
Characterization of the Putative Acylated Cellulose Synthase Operon in Komagataeibacter xylinus E25

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
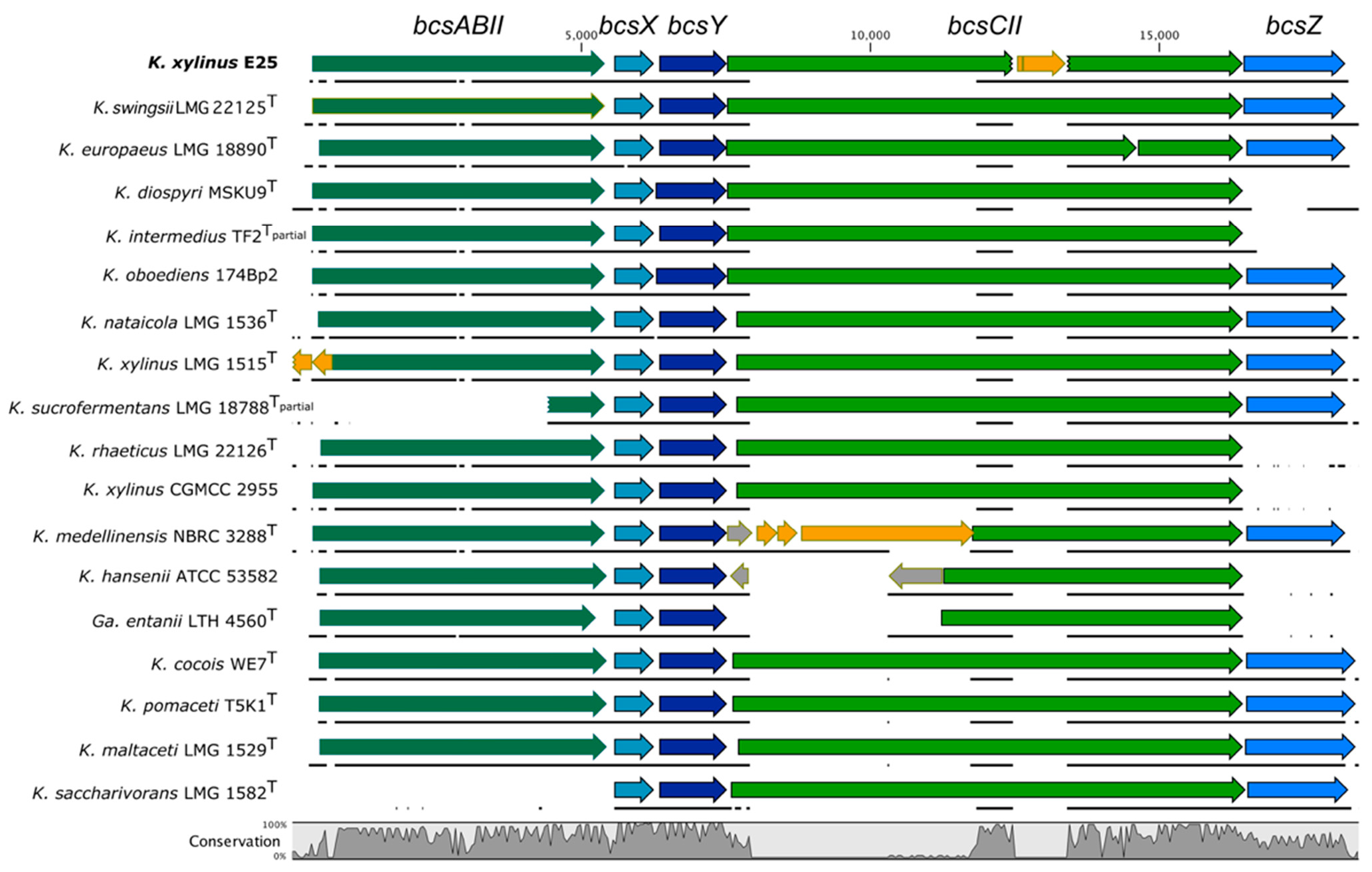
2.1. Diversity of the BcsII Operon in the Genomes of the Komagataeibacter Genus
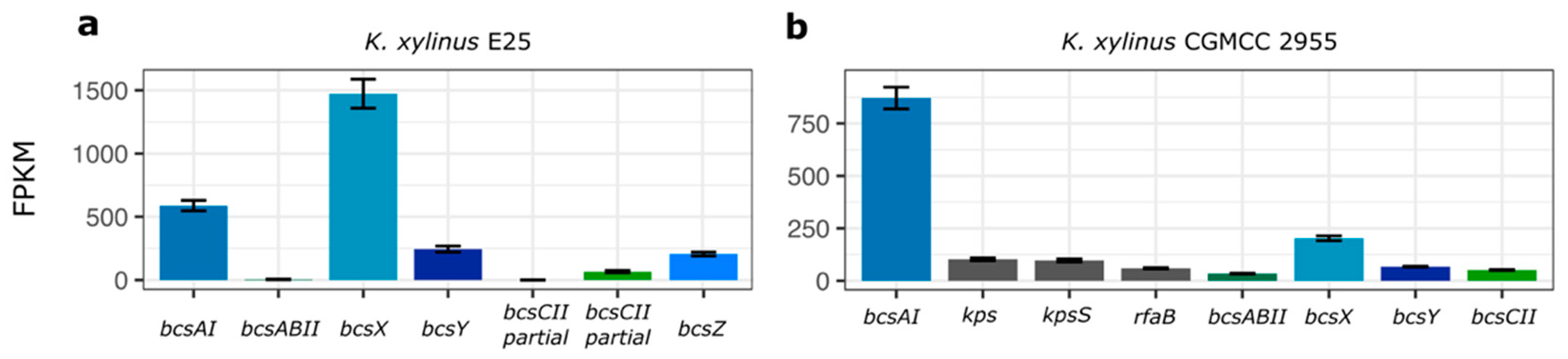
2.2. Expression Profile of the BcsII Operon
2.3. Function Prediction, Sequence Similarity and Homology Search for the BcsII Proteins
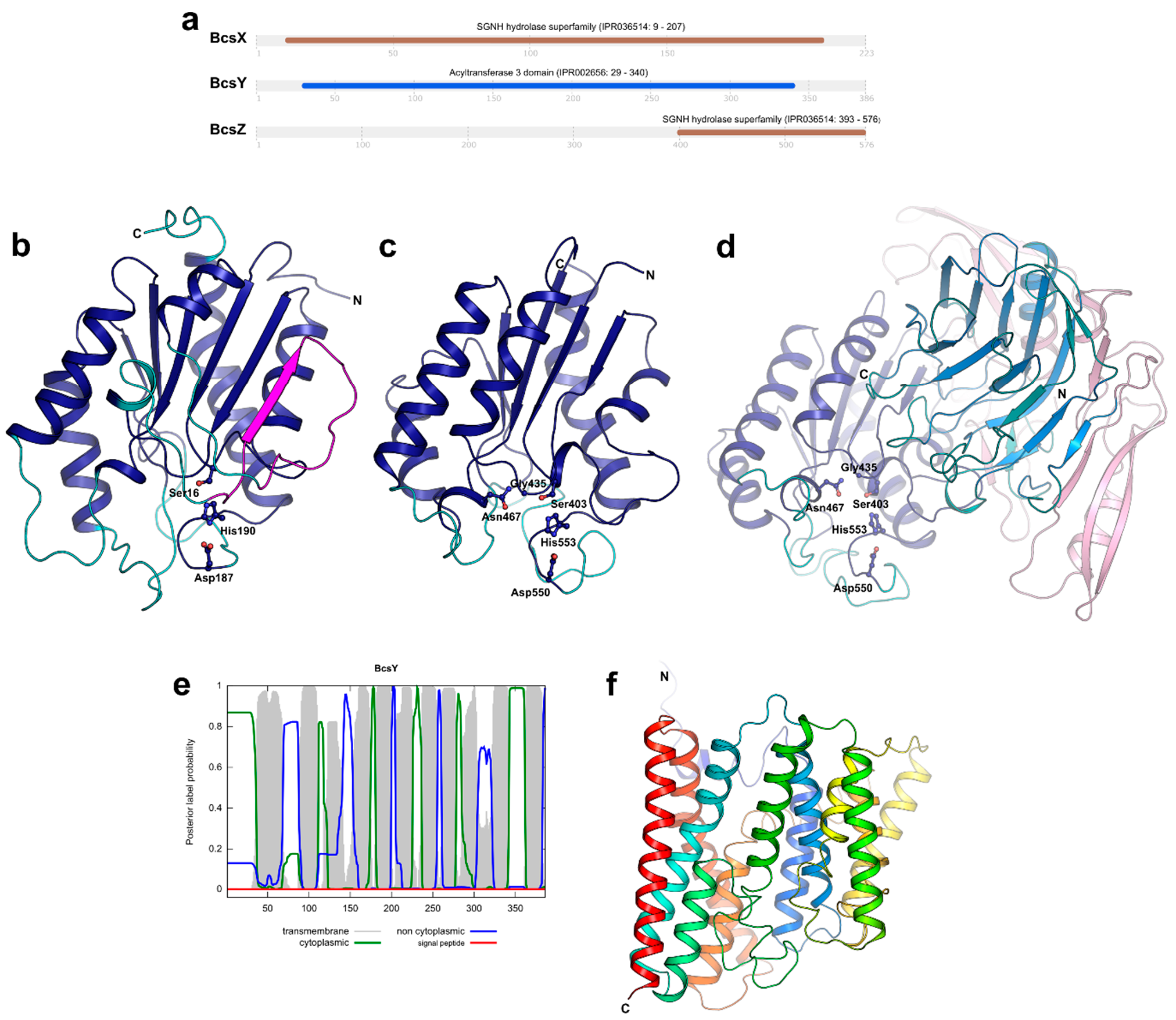
2.4. Prediction of BcsX, BcsY and BcsZ Subcellular Localization
2.5. Three-Dimensional Structure Prediction for BcsX, BcsY and BcsZ
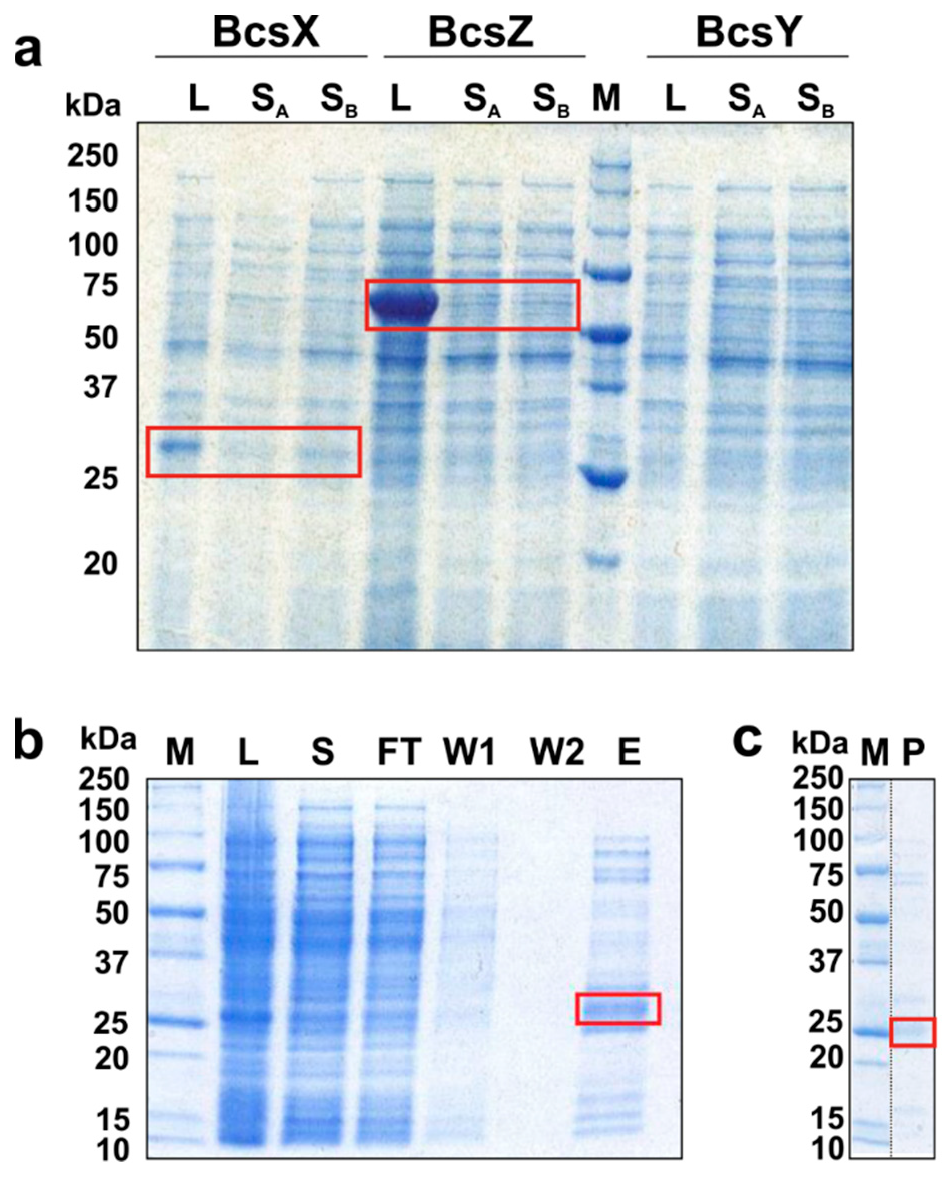
2.6. Overexpression and Solubility Tests of the BcsII Operon Proteins, BcsX Purification and Crystallization
3. Materials and Methods
3.1. Bioinformatical Analysis of Genome and Transcriptome Data
3.2. Functional and Structural Prediction
3.3. Overexpression and Solubility Tests of the BcsII Operon Proteins
3.4. Production and Purification of BcsX Protein
3.5. Protein Crystallization and Data Collection
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Szustak, M.; Gendaszewska-Darmach, E. Nanocellulose-Based Scaffolds for Chondrogenic Differentiation and Expansion. Front. Bioeng. Biotechnol. 2021, 9, 733. [Google Scholar] [CrossRef] [PubMed]
- Ludwicka, K.; Kaczmarek, M.; Białkowska, A. Bacterial Nanocellulose—A Biobased Polymer for Active and Intelligent Food Packaging Applications: Recent Advances and Developments. Polymers 2020, 12, 2209. [Google Scholar] [CrossRef] [PubMed]
- Gorgieva, S.; Trček, J. Bacterial Cellulose: Production, Modification and Perspectives in Biomedical Applications. Nanomaterials 2019, 9, 1352. [Google Scholar] [CrossRef] [Green Version]
