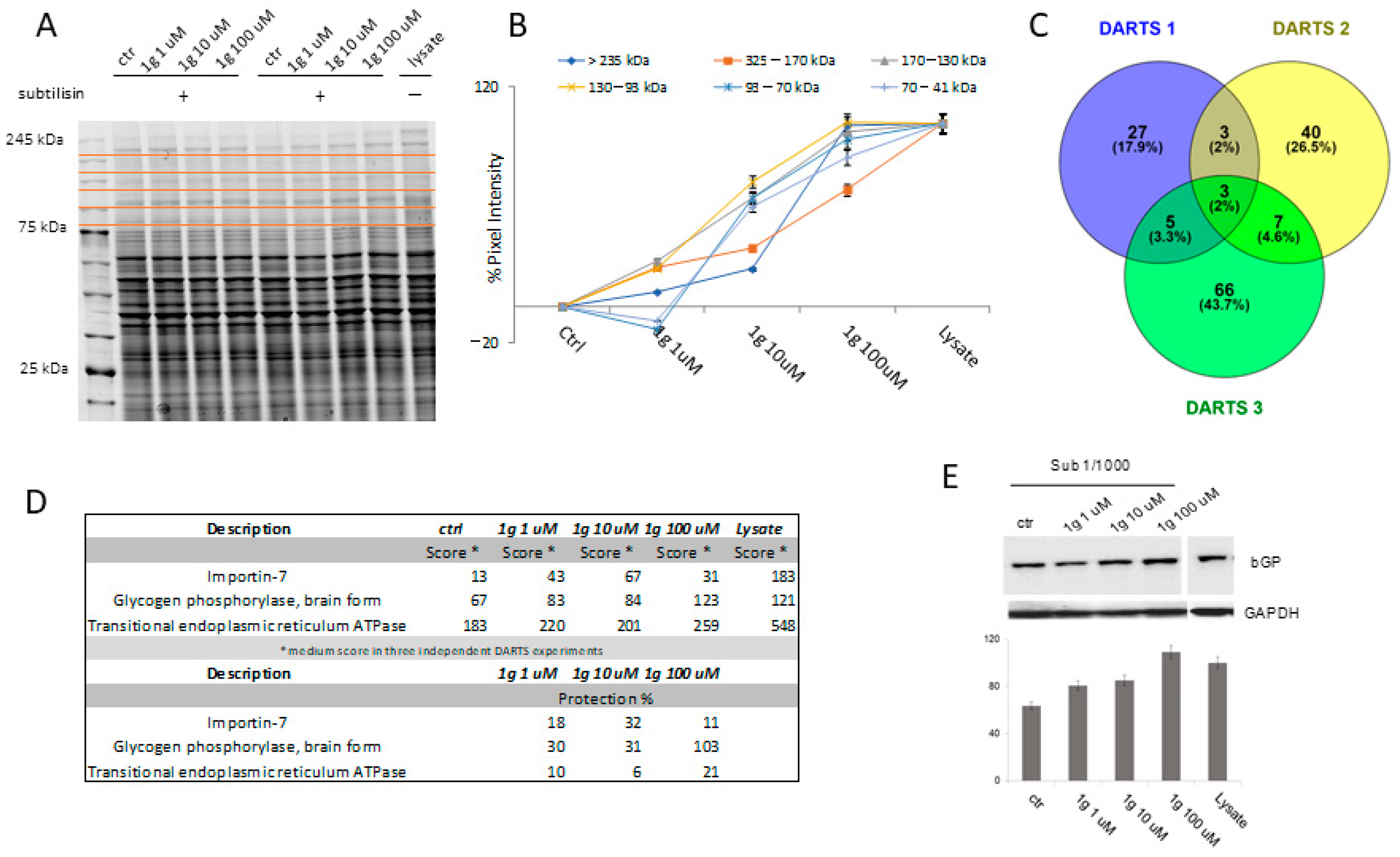
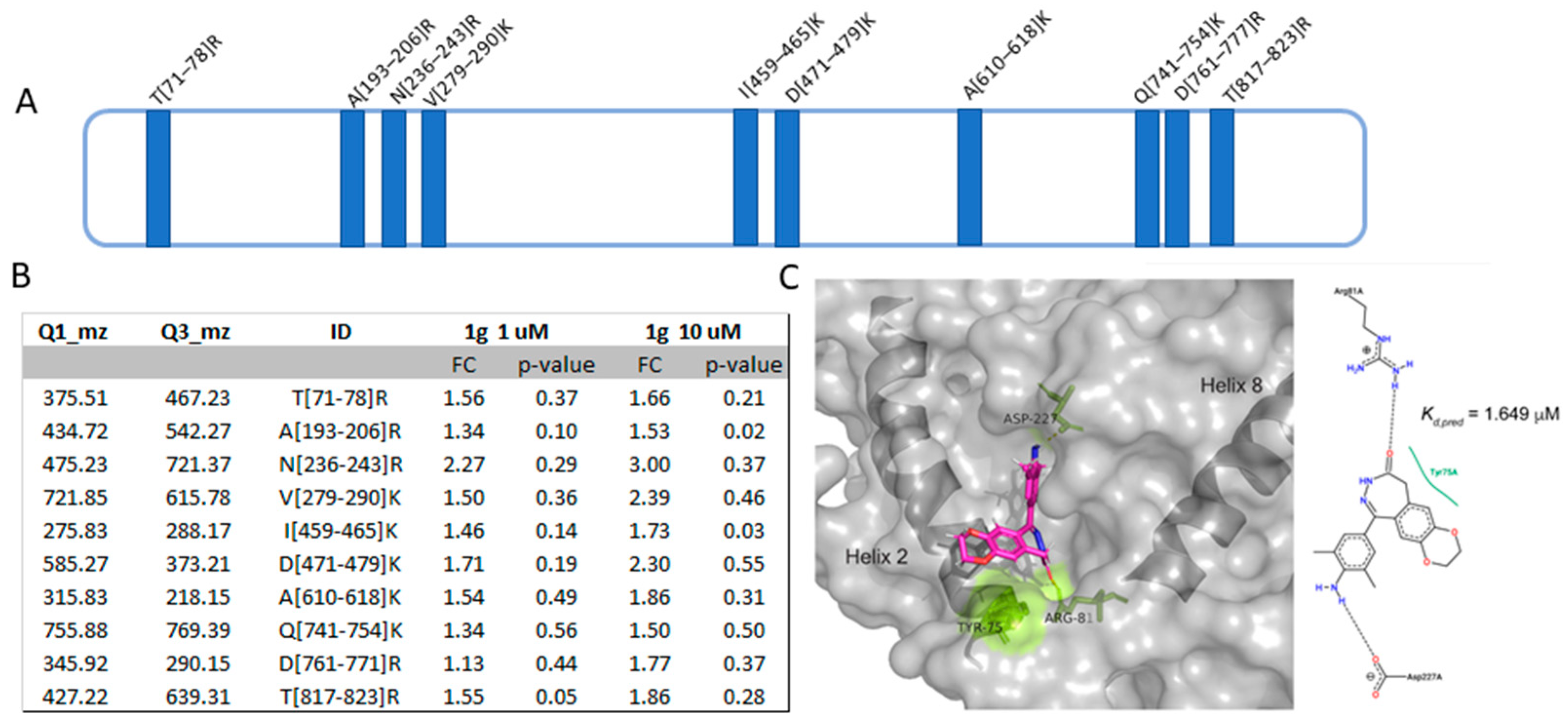
2.5. Target Identification of 1g by a Proteomic Platform
2.5.1. Identification of 1g Cellular Partner(s) through DARTS
2.5.2. Analysis of the Interaction Features at the Basis of 1g-PYGB Binding by t-LiP MRM
2.5.3. Molecular Docking Analysis of PYGB/1g Complex
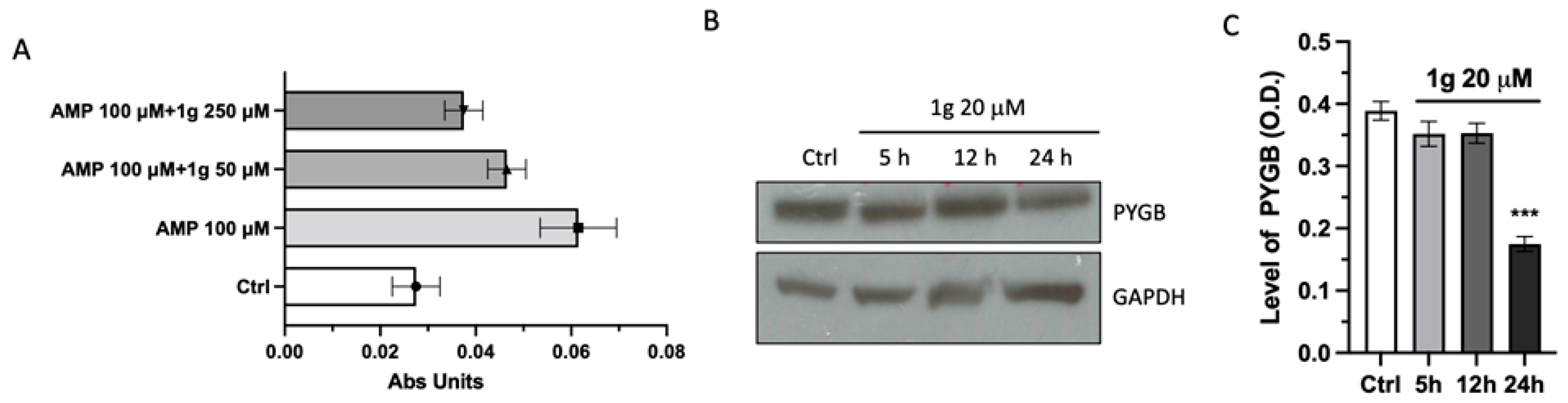
2.6. 1g Negatively Affected PYGB Expression and Activity
3. Discussion
4. Materials and Methods
4.1. 1g PAMPA Assays
4.2. Cell Cultures
4.3. Determination of Cell Growth Inhibition
4.4. Cell Cycle Analysis
4.5. Western Blot Analysis
4.6. Identification of 1g Cellular Targets
4.7. Validation of DARTS Results via Immunoblotting
4.8. t-LiP-MRM Analysis
4.9. Molecular Docking Analysis
4.10. Activity Assay
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ostrom, Q.T.; Gittleman, H.; Liao, P.; Rouse, C.; Chen, Y.; Dowling, J.; Wolinsky, Y.; Kruchko, C.; Barnholtz-Sloan, J. CBTRUS Statistical Report: Primary Brain and Central Nervous System Tumors Diagnosed in the United States in 2007–2011. Neuro. Oncol. 2014, 16 (Suppl. 4), iv1–iv63. [Google Scholar] [CrossRef] [PubMed]
- McLendon, R.; Friedman, A.; Bigner, D.; Van Meir, E.G.; Brat, D.J.; Mastrogianakis, G.M.; Olson, J.J.; Mikkelsen, T.; Lehman, N.; Aldape, K.; et al. Comprehensive Genomic Characterization Defines Human Glioblastoma Genes and Core Pathways. Nature 2008, 455, 1061–1068. [Google Scholar] [CrossRef]
- Jhanwar-Uniyal, M.; Labagnara, M.; Friedman, M.; Kwasnicki, A.; Murali, R. Glioblastoma: Molecular Pathways, Stem Cells and Therapeutic Targets. Cancers 2015, 7, 538–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuccarini, M.; Giuliani, P.; Ziberi, S.; Carluccio, M.; Di Iorio, P.; Caciagli, F.; Ciccarelli, R. The Role of Wnt Signal in Glioblastoma Development and Progression: A Possible New Pharmacological Target for the Therapy of This Tumor. Genes 2018, 9, 105. [Google Scholar] [CrossRef] [Green Version]
