
Molecular Mechanisms of Drug Resistance in Staphylococcus aureus

 ,
,  and
and
Abstract
:1. Introduction
2. Resistance to Beta-Lactam Antibiotics
2.1. Synthesis of Beta-Lactamases
2.2. PBP2A Synthesis (Methicillin-Resistance)
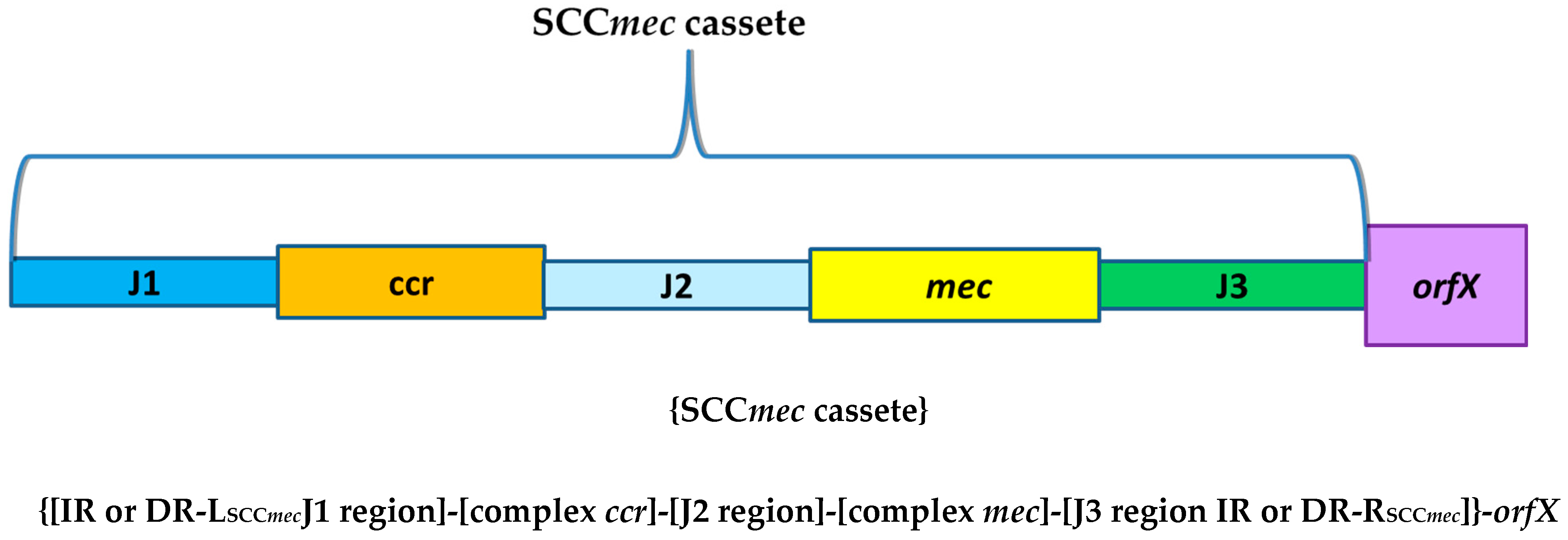
SCCmec Chromosomal Cassettes
2.3. Mutation-Dependent Modification of PBP Proteins
3. Resistance to Glycopeptides and Lipoglycopeptides
4. Resistance to Oxazolidinones
5. Resistance to Macrolides, Lincosamides, Ketolides and Streptogramins B
6. Resistance to Aminoglycosides and Spectinomycin
7. Resistance to Fluoroquinolones
8. Resistance to Tetracyclines
9. Resistance to Mupirocin
10. Resistance to Fusidic Acid
11. Resistance to Daptomycin
12. Other Antibiotics
12.1. Resistance to Streptogramins A and Quinupristin-Dalfopristin
12.2. Resistance to Rifampicin
12.3. Resistance to Chloramphenicol
12.4. Resistance to Fosfomycin
12.5. Resistance to Trimethoprim
13. Molecular Epidemiology of MRSA
14. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| aa | amino acids |
| AAC | aminoglycoside acetyltransferase |
| ABC | ATP-binding cassette transporter system |
| APH | aminoglycoside phosphotransferase |
| Bp | base pair |
| CA-MRSA | community acquired MRSA |
| CC | clonal complex |
| CFU | colony forming unit |
| CLSI | Clinical and Laboratory Standards Institute |
| CNS | coagulase negative Staphylococci |
| DHA | drug:H+ antiporter |
| EF-G | elongation factor G |
| EUCAST | European Committee on Antimicrobial SusceptibilityTesting |
| GI | genomic island |
| GISA | glycopeptide intermediate S. aureus |
| GlcNAc | N-acetylglucosamine |
| GRSA | glycopeptide resistant S. aureus |
| GSSA | glycopeptide sensitive S. aureus |
| HA-MRSA | hospital acquired or health care acquired MRSA |
| h-GISA | heterogeneous glycopeptide intermediate S. aureus |
| h-VISA | heterogeneous vancomycin intermediate S. aureus |
| In | integron |
| IS | insertion sequence |
| LA-MRSA | livestock associated MRSA |
| MBC | minimal bactericidal concentration |
| MFS | major facilitator superfamily; |
| MIC | minimal inhibitory concentration |
| MIC50 | MIC for 50% of strains tested |
| MIC90 | MIC for 90% of strains tested |
| MLS-B | macrolide-lincosamides-streptogramin B |
| MLST | multilocus sequence typing |
| MPH | macrolide phosphotransferase |
| MRSA | methicillin resistant S. aureus |
| MSSA | methicillin sensitive S. aureus |
| MurNAc | N-acetylmuramic acid |
| orf | open reading frame |
| p | plasmid |
| PAI | pathogenicity island |
| PBP | penicillin binding protein |
| PFGE | pulse field gel electrophoresis |
| PVL | Panton–Valentine leukocidin |
| PMF | proton-motive force |
| RBS | ribosome binding site |
| RND | resistance-nodulation-cell division superfamily |
| RPPs | ribosomal protection proteins |
| S. aureus SCV | S. aureus small colony variants |
| SCCmec | staphylococcal chromosomal cassette mec |
| Spa | S. aureus protein A |
| ST | sequence type |
| TMS | transmembrane segments |
| TRSA | teicoplanin resistant S. aureus |
| Tn | transposon |
| VISA | vancomycin intermediate S. aureus |
| VRSA | vancomycin resistant S. aureus |
| VSSA | vancomycin sensitive S. aureus |
References
- De Oliveira, D.M.P.; Forde, B.M.; Kidd, T.J.; Harris, P.N.A.; Schembri, M.A.; Beatson, S.A.; Paterson, D.L.; Walker, M.J. Antimicrobial resistance in ESKAPE pathogens. Clin. Microbiol. Rev. 2020, 33, e00181-19. [Google Scholar] [CrossRef] [PubMed]
- Hryniewicz, W. Epidemiology of MRSA. Infection 1999, 27, S13–S16. [Google Scholar] [CrossRef] [PubMed]
- Mlynarczyk, A.; Mlynarczyk, B.; Kmera-Muszynska, M.; Majewski, S.; Mlynarczyk, G. Mechanisms of the resistance and tolerance to beta-lactam and glycopeptide antibiotics in pathogenic gram-positive cocci. Mini Rev. Med. Chem. 2009, 9, 1527–1537. [Google Scholar] [CrossRef] [PubMed]
- Saha, B.; Singh, A.K.; Ghosh, A.; Bal, M. Identification and characterization of a vancomycin-resistant Staphylococcus aureus isolated from Kolkata (South Asia). J. Med. Microbiol. 2008, 57, 72–79. [Google Scholar] [CrossRef]
- Zhu, W.; Clark, N.C.; McDougal, L.K.; Hageman, J.; McDonald, L.C.; Patel, J.B. Vancomycin-resistant Staphylococcus aureus isolates associated with Inc18-like vanA plasmids in Michigan. Antimicrob. Agents Chemother. 2008, 52, 452–457. [Google Scholar] [CrossRef] [Green Version]
- Limbago, B.M.; Kallen, A.J.; Zhu, W.; Eggers, P.; McDougal, L.K.; Albrecht, V.S. Report of the 13th vancomycin-resistant Staphylococcus aureus isolate from the United States. J. Clin. Microbiol. 2014, 52, 998–1002. [Google Scholar] [CrossRef] [Green Version]
- McGuinness, W.A.; Malachowa, N.; DeLeo, F.R. Vancomycin resistance in Staphylococcus aureus. Yale J. Biol. Med. 2017, 90, 269–281. [Google Scholar]
- Shi, L.; Zheng, M.; Xiao, Z.; Asakura, M.; Su, J.; Li, L.; Yamasaki, S. Unnoticed spread of class 1 integrons in gram-positive clinical strains isolated in Guangzhou, China. Microbiol. Immunol. 2006, 50, 463–467. [Google Scholar] [CrossRef]
- Xu, Z.; Li, L.; Alam, M.J.; Zhang, L.; Yamasaki, S.; Shi, L. First confirmation of integron-bearing methicillin-resistant Staphylococcus aureus. Curr. Microbiol. 2008, 57, 264–268. [Google Scholar] [CrossRef]
- Butrico, C.E.; Cassat, J.E. Quorum sensing and toxin production in Staphylococcus aureus osteomyelitis: Pathogenesis and paradox. Toxins 2020, 12, 516. [Google Scholar] [CrossRef]
- Bush, K.; Jacoby, G.A. Updated functional classification of beta-lactamases. Antimicrob. Agents Chemother. 2010, 54, 969–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Álvarez, L.; Holden, M.T.; Lindsay, H.; Webb, C.R.; Brown, D.F.; Curran, M.D.; Walpole, E.; Brooks, K.; Pickard, D.J.; Teale, C.; et al. Meticillin-resistant Staphylococcus aureus with a novel mecA homologue in human and bovine populations in the UK and Denmark: A descriptive study. Lancet Infect. Dis. 2011, 11, 595–603. [Google Scholar] [CrossRef] [Green Version]
- Młynarczyk, A.; Młynarczyk, G.; Jeljaszewicz, J. The genome of Staphylococcus aureus: A review. Zentralbl. Bakteriol. 1998, 287, 277–314. [Google Scholar] [CrossRef]
- Livermore, D.M. β-Lactamases in laboratory and clinical resistance. Clin. Microbiol. Rev. 1995, 8, 557–584. [Google Scholar] [CrossRef]
- Balslev, U.; Bremmelgaard, A.; Svejgaard, E.; Havstreym, J.; Westh, H. An outbreak of borderline oxacillin-resistant Staphylococcus aureus (BORSA) in a dermatological unit. Microb. Drug Resist. 2005, 11, 78–81. [Google Scholar] [CrossRef]
- European Committee on Antimicrobial Susceptibility Testing (EUCAST) v. 12.0. 2022. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Breakpoint_tables/v_12.0_Breakpoint_Tables.pdf (accessed on 30 May 2022).
- Standards M100-S32; Performance Standarts for Antimicrobial Susceptibility Testing. Twenty-First Information Supplement. Clinical and Laboratory Standarts Institute (CLSI): Wayne, PA, USA, 2022.
- Baba, T.; Bae, T.; Schneewind, O.; Takeuchi, F.; Hiramatsu, K. Genome sequence of Staphylococcus aureus strain Newman and comparative analysis of staphylococcal genomes: Polymorphism and evolution of two major pathogenicity islands. J. Bacteriol. 2008, 190, 300–310. [Google Scholar] [CrossRef] [Green Version]
- Moreillon, P. New and emerging treatment of Staphylococcus aureus infections in the hospital setting. Clin. Microbiol. Infect. 2008, 14, 32–41. [Google Scholar] [CrossRef] [Green Version]
- Ito, T.; Okuma, K.; Ma, X.X.; Yuzawa, H.; Hiramatsu, K. Insights on antibiotic resistance of Staphylococcus aureus from its whole genome: Genomic island SCC. Drug Resist. Updat. 2003, 6, 41–52. [Google Scholar] [CrossRef]
- Taxonomy Browser (Staphylococcus). Available online: https://www.ncbi.nlm.nih.gov/Taxonomy/Browser/wwwtax.cgi (accessed on 3 February 2022).
