
Antibodies Regulate Dual-Function Enzyme IYD to Induce Functional Synergy between Metabolism and Thermogenesis



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. H3 Ab Induces Weight Gain
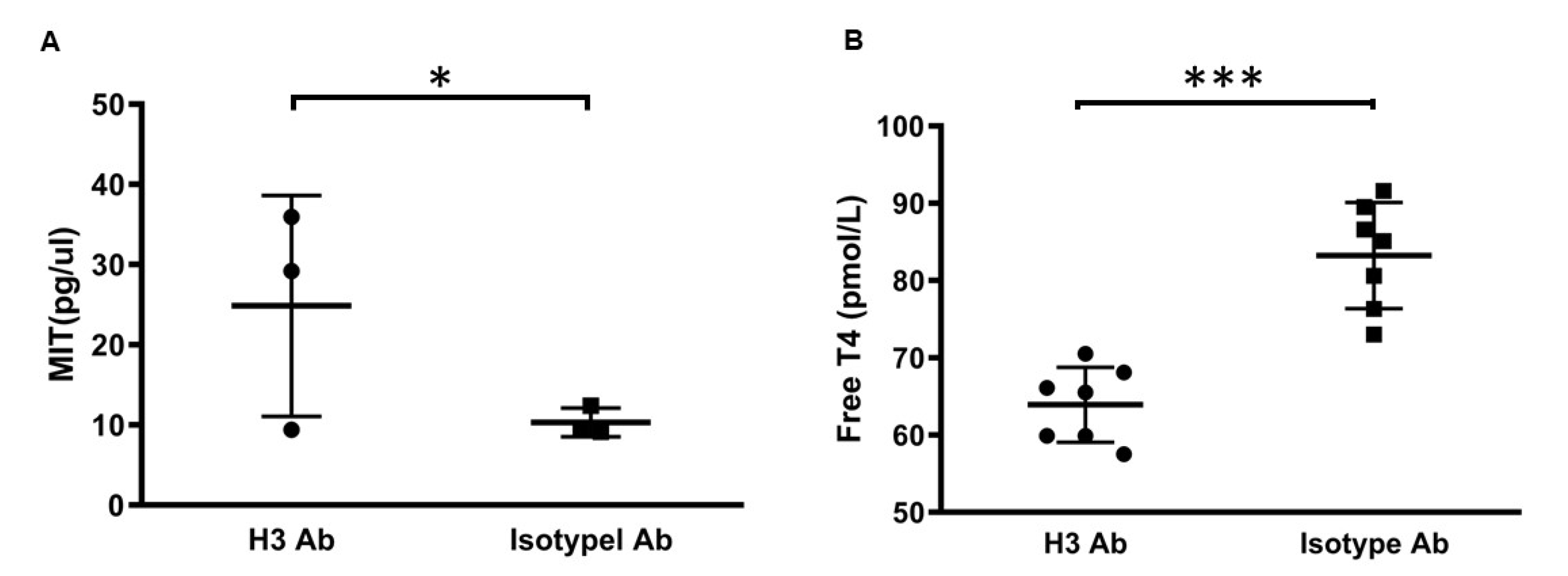
2.2. H3 Ab Controls Thyroid Hormone
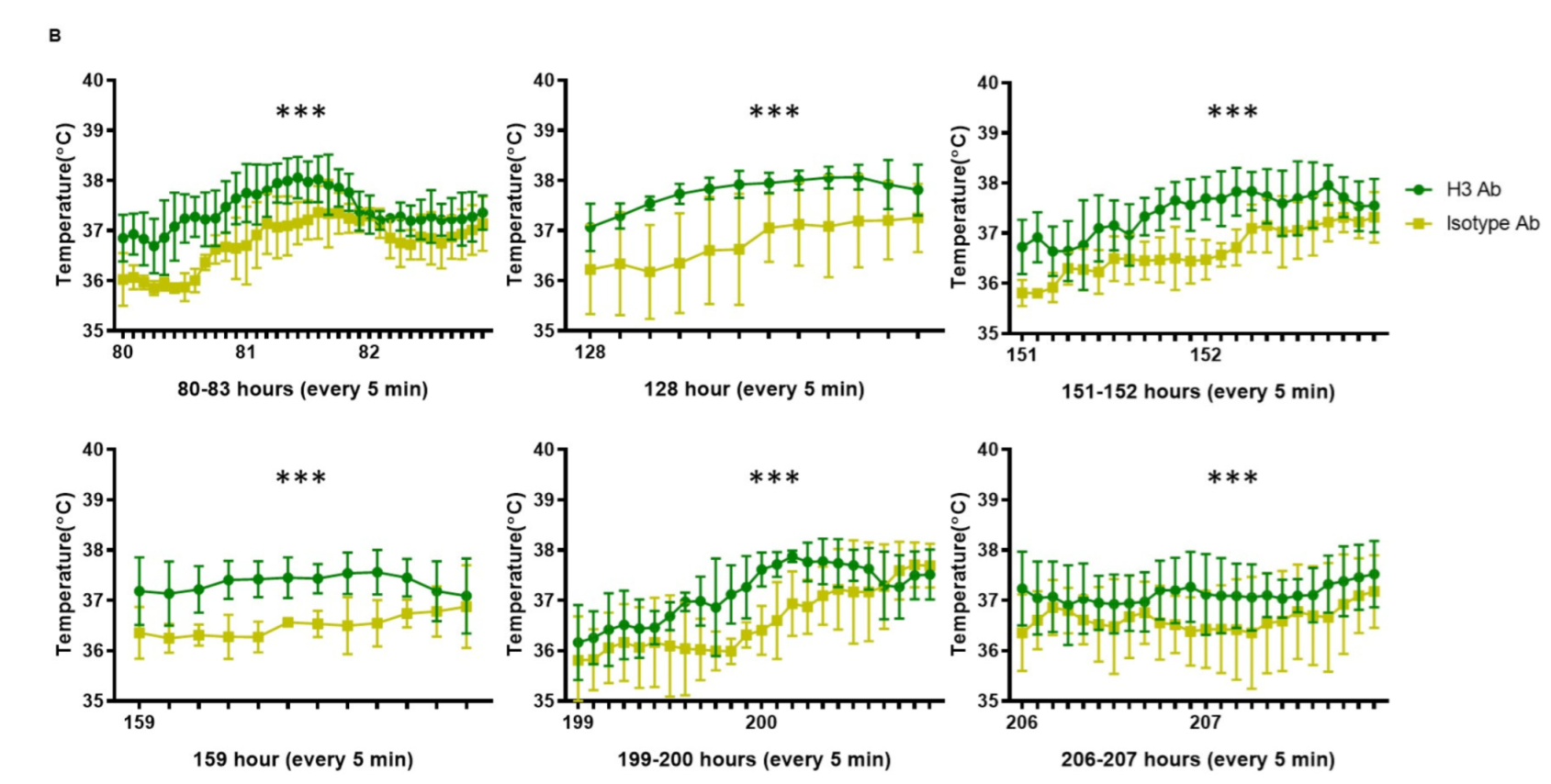
2.3. H3 Ab Increases Core Body Temperature
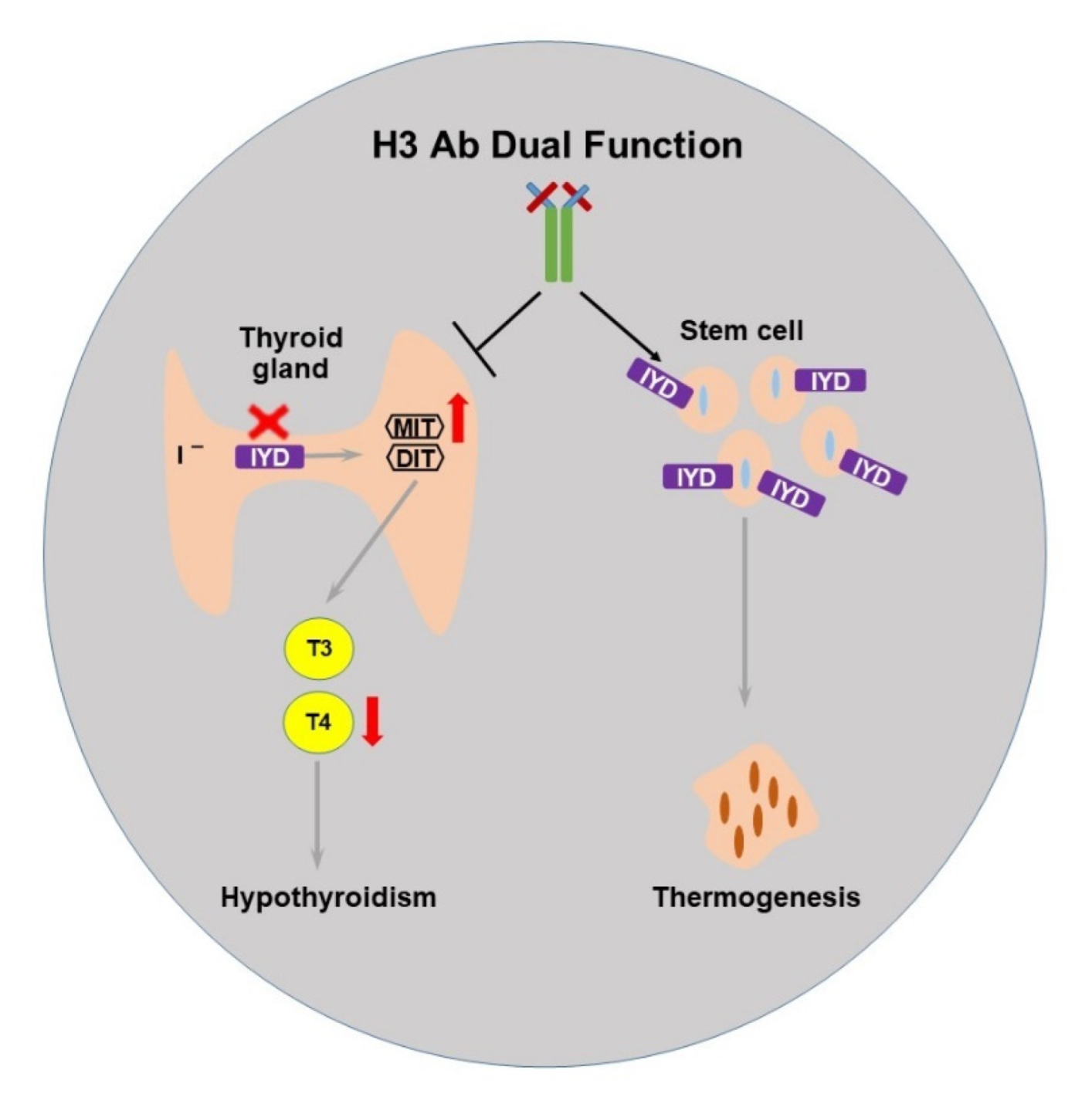
2.4. The Selected H3 Antibody Has Dual Function
3. Discussion
4. Materials and Methods
4.1. Study Design
4.2. Mice and Cell Lines
4.3. Telemetry
4.4. Treatment with Isolated Antibody
4.5. MIT Assay
4.6. Free T4 Assay
4.7. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sociale, M.; Wulf, A.L.; Breiden, B.; Klee, K.; Thielisch, M.; Eckardt, F.; Sellin, J.; Bulow, M.H.; Lobbert, S.; Weinstock, N.; et al. Ceramide Synthase Schlank Is a Transcriptional Regulator Adapting Gene Expression to Energy Requirements. Cell Rep. 2018, 22, 967–978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallen, A.; Cooper, A.J.; Jamie, J.F.; Karuso, P. Insights into Enzyme Catalysis and Thyroid Hormone Regulation of Cerebral Ketimine Reductase/mu-Crystallin Under Physiological Conditions. Neurochem. Res. 2015, 40, 1252–1266. [Google Scholar] [CrossRef] [PubMed]