- Peñaranda-Fajardo, N.M.; Meijer, C.; Liang, Y.; Dijkstra, B.M.; Aguirre-Gamboa, R.; den Dunnen, W.F.A.; Kruyt, F.A.E. ER Stress and UPR Activation in Glioblastoma: Identification of a Noncanonical PERK Mechanism Regulating GBM Stem Cells through SOX2 Modulation. Cell Death Dis. 2019, 10, 690. [Google Scholar] [CrossRef]
- Alexandru, O.; Horescu, C.; Sevastre, A.S.; Cioc, C.E.; Baloi, C.; Oprita, A.; Dricu, A. Receptor Tyrosine Kinase Targeting in Glioblastoma: Performance, Limitations and Future Approaches. Contemp. Oncol. 2020, 24, 55–66. [Google Scholar] [CrossRef]
- Sevastre, A.-S.; Costachi, A.; Tataranu, L.; Brandusa, C.; Artene, S.; Stovicek, O.; Alexandru, O.; Danoiu, S.; Sfredel, V.; Dricu, A. Glioblastoma Pharmacotherapy: A Multifaceted Perspective of Conventional and Emerging Treatments (Review). Exp. Ther. Med. 2021, 22, 1408. [Google Scholar] [CrossRef]
- Chang, L.; Ying, J.; Zhimin, F. The mechanism of Warburg Effect-Induced Chemoresistance in Cancer. Front. Oncol. 2021, 11, 698023. [Google Scholar] [CrossRef]
- Grasso, S.; De Sarro, G.; De Sarro, A.; Micale, N.; Zappala, M.; Puia, G.; Baraldi, M.; De Micheli, C. Synthesis and Anticonvulsant Activity of Novel and Potent 2,3-Benzodiazepine AMPA/Kainate Receptor Antagonists. J. Med. Chem. 1999, 42, 4414–4421. [Google Scholar] [CrossRef]
- Parenti, S.; Casagrande, G.; Montanari, M.; Espahbodinia, M.; Ettari, R.; Grande, A.; Corsi, L. A Novel 2,3-Benzodiazepine-4-One Derivative AMPA Antagonist Inhibits G2/M Transition and Induces Apoptosis in Human Leukemia Jurkat T Cell Line. Life Sci. 2016, 152, 117–125. [Google Scholar] [CrossRef]
- Pirani, V.; Métivier, M.; Gallaud, E.; Thomas, A.; Ku, S.; Chretien, D.; Ettari, R.; Giet, R.; Corsi, L.; Benaud, C. A Novel Benzodiazepine Derivative That Suppresses Microtubule Dynamics and Impairs Mitotic Progression. J. Cell Sci. 2020, 133, jcs239244. [Google Scholar] [CrossRef] [PubMed]
- Rix, U.; Superti-Furga, G. Target Profiling of Small Molecules by Chemical Proteomics. Nat. Chem. Biol. 2009, 5, 616–624. [Google Scholar] [CrossRef] [PubMed]