- Cai, Y.; Zheng, L.; Lu, Y.; Zhao, X.; Sun, Y.; Tang, X.; Xiao, J.; Wang, C.; Tong, C.; Zhao, L.; et al. Inducible resistance to β-lactams in oxacillin-susceptible mecA1-positive Staphylococcus sciuri isolated from retail pork. Front. Microbiol. 2021, 12, 721426. [Google Scholar] [CrossRef]
- Lakhundi, S.; Zhang, K. Methicillin-resistant Staphylococcus aureus: Molecular characterization, evolution, and epidemiology. Clin. Microbiol. Rev. 2018, 31, e00020-18. [Google Scholar] [CrossRef] [Green Version]
- Miragaia, M. Factors contributing to the evolution of mecA-mediated β-lactam resistance in Staphylococci: Update and new insights from whole genome sequencing (WGS). Front. Microbiol. 2018, 9, 2723. [Google Scholar] [CrossRef] [Green Version]
- Becker, K.; van Alen, S.; Idelevich, E.A.; Schleimer, N.; Seggewiß, J.; Mellmann, A.; Kaspar, U.; Peters, G. Plasmid-encoded transferable mecB-mediated methicillin resistance in Staphylococcus aureus. Emerg. Infect. Dis. 2018, 24, 242–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, T.; Ma, X.X.; Takeuchi, F.; Okuma, K.; Yuzawa, H.; Hiramatsu, K. Novel type V staphylococcal cassette chromosome mec driven by a novel cassette chromosome recombinase, ccrC. Antimicrob. Agents Chemother. 2004, 48, 2637–2651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milheiriço, C.; Oliveira, D.C.; de Lencastre, H. Update to the multiplex PCR strategy for assignment of mec element types in Staphylococcus aureus. Antimicrob. Agents Chemother. 2007, 51, 3374–3377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milheiriço, C.; Oliveira, D.C.; de Lencastre, H. Multiplex PCR strategy for subtyping the staphylococcal cassette chromosome mec type IV in methicillin-resistant Staphylococcus aureus: “SCCmec IV multiplex”. J. Antimicrob. Chemother. 2007, 60, 42–48. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, D.C.; de Lencastre, H. Methicillin-resistance in Staphylococcus aureus us not affected by the overexpression in trans of the mecA gene repressor: A surprising observation. PLoS ONE 2011, 6, e23287. [Google Scholar] [CrossRef]
- Oliveira, D.C.; Tomasz, A.; de Lencastre, H. The evolution of pandemic clones of methicillin-resistant Staphylococcus aureus: Identification of two ancestral genetic backgrounds and the associated mec elements. Microb. Drug Resist. 2001, 7, 349–361. [Google Scholar] [CrossRef]
- Oliveira, D.C.; de Lencastre, H. Multiplex PCR strategy for rapid identification of structural types and variants of the mec element in methicillin- resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2002, 46, 2155–2161. [Google Scholar] [CrossRef] [Green Version]
- Shore, A.C.; Rossney, A.S.; Keane, C.T.; Enright, M.C.; Coleman, D.C. Seven novel variants of the staphylococcal chromosomal cassette mec in methicillin-resistant Staphylococcus aureus isolates from Ireland. Antimicrob. Agents Chemother. 2005, 49, 2070–2083. [Google Scholar] [CrossRef] [Green Version]
- International Working Group on the Classification of Staphylococcal Cassette Chromosome Elements (IWG-CSCCE). Classification of Staphylococcal cassette chromosome mec (SCCmec): Guidelines for reporting novel SCCmec elements. Antimicrob. Agents Chemother. 2009, 53, 4961–4967. [Google Scholar] [CrossRef] [Green Version]
- International Working Group on the Classification of Staphylococcal Cassette Chromosome (SCC) Elements (IWG-CSCCE). Available online: https://www.sccmec.org/index.php/en/component/search/?searchword=sccmec%20type1&ordering=newest&searchphrase=any (accessed on 3 February 2022).
- Urushibara, N.; Paul, S.K.; Hossain, M.A.; Kawaguchiya, M.; Kobayashi, N. Analysis of staphylococcal cassette chromosome mec in Staphylococcus haemolyticus and Staphylococcus sciuri: Identification of a novel ccr gene complex with a newly identified ccrA allotype (ccrA7). Microb. Drug Resist. 2011, 17, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Turlej, A.; Hryniewicz, W.; Empel, J. Staphylococcal cassette chromosome mec (SCCmec) classification and typing methods: An overview. Pol. J. Microbiol. 2011, 60, 95–103. [Google Scholar] [CrossRef]
- Chongtrakool, P.; Ito, T.; Ma, X.X.; Kondo, Y.; Trakulsomboon, S.; Tiensasitorn, C.; Jamklang, M.; Chavalit, T.; Song, J.H.; Hiramatsu, K. Staphylococcal cassette chromosome mec (SCCmec) typing of methicillin-resistant Staphylococcus aureus strains isolated in 11 Asian countries: A proposal for a new nomenclature for SCCmec elements. Antimicrob. Agents Chemother. 2006, 50, 1001–1012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katayama, Y.; Ito, T.; Hiramatsu, K. Genetic organization of the chromosome region surrounding mecA in clinical staphylococcal strains: Role of IS431-mediated mecI deletion in expression of resistance in mecA-carrying, low-level methicillin- resistant Staphylococcus haemolyticus. Antimicrob. Agents Chemother. 2001, 45, 1955–1963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chlebowicz, M.A.; Nganou, K.; Kozytska, S.; Arends, J.P.; Engelmann, S.; Grundmann, H.; Ohlsen, K.; van Dijl, J.M.; Buist, G. Recombination between ccrC genes in a type V (5C2&5) staphylococcal cassette chromosome mec (SCCmec) of Staphylococcus aureus ST398 leads to conversion from methicillin resistance to methicillin susceptibility in vivo. Antimicrob. Agents Chemother. 2010, 54, 783–791. [Google Scholar] [CrossRef] [Green Version]
- Zong, Z.; Lü, X. Characterization of a new SCCmec element in Staphylococcus cohnii. PLoS ONE 2010, 5, e14016. [Google Scholar] [CrossRef]
- Worthing, K.A.; Schwendener, S.; Perreten, V.; Saputra, S.; Coombs, G.W.; Pang, S.; Davies, M.R.; Abraham, S.; Trott, D.J.; Norris, J.M. Characterization of staphylococcal cassette chromosome mec elements from methicillin-resistant Staphylococcus pseudintermedius infections in australian animals. mSphere 2018, 3, e00491-18. [Google Scholar] [CrossRef] [Green Version]
- Zong, Z.; Peng, C.; Lü, X. Diversity of SCCmec elements in methicillin-resistant coagulase-negative staphylococci clinical isolates. PLoS ONE 2011, 6, e20191. [Google Scholar] [CrossRef]
- Ito, T.; Katayama, Y.; Asada, K.; Mori, N.; Tsutsumimoto, K.; Tiensasitorn, C.; Hiramatsu, K. Structural comparison of three types of staphylococcal cassette chromosome mec integrated in the chromosome in methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2001, 45, 1323–1336. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.X.; Ito, T.; Tiensasitorn, C.; Jamklang, M.; Chongtrakool, P.; Boyle-Vavra, S.; Daum, R.S.; Hiramatsu, K. Novel type of staphylococcal cassette chromosome mec identified in community-acquired methicillin-resistant Staphylococcus aureus strains. Antimicrob. Agents Chemother. 2002, 46, 1147–1152. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Skov, R.L.; Han, X.; Larsen, A.R.; Larsen, J.; Sørum, M.; Wulf, M.; Voss, A.; Hiramatsu, K.; Ito, T. Novel types of staphylococcal cassette chromosome mec elements identified in clonal complex 398 methicillin-resistant Staphylococcus aureus strains. Antimicrob. Agents Chemother. 2011, 55, 3046–3050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berglund, C.; Ito, T.; Ikeda, M.; Ma, X.X.; Söderquist, B.; Hiramatsu, K. Novel type of staphylococcal cassette chromosome mec in a methicillin-resistant Staphylococcus aureus strain isolated in Sweden. Antimicrob. Agents Chemother. 2008, 52, 3512–3516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, K.; McClure, J.A.; Elsayed, S.; Conly, J.M. Novel staphylococcal cassette chromosome mec type carrying class a mec and type 4 ccr gene complexes in a canadian epidemic strain of methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2009, 53, 531–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Z.; Li, F.; Liu, D.; Xue, H.; Zhao, X. Novel type XII staphylococcal cassette chromosome mec harboring a new cassette chromosome recombinase, CcrC2. Antimicrob. Agents Chemother. 2015, 59, 7597–7601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baig, S.; Johannesen, T.B.; Overballe-Petersen, S.; Larsen, J.; Larsen, A.R.; Stegger, M. Novel SCCmec type XIII (9A) identified in an ST152 methicillin-resistant Staphylococcus aureus. Infect. Genet. Evol. 2018, 61, 74–76. [Google Scholar] [CrossRef] [PubMed]
- Urushibara, N.; Aung, M.S.; Kawaguchiya, M.; Kobayashi, N. Novel staphylococcal cassette chromosome mec (SCCmec) type XIV (5A) and a truncated SCCmec element in SCC composite islands carrying speG in ST5 MRSA in Japan. J. Antimicrob. Chemother. 2020, 75, 46–50. [Google Scholar] [CrossRef]
- Oliveira, D.C.; Milheiriço, C.; de Lencastre, H. Redefining a structural variant of staphylococcal cassette chromosome mec, SCCmec type VI. Antimicrob. Agents Chemother. 2006, 50, 3457–3459. [Google Scholar] [CrossRef] [Green Version]
- Uehara, Y. Current status of staphylococcal cassette chromosome mec (SCCmec). Antibiotics 2022, 11, 86. [Google Scholar] [CrossRef]
- Ito, T.; Katayama, Y.; Hiramatsu, K. Cloning and nucleotide sequence determination of the entire mec DNA of pre-methicillin-resistant Staphylococcus aureus N315. Antimicrob. Agents Chemother. 1999, 43, 1449–1458. [Google Scholar] [CrossRef] [Green Version]
- Kondo, Y.; Ito, T.; Ma, X.X.; Watanabe, S.; Kreiswirth, B.N.; Etienne, J.; Hiramatsu, K. Combination of multiplex PCRs for staphylococcal cassette chromosome mec type assignment: Rapid identification system for mec, ccr, and major differences in junkyard regions. Antimicrob. Agents Chemother. 2007, 51, 264–274. [Google Scholar] [CrossRef] [Green Version]
- Hisata, K.; Kuwahara-Arai, K.; Yamanoto, M.; Ito, T.; Nakatomi, Y.; Cui, L.; Baba, T.; Terasawa, M.; Sotozono, C.; Kinoshita, S.; et al. Dissemination of methicillin-resistant staphylococci among healthy japanese children. J. Clin. Microbiol. 2005, 43, 3364–3372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, X.; Ito, T.; Takeuchi, F.; Ma, X.X.; Takasu, M.; Uehara, Y.; Oliveira, D.C.; de Lencastre, H.; Hiramatsu, K. Identification of a novel variant of staphylococcal cassette chromosome mec, type II.5, and Its truncated form by insertion of putative conjugative transposon Tn6012. Antimicrob. Agents Chemother. 2009, 53, 2616–2619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berglund, C.; Ito, T.; Ma, X.X.; Ikeda, M.; Watanabe, S.; Soderquist, B.; Hiramatsu, K. Genetic diversity of methicillin-resistant Staphylococcus aureus carrying type IV SCCmec in Orebro county and the western region of Sweden. J. Antimicrob. Chemother. 2009, 63, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-S.; Song, W.; Kim, H.-S.; Cho, H.C.; Lee, K.M.; Choi, M.-S.; Kim, E.-C. Association between the methicillin resistance of clinical isolates of Staphylococcus aureus, their staphylococcal cassette chromosome mec (SCCmec) subtype classification, and their toxin gene profiles. Diagn. Microbiol. Infect. Dis. 2006, 56, 289–295. [Google Scholar] [CrossRef]