- Jankau, J.; Błażyńska-Spychalska, A.; Kubiak, K.; Jędrzejczak-Krzepkowska, M.; Pankiewicz, T.; Ludwicka, K.; Dettlaff, A.; Pęksa, R. Bacterial Cellulose Properties Fulfilling Requirements for a Biomaterial of Choice in Reconstructive Surgery and Wound Healing. Front. Bioeng. Biotechnol. 2022, 9, 1492. [Google Scholar] [CrossRef]
- Dourado, F.; Ryngajllo, M.; Jedrzejczak-Krzepkowska, M.; Bielecki, S.; Gama, M. Taxonomic Review and Microbial Ecology in Bacterial NanoCellulose Fermentation. Bact. Nanocellul. 2016, 1–17. [Google Scholar]
- Ryngajłło, M.; Jędrzejczak-Krzepkowska, M.; Kubiak, K.; Ludwicka, K.; Bielecki, S. Towards Control of Cellulose Biosynthesis by Komagataeibacter Using Systems-Level and Strain Engineering Strategies: Current Progress and Perspectives. Appl. Microbiol. Biotechnol. 2020, 104, 6565–6585. [Google Scholar] [CrossRef]
- Ryngajłło, M.; Kubiak, K.; Jędrzejczak-Krzepkowska, M.; Jacek, P.; Bielecki, S. Comparative Genomics of the Komagataeibacter Strains—Efficient Bionanocellulose Producers. Microbiologyopen 2019, 8, e00731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicolas, W.J.; Ghosal, D.; Tocheva, E.I.; Meyerowitz, E.M.; Jensen, G.J. Structure of the Bacterial Cellulose Ribbon and Its Assembly-Guiding Cytoskeleton by Electron Cryotomography. J. Bacteriol. 2021, 203, e00371-20. [Google Scholar] [CrossRef]
- Cielecka, I.; Ryngajłło, M.; Maniukiewicz, W.; Bielecki, S. Response Surface Methodology-Based Improvement of the Yield and Differentiation of Properties of Bacterial Cellulose by Metabolic Enhancers. Int. J. Biol. Macromol. 2021, 187, 584–593. [Google Scholar] [CrossRef]
- Gullo, M.; La China, S.; Falcone, P.M.; Giudici, P. Biotechnological Production of Cellulose by Acetic Acid Bacteria: Current State and Perspectives. Appl. Microbiol. Biotechnol. 2018, 102, 6885–6898. [Google Scholar] [CrossRef]