- Ceccacci, S.; Deitersen, J.; Mozzicafreddo, M.; Morretta, E.; Proksch, P.; Wesselborg, S.; Stork, B.; Monti, M.C. Carbamoyl-Phosphate Synthase 1 as a Novel Target of Phomoxanthone A, a Bioactive Fungal Metabolite. Biomolecules 2020, 10, 846. [Google Scholar] [CrossRef]
- Morretta, E.; Esposito, R.; Festa, C.; Riccio, R.; Casapullo, A.; Monti, M.C. Discovering the Biological Target of 5-Epi-Sinuleptolide Using a Combination of Proteomic Approaches. Mar. Drugs 2017, 15, 312. [Google Scholar] [CrossRef] [PubMed]
- Morretta, E.; Belvedere, R.; Petrella, A.; Spallarossa, A.; Rapetti, F.; Bruno, O.; Brullo, C.; Monti, M.C. Novel Insights on the Molecular Mechanism of Action of the Anti-Angiogenic Pyrazolyl-Urea GeGe-3 by Functional Proteomics. Bioorg. Chem. 2021, 115, 105168. [Google Scholar] [CrossRef] [PubMed]
- Gelinas, R.P.; Froman, B.E.; McElroy, F.; Tait, R.C.; Gorin, F.A. Human Brain Glycogen Phosphorylase: Characterization of Fetal CDNA and Genomic Sequences. Brain Res. Mol. Brain Res. 1989, 6, 177–185. [Google Scholar] [CrossRef]
- Pudil, R.; Vašatová, M.; Lenčo, J.; Tichý, M.; Řeháček, V.; Fučíková, A.; Horáček, J.M.; Vojáček, J.; Pleskot, M.; Stulík, J.; et al. Plasma Glycogen Phosphorylase BB Is Associated with Pulmonary Artery Wedge Pressure and Left Ventricle Mass Index in Patients with Hypertrophic Cardiomyopathy. Clin. Chem. Lab. Med. 2010, 48, 1193–1195. [Google Scholar] [CrossRef]
- Lillpopp, L.; Tzikas, S.; Ojeda, F.; Zeller, T.; Baldus, S.; Bickel, C.; Sinning, C.R.; Wild, P.S.; Genth-Zotz, S.; Warnholtz, A.; et al. Prognostic Information of Glycogen Phosphorylase Isoenzyme BB in Patients with Suspected Acute Coronary Syndrome. Am. J. Cardiol. 2012, 110, 1225–1230. [Google Scholar] [CrossRef]
- Zhou, P.; Chi, H. AGK Unleashes CD8+ T Cell Glycolysis to Combat Tumor Growth. Cell Metab. 2019, 30, 233–234. [Google Scholar] [CrossRef]
- Cui, G.; Wang, H.; Liu, W.; Xing, J.; Song, W.; Zeng, Z.; Liu, L.; Wang, H.; Wang, X.; Luo, H.; et al. Glycogen Phosphorylase B Is Regulated by MiR101-3p and Promotes Hepatocellular Carcinoma Tumorigenesis. Front. Cell Dev. Biol. 2020, 8, 105168. [Google Scholar] [CrossRef]