- Kuo, Y.W.; Trottier, O.; Mahamdeh, M.; Howard, J. Spastin is a dual-function enzyme that severs microtubules and promotes their regrowth to increase the number and mass of microtubules. Proc. Natl. Acad. Sci. USA 2019, 116, 5533–5541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geoghegan, J.C.; Diedrich, G.; Lu, X.; Rosenthal, K.; Sachsenmeier, K.F.; Wu, H.; Dall’Acqua, W.F.; Damschroder, M.M. Inhibition of CD73 AMP hydrolysis by a therapeutic antibody with a dual, non-competitive mechanism of action. MAbs 2016, 8, 454–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, K.H.; Arlian, B.M.; Lin, C.W.; Jin, H.Y.; Kang, G.H.; Lee, S.; Lee, P.C.; Lerner, R.A. Agonist Antibody Converts Stem Cells into Migrating Brown Adipocyte-Like Cells in Heart. Cells 2020, 9, 256. [Google Scholar] [CrossRef] [Green Version]
- Han, K.H.; Arlian, B.M.; Macauley, M.S.; Paulson, J.C.; Lerner, R.A. Migration-based selections of antibodies that convert bone marrow into trafficking microglia-like cells that reduce brain amyloid beta. Proc. Natl. Acad. Sci. USA 2018, 115, E372–E381. [Google Scholar] [CrossRef] [Green Version]
- Han, K.H.; Gonzalez-Quintial, R.; Peng, Y.; Baccala, R.; Theofilopoulos, A.N.; Lerner, R.A. An agonist antibody that blocks autoimmunity by inducing anti-inflammatory macrophages. FASEB J. 2016, 30, 738–747. [Google Scholar] [CrossRef] [Green Version]