- Valera, M.J.; Mas, A.; Streit, W.R.; Mateo, E. GqqA, a Novel Protein in Komagataeibacter Europaeus Involved in Bacterial Quorum Quenching and Cellulose Formation. Microb. Cell Fact. 2016, 15, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsutani, M.; Ito, K.; Azuma, Y.; Ogino, H.; Shirai, M.; Yakushi, T.; Matsushita, K. Adaptive Mutation Related to Cellulose Producibility in Komagataeibacter Medellinensis (Gluconacetobacter Xylinus) NBRC 3288. Appl. Microbiol. Biotechnol. 2015, 99, 7229–7240. [Google Scholar] [CrossRef] [PubMed]
- Ishiya, K.; Kosaka, H.; Inaoka, T.; Kimura, K.; Nakashima, N. Comparative Genome Analysis of Three Komagataeibacter Strains Used for Practical Production of Nata-de-Coco. Front. Microbiol. 2022, 12, 4308. [Google Scholar] [CrossRef] [PubMed]
- Jedrzejczak-Krzepkowska, M.; Kubiak, K.; Ludwicka, K.; Bielecki, S. Chapter 2 – Bacterial NanoCellulose Synthesis, Recent Findings. In Bacterial Nanocellulose: From Biotechnology to Bio-Economy; Gama, M., Dourado, F., Bielecki, S., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 19–46. ISBN 9780444634580. [Google Scholar]
- Uto, T.; Ikeda, Y.; Sunagawa, N.; Tajima, K.; Yao, M.; Yui, T. Molecular Dynamics Simulation of Cellulose Synthase Subunit D Octamer with Cellulose Chains from Acetic Acid Bacteria: Insight into Dynamic Behaviors and Thermodynamics on Substrate Recognition. J. Chem. Theory Comput. 2021, 17, 488–496. [Google Scholar] [CrossRef] [PubMed]
- Römling, U.; Galperin, M.Y. Bacterial Cellulose Biosynthesis: Diversity of Operons, Subunits, Products, and Functions. Trends Microbiol. 2015, 23, 545–557. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Arriaga, A.M.; del Cerro, C.; Urbina, L.; Eceiza, A.; Corcuera, M.A.; Retegi, A.; Auxiliadora Prieto, M. Genome Sequence and Characterization of the Bcs Clusters for the Production of Nanocellulose from the Low PH Resistant Strain Komagataeibacter Medellinensis ID13488. Microb. Biotechnol. 2019, 12, 620–632. [Google Scholar] [CrossRef] [Green Version]