- Tashima, S.; Shimada, S.; Yamaguchi, K.; Tsuruta, J.; Ogawa, M. Expression of Brain-Type Glycogen Phosphorylase Is a Potentially Novel Early Biomarker in the Carcinogenesis of Human Colorectal Carcinomas. Am. J. Gastroenterol. 2000, 95, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S., II; Lee, H.J.; Lee, S.E.; Hong, S.K.; Byun, S.S.; Lee, S.C.; Choe, G. Value of MR-US Fusion in Guidance of Repeated Prostate Biopsy in Men with PSA < 10 ng/mL. Clin. Imaging 2019, 53, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.K.; Kim, J.H.; Lee, C.H.; Kim, J.M.; Kang, C.D.; Kim, Y.D.; Choi, K.U.; Kim, H.W.; Kim, J.Y.; Park, D.Y.; et al. Clinicopathological Significance of BGP Expression in Non-Small-Cell Lung Carcinoma: Relationship with Histological Type, Microvessel Density and Patients’ Survival. Pathology 2006, 38, 555–560. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.Y.; Hua, C.H.; Li, C.H.; Zheng, R.Z.; Li, X.Y. High PYGL Expression Predicts Poor Prognosis in Human Gliomas. Front. Neurol. 2021, 12, 652931. [Google Scholar] [CrossRef]
- Festa, C.; Finamore, C.; Marchianò, S.; Di Leva, F.S.; Carino, A.; Monti, M.C.; Del Gaudio, F.; Ceccacci, S.; Limongelli, V.; Zampella, A.; et al. Investigation around the Oxadiazole Core in the Discovery of a New Chemotype of Potent and Selective FXR Antagonists. ACS Med. Chem. Lett. 2019, 10, 504–510. [Google Scholar] [CrossRef]
- Mathieu, C.; Dupret, J.M.; Rodrigues Lima, F. The Structure of Brain Glycogen Phosphorylase-from Allosteric Regulation Mechanisms to Clinical Perspectives. FEBS J. 2017, 284, 546–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crerar, M.M.; Karlsson, O.; Fletterick, R.J.; Hwang, P.K. Chimeric Muscle and Brain Glycogen Phosphorylases Define Protein Domains Governing Isozyme-Specific Responses to Allosteric Activation. J. Biol. Chem. 1995, 270, 13748–13756. [Google Scholar] [CrossRef] [Green Version]
- Fareed, M.M.; Pike, L.R.G.; Bang, A.; Huynh, M.A.; Taylor, A.; Spektor, A.; Awad, M.M.; Ott, P.A.; Krishnan, M.; Balboni, T.A.; et al. Palliative Radiation Therapy for Vertebral Metastases and Metastatic Cord Compression in Patients Treated with Anti-PD-1 Therapy. Front. Oncol. 2019, 9, 199. [Google Scholar] [CrossRef]