- Kwon, N.H.; Park, K.T.; Moon, J.S.; Jung, W.K.; Kim, S.H.; Kim, J.M.; Hong, S.K.; Koo, H.C.; Joo, Y.S.; Park, Y.H. Staphylococcal cassette chromosome mec (SCCmec) characterization and molecular analysis for methicillin-resistant Staphylococcus aureus and novel SCCmec subtype IVg isolated from bovine milk in Korea. J. Antimicrob. Chemother. 2005, 56, 624–632. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.X.; Ito, T.; Chongtrakool, P.; Hiramatsu, K. Predom inance of clones carrying Panton-Valentine leukocidin genes among methicillin-resistant Staphylococcus aureus strains isolated in Japanese hospitals from 1979 to 1985. J. Clin. Microbiol. 2006, 44, 4515–4527. [Google Scholar] [CrossRef] [Green Version]
- Mongkolrattanothai, K.; Boyle, S.; Murphy, T.V.; Daum, R.S. Novel non-mecA-containing staphylococcal chromosomal cassette composite island containing pbp4 and tagF genes in a commensal staphylococcal species: A possible reservoir for antibiotic resistance islands in Staphylococcus aureus. Antimicrob. Agents Chemother. 2004, 48, 1823–1836. [Google Scholar] [CrossRef] [Green Version]
- Okuma, K.; Iwakawa, K.; Turnidge, J.D.; Grubb, W.B.; Bell, J.M.; O’Brien, F.G.; Coombs, G.W.; Pearman, J.W.; Tenover, F.C.; Kapi, M.; et al. Dissemination of new methicillin-resistant Staphylococcus aureus clones in the community. J. Clin. Microbiol. 2002, 40, 4289–4294. [Google Scholar] [CrossRef] [Green Version]
- van Hal, S.J.; Steinig, E.J.; Andersson, P.; Holden, M.T.G.; Harris, S.R.; Nimmo, G.R.; Williamson, D.A.; Heffernan, H.; Ritchie, S.R.; Kearns, A.M.; et al. Global scale dissemination of ST93: A divergent Staphylococcus aureus epidemic lineage that has recently emerged from remote northern Australia. Front. Microbiol. 2018, 9, 1453. [Google Scholar] [CrossRef]
- Harris, T.M.; Bowen, A.C.; Holt, D.C.; Sarovich, D.S.; Stevens, K.; Currie, B.J.; Howden, B.P.; Carapetis, J.R.; Giffard, P.M.; Tong, S.Y.C. Investigation of trimethoprim/sulfamethoxazole resistance in an emerging sequence type 5 methicillin-resistant Staphylococcus aureus clone reveals discrepant resistance reporting. Clin. Microbiol. Infect. 2018, 24, 1027–1029. [Google Scholar] [CrossRef] [Green Version]
- Iwao, Y.; Takano, T.; Higuchi, W.; Yamamoto, T. A new staphylococcal cassette chromosome mec IV encoding a novel cell-wall-anchored surface protein in a major ST8 community-acquired methicillin-resistant Staphylococcus aureus clone in Japan. J. Infect. Chemother. 2012, 18, 96–104. [Google Scholar] [CrossRef] [PubMed]
- McGuinness, S.L.; Holt, D.C.; Harris, T.M.; Wright, C.; Baird, R.; Giffard, P.M.; Bowen, A.C.; Tong, S.Y.C. Clinical and molecular epidemiology of an emerging Panton-Valentine leukocidin-positive ST5 methicillin-resistant Staphylococcus aureus clone in northern Australia. mSphere 2021, 6, e00651-20. [Google Scholar] [CrossRef] [PubMed]
- Hisata, K.; Ito, T.; Jin, J.; Li, S.; Watanabe, S.; Hiramatsu, K.; Matsunaga, N.; Komatsu, M.; Shimizu, T. Dissemination of multiple MRSA clones among community-associated methicillin-resistant Staphylococcus aureus infections from Japanese children with impetigo. J. Infect. Chemother. 2011, 17, 609–621. [Google Scholar] [CrossRef]
- Jorgensen, J.H. Mechanisms of methicillin resistance in Staphylococcus aureus and methods for laboratory detections. Infect. Control. Hosp. Epidemiol. 1991, 12, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Tomasz, A.; Drugeon, H.B.; de Lencastre, H.M.; Jabes, D.; McDougall, L.; Bille, J. New mechanism for methicillin resistance in Staphylococcus aureus: Clinical isolates that lack the PBP 2a gene and contain normal penicillin-binding proteins with modified penicillin-binding capacity. Antimicrob. Agents Chemother. 1989, 33, 1869–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Argudín, M.A.; Roisin, S.; Nienhaus, L.; Dodémont, M.; de Mendonça, R.; Nonhoff, C.; Deplano, A.; Denis, O. Genetic diversity among Staphylococcus aureus isolates showing oxacillin and/or cefoxitin resistance not linked to the presence of mec genes. Antimicrob. Agents Chemother. 2018, 62, e00091-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banerjee, R.; Gretes, M.; Harlem, C.; Basuino, L.; Chambers, H.F. A mecA-negative strain of methicillin-resistant Staphylococcus aureus with high-level β-lactam resistance contains mutations in three genes. Antimicrob. Agents Chemother. 2010, 54, 4900–4902. [Google Scholar] [CrossRef] [Green Version]
- Greninger, A.L.; Chatterjee, S.S.; Chan, L.C.; Hamilton, S.M.; Chambers, H.F.; Chiu, C.Y. Whole-genome sequencing of methicillin-resistant Staphylococcus aureus resistant to fifth-generation cephalosporins reveals potential non-mecA mechanisms of resistance. PLoS ONE 2016, 11, e0149541. [Google Scholar] [CrossRef]
- Hodille, E.; Delouere, L.; Bouveyron, C.; Meugnier, H.; Bes, M.; Tristan, A.; Laurent, F.; Vandenesch, F.; Lina, G.; Dumitrescu, O. In vitro activity of ceftobiprole on 440 Staphylococcus aureus strains isolated from bronchopulmonary infections. Med. Mal. Infect. 2017, 47, 152–157. [Google Scholar] [CrossRef]
- Morroni, G.; Brenciani, A.; Brescini, L.; Fioriti, S.; Simoni, S.; Pocognoli, A.; Mingoia, M.; Giovanetti, E.; Barchiesi, F.; Giacometti, A.; et al. High rate of ceftobiprole resistance among clinical methicillin-resistant Staphylococcus aureus isolates from a hospital in central Italy. Antimicrob. Agents Chemother. 2018, 62, e01663-18. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.-T.; Chen, E.-Z.; Yang, L.; Peng, C.; Wang, Q.; Xu, Z.; Chen, D.-Q. Emerging resistance mechanisms for 4 types of common anti-MRSA antibiotics in Staphylococcus aureus: A comprehensive review. Microb. Pathog. 2021, 156, 104915. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Yoon, E.-J.; Kim, D.; Kim, J.W.; Lee, K.-J.; Kim, H.S.; Kim, Y.R.; Shin, J.H.; Shin, J.H.; Shin, K.S.; et al. Ceftaroline resistance by clone-specific polymorphism in penicillin-binding protein 2a of methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2018, 62, e00485-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, D.; Debabov, D.; Hartsell, T.L.; Cano, R.J.; Adams, S.; Schuyler, J.A.; McMillan, R.; Pace, J.L. Approved glycopeptide antibacterial drugs: Mechanism of action and resistance. Cold Spring Harb. Perspect. Med. 2016, 6, a026989. [Google Scholar] [CrossRef] [Green Version]
- Courvalin, P. Vancomycin resistance in gram-positive cocci. Clin. Infect. Dis. 2006, 42, S25–S34. [Google Scholar] [CrossRef] [PubMed]
- Novais, C.; Freitas, A.R.; Sousa, J.C.; Baquero, F.; Coque, T.M.; Peixe, L.V. Diversity of Tn1546 and its role in the dissemination of vancomycin-resistant enterococci in Portugal. Antimicrob. Agents Chemother. 2008, 52, 1001–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perichon, B.; Courvalin, P. Heterologous expression of the enterococcal vanA operon in methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2004, 48, 4281–4285. [Google Scholar] [CrossRef] [Green Version]
- Centers for Disease Control and Prevention. Vancomycin-resistant Staphylococcus aureus--New York, 2004. MMWR Morb. Mortal. Wkly. Rep. 2004, 53, 322–323. [Google Scholar]
- Weigel, L.M.; Donlan, R.M.; Shin, D.H.; Jensen, B.; Clark, N.C.; McDougal, L.K.; Zhu, W.; Musser, K.A.; Thompson, J.; Kohlerschmidt, D.; et al. High-level vancomycin-resistant Staphylococcus aureus isolates associated with a polymicrobial biofilm. Antimicrob. Agents Chemother. 2007, 51, 231–238. [Google Scholar] [CrossRef] [Green Version]
- Weigel, L.M.; Clewell, D.B.; Gill, S.R.; Clark, N.C.; McDougal, L.K.; Flannagan, S.E.; Kolonay, J.F.; Shetty, J.; Killgore, G.E.; Tenover, F.C. Genetic analysis of a high-level vancomycin-resistant isolate of Staphylococcus aureus. Science 2003, 302, 1569–1571. [Google Scholar] [CrossRef]
- Bakthavatchalam, Y.D.; Babu, P.; Munusamy, E.; Dwarakanathan, H.T.; Rupali, P.; Zervos, M.; Victor, P.J.; Veeraraghavan, B. Genomic insights on heterogeneous resistance to vancomycin and teicoplanin in methicillin-resistant Staphylococcus aureus: A first report from South India. PLoS ONE 2019, 14, e0227009. [Google Scholar] [CrossRef] [Green Version]
- Szymanek-Majchrzak, K.; Mlynarczyk, A.; Mlynarczyk, G. Characteristics of glycopeptide-resistant Staphylococcus aureus strains isolated from inpatients of three teaching hospitals in Warsaw, Poland. Antimicrob. Resist. Infect. Control 2018, 7, 105. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Sun, X.; Chang, W.; Dai, Y.; Ma, X. Systematic review and meta-analysis of the epidemiology of vancomycin-intermediate and heterogeneous vancomycin-intermediate Staphylococcus aureus isolates. PLoS ONE 2015, 10, e0136082. [Google Scholar] [CrossRef] [PubMed]
- Sakurada, M.; Sumi, H.; Kaji, K.; Kobayashi, N.; Sakai, Y.; Aung, M.S.; Urushibara, N. Pacemaker-associated infection caused by ST81/SCCmec IV methicillin-resistant, vancomycin-intermediate Staphylococcus aureus in Japan. New Microbes New Infect. 2020, 35, 100656. [Google Scholar] [CrossRef]
- Katayama, Y.; Sekine, M.; Hishinuma, T.; Aiba, Y.; Hiramatsu, K. Complete reconstitution of the vancomycin-intermediate Staphylococcus aureus phenotype of strain Mu50 in vancomycin-susceptible S. aureus. Antimicrob. Agents Chemother. 2016, 60, 3730–3742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.W.; Lee, K.J. Single-nucleotide polymorphisms in a vancomycin-resistant Staphylococcus aureus strain based on whole-genome sequencing. Arch. Microbiol. 2020, 202, 2255–2261. [Google Scholar] [CrossRef]
- Picazo, J.J.; Betriu, C.; Culebras, E.; Rodríguez-Avial, I.; Gómez, M.; López-Fabal, F.; Vira, G. Methicillin-resistant Staphylococcus aureus: Changes in the susceptibility pattern to daptomycin during a 10-year period (2001–2010). Rev. Esp. Quimioter. 2011, 24, 107–111. [Google Scholar]
- Jansen, A.; Türck, M.; Szekat, C.; Nagel, M.; Clever, I.; Bierbaum, G. Role of insertion elements and yycFG in the development of decreased susceptibility to vancomycin in Staphylococcus aureus. Int. J. Med. Microbiol. 2007, 297, 205–215. [Google Scholar] [CrossRef]
- Młynarczyk, G.; Młynarczyk, A.; Zabicka, D.; Jeljaszewicz, J. Lysogenic conversion as a factor influencing the vancomycin tolerance phenomenon in Staphylococcus aureus. J. Antimicrob. Chemother. 1997, 40, 136–137. [Google Scholar] [CrossRef] [Green Version]