- Guler, R.; Thatikonda, N.; Ghani, H.A.; Hedhammar, M.; Lofblom, J. VEGFR2-Specific Ligands Based on Affibody Molecules Demonstrate Agonistic Effects when Tetrameric in the Soluble Form or Immobilized via Spider Silk. ACS Biomater. Sci. Eng. 2019, 5, 6474–6484. [Google Scholar] [CrossRef] [Green Version]
- Molina, M.F.; Papendieck, P.; Sobrero, G.; Balbi, V.A.; Belforte, F.S.; Martinez, E.B.; Adrover, E.; Olcese, M.C.; Chiesa, A.; Miras, M.B.; et al. Mutational screening of the TPO and DUOX2 genes in Argentinian children with congenital hypothyroidism due to thyroid dyshormonogenesis. Endocrine 2022, 77, 86–101. [Google Scholar] [CrossRef]
- Olker, J.H.; Korte, J.J.; Haselman, J.T.; Hornung, M.W.; Degitz, S.J. Cross-species comparison of chemical inhibition of human and Xenopus iodotyrosine deiodinase. Aquat. Toxicol. 2022, 249, 106227. [Google Scholar] [CrossRef]
- Chaker, L.; Bianco, A.C.; Jonklaas, J.; Peeters, R.P. Hypothyroidism. Lancet 2017, 390, 1550–1562. [Google Scholar] [CrossRef]
- Virili, C.; Antonelli, A.; Santaguida, M.G.; Benvenga, S.; Centanni, M. Gastrointestinal Malabsorption of Thyroxine. Endocr. Rev. 2019, 40, 118–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burniat, A.; Pirson, I.; Vilain, C.; Kulik, W.; Afink, G.; Moreno-Reyes, R.; Corvilain, B.; Abramowicz, M. Iodotyrosine deiodinase defect identified via genome-wide approach. J. Clin. Endocrinol. Metab. 