- Brandão, P.R.; Crespo, M.T.B.; Nascimento, F.X. Phylogenomic and Comparative Analyses Support the Reclassification of Several Komagataeibacter Species as Novel Members of the Novacetimonas Gen. Nov. and Bring New Insights into the Evolution of Cellulose Synthase Genes. Int. J. Syst. Evol. Microbiol. 2022, 72, 5252. [Google Scholar] [CrossRef]
- Bimmer, M.; Mientus, M.; Klingl, A.; Ehrenreich, A.; Liebl, W. The Roles of the Various Cellulose Biosynthesis Operons in Komagataeibacter Hansenii ATCC 23769. Appl. Environ. Microbiol. 2022, 88, e02460-21. [Google Scholar] [CrossRef]
- Florea, M.; Reeve, B.; Abbott, J.; Freemont, P.S.; Ellis, T. Genome Sequence and Plasmid Transformation of the Model High-Yield Bacterial Cellulose Producer Gluconacetobacter Hansenii ATCC 53582. Sci. Rep. 2016, 6, 23635. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Xu, X.; Chen, X.; Yuan, F.; Sun, B.; Xu, Y.; Yang, J.; Sun, D. Complete Genome Sequence of the Cellulose-Producing Strain Komagataeibacter Nataicola RZS01. Sci. Rep. 2017, 7, 4431. [Google Scholar] [CrossRef] [Green Version]
- Umeda, Y.; Hirano, A.; Ishibashi, M.; Akiyama, H.; Onizuka, T.; Ikeuchi, M.; Inoue, Y. Cloning of Cellulose Synthase Genes from Acetobacter Xylinum JCM 7664: Implication of a Novel Set of Cellulose Synthase Genes. DNA Res. 1999, 6, 109–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coucheron, D.H. A Family of IS1031 Elements in the Genome of Acetobacter Xylinum: Nucleotide Sequences and Strain Distribution. Mol. Microbiol. 1993, 9, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Gopu, G.; Govindan, S. Production of Bacterial Cellulose from Komagataeibacter Saccharivorans Strain BC1 Isolated from Rotten Green Grapes. Prep. Biochem. Biotechnol. 2018, 48, 842–852. [Google Scholar] [CrossRef] [PubMed]
- Abol-Fotouh, D.; Hassan, M.A.; Shokry, H.; Roig, A.; Azab, M.S.; Kashyout, A.E.H.B. Bacterial Nanocellulose from Agro-Industrial Wastes: Low-Cost and Enhanced Production by Komagataeibacter Saccharivorans MD1. Sci. Rep. 2020, 10, 3491. [Google Scholar] [CrossRef] [Green Version]