- Colca, J.R.; McDonald, W.G.; Waldon, D.J.; Thomasco, L.M.; Gadwood, R.C.; Lund, E.T.; Cavey, G.S.; Mathews, W.R.; Adams, L.D.; Cecil, E.T.; et al. Cross-linking in the living cell locates the site of action of oxazolidinone antibiotics. J. Biol. Chem. 2003, 278, 21972–21979. [Google Scholar] [CrossRef] [Green Version]
- Shen, T.; Penewit, K.; Waalkes, A.; Xu, L.; Salipante, S.J.; Nath, A.; Werth, B.J. Identification of a novel tedizolid resistance mutation in rpoB of MRSA after in vitro serial passage. J. Antimicrob. Chemother. 2021, 76, 292–296. [Google Scholar] [CrossRef]
- Hill, R.L.R.; Kearns, A.M.; Nash, J.; North, S.E.; Pike, R.; Newson, T.; Woodford, N.; Calver, R.; Livermore, D.M. Linezolid-resistant ST36 methicillin-resistant Staphylococcus aureus associated with prolonged linezolid treatment in two paediatric cystic fibrosis patients. J. Antimicrob. Chemother. 2010, 65, 442–445. [Google Scholar] [CrossRef] [PubMed]
- Meka, V.G.; Pillai, S.K.; Sakoulas, G.; Wennersten, C.; Venkataraman, L.; DeGirolami, P.C.; Eliopoulos, G.M.; Moellering, R.C., Jr.; Gold, H.S. Linezolid resistance in sequential Staphylococcus aureus isolates associated with a T2500A mutation in the 23S rRNA gene and loss of a single copy of rRNA. J. Infect. Dis. 2004, 190, 311–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendes, R.E.; Deshpande, L.M.; Castanheira, M.; Dipersio, J.; Saubolle, M.; Jones, R.N. First report of cfr-mediated resistance to linezolid in human staphylococcal clinical isolates recovered in the United States. Antimicrob. Agents Chemother. 2008, 52, 2244–2246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kehrenberg, C.; Aarestrup, F.M.; Schwarz, S. IS21-558 insertion sequences are involved in the mobility of the multiresistance gene cfr. Antimicrob. Agents Chemother. 2007, 51, 483–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shore, A.C.; Brennan, O.M.; Ehricht, R.; Monecke, S.; Schwarz, S.; Slickers, P.; Coleman, D.C. Identification and characterization of the multidrug resistance gene cfr in a Panton-Valentine leukocidin-positive sequence type 8 methicillin-resistant Staphylococcus aureus IVa (USA300) isolate. Antimicrob. Agents Chemother. 2010, 54, 4978–4984. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Wang, C.; Schwarz, S.; Liu, W.; Yang, Q.; Luan, T.; Wang, L.; Liu, S.; Zhang, W. Identification of a novel tetracycline resistance gene, tet(63), located on a multiresistance plasmid from Staphylococcus aureus. J. Antimicrob. Chemother. 2021, 76, 576–581. [Google Scholar] [CrossRef]
- Antonelli, A.; D’Andrea, M.M.; Brenciani, A.; Galeotti, C.L.; Morroni, G.; Pollini, S.; Varaldo, P.E.; Rossolini, G.M. Characterization of poxtA, a novel phenicol-oxazolidinone-tetracycline resistance gene from an MRSA of clinical origin. J. Antimicrob. Chemother. 2018, 73, 1763–1769. [Google Scholar] [CrossRef] [Green Version]
- Venugopal, A.A.; Johnson, S. Fidaxomicin: A novel macrocyclic antibiotic approved for treatment of Clostridium difficile infection. Clin. Infect. Dis. 2012, 54, 568–574. [Google Scholar] [CrossRef] [Green Version]
- Mlynarczyk, B.; Mlynarczyk, A.; Kmera-Muszynska, M.; Majewski, S.; Mlynarczyk, G. Mechanisms of resistance to antimicrobial drugs in pathogenic Gram-positive cocci. Mini Rev. Med. Chem. 2010, 10, 928–937. [Google Scholar] [CrossRef]
- Weisblum, B. Macrolide resistance. Drug Resist. Update 1998, 1, 29–41. [Google Scholar] [CrossRef]
- Chen, L.; Mediavilla, J.R.; Smyth, D.S.; Chavda, K.D.; Ionescu, R.; Roberts, R.B.; Robinson, D.A.; Kreiswirth, B.N. Identification of a novel transposon (Tn6072) and a truncated staphylococcal cassette chromosome mec element in methicillin-resistant Staphylococcus aureus ST239. Antimicrob. Agents Chemother. 2010, 54, 3347–3354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horinouchi, S.; Weisblum, B. Nucleotide sequence and functional map of pE194, a plasmid that specifies inducible resistance to macrolide, lincosamide, and streptogramin type B antibodies. J. Bacteriol. 1982, 150, 804–814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catchpole, I.; Thomas, C.; Davies, A.; Dyke, K.G. The nucleotide sequence of Staphylococcus aureus plasmid pT48 conferring inducible macrolide-lincosamide-streptogramin B resistance and comparison with similar plasmids expressing constitutive resistance. Microbiology 1988, 134, 697–709. [Google Scholar] [CrossRef] [Green Version]
- Projan, S.J.; Monod, M.; Narayanan, C.S.; Dubnau, D. Replication properties of pIM13, a naturally occurring plasmid found in Bacillus subtilis, and of its close relative pE5, a plasmid native to Staphylococcus aureus. J. Bacteriol. 1987, 169, 5131–5139. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, S.S.; Murphy, E.; Gamon, M.R.; Bastos, M.C. pRJ5: A naturally occurring Staphylococcus aureus plasmid expressing constitutive macrolide-lincosamide-streptogramin B resistance contains a tandem duplication in the leader region of the ermC gene. J. Gen. Microbiol. 1993, 139, 1461–1467. [Google Scholar] [CrossRef] [Green Version]
- Catchpole, I.; Dyke, K.G.H. A Staphylococcus aureus plasmid that specifies constitutive macrolide-lincosamide-streptogramin B resistance contains a novel deletion in the ermC attenuator. FEMS Microbiol. Lett. 1990, 69, 43–48. [Google Scholar] [CrossRef]
- Needham, C.; Noble, W.C.; Dyke, K.G. The staphylococcal insertion sequence IS257 is active. Plasmid 1995, 34, 198–205. [Google Scholar] [CrossRef]
- Diep, B.A.; Carleton, H.A.; Chang, R.F.; Sensabaugh, G.F.; Perdreau-Remington, F. Roles of 34 virulence genes in the evolution of hospital- and community-associated strains of methicillin-resistant Staphylococcus aureus. J. Infect. Dis. 2006, 193, 1495–1503. [Google Scholar] [CrossRef] [Green Version]
- Steward, C.D.; Raney, P.M.; Morrell, A.K.; Williams, P.P.; McDougal, L.K.; Jevitt, L.; McGowan, J.E., Jr.; Tenover, F.C. Testing for induction of clindamycin resistance in erythromycin-resistant isolates of Staphylococcus aureus. J. Clin. Microbiol. 2005, 43, 1716–1721. [Google Scholar] [CrossRef] [Green Version]
- Malhotra-Kumar, S.; Mazzariol, A.; Van Heirstraeten, L.; Lammens, C.; de Rijk, P.; Cornaglia, G.; Goossens, H. Unusual resistance patterns in macrolide-resistant Streptococcus pyogenes harbouring erm(A). J. Antimicrob. Chemother. 2009, 63, 42–46. [Google Scholar] [CrossRef] [Green Version]
- Matsuoka, M.; Inoue, M.; Nakajima, Y.; Endo, Y. New erm gene in Staphylococcus aureus clinical isolates. Antimicrob. Agents Chemother. 2002, 46, 211–215. [Google Scholar] [CrossRef] [Green Version]
- Argudín, M.A.; Tenhagen, B.-A.; Fetsch, A.; Sachsenröder, J.; Käsbohrer, A.; Schroeter, A.; Hammerl, J.A.; Hertwig, S.; Helmuth, R.; Bräunig, J.; et al. Virulence and resistance determinants of German Staphylococcus aureus ST398 isolates from nonhuman sources. Appl. Environ. Microbiol. 2011, 77, 3052–3060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takano, T.; Higuchi, W.; Otsuka, T.; Baranovich, T.; Enany, S.; Saito, K.; Isobe, H.; Dohmae, S.; Ozaki, K.; Takano, M.; et al. Novel characteristics of community-acquired methicillin-resistant Staphylococcus aureus strains belonging to multilocus sequence type 59 in Taiwan. Antimicrob. Agents Chemother. 2008, 52, 837–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luna, V.A.; Heiken, M.; Judge, K.; Ulep, C.; Van Kirk, N.; Luis, H.; Bernardo, M.; Leitao, J.; Roberts, M.C. Distribution of mef(A) in gram-positive bacteria from healthy portuguese children. Antimicrob. Agents Chemother. 2002, 46, 2513–2517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lozano, C.; Aspiroz, C.; Ara, M.; Gómez-Sanz, E.; Zarazaga, M.; Torres, C. Methicillin-resistant Staphylococcus aureus (MRSA) ST398 in a farmer with skin lesions and in pigs of his farm: Clonal relationship and detection of lnu(A) gene. Clin. Microbiol. Infect. 2011, 17, 923–927. [Google Scholar]
- Lozano, C.; Rezusta, A.; Gómez, P.; Gómez-Sanz, E.; Báez, N.; Martin-Saco, G.; Zarazaga, M.; Torres, C. High prevalence of spa types associated with the clonal lineage CC398 among tetracycline-resistant methicillin-resistant Staphylococcus aureus strains in a Spanish hospital. J. Antimicrob. Chemother. 2012, 67, 330–334. [Google Scholar] [CrossRef] [Green Version]
- Schmitz, F.J.; Petridou, J.; Fluit, A.C.; Hadding, U.; Peters, G.; von Eiff, C. Distribution of macrolide-resistance genes in Staphylococcus aureus blood-culture isolates from fifteen german university hospitals. Eur. J. Clin. Microbiol. Infect. Dis. 2000, 19, 385–387. [Google Scholar] [CrossRef]
- Schmitz, F.-J.; Sadurski, R.; Kray, A.; Boos, M.; Geisel, R.; Köhrer, K.; Verhoef, J.; Fluit, A.C. Prevalence of macrolide-resistance genes in Staphylococcus aureus and Enterococcus faecium isolates from 24 European university hospitals. J. Antimicrob. Chemother. 2000, 45, 891–894. [Google Scholar] [CrossRef] [Green Version]
- Cassone, M.; D’Andrea, M.M.; Iannelli, F.; Oggioni, M.R.; Rossolini, G.M.; Pozzi, G. DNA microarray for detection of macrolide resistance genes. Antimicrob. Agents Chemother. 2006, 50, 2038–2041. [Google Scholar] [CrossRef] [Green Version]
- Qin, X.; Poon, B.; Kwong, J.; Niles, D.; Schmidt, B.Z.; Rajagopal, L.; Gantt, S. Two paediatric cases of skin and soft-tissue infections due to clindamycin-resistant Staphylococcus aureus carrying a plasmid-encoded vga(A) allelic variant for a putative efflux pump. Int. J. Antimicrob. Agents 2011, 38, 81–83. [Google Scholar] [CrossRef]
- Wang, B.; Xu, Y.; Zhao, H.; Wang, X.; Rao, L.; Guo, Y.; Yi, X.; Hu, L.; Chen, S.; Han, L.; et al. Methicillin-resistant Staphylococcus aureus in China: A multicenter longitudinal study and whole-genome sequencing. Emerg. Microbes Infect. 2022, 11, 532–542. [Google Scholar] [CrossRef] [PubMed]
- Schnellmann, C.; Gerber, V.; Rossano, A.; Jaquier, V.; Panchaud, Y.; Doherr, M.G.; Thomann, A.; Straub, R.; Perreten, V. Presence of new mecA and mph(C) variants conferring antibiotic resistance in Staphylococcus spp. isolated from the skin of horses before and after clinic admission. J. Clin. Microbiol. 2006, 44, 4444–4454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loeza-Lara, P.D.; Soto-Huipe, M.; Baizabal-Aguirre, V.M.; Ochoa-Zarzosa, A.; Valdez-Alarcón, J.J.; Cano-Camacho, H.; López-Meza, J.E. pBMSa1, a plasmid from a dairy cow isolate of Staphylococcus aureus, encodes a lincomycin resistance determinant and replicates by the rolling-circle mechanism. Plasmid 2004, 52, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Luthje, P.; Schwarz, S. Molecular basis of resistance to macrolides and lincosamides among staphylococci and streptococci from various animal sources collected in the resistance monitoring program BfT-Germ. Vet. Int. J. Antimicrob. Agents 2007, 29, 528–535. [Google Scholar] [CrossRef]