- Burster, T.; Traut, R.; Yermekkyzy, Z.; Mayer, K.; Westhoff, M.A.; Bischof, J.; Knippschild, U. Critical View of Novel Treatment Strategies for Glioblastoma: Failure and Success of Resistance Mechanisms by Glioblastoma Cells. Front. Cell Dev. Biol. 2021, 9, 2290. [Google Scholar] [CrossRef]
- Lew, C.R.; Guin, S.; Theodorescu, D. Targeting Glycogen Metabolism in Bladder Cancer. Nat. Rev. Urol. 2015, 12, 383–391. [Google Scholar] [CrossRef] [Green Version]
- Papadopoli, D.; Pollak, M.; Topisirovic, I. The Role of GSK3 in Metabolic Pathway Perturbations in Cancer. Biochim. Biophys. Acta Mol. Cell Res. 2021, 1868, 119059. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Zhao, Z.; Arooj, S.; Zheng, T.; Liao, G. Lapatinib Plus Local Radiation Therapy for Brain Metastases From HER-2 Positive Breast Cancer Patients and Role of Trastuzumab: A Systematic Review and Meta-Analysis. Front. Oncol. 2020, 10, 576926. [Google Scholar] [CrossRef] [PubMed]
- Favaro, E.; Bensaad, K.; Chong, M.G.; Tennant, D.A.; Ferguson, D.J.P.; Snell, C.; Steers, G.; Turley, H.; Li, J.L.; Günther, U.L.; et al. Glucose Utilization via Glycogen Phosphorylase Sustains Proliferation and Prevents Premature Senescence in Cancer Cells. Cell Metab. 2012, 16, 751–764. [Google Scholar] [CrossRef] [Green Version]
- Altemus, M.A.; Goo, L.E.; Little, A.C.; Yates, J.A.; Cheriyan, H.G.; Wu, Z.F.; Merajver, S.D. Breast Cancers Utilize Hypoxic Glycogen Stores via PYGB, the Brain Isoform of Glycogen Phosphorylase, to Promote Metastatic Phenotypes. PLoS ONE 2019, 14, e0220973. [Google Scholar] [CrossRef] [Green Version]
- Xia, B.; Zhang, K.; Liu, C. PYGB Promoted Tumor Progression by Regulating Wnt/β-Catenin Pathway in Gastric Cancer. Technol. Cancer Res. Treat. 2020, 19, 1533033820926592. [Google Scholar] [CrossRef]
- Zhan, Y.; Chen, R.; Wang, T.; Shan, S.; Zhu, H. Glycogen Phosphorylase B Promotes Cell Proliferation and Migration through PI3K/AKT Pathway in Non-Small Cell Lung Cancer. Exp. Lung Res. 2021, 47, 111–120. [Google Scholar] [CrossRef]