- Ryngajłło, M.; Jacek, P.; Cielecka, I.; Kalinowska, H.; Bielecki, S. Effect of Ethanol Supplementation on the Transcriptional Landscape of Bionanocellulose Producer Komagataeibacter Xylinus E25. Appl. Microbiol. Biotechnol. 2019, 103, 6673–6688. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.H.; Liu, Q.J.; Sun, X.W.; Li, X.J.; Liu, M.; Jia, S.R.; Xie, Y.Y.; Zhong, C. Tailoring Bacterial Cellulose Structure through CRISPR Interference-Mediated Downregulation of GalU in Komagataeibacter Xylinus CGMCC 2955. Biotechnol. Bioeng. 2020, 117, 2165–2176. [Google Scholar] [CrossRef]
- Spiers, A.J.; Bohannon, J.; Gehrig, S.M.; Rainey, P.B. Biofilm Formation at the Air-Liquid Interface by the Pseudomonas Fluorescens SBW25 Wrinkly Spreader Requires an Acetylated Form of Cellulose. Mol. Microbiol. 2003, 50, 15–27. [Google Scholar] [CrossRef]
- Kruis, A.J.; Bohnenkamp, A.C.; Patinios, C.; van Nuland, Y.M.; Levisson, M.; Mars, A.E.; van den Berg, C.; Kengen, S.W.M.; Weusthuis, R.A. Microbial Production of Short and Medium Chain Esters: Enzymes, Pathways, and Applications. Biotechnol. Adv. 2019, 37, 107407. [Google Scholar] [CrossRef]
- Park, Y.C.; Shaffer, C.E.H.; Bennett, G.N. Microbial Formation of Esters. Appl. Microbiol. Biotechnol. 2009, 85, 13–25. [Google Scholar] [CrossRef]
- PDBeFold (SSM). Available online: http://www.ebi.ac.uk/msd-srv/ssm/ (accessed on 2 June 2022).
- Kim, K.; Ryu, B.H.; Kim, S.S.; An, D.R.; Ngo, T.D.; Pandian, R.; Kim, K.K.; Kim, T.D. Structural and Biochemical Characterization of a Carbohydrate Acetylesterase from Sinorhizobium Meliloti 1021. FEBS Lett. 2015, 589, 117–122. [Google Scholar] [CrossRef] [Green Version]