- Wondrack, L.; Massa, M.; Yang, B.V.; Sutcliffe, J. Clinical strain of Staphylococcus aureus inactivates and causes efflux of macrolides. Antimicrob. Agents Chemother. 1996, 40, 992–998. [Google Scholar] [CrossRef] [Green Version]
- Allignet, J.; Liassine, N.; El Solh, N. Characterization of a staphylococcal plasmid related to pUB110 and carrying two novel genes, vatC and vgbB, encoding resistance to streptogramins A and B and similar antibiotics. Antimicrob. Agents Chemother. 1998, 42, 1794–1798. [Google Scholar] [CrossRef] [Green Version]
- Haroche, J.; Morvan, A.; Davi, M.; Allignet, J.; Bimet, F.; El Solh, N. Clonal diversity among streptogramin A-resistant Staphylococcus aureus isolates collected in french hospitals. J. Clin. Microbiol. 2003, 41, 586–591. [Google Scholar] [CrossRef] [Green Version]
- Mukhtar, T.A.; Koteva, K.P.; Hughes, D.W.; Wright, G.D. Vgb from Staphylococcus aureus inactivates streptogramin B antibiotics by an elimination mechanism not hydrolysis. Biochemistry 2001, 40, 8877–8886. [Google Scholar] [CrossRef]
- Malbruny, B.; Canu, A.; Bozdogan, B.; Fantin, B.; Zarrouk, V.; Dutka-Malen, S.; Feger, C.; Leclercq, R. Resistance to quinupristin-dalfopristin due to mutation of L22 ribosomal protein in Staphylococcus aureus. Antimicrob. Agents Chemother. 2002, 46, 2200–2207. [Google Scholar] [CrossRef] [Green Version]
- Prunier, A.L.; Trong, H.N.; Tande, D.; Segond, C.; Leclercq, R. Mutation of L4 ribosomal protein conferring unusual macrolide resistance in two independent clinical isolates of Staphylococcus aureus. Microb. Drug Resist. 2005, 11, 18–20. [Google Scholar] [CrossRef]
- Chandrakanth, R.; Raju, S.; Patil, S.A. Aminoglycoside-resistance mechanisms in multidrug-resistant Staphylococcus aureus clinical isolates. Curr. Microbiol. 2008, 56, 558–562. [Google Scholar] [CrossRef] [PubMed]
- Emaneini, M.; Taherikalani, M.; Eslampour, M.A.; Sedaghat, H.; Aligholi, M.; Jabalameli, F.; Shahsavan, S.; Sotoudeh, N. Phenotypic and genotypic evaluation of aminoglycoside resistance in clinical isolates of staphylococci in Tehran, Iran. Microb. Drug Resist. 2009, 15, 129–132. [Google Scholar] [CrossRef] [PubMed]
- Rossolini, G.M.; Mantengoli, E. Antimicrobial resistance in Europe and its potential impact on empirical therapy. Clin. Microbiol. Infect. 2008, 14, 2–8. [Google Scholar] [CrossRef] [Green Version]
- Clark, J.A.; Burgess, D.S. Plazomicin: A new aminoglycoside in the fight against antimicrobial resistance. Ther. Adv. Infect. Dis. 2020, 7, 2049936120952604. [Google Scholar] [CrossRef] [PubMed]
- Melter, O.; Radojevič, B. Small colony variants of Staphylococcus aureus—Review. Folia Microbiol. 2010, 55, 548–558. [Google Scholar] [CrossRef] [PubMed]
- Ardic, N.; Sareyyupoglu, B.; Ozyurt, M.; Haznedaroglu, T.; Ilga, U. Investigation of aminoglycoside modifying enzyme genes in methicillin-resistant staphylococci. Microbiol. Res. 2006, 161, 49–54. [Google Scholar] [CrossRef]
- Ida, T.; Okamoto, R.; Shimauchi, C.; Okubo, T.; Kuga, A.; Inoue, M. Identification of aminoglycoside-modifying enzymes by susceptibility testing: Epidemiology of methicillin-resistant Staphylococcus aureus in Japan. J. Clin. Microbiol. 2001, 39, 3115–3121. [Google Scholar] [CrossRef] [Green Version]
- Raju, S.; Oli, A.K.; Patil, S.A.; Chandrakanth, R.K. Prevalence of multidrug-resistant Staphylococcus aureus in diabetics clinical samples. World J. Microbiol. Biotechnol. 2010, 26, 171–176. [Google Scholar] [CrossRef]
- Yadegar, A.; Sattari, M.; Mozafari, N.A.; Goudarzi, G.R. Prevalence of the genes encoding aminoglycoside-modifying enzymes and methicillin resistance among clinical isolates of Staphylococcus aureus in Tehran, Iran. Microb. Drug Resist. 2009, 15, 109–113. [Google Scholar] [CrossRef]
- Szymanek-Majchrzak, K.; Mlynarczyk, A.; Kawecki, D.; Pacholczyk, M.; Durlik, M.; Deborska-Materkowska, D.; Paczek, L.; Mlynarczyk, G. Resistance to aminoglycosides of methicillin-resistant strains of Staphylococcus aureus, originating in the surgical and transplantation wards of the Warsaw clinical center—A retrospective analysis. Transplant. Proc. 2018, 50, 2170–2175. [Google Scholar] [CrossRef]
- Lowy, D. Antimicrobial resistance: The example of Staphylococcus aureus. J. Clin. Investig. 2003, 111, 1265–1273. [Google Scholar] [CrossRef] [PubMed]
- Ida, T.; Okamoto, R.; Nonoyama, M.; Irinoda, K.; Kurazono, M.; Inoue, M. Antagonism between aminoglycosides and beta-lactams in a methicillin-resistant Staphylococcus aureus isolate involves induction of an aminoglycoside-modifying enzyme. Antimicrob. Agents Chemother. 2002, 46, 1516–1521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujimura, S.; Tokue, Y.; Takahashi, H.; Kobayashi, T.; Gomi, K.; Abe, T.; Nukiwa, T.; Watanabe, A. Novel arbekacin- and amikacin-modifying enzyme of methicillin-resistant Staphylococcus aureus. FEMS Microbiol. Lett. 2000, 190, 299–303. [Google Scholar] [CrossRef]
- Derbise, A.; de Cespedes, G.; El Solh, N. Nucleotide sequence of the Staphylococcus aureus transposon, Tn5405, carrying aminoglycosides resistance genes. J. Basic Microbiol. 1997, 37, 379–384. [Google Scholar] [CrossRef]
- Derbise, A.; Dyke, K.G.; El Solh, N. Characterization of a Staphylococcus aureus transposon, Tn5405, located within Tn5404 and carrying the aminoglycoside resistance genes, aphA-3 and aadE. Plasmid 1996, 35, 174–188. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Li, L.; Shirtliff, M.E.; Peters, B.M.; Li, B.; Peng, Y.; Alam, M.J.; Yamasaki, S.; Shi, L. Resistance class 1 integron in clinical methicillin-resistant Staphylococcus aureus strains in southern China, 2001–2006. Clin. Microbiol. Infect. 2011, 17, 714–718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, Y.; Onodera, Y.; Lee, J.C.; Hooper, D.C. NorB, an efflux pump in Staphylococcus aureus strain MW2, contributes to bacterial fitness in abscesses. J. Bacteriol. 2008, 190, 7123–7129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, J.; Thanassi, J.A.; Thoma, C.L.; Bradbury, B.J.; Deshpande, M.; Pucci, M.J. Dual targeting of DNA gyrase and topoisomerase IV: Target interactions of heteroaryl isothiazolones in Staphylococcus aureus. Antimicrob. Agents Chemother. 2007, 51, 2445–2453. [Google Scholar] [CrossRef] [Green Version]
- Hooper, D.C. Mechanisms of quinolone resistance. In Gram-Positive Pathogens; Fischetti, V.A., Novick, R.P., Ferretti, J.J., Portnoy, D.A., Rood, J.I., Eds.; ASM Press: Washington, WA, USA, 2006; pp. 821–833. [Google Scholar]
- Guay, G.G.; Khan, S.A.; Rothstein, D.M. The tet(K) gene of plasmid pT181 of Staphylococcus aureus encodes an efflux protein that contains 14 transmembrane helices. Plasmid 1993, 30, 163–166. [Google Scholar] [CrossRef]
- Noguchi, N.; Aoki, T.; Sasatsu, M.; Kono, M.; Shishido, K.; Ando, T. Determination of the complete nucleotide sequence of pNS1, a staphylococcal tetracycline-resistance plasmid propagated in Bacillus subtilis. FEMS Microbiol. Lett. 1986, 37, 283–288. [Google Scholar] [CrossRef]
- Moon, K.H.; Kim, W.K.; Yoon, S.J.; Kim, M.; Shin, C.K.; Im, S.H. Association of tet gene with partial sequence of IS431mec in tetracycline resistance plasmid pKH1. Arch. Pharmacal Res. 1996, 19, 171–172. [Google Scholar]
- Truong-Bolduc, Q.C.; Dunman, P.M.; Strahilevitz, J.; Projan, S.J.; Hooper, D.C. MgrA is a multiple regulator of two new efflux pumps in Staphylococcus aureus. J. Bacteriol. 2005, 187, 2395–2405. [Google Scholar] [CrossRef] [Green Version]
- Trzciński, K.; Cooper, B.S.; Hryniewicz, W.; Dowson, C.G. Expression of resistance to tetracyclines in strains of methicillin-resistant Staphylococcus aureus. J. Antimicrob. Chemother. 2000, 45, 763–770. [Google Scholar] [CrossRef] [Green Version]
- McDougal, L.K.; Fosheim, G.E.; Nicholson, A.; Bulens, S.N.; Limbago, B.M.; Shearer, J.E.; Summers, A.O.; Patel, J.B. Emergence of resistance among USA300 methicillin-resistant Staphylococcus aureus isolates causing invasive disease in the United States. Antimicrob. Agents Chemother. 2010, 54, 3804–3811. [Google Scholar] [CrossRef] [Green Version]
- De Vries, L.E.; Christensen, H.; Skov, R.L.; Aarestrup, F.M.; Agersø, Y. Diversity of the tetracycline resistance gene tet(M) and identification of Tn916- and Tn5801-like (Tn6014) transposons in Staphylococcus aureus from humans and animals. J. Antimicrob. Chemother. 2009, 64, 490–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dyke, K.G.H.; Curnock, S.P.; Golding, M.; Noble, W.C. Cloning of the gene conferring resistance to mupirocin in Staphylococcus aureus FEMS Microbiol. Lett. 1991, 77, 195–198. [Google Scholar]
- Yoo, J.I.; Shin, E.S.; Chung, G.T.; Lee, K.M.; Yoo, J.S.; Lee, Y.S. Restriction fragment length polymorphism (RFLP) patterns and sequence analysis of high-level mupirocin-resistant meticillin-resistant staphylococci. Int. J. Antimicrob. Agents 2010, 35, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Cadilla, A.; David, M.Z.; Daum, R.S.; Boyle-Vavra, S. Association of high-level mupirocin resistance and multi-drug resistant methicillin-resistant Staphylococcus aureus at an academic center in the midwestern United States. J. Clin. Microbiol. 2011, 49, 95–100. [Google Scholar] [CrossRef] [Green Version]
- Caffrey, A.R.; Quilliam, B.J.; LaPlante, K.L. Risk factors associated with mupirocin resistance in meticillin-resistant Staphylococcus aureus. J. Hosp. Infect. 2010, 76, 206–210. [Google Scholar] [CrossRef]
- Perez-Roth, E.; Kwong, S.M.; Alcoba-Florez, J.; Firth, N.; Mendez-Alvarez, S. Complete nucleotide sequence and comparative analysis of pPR9, a 41.7-kilobase conjugative staphylococcal multiresistance plasmid conferring high-level mupirocin resistance. Antimicrob. Agents Chemother. 2010, 54, 2252–2257. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, A.K.; Skov, R.L.; Venezia, R.A.; Johnson, J.K.; Stender, H. Evaluation of mupA EVIGENE assay for determination of high-level mupirocin resistance in Staphylococcus aureus. J. Clin. Microbiol. 2010, 48, 4253–4255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seah, C.; Alexander, D.C.; Louie, L.; Simor, A.; Low, D.E.; Longtin, J.; Melano, R.G. MupB, a new high-level mupirocin resistance mechanism in Staphylococcus aureus. Antimicrob. Agents Chemother. 2012, 56, 1916–1920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurdle, J.G.; O’Neill, A.J.; Chopra, I. The isoleucyl-tRNA synthetase mutation V588F conferring mupirocin resistance in glycopeptide-intermediate Staphylococcus aureus is not associated with a significant fitness burden. J. Antimicrob. Chemother. 2004, 53, 102–104. [Google Scholar] [CrossRef] [PubMed]