- Ornelas, I.M.; Silva, T.M.; Fragel-Madeira, L.; Ventura, A.L.M. Inhibition of PI3K/Akt Pathway Impairs G2/M Transition of Cell Cycle in Late Developing Progenitors of the Avian Embryo Retina. PLoS ONE 2013, 8, e53517. [Google Scholar] [CrossRef] [Green Version]
- Park, C.; Cha, H.J.; Lee, H.; Hwang-Bo, H.; Ji, S.Y.; Kim, M.Y.; Hong, S.H.; Jeong, J.W.; Han, M.H.; Choi, S.H.; et al. Induction of G2/M Cell Cycle Arrest and Apoptosis by Genistein in Human Bladder Cancer T24 Cells through Inhibition of the ROS-Dependent PI3k/Akt Signal Transduction Pathway. Antioxidants 2019, 8, 327. [Google Scholar] [CrossRef] [Green Version]
- Sarbassov, D.D.; Guertin, D.A.; Ali, S.M.; Sabatini, D.M. Phosphorylation and Regulation of Akt/PKB by the Rictor-MTOR Complex. Science 2005, 307, 1098–1101. [Google Scholar] [CrossRef] [Green Version]
- Jacinto, E.; Facchinetti, V.; Liu, D.; Soto, N.; Wei, S.; Jung, S.Y.; Huang, Q.; Qin, J.; Su, B. SIN1/MIP1 Maintains Rictor-MTOR Complex Integrity and Regulates Akt Phosphorylation and Substrate Specificity. Cell 2006, 127, 125–137. [Google Scholar] [CrossRef] [Green Version]
- Novelli, A.; Reilly, J.A.; Lysko, P.G.; Henneberry, R.C. Glutamate Becomes Neurotoxic via the N-Methyl-D-Aspartate Receptor When Intracellular Energy Levels Are Reduced. Brain Res. 1988, 451, 205–212. [Google Scholar] [CrossRef]
- Manfredini, R.; Balestri, R.; Tagliafico, E.; Trevisan, F.; Pizzanelli, M.; Grande, A.; Barbieri, D.; Zucchini, P.; Citro, G.; Franceschi, C.; et al. Antisense inhibition of c-fes proto-oncogene blocks PMA-induced macrophage differentiation in HL60 and in FDC-P1/MAC-11 cells. Blood 1997, 89, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Gemelli, C.; Dongmo, B.M.; Ferrarini, F.; Grande, A.; Corsi, L. Cytotoxic effect of hemin in colonic epithelial cell line: Involvement of 18 kDa translocator protein (TSPO). Life Sci. 2014, 107, 14–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shevchenko, A.; Tomas, H.; Havliš, J.; Olsen, J.V.; Mann, M. In-Gel Digestion for Mass Spectrometric Characterization of Proteins and Proteomes. Nat. Protoc. 2006, 1, 2856–2860. [Google Scholar] [CrossRef]