- Mathews, I.; Soltis, M.; Saldajeno, M.; Ganshaw, G.; Sala, R.; Weyler, W.; Cervin, M.A.; Whited, G.; Bott, R. Structure of a Novel Enzyme That Catalyzes Acyl Transfer to Alcohols in Aqueous Conditions. Biochemistry 2007, 46, 8969–8979. [Google Scholar] [CrossRef] [PubMed]
- Kovačić, F.; Granzin, J.; Wilhelm, S.; Kojić-Prodić, B.; Batra-Safferling, R.; Jaeger, K.E. Structural and Functional Characterisation of TesA - a Novel Lysophospholipase A from Pseudomonas Aeruginosa. PLoS ONE 2013, 8, e69125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brzuszkiewicz, A.; Nowak, E.; Dauter, Z.; Dauter, M.; Cieśliski, H.; Dlługolłȩcka, A.; Kur, J. Structure of EstA Esterase from Psychrotrophic Pseudoalteromonas Sp. 643A Covalently Inhibited by Monoethylphosphonate. Acta Crystallogr. Sect. F. Struct. Biol. Cryst. Commun. 2009, 65, 862–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo, Y.C.; Lin, S.C.; Shaw, J.F.; Liaw, Y.C. Crystal Structure of Escherichia Coli Thioesterase I/Protease I/Lysophospholipase L1: Consensus Sequence Blocks Constitute the Catalytic Center of SGNH-Hydrolases through a Conserved Hydrogen Bond Network. J. Mol. Biol. 2003, 330, 539–551. [Google Scholar] [CrossRef]
- Watanabe, M.; Fukada, H.; Inoue, H.; Ishikawa, K. Crystal Structure of an Acetylesterase from Talaromyces Cellulolyticus and the Importance of a Disulfide Bond near the Active Site. FEBS Lett. 2015, 589, 1200–1206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, K.; Duquette, M.; Liu, J.H.; Zhang, R.; Joachimiak, A.; Wang, J.H.; Lawler, J. The Structures of the Thrombospondin-1 N-Terminal Domain and Its Complex with a Synthetic Pentameric Heparin. Structure 2006, 14, 33. [Google Scholar] [CrossRef] [Green Version]
- Geethanandan, K.; Abhilash, J.; Bharath, S.R.; Sadasivan, C.; Haridas, M. X-Ray Structure of a Galactose-Specific Lectin from Spatholobous Parviflorous. Int. J. Biol. Macromol. 2011, 49, 992–998. [Google Scholar] [CrossRef]