- Lannergard, J.; Norstrom, T.; Hughes, D. Genetic determinants of resistance to fusidic acid among clinical bacteremia isolates of Staphylococcus aureus. Antimicrob. Agents Chemother. 2009, 53, 2059–2065. [Google Scholar] [CrossRef] [Green Version]
- Mairi, A.; Touati, A.; Pantel, A.; Martinez, A.Y.; Ahmim, M.; Sotto, A.; Dunyach-Remy, C.; Lavigne, J.P. First report of CC5-MRSA-IV-SCCfus “maltese clone” in Bat Guano. Microorganisms 2021, 9, 2264. [Google Scholar] [CrossRef]
- Lacey, R.W.; Rosdahl, V.T. An unusual “penicillinase plasmid” in Staphylococcus aureus; evidence for its transfer under natural conditions. J. Med. Microbiol. 1974, 7, 1–9. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, F.G.; Price, C.; Grubb, W.B.; Gustafson, J.E. Genetic characterization of the fusidic acid and cadmium resistance determinants of Staphylococcus aureus plasmid pUB101. J. Antimicrob. Chemother. 2002, 50, 313–321. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, A.J.; Chopra, I. Molecular basis of fusB-mediated resistance to fusidic acid in Staphylococcus aureus. Mol. Microbiol. 2006, 59, 664–676. [Google Scholar] [CrossRef]
- O’Neill, A.J.; Larsen, A.R.; Skov, R.; Henriksen, A.S.; Chopra, I. Characterization of the epidemic european fusidic acid-resistant impetigo clone of Staphylococcus aureus. J. Clin. Microbiol. 2007, 45, 1505–1510. [Google Scholar] [CrossRef] [Green Version]
- Mehta, S.; Cuirolo, A.X.; Plata, K.B.; Riosa, S.; Silverman, J.A.; Rubio, A.; Rosato, R.R.; Rosato, A.E. VraSR two-component regulatory system contributes to mprF-mediated decreased susceptibility to daptomycin in in vivo-selected clinical strains of methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2012, 56, 92–102. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.J.; Kreiswirth, B.N.; Sakoulas, G.; Yeaman, M.R.; Xiong, Y.Q.; Sawa, A.; Bayer, A.S. Enhanced expression of dltABCD is associated with the development of daptomycin nonsusceptibility in a clinical endocarditis isolate of Staphylococcus aureus. J. Infect. Dis. 2009, 200, 1916–1920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.J.; Xiong, Y.Q.; Dunman, P.M.; Schrenzel, J.; Franc, P.; Peschel, A.; Bayer, A.S. Regulation of mprF in daptomycin-nonsusceptible Staphylococcus aureus strains. Antimicrob. Agents Chemother. 2009, 53, 2636–2637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, A.; Grein, F.; Otto, A.; Gries, K.; Orlov, D.; Zarubaev, V.; Girard, M.; Sher, X.; Shamova, O.; Roemer, T.; et al. Differential daptomycin resistance development in Staphylococcus aureus strains with active and mutated gra regulatory systems. Int. J. Med. Microbiol. 2018, 308, 335–348. [Google Scholar] [CrossRef]
- Allignet, J.; Loncle, V.; Simenel, C.; Delepierre, M.; El Solh, N. Sequence of a staphylococcal gene, vat, encoding an acetyltransferase inactivating the A-type compounds of virginiamycin-like antibiotics. Gene 1993, 130, 91–98. [Google Scholar] [CrossRef]
- Allignet, J.; El Solh, N. Diversity among the gram-positive acetyltransferases inactivating streptogramin A and structurally related compounds and characterization of a new staphylococcal determinant, vatB. Antimicrob. Agents Chemother. 1995, 39, 2027–2036. [Google Scholar] [CrossRef] [Green Version]
- Haroche, J.; Allignet, J.; El Solh, N. Tn5406, a new staphylococcal transposon conferring resistance to streptogramin a and related compounds including dalfopristin. Antimicrob. Agents Chemother. 2002, 46, 2337–2343. [Google Scholar] [CrossRef] [Green Version]
- Haroche, J.; Allignet, J.; Buchrieser, C.; El Solh, N. Characterization of a variant of vga(A) conferring resistance to streptogramin A and related compounds. Antimicrob. Agents Chemother. 2000, 44, 2271–2275. [Google Scholar] [CrossRef] [Green Version]
- Kadlec, K.; Schwarz, S. Novel ABC transporter gene, vga(C), located on a multiresistance plasmid from a porcine methicillin-resistant Staphylococcus aureus ST398 strain. Antimicrob. Agents Chemother. 2009, 53, 3589–3591. [Google Scholar] [CrossRef] [Green Version]
- Schwendener, S.; Perreten, V. New transposon Tn6133 in methicillin-resistant Staphylococcus aureus ST398 contains vga(E), a novel streptogramin a, pleuromutilin, and lincosamide resistance gene. Antimicrob. Agents Chemother. 2011, 55, 4900–4904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allington, D.R.; Rivey, M.P. Quinupristin/dalfopristin: A therapeutic review. Clin. Ther. 2001, 23, 24–44. [Google Scholar] [CrossRef]
- Pankuch, G.A.; Jacobs, M.R.; Appelbaum, P.C. Postantibiotic effect and postantibiotic sub-MIC effect of quinupristin-dalfopristin against gram-positive and -negative organisms. Antimicrob. Agents Chemother. 1998, 42, 3028–3031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aubry-Damon, H.; Soussy, C.J.; Courvalin, P. Characterization of mutations in the rpoB gene that confer rifampin resistance in Staphylococcus aureus. Antimicrob. Agents Chemother. 1998, 42, 2590–2594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sekiguchi, J.-I.; Fujino, T.; Araake, M.; Toyota, E.; Kudo, K.; Saruta, K.; Kirikae, T.; Yoshikura, H.; Kuratsuji, T. Emergence of rifampicin resistance in methicillin-resistant Staphylococcus aureus in tuberculosis wards. J. Infect. Chemother. 2006, 12, 47–50. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, S.; Witte, D.G. Phenicol resistance. In Frontiers in Antimicrobial Resistance: Atribute to Stuart B. Levy; White, D.G., Alekshun, M.N., McDermott, P.F., Eds.; ASM Press: Washington, WA, USA, 2005; pp. 124–147. [Google Scholar]
- Projan, S.J.; Kornblum, J.; Moghazeh, S.L.; Edelman, I.; Gennaro, M.L.; Novick, R.P. Comparative sequence and functional analysis of pT181 and pC221, cognate plasmid replicons from Staphylococcus aureus. Mol. Gen. Genet. 1985, 199, 452–464. [Google Scholar] [CrossRef]
- Tennent, J.M.; May, J.W.; Skurray, R.A. Characterisation of chloramphenicol resistance plasmids of Staphylococcus aureus and S. epidermidis by restriction enzyme mapping techniques. J. Med. Microbiol. 1986, 22, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Brenner, D.G.; Shaw, W.V. The use of synthetic oligonucleotides with universal templates for rapid DNA sequencing: Results with staphylococcal replicon pC221. EMBO J. 1985, 4, 561–568. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, S.; Cardoso, M. Nucleotide sequence and phylogeny of a chloramphenicol acetyltransferase encoded by the plasmid pSCS7 from Staphylococcus aureus. Antimicrob. Agents Chemother. 1991, 35, 1551–1556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarz, S.; Spies, U.; Cardoso, M. Cloning and sequence analysis of a plasmid-encoded chloramphenicol acetyltransferase gene from Staphylococcus intermedius. J. Gen. Microbiol. 1991, 137, 977–981. [Google Scholar] [CrossRef] [Green Version]
- Smith, M.C.; Thomas, C.D. An accessory protein is required for relaxosome formation by small staphylococcal plasmids. J. Bacteriol. 2004, 186, 3363–3373. [Google Scholar] [CrossRef] [Green Version]
- Bhakta, M.; Bal, M. Identification and characterization of a shuttle plasmid with antibiotic resistance gene from Staphylococcus aureus. Curr. Microbiol. 2003, 46, 413–417. [Google Scholar] [CrossRef]
- Wright, G.D. Bacterial resistance to antibiotics: Enzymatic degradation and modification. Adv. Drug Deliv. Rev. 2005, 57, 1451–1470. [Google Scholar] [CrossRef] [PubMed]
- Neoh, H.M.; Cui, L.; Yuzawa, H.; Takeuchi, F.; Matsuo, M.; Hiramatsu, K. Mutated response regulator graR is responsible for phenotypic conversion of Staphylococcus aureus from heterogeneous vancomycin-intermediate resistance to vancomycin-intermediate resistance. Antimicrob. Agents Chemother. 2008, 52, 45–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouch, D.A.; Messerotti, L.J.; Loo, L.S.; Jackson, C.A.; Skurray, R.A. Trimethoprim resistance transposon Tn4003 from Staphylococcus aureus encodes genes for a dihydrofolate reductase and thymidylate synthetase flanked by three copies of IS257. Mol. Microbiol. 1989, 3, 161–175. [Google Scholar] [CrossRef]
- Burdeska, A.; Ott, M.; Bannwarth, W.; Then, R.L. Identical genes for trimethoprim-resistant dihydrofolate reductasefrom Staphylococcus aureus in Australia and central Europe. FEBS Lett. 1990, 266, 159–162. [Google Scholar] [CrossRef] [Green Version]
- Kadlec, K.; Schwarz, S. Identification of a novel trimethoprim resistance gene, dfrK, in a methicillin-resistant Staphylococcus aureus ST398 strain and its physical linkage to the tetracycline resistance gene tet(L). Antimicrob. Agents Chemother. 2009, 53, 776–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadlec, K.; Schwarz, S. Identification of a plasmid-borne resistance gene cluster comprising the resistance genes erm(T), dfrK, and tet(L) in a porcine methicillin-resistant Staphylococcus aureus ST398 strain. Antimicrob. Agents Chemother. 2010, 54, 915–918. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Chlebowicz-Flissikowska, M.A.; Wang, M.; Murguia, E.V.; de Jong, A.; Becher, D.; Maaß, S.; Buist, G.; van Dijl, J.M. Exoproteomic profiling uncovers critical determinants for virulence of livestock-associated and human-originated Staphylococcus aureus ST398 strains. Virulence 2020, 11, 947–963. [Google Scholar] [CrossRef]
- Amdahl, H.; Haapasalo, K.; Tan, L.; Meri, T.; Kuusela, P.I.; van Strijp, J.A.; Rooijakkers, S.; Jokiranta, T.S. Staphylococcal protein Ecb impairs complement receptor-1 mediated recognition of opsonized bacteria. PLoS ONE 2017, 12, e0172675. [Google Scholar] [CrossRef] [Green Version]
- Lamret, F.; Varin-Simon, J.; Velard, F.; Terryn, C.; Mongaret, C.; Colin, M.; Gangloff, S.C.; Reffuveille, F. Staphylococcus aureus strain-dependent biofilm formation in bone-like environment. Front. Microbiol. 2021, 12, 714994. [Google Scholar] [CrossRef]
- Langley, R.J.; Ting, Y.T.; Clow, F.; Young, P.G.; Radcliff, F.J.; Choi, J.M.; Sequeira, R.P.; Holtfreter, S.; Baker, H.; Fraser, J.D. Staphylococcal enterotoxin-like X (SElX) is a unique superantigen with functional features of two major families of staphylococcal virulence factors. PLoS Pathog. 2017, 13, e1006549. [Google Scholar] [CrossRef] [Green Version]
- Porayath, C.; Suresh, M.K.; Biswas, R.; Nair, B.G.; Mishra, N.; Pal, S. Autolysin mediated adherence of Staphylococcus aureus with fibronectin, gelatin and heparin. Int. J. Biol. Macromol. 2018, 110, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Hirschhausen, N.; Schlesier, T.; Peters, G.; Heilmann, C. Characterization of the modular design of the autolysin/adhesin Aaa from Staphylococcus aureus. PLoS ONE 2012, 7, e40353. [Google Scholar] [CrossRef] [PubMed]