- Mathieu, C.; de la Sierra-Gallay, I.L.; Duval, R.; Xu, X.; Cocaign, A.; Léger, T.; Woffendin, G.; Camadro, J.-M.; Etchebest, C.; Haouz, A.; et al. Rodrigues-Lima. Insights into Brain Glycogen Metabolism: The structure of human brain glycogen phosphorylase. J. Biol. Chem. 2016, 291, 18072–18083. [Google Scholar] [CrossRef] [Green Version]
- Del Gaudio, F.; Pollastro, F.; Mozzicafreddo, M.; Riccio, R.; Minassi, A.; Monti, M.C. Chemoproteomic Fishing Identifies Arzanol as a Positive Modulator of Brain Glycogen Phosphorylase. Chem. Commun. 2018, 54, 12863–12866. [Google Scholar] [CrossRef]
- Grosdidier, A.; Zoete, V.; Michielin, O. SwissDock, a Protein-Small Molecule Docking Web Service Based on EADock DSS. Nucleic Acids Res. 2011, 39, W270–W277. [Google Scholar] [CrossRef] [Green Version]
- Stierand, K.; Maaß, P.C.; Rarey, M. Molecular Complexes at a Glance: Automated Generation of Two-Dimensional Complex Diagrams. Bioinformatics 2006, 22, 1710–1716. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferraro, G.; Mozzicafreddo, M.; Ettari, R.; Corsi, L.; Monti, M.C. A Proteomic Platform Unveils the Brain Glycogen Phosphorylase as a Potential Therapeutic Target for Glioblastoma Multiforme. Int. J. Mol. Sci. 2022, 23, 8200. https://doi.org/10.3390/ijms23158200
Ferraro G, Mozzicafreddo M, Ettari R, Corsi L, Monti MC. A Proteomic Platform Unveils the Brain Glycogen Phosphorylase as a Potential Therapeutic Target for Glioblastoma Multiforme. International Journal of Molecular Sciences. 2022; 23(15):8200. https://doi.org/10.3390/ijms23158200
Chicago/Turabian StyleFerraro, Giusy, Matteo Mozzicafreddo, Roberta Ettari, Lorenzo Corsi, and Maria Chiara Monti. 2022. "A Proteomic Platform Unveils the Brain Glycogen Phosphorylase as a Potential Therapeutic Target for Glioblastoma Multiforme" International Journal of Molecular Sciences 23, no. 15: 8200. https://doi.org/10.3390/ijms23158200