- Hamelryck, T.W.; Loris, R.; Bouckaert, J.; Dao-Thi, M.H.; Strecker, G.; Imberty, A.; Fernandez, E.; Wyns, L.; Etzler, M.E. Carbohydrate Binding, Quaternary Structure and a Novel Hydrophobic Binding Site in Two Legume Lectin Oligomers from Dolichos Biflorus. J. Mol. Biol. 1999, 286, 1161–1177. [Google Scholar] [CrossRef] [Green Version]
- Audette, G.F.; Vandonselaar, M.; Delbaere, L.T.J. The 2.2 A Resolution Structure of the O(H) Blood-Group-Specific Lectin I from Ulex Europaeus. J. Mol. Biol. 2000, 304, 423–433. [Google Scholar] [CrossRef] [Green Version]
- Delbaere, L.T.J.; Vandonselaar, M.; Prasad, L.; Quail, J.W.; Wilson, K.S.; Dauter, Z. Structure of the Lectin IV of Griffonia Simplicifolia and Its Complex with the Lewis b Human Blood Group Determinant at 2·0 Å Resolution. J. Mol. Biol. 1993, 230, 950–965. [Google Scholar] [CrossRef]
- Lis, H.; Sharon, N. Lectins: Carbohydrate-Specific Proteins That Mediate Cellular Recognition. Chem. Rev. 1998, 98, 637–674. [Google Scholar] [CrossRef] [PubMed]
- Hirata, K.; Shinzawa-Itoh, K.; Yano, N.; Takemura, S.; Kato, K.; Hatanaka, M.; Muramoto, K.; Kawahara, T.; Tsukihara, T.; Yamashita, E.; et al. Determination of Damage-Free Crystal Structure of an X-Ray-Sensitive Protein Using an XFEL. Nat. Methods 2014, 11, 734–736. [Google Scholar] [CrossRef] [PubMed]
- Gordiyenko, Y.; Llácer, J.L.; Ramakrishnan, V. Structural Basis for the Inhibition of Translation through EIF2α Phosphorylation. Nat. Commun. 2019, 10, 2640. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- InterProScan. Available online: https://www.ebi.ac.uk/interpro/search/sequence/ (accessed on 9 December 2021).
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and Sensitive Protein Alignment Using DIAMOND. Nat. Methods 2014 121 2014, 12, 59–60. [Google Scholar] [CrossRef] [PubMed]
- NCBI Blastp. Available online: https://blast.ncbi.nlm.nih.gov/blast.Cgi?PAGE=Proteins (accessed on 12 June 2022).