- Foster, T.J. Surface proteins of Staphylococcus aureus. Microbiol. Spectr. 2019, 7, 4. [Google Scholar] [CrossRef]
- Foster, T.J. The MSCRAMM family of cell-wall-anchored surface proteins of gram-positive cocci. Trends Microbiol. 2019, 27, 927–941. [Google Scholar] [CrossRef]
- Viela, F.; Speziale, P.; Pietrocola, G.; Dufrêne, Y.F. Bacterial pathogens under high-tension: Staphylococcus aureus adhesion to von Willebrand factor is activated by force. Microb. Cell 2019, 6, 321–323. [Google Scholar] [CrossRef]
- Ohnishi, Y.; Okino, N.; Ito, M.; Imayama, S. Ceramidase activity in bacterial skin flora as a possible cause of ceramide deficiency in atopic dermatitis. Clin. Diagn. Lab. Immunol. 1999, 6, 101–104. [Google Scholar] [CrossRef] [Green Version]
- Pickering, A.C.; Yebra, G.; Gong, X.; Goncheva, M.I.; Wee, B.A.; MacFadyen, A.C.; Muehlbauer, L.F.; Alves, J.; Cartwright, R.A.; Paterson, G.K.; et al. Evolutionary and functional analysis of coagulase positivity among the Staphylococci. mSphere 2021, 6, e0038121. [Google Scholar] [CrossRef]
- Courjon, J.; Munro, P.; Benito, Y.; Visvikis, O.; Bouchiat, C.; Boyer, L.; Doye, A.; Lepidi, H.; Ghigo, E.; Lavigne, J.P.; et al. EDIN-B promotes the translocation of Staphylococcus aureus to the bloodstream in the course of pneumonia. Toxins 2015, 7, 4131–4142. [Google Scholar] [CrossRef] [Green Version]
- Wright, J.A.; Nair, S.P. The lipoprotein components of the Isd and Hts transport systems are dispensable for acquisition of heme by Staphylococcus aureus. FEMS Microbiol. Lett. 2012, 329, 177–185. [Google Scholar] [CrossRef] [Green Version]
- Conroy, B.S.; Grigg, J.C.; Kolesnikov, M.; Morales, L.D.; Murphy, M.E.P. Staphylococcus aureus heme and siderophore-iron acquisition pathways. BioMetals 2019, 32, 409–424. [Google Scholar] [CrossRef]
- Sun, L.; Wu, D.; Chen, Y.; Wang, Q.; Wang, H.; Yu, Y. Characterization of a PVL-negative community-acquired methicillin-resistant Staphylococcus aureus strain of sequence type 88 in China. Int. J. Med. Microbiol. 2017, 307, 346–352. [Google Scholar] [CrossRef] [PubMed]
- Uehara, Y.; Sasaki, T.; Baba, T.; Lu, Y.; Imajo, E.; Sato, Y.; Tanno, S.; Furuichi, M.; Kawada, M.; Hiramatsu, K. Regional outbreak of community-associated methicillin-resistant Staphylococcus aureus ST834 in Japanese children. BMC Infect. Dis. 2019, 19, 35. [Google Scholar] [CrossRef] [PubMed]
- Heuer, H.; Krogerrecklenfort, E.; Wellington, E.M.; Egan, S.; Elsas, J.D.; Overbeek, L.; Collard, J.M.; Guillaume, G.; Karagouni, A.D.; Nikolakopoulou, T.L.; et al. Gentamicin resistance genes in environmental bacteria: Prevalence and transfer. FEMS Microbiol. Ecol. 2002, 42, 289–302. [Google Scholar] [CrossRef] [PubMed]
- Strauß, L.; Stegger, M.; Akpaka, P.E.; Alabi, A.; Breurec, S.; Coombs, G.; Egyir, B.; Larsen, A.R.; Laurent, F.; Monecke, S.; et al. Origin, evolution, and global transmission of community-acquired Staphylococcus aureus ST8. Proc. Natl. Acad. Sci. USA 2017, 114, E10596–E10604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baig, S.; Larsen, A.R.; Simões, P.M.; Laurent, F.; Johannesen, T.B.; Lilje, B.; Tristan, A.; Schaumburg, F.; Egyir, B.; Cirkovic, I.; et al. Evolution and population dynamics of clonal complex 152 community-associated methicillin-resistant Staphylococcus aureus. mSphere 2020, 5, e00226-20. [Google Scholar] [CrossRef]
- Ogura, K.; Kaji, D.; Sasaki, M.; Otsuka, Y.; Takemoto, N.; Miyoshi-Akiyama, T.; Kikuchi, K. Predominance of ST8 and CC1/spa-t1784 methicillin-resistant Staphylococcus aureus isolates in Japan and their genomic characteristics. J. Glob. Antimicrob. Resist. 2022, 28, 195–202. [Google Scholar] [CrossRef]
- D’Souza, N.; Rodrigues, C.; Mehta, A. Molecular characterization of methicillin-resistant Staphylococcus aureus with emergence of epidemic clones of sequence type (ST) 22 and ST 772 in Mumbai. J. Clin. Microbiol. 2010, 48, 1806–1811. [Google Scholar] [CrossRef] [Green Version]
- Broderick, D.; Brennan, G.I.; Drew, R.J.; O’Connell, B. Epidemiological typing of methicillin resistant Staphylococcus aureus recovered from patients attending a maternity hospital in Ireland 2014–2019. Infect. Prev. Pract. 2021, 3, 100124. [Google Scholar] [CrossRef]
- Senok, A.; Ehricht, R.; Monecke, S.; Al-Saedan, R.; Somily, A. Molecular characterization of methicillin-resistant Staphylococcus aureus in nosocomial infections in a tertiary-care facility: Emergence of new clonal complexes in Saudi Arabia. New Microbes New Infect. 2016, 14, 13–18. [Google Scholar] [CrossRef] [Green Version]
- Earls, M.R.; Coleman, D.C.; Brennan, G.I.; Fleming, T.; Monecke, S.; Slickers, P.; Ehricht, R.; Shore, A.C. Intra-hospital, inter-hospital and intercontinental spread of ST78 MRSA from two neonatal intensive care unit outbreaks established using whole-genome sequencing. Front. Microbiol. 2018, 9, 1485. [Google Scholar] [CrossRef]
- Kikuta, H.; Shibata, M.; Nakata, S.; Yamanaka, T.; Sakata, H.; Akizawa, K.; Kobayashi, K. Predominant dissemination of PVL-negative CC89 MRSA with SCCmec Type II in children with impetigo in Japan. Int. J. Pediatr. 2011, 2011, 143872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tristan, A.; Bes, M.; Meugnier, H.; Lina, G.; Bozdogan, B.; Courvalin, P.; Reverdy, M.E.; Enright, M.C.; Vandenesch, F.; Etienne, J. Global distribution of Panton-Valentine leucocidin–positive methicillin-resistant Staphylococcus aureus, 2006. Emerg. Infect. Dis. 2007, 13, 594–600. [Google Scholar] [CrossRef] [PubMed]
- Egyir, B.; Guardabassi, L.; Sørum, M.; Nielsen, S.S.; Kolekang, A.; Frimpong, E.; Addo, K.K.; Newman, M.J.; Larsen, A.R. Molecular epidemiology and antimicrobial susceptibility of clinical Staphylococcus aureus from healthcare institutions in Ghana. PLoS ONE 2014, 9, e89716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monecke, S.; Aamot, H.V.; Stieber, B.; Ruppelt, A.; Ehricht, R. Characterization of PVL-positive MRSA from Norway. APMIS 2014, 122, 580–584. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Zhou, W.; Yin, Z.; Zhang, S.; Chen, Y.; Shen, P.; Ji, J.; Chen, W.; Zheng, B.; Xiao, Y. The genetic feature and virulence determinant of highly virulent community-associated MRSA ST338-SCCmec Vb in China. Emerg. Microbes Infect. 2021, 10, 1052–1064. [Google Scholar] [CrossRef]
- Boswihi, S.S.; Udo, E.E.; AlFouzan, W. Antibiotic resistance and typing of the methicillin-resistant Staphylococcus aureus clones in Kuwait hospitals, 2016-2017. BMC Microbiol. 2020, 20, 314. [Google Scholar] [CrossRef]
- Xiao, M.; Wang, H.; Zhao, Y.; Mao, L.-L.; Brown, M.; Yu, Y.-S.; O’Sullivan, M.V.N.; Kong, F.; Xu, Y.-C. National surveillance of methicillin-resistant Staphylococcus aureus in China highlights a still-evolving epidemiology with 15 novel emerging multilocus sequence types. J. Clin. Microbiol. 2013, 51, 3638–3644. [Google Scholar] [CrossRef] [Green Version]
- Neradova, K.; Fridrichova, M.; Jakubu, V.; Pomorska, K.; Zemlickova, H. Epidemiological characteristics of methicillin-resistant Staphylococcus aureus isolates from bloodstream cultures at University Hospital in the Czech Republic. Folia Microbiol. 2020, 65, 615–622. [Google Scholar] [CrossRef] [Green Version]
- Mlynarczyk, A.; Szymanek-Majchrzak, K.; Grzybowska, W.; Durlik, M.; Deborska-Materkowska, D.; Paczek, L.; Chmura, A.; Swoboda-Kopec, E.; Tyski, S.; Mlynarczyk, G. Molecular and phenotypic characteristics of methicillin-resistant Staphylococcus aureus strains isolated from hospitalized patients in transplantation wards. Transplant. Proc. 2014, 46, 2579–2582. [Google Scholar] [CrossRef]
- Szymanek-Majchrzak, K.; Mlynarczyk, A.; Dobrzaniecka, K.; Majchrzak, K.; Mierzwinska-Nastalska, E.; Chmura, A.; Kwiatkowski, A.; Durlik, M.; Deborska-Materkowska, D.; Paczek, L.; et al. Epidemiological and drug-resistance types of methicillin-resistant Staphylococcus aureus strains isolated from surgical and transplantation ward patients during 2010 to 2011. Transplant. Proc. 2016, 48, 1414–1417. [Google Scholar] [CrossRef]
- Yu, F.; Cienfuegos-Gallet, A.V.; Cunningham, M.H.; Jin, Y.; Wang, B.; Kreiswirth, B.N.; Chen, L. Molecular evolution and adaptation of livestock-associated methicillin-resistant Staphylococcus aureus (LA-MRSA) sequence type 9. mSystems 2021, 6, e0049221. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Jia, M.; Cheng, Y.; Zhang, W.; Lu, Q.; Guo, Y.; Wen, G.; Shao, H.; Luo, Q. First report of ST9-MRSA-XII from a chicken farm in China. J. Glob. Antimicrob. Resist. 2021, 27, 292–293. [Google Scholar] [CrossRef] [PubMed]
- Juhász-Kaszanyitzky, E.; Jánosi, S.; Somogyi, P.; Dán, A.; van der Graaf-van Bloois, L.; van Duijkeren, E.; Wagenaar, J.A. MRSA transmission between cows and humans. Emerg. Infect. Dis. 2007, 13, 630–632. [Google Scholar] [CrossRef]
- Hau, S.J.; Kellner, S.; Eberle, K.C.; Waack, U.; Brockmeier, S.L.; Haan, J.S.; Davies, P.R.; Frana, T.; Nicholson, T.L. Methicillin-resistant Staphylococcus aureus sequence type (ST) 5 isolates from health care and agricultural sources adhere equivalently to human keratinocytes. Appl. Environ. Microbiol. 2018, 84, e02073-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, G.; Matuszewska, M.; Jia, M.; Zhou, J.; Ba, X.; Duan, J.; Zhang, C.; Zhao, J.; Tao, M.; Fan, J.; et al. A survey of chinese pig farms and human healthcare isolates reveals separate human and animal methicillin-resistant Staphylococcus aureus populations. Adv. Sci. 2022, 9, 2103388. [Google Scholar] [CrossRef]
- Gómez-Sanz, E.; Torres, C.; Lozano, C.; Fernández-Pérez, R.; Aspiroz, C.; Ruiz-Larrea, F.; Zarazaga, M. Detection, molecular characterization, and clonal diversity of methicillin-resistant Staphylococcus aureus CC398 and CC97 in spanish slaughter pigs of different age groups. Foodborne Pathog. Dis. 2010, 7, 1269–1277. [Google Scholar] [CrossRef]
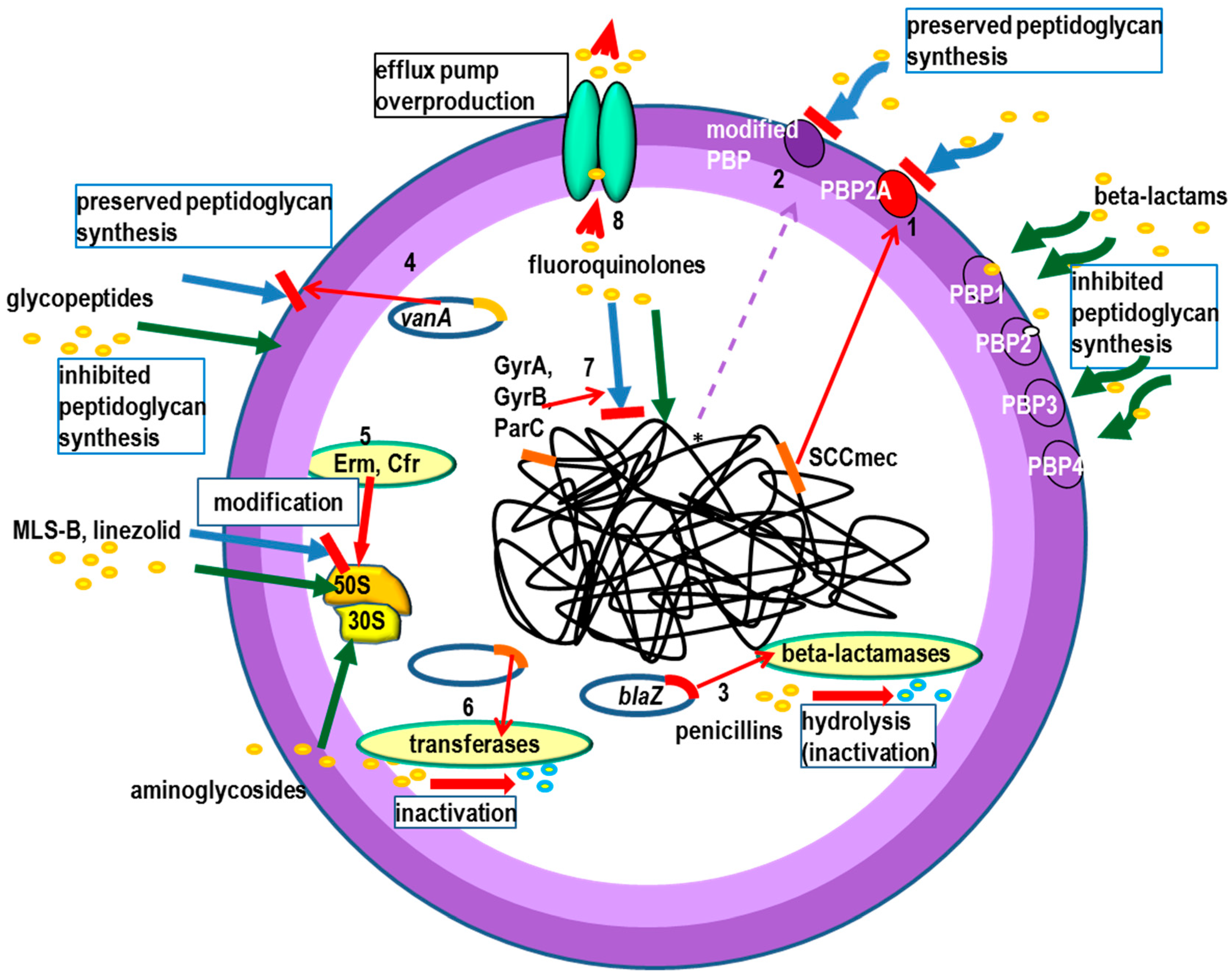
 , mechanisms of action—green arrows. Resistance to beta lactams: 1. Production of penicillin-binding protein PBP2A, 2. * mutations in PBP genes—rare (MODSA), 3. beta-lactamases production -usually narrow substrate spectrum. Glycopeptide resistance: 4. VanA operon (modification of the antibiotic binding site), Linezolid resistance: 5. adenylyl-N-methyltransferase Cfr-modification 23S rRNA of bacterial ribosome. Resistance to MLS-B (macrolides, lincosamides and streptogramins B): 5. Erm—erythromycin ribosome methylation. Aminoglycosides resistance: 6. antibiotics inactivation by tansferases. Fluoroinolones resistance: 7. mutations in gyrA and gyrB (topoisomerase II) and parC (grlA) and parE (topoisomerase IV) genes (modification of the antibiotic binding site), 8. removal from the bacterial cell by the efflux pump.