2012, 97, E1276–E1283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Zhang, X.; Jiang, Y.; Bao, S.; Shan, Z.; Teng, W. Expression of Iodotyrosine Deiodinase in Thyroid and Other Organs in Iodine-Deficient and Iodine-Excess Rats. Biol. Trace Elem. Res. 2015, 167, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.Y.; Leonard, J.L.; Davis, P.J. Molecular aspects of thyroid hormone actions. Endocr. Rev. 2010, 31, 139–170. [Google Scholar] [CrossRef] [Green Version]
- Mullur, R.; Liu, Y.Y.; Brent, G.A. Thyroid hormone regulation of metabolism. Physiol. Rev. 2014, 94, 355–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritter, M.J.; Amano, I.; Hollenberg, A.N. Thyroid Hormone Signaling and the Liver. Hepatology 2020, 72, 742–752. [Google Scholar] [CrossRef]
- Rousset, B.; Dupuy, C.; Miot, F.; Dumont, F. Chapter 2 Thyroid Hormone Synthesis And Secretion; MDText.com, Inc.: South Dartmouth, MA, USA, 2000. [Google Scholar]
- Chaker, L.; Razvi, S.; Bensenor, I.M.; Azizi, F.; Pearce, E.N.; Peeters, R.P. Hypothyroidism. Nat. Rev. Dis. Primers 2022, 8, 30. [Google Scholar] [CrossRef]
- Hegedus, L.; Bianco, A.C.; Jonklaas, J.; Pearce, S.H.; Weetman, A.P.; Perros, P. Primary hypothyroidism and quality of life. Nat. Rev. Endocrinol. 2022, 18, 230–242. [Google Scholar] [CrossRef] [PubMed]
- Wade, G.; McGahee, A.; Ntambi, J.M.; Simcox, J. Lipid Transport in Brown Adipocyte Thermogenesis. Front. Physiol. 2021, 12, 787535. [Google Scholar] [CrossRef] [PubMed]