- Teufel, F.; Almagro Armenteros, J.J.; Johansen, A.R.; Gíslason, M.H.; Pihl, S.I.; Tsirigos, K.D.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 6.0 Predicts All Five Types of Signal Peptides Using Protein Language Models. Nat. Biotechnol. 2022, 40, 1023–1025. [Google Scholar] [CrossRef] [PubMed]
- Käll, L.; Krogh, A.; Sonnhammer, E.L.L. A Combined Transmembrane Topology and Signal Peptide Prediction Method. J. Mol. Biol. 2004, 338, 1027–1036. [Google Scholar] [CrossRef] [PubMed]
- Käll, L.; Krogh, A.; Sonnhammer, E.L.L. Advantages of Combined Transmembrane Topology and Signal Peptide Prediction-the Phobius Web Server. Nucleic Acids Res. 2007, 35, W429–W432. [Google Scholar] [CrossRef] [Green Version]
- Tsirigos, K.D.; Elofsson, A.; Bagos, P.G. PRED-TMBB2: Improved Topology Prediction and Detection of Beta-Barrel Outer Membrane Proteins. Bioinformatics 2016, 32, i665–i671. [Google Scholar] [CrossRef] [Green Version]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Pietrzyk-Brzezinska, A.J.; Absmeier, E.; Klauck, E.; Wen, Y.; Antelmann, H.; Wahl, M.C. Crystal Structure of the Escherichia Coli DExH-Box NTPase HrpB. Structure 2018, 26, 1462–1473. [Google Scholar] [CrossRef] [Green Version]
- Pietrzyk-Brzezinska, A.J.; Cociurovscaia, A. Structures of the TetR-like Transcription Regulator RcdA Alone and in Complexes with Ligands. Proteins Struct. Funct. Bioinforma. 2022, 90, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Mueller, U.; Förster, R.; Hellmig, M.; Huschmann, F.U.; Kastner, A.; Malecki, P.; Pühringer, S.; Röwer, M.; Sparta, K.; Steffien, M.; et al. The Macromolecular Crystallography Beamlines at BESSY II of the Helmholtz-Zentrum Berlin: Current Status and Perspectives. Eur. Phys. J. Plus 2015, 130. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PDB ID | Enzyme | Rmsd (Å) | Cα Atoms Aligned | Reference (Homolog Structure) |
|---|---|---|---|---|
| Structural homologs of BcsX | ||||
| 4TX1 | Sinorhizobium meliloti carbohydrate acetylesterase | 2.63 | 171 | [32] |
| 3DCI | Agrobacterium tumefaciens putative arylesterase | 2.67 | 168 | n/a |
| 2Q0Q | Mycobacterium smegmatis serine esterase | 2.47 | 160 | [33] |
| 4JGG | Pseudomonas aeruginosa serine esterase | 2.57 | 154 | [34] |
| 3HP4 | Pseudoalteromonas sp. 643A serine esterase | 2.31 | 148 | [35] |
| Structural homologs of BcsZ-CTD | ||||
| 1IVN | Escherichia coli multifunctional enzyme (activities of thioesterase, esterase, arylesterase, protease and lysophospholipase) | 1.92 | 147 | [36] |
| 3P94 | Parabacteroides distasonis GDSL-like lipase | 1.94 | 151 | n/a |
| 5B5S | Talaromyces cellulolyticus carbohydrate esterase | 2.01 | 152 | [37] |
| 4TX1 | Sinorhizobium meliloti carbohydrate acetylesterase | 2.36 | 159 | [32] |
| 2Q0Q | Mycobacterium smegmatis serine esterase | 2.10 | 151 | [33] |
| Structural homologs of BcsZ middle part (residues 192–382) | ||||
| 1Z78 | Homo sapiens thrombospondin-NTD | 3.21 | 163 | [38] |
| 3IPV | Spatholobus parviflorus seed lectin | 3.39 | 165 | [39] |
| 1LU1 | Vigna unguiculata seed lectin | 3.42 | 161 | [40] |
| 1FX5 | Ulex europaeus lectin I | 3.15 | 149 | [41] |
| 1GSL | Griffonia simplicifolia lectin | 3.24 | 150 | [42] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szymczak, I.; Pietrzyk-Brzezińska, A.J.; Duszyński, K.; Ryngajłło, M. Characterization of the Putative Acylated Cellulose Synthase Operon in Komagataeibacter xylinus E25. Int. J. Mol. Sci. 2022, 23, 7851. https://doi.org/10.3390/ijms23147851
Szymczak I, Pietrzyk-Brzezińska AJ, Duszyński K, Ryngajłło M. Characterization of the Putative Acylated Cellulose Synthase Operon in Komagataeibacter xylinus E25. International Journal of Molecular Sciences. 2022; 23(14):7851. https://doi.org/10.3390/ijms23147851
Chicago/Turabian StyleSzymczak, Izabela, Agnieszka J. Pietrzyk-Brzezińska, Kajetan Duszyński, and Małgorzata Ryngajłło. 2022. "Characterization of the Putative Acylated Cellulose Synthase Operon in Komagataeibacter xylinus E25" International Journal of Molecular Sciences 23, no. 14: 7851. https://doi.org/10.3390/ijms23147851