, mechanisms of action—green arrows. Resistance to beta lactams: 1. Production of penicillin-binding protein PBP2A, 2. * mutations in PBP genes—rare (MODSA), 3. beta-lactamases production -usually narrow substrate spectrum. Glycopeptide resistance: 4. VanA operon (modification of the antibiotic binding site), Linezolid resistance: 5. adenylyl-N-methyltransferase Cfr-modification 23S rRNA of bacterial ribosome. Resistance to MLS-B (macrolides, lincosamides and streptogramins B): 5. Erm—erythromycin ribosome methylation. Aminoglycosides resistance: 6. antibiotics inactivation by tansferases. Fluoroinolones resistance: 7. mutations in gyrA and gyrB (topoisomerase II) and parC (grlA) and parE (topoisomerase IV) genes (modification of the antibiotic binding site), 8. removal from the bacterial cell by the efflux pump.
, mechanisms of action—green arrows. Resistance to beta lactams: 1. Production of penicillin-binding protein PBP2A, 2. * mutations in PBP genes—rare (MODSA), 3. beta-lactamases production -usually narrow substrate spectrum. Glycopeptide resistance: 4. VanA operon (modification of the antibiotic binding site), Linezolid resistance: 5. adenylyl-N-methyltransferase Cfr-modification 23S rRNA of bacterial ribosome. Resistance to MLS-B (macrolides, lincosamides and streptogramins B): 5. Erm—erythromycin ribosome methylation. Aminoglycosides resistance: 6. antibiotics inactivation by tansferases. Fluoroinolones resistance: 7. mutations in gyrA and gyrB (topoisomerase II) and parC (grlA) and parE (topoisomerase IV) genes (modification of the antibiotic binding site), 8. removal from the bacterial cell by the efflux pump.
, mechanisms of action—green arrows. Resistance to beta lactams: 1. Production of penicillin-binding protein PBP2A, 2. * mutations in PBP genes—rare (MODSA), 3. beta-lactamases production -usually narrow substrate spectrum. Glycopeptide resistance: 4. VanA operon (modification of the antibiotic binding site), Linezolid resistance: 5. adenylyl-N-methyltransferase Cfr-modification 23S rRNA of bacterial ribosome. Resistance to MLS-B (macrolides, lincosamides and streptogramins B): 5. Erm—erythromycin ribosome methylation. Aminoglycosides resistance: 6. antibiotics inactivation by tansferases. Fluoroinolones resistance: 7. mutations in gyrA and gyrB (topoisomerase II) and parC (grlA) and parE (topoisomerase IV) genes (modification of the antibiotic binding site), 8. removal from the bacterial cell by the efflux pump.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class of mec Gene Complex | mec Complex | SCCmec in S. aureus |
|---|---|---|
| A | IS431-mecA-mecR1-mecI | II, III, VIII, XIII, XIV |
| B | IS431-mecA-ΔmecR1-ΨIS1272 | I, IV, VI |
| B2 | IS431-mecA-ΔmecR1-Tn4001-ΨIS1272 | IV |
| C1 | IS431→-mecA-ΔmecR1-IS431→ | VII, X |
| C2 | IS431→-mecA-ΔmecR1-IS431← | V, IX, XII |
| E | blaZ-mecC-mecR1c-mecIc | XI |
| Number of ccr Gene Complex | Gene of ccr | Type of SCCmec |
|---|---|---|
| 1 | A1B1 | I, IX |
| 2 | A2B2 | II, IV |
| 3 | A3B3 | III |
| 4 | A4B4 | VI, VIII |
| 5 | C1 * | V, VII, XIV |
| 7 | A1B6 | X |
| 8 | A1B3 | XI |
| 9 | C2 | XII, XIII |
| SCCmec Type | Representative Strain | Isolated in | GenBank Accession | SCCmec (kb) | ccr Complex | mec Complex | Other Genes and Genetic Elements |
|---|---|---|---|---|---|---|---|
| No. | in SCCmec | ||||||
| I | NCTC10442 (JCSC9884) | England;1961 | AB033763 | 34.4 | 1 | B | |
| II | N315 (JCSC9885) | Japan; 1981 | D86934 | 53.0 | 2 | A | pUB110, Tn554 |
| III | 85/2082 (JCSC9889) | New Zealand; 1985 | AB037671 | 66.9 | 3 | A | SCCHg, ΨTn554, pT181 |
| IV | CA05 (JCSC9890) | USA; 1999 | AB063172 | 24.3 | 2 | B | - |
| V | WIS (JCSC9897) | Australia; 1995 | AB121219 | 27.6 | 5 | C2 | hsdR, hsdS, hsdM |
| VI | HDE288 (JCSC9900) | Portugal; 1996 | AF411935 | 23.0 | 4 | B | - |
| VII | P5747/2002 (JCSC9900) | Sweden; 2002 | AB373032 | 32.4 | 5 | C1 | hsdR, hsdM |
| VIII | C10682 (JCSC9902) | Canada; 2003 | FJ390057 | 32.1 | 4 | A | Tn554 |
| IX | JCSC6943 (JCSC9903) | Thailand; 2006 | AB505628 | 43.7 | 1 | C2 | arsDARBC, cadDX arsRBC, cadDX |
| X | JCSC6945 (JCSC9904) | Canada; 2006 | AB505630 | 50.8 | 7 | C1 | |
| XI | LGA251 (JCSC9905) | England; 2007 | FR821779 | 29.4 | 8 | E | arsRBC, blaZ |
| XII | BA01611 | China; 2015 | KR187111 | 49.3 | 9 | C2 | ΨSCCBA01611 |
| XIII | 55-99-44 | Denmark; 2018 | MG674089 | 29.2 | 9 | A | Tn4001 |
| XIV | SC792 (JCSC11500) | Japan; 2013–2014 | LC440647 | 81.5 | 5 | A | ΨSCCpls; ACME II’; SCCSC640 |
| CC | Clone | Spa Type | Agr Type | PVL | Other Name of Clone |
|---|---|---|---|---|---|
| 1 | ST1-IV/V | t125; t127; t128; t175; t273; t558; t1178; t1272; t1274 t1784; t5388 | 3 | +/- | USA400; MW2; WA MRSA 1/45, 1/57; PFGE-1I; cMRSA; USA400 ORSA IV |
| 1 | ST772-V | t345; t345; t657; t1839; t3387; t5414; t10795; | 2 | + | Bengal Bay Clone; WA MRSA 60 |
| 5 | ST5-IV/IV+ SCCfus/V/VI | t001; t002; t003; t311; t450; t1277; t2460 | 2 | + | Peadiatric; Maltese; USA800; HDE288; Portoguese peadiatric |
| 8 | ST8-IV | t008; t024; t064; t068; t112; t121; t451; t622; t1476 | 1 | + | USA300; USA300-0114; USA300vLA; CMRSA10; PFGE-B; CA-MRSA/J |
| 8 | ST72-IV/V | t126; t148; t324; t537; t664 | 1 | +/- | USA700 ORSA IV; |
| 8 | ST612-IV | t1257 | 1 | - | PFGE-A6 |
| 8 | ST2021-V | t024 | + | ||
| 9 | ST834-IV | t1379; t9624 | - | ||
| 15 | ST15-IV | t084/t085 | 2 | + | |
| 22 | ST22-IV/V | t005; t022; t032; t223; t310; t891 | 1 | +/- | UK EMRSA-15, Barnim; PFGE-B |
| 22 | ST766-V | t1276 | 1 | + | |
| 30 | ST30-IV | t019; t021; t318; t975; t1273 | 3 | + | Oceania Southwest Pacific; Uruguayan 6; Mexican; USA1100; Southwest Pacific; PFGE-N; HKU-100 |
| 45 | ST45-IV/V | t004/t026/t040 | 1 | PFGE-E | |
| 59 | ST59-IV/V/VII | t163; t172; t216; t316; t437; t528; t976; t3523 | 1 | +/- | USA1000; HKU200; Western Australia MRSA-9, -15, -52, -55, -56, -73; Taiwan; Asian-Pacific, PFGE-A |
| 59 | ST87-IVb (2B) | t216 | 1 | - | Western Australia MRSA-24 |
| 59 | ST338-IV/V | t437; t441 | 1 | + | |
| 80 | ST80-IV | t044; t131; t359; t376; t639; t1199; t1200; t1201; t1206 | 3 | +/- | European; PFGE-G2; cMRSA |
| 88 | ST78-IV | t186; t690; t786; t1598; t2832; t3205 | - | ||
| 88 | ST88-IV | t168/t186/t690/t729 | 3 | +/- | African; PFGE-J |
| 89 | ST89-IV | - | PFGE-1B | ||
| 89 | ST91-IV | t416/t604 | PFGE-3B | ||
| 93 | ST93-IV | t202 | 3 | + | Queensland; PFGE-E |
| 121 | ST121-V | t159/t314 | + | ||
| 152 | ST152-V | t355 | + | Balkan Region | |
| 152 | ST789-IV | t547 | + | PFGE-1B |
| CC | Clone | Spa Type | Agr Type | Other Names |
|---|---|---|---|---|
| 5 | ST5-I/II | t001; t002; t003; t214; t242; t311; t586; t2460 | 2 | UK EMRSA-3; Southern German MRSA, Rhine Hesse MRSA, Cordobes/ Chilean; PFGE-C; Geraldine; Pediatric; New York/Japan; USA100, CMRSA2; GISA |
| 5 | ST225-II | t003; t014; t151, t1282; t1623 | Rhine Hesse MRSA, EMRSA-3, New York | |
| 5 | ST228-I | t001; t023 | 2 | Southern German MRSA, Rhine Hesse MRSA, EMRSA-3, New York |
| 5 | ST764-II | t002; t1064 | ||
| 5 | ST2590-II | t002 | 2 | |
| 5 | ST105-II | t002 | 2 | |
| 8 | ST8-II/IV | t008; t064; t068; t190 | 1 | Irish-1; UK EMRSA-2/-6/; USA500 ORSA IV, USA500 ORSA II, ST8 ORSA I, ST8 ORSA IV, ST8 ORSA III; Archaic/Iberian |
| 8 | ST239-III | t030; t037 | 1 | Hungarian; Brazilian/Hungarian; UK EMRSA-1/-4/-11; Vienna; Australian, AUS-2, AUS-3 (2000); East Australian; PFGE-B; CC8/239; ST239 ORSA III; Eurasian; Brazilian; Portuguese; PFGE-B |
| 8 | ST240-III | t037 | ST240 ORSA III, | |
| 8 | ST241-II/III | t037; t138 | 1 | Finland-UK |
| 8 | ST247-I | t008; t051; t052; t054 | 1 | Iberian, UK EMRSA-5/-7/-17; PFGE-A; ST247 ORSA I |
| 8 | ST250-I | t008; t194; t292 | 1/4 | Archaic, ST250 ORSA I; EMRSA-8 |
| 8 | ST254-I/IV | t009 | 1 | UK EMRSA-10, Hannover MRSA |
| 22 | ST22-III/IV/V | t022; t032; t223 | 1/2 | PFGE-B |
| 30 | ST30-I | t018; t019; t037; t268; t318 | 3 | EMRSA-16, USA200 ORSA II |
| 30 | ST36-II | t018; t268 | 3 | UK EMRSA-16; USA200; CMRSA4/8/9 |
| 45 | ST45-II/IV | t004; t015; t026; t038; t445 | 1/4 | USA600; CMRSA1; Berlin MRSA; USA600 ORSA II; USA600 ORSA IV |
| 89 | ST89-II | t3520 | 3 |
| CC | Clone | Spa Type | Agr Type | Other Name |
|---|---|---|---|---|
| 1 | ST1-IVa | t125; t127; t128; t1178 | 3 | USA400 |
| 5 | ST5-IV | t002; t003; t311 | 2 | PFGE-I |
| 9 | ST9-III/IV/V/XII/IV+XII | t099; t100; t193; t411; t464, t526, t587, t800; t899; t1334, t1430; t2315; t2700; t3446; t4132; t4358; t4794; t13493; t29922 | 2 | GER-MRSA-ST9, CHN-MRSA-ST9 |
| 97 | ST97-IV/V | t1234 | 1 | |
| 97 | ST1379-V | t3992 | 1 | |
| 130 | ST130-XI | t373, t843 | ||
| 398 | ST398-IV/V/VII | t011; t034; t571; t1197; t1250; t1255; t1451; t1456; t1928; t2510 | 1 | GER-MRSA-ST398, CHN-MRSA-ST398 |
| 398 | ST1232-V | t034 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mlynarczyk-Bonikowska, B.; Kowalewski, C.; Krolak-Ulinska, A.; Marusza, W. Molecular Mechanisms of Drug Resistance in Staphylococcus aureus. Int. J. Mol. Sci. 2022, 23, 8088. https://doi.org/10.3390/ijms23158088
Mlynarczyk-Bonikowska B, Kowalewski C, Krolak-Ulinska A, Marusza W. Molecular Mechanisms of Drug Resistance in Staphylococcus aureus. International Journal of Molecular Sciences. 2022; 23(15):8088. https://doi.org/10.3390/ijms23158088
Chicago/Turabian StyleMlynarczyk-Bonikowska, Beata, Cezary Kowalewski, Aneta Krolak-Ulinska, and Wojciech Marusza. 2022. "Molecular Mechanisms of Drug Resistance in Staphylococcus aureus" International Journal of Molecular Sciences 23, no. 15: 8088. https://doi.org/10.3390/ijms23158088