- Cohen, P.; Kajimura, S. The cellular and functional complexity of thermogenic fat. Nat. Rev. Mol. Cell Biol. 2021, 22, 393–409. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Avery, J.; Yin, A.; Singh, A.M.; Cliff, T.S.; Yin, H.; Dalton, S. Generation of Functional Brown Adipocytes from Human Pluripotent Stem Cells via Progression through a Paraxial Mesoderm State. Cell Stem Cell 2020, 27, 784–797.e711. [Google Scholar] [CrossRef] [PubMed]
- Shao, M.; Wang, Q.A.; Song, A.; Vishvanath, L.; Busbuso, N.C.; Scherer, P.E.; Gupta, R.K. Cellular Origins of Beige Fat Cells Revisited. Diabetes 2019, 68, 1874–1885. [Google Scholar] [CrossRef] [PubMed]
- Min, S.Y.; Desai, A.; Yang, Z.; Sharma, A.; DeSouza, T.; Genga, R.M.J.; Kucukural, A.; Lifshitz, L.M.; Nielsen, S.; Scheele, C.; et al. Diverse repertoire of human adipocyte subtypes develops from transcriptionally distinct mesenchymal progenitor cells. Proc. Natl. Acad. Sci. USA 2019, 116, 17970–17979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuko, O.O.; Saito, M. Brown Fat as a Regulator of Systemic Metabolism beyond Thermogenesis. Diabetes Metab. J. 2021, 45, 840–852. [Google Scholar] [CrossRef]
- Afink, G.; Kulik, W.; Overmars, H.; de Randamie, J.; Veenboer, T.; van Cruchten, A.; Craen, M.; Ris-Stalpers, C. Molecular characterization of iodotyrosine dehalogenase deficiency in patients with hypothyroidism. J. Clin. Endocrinol. Metab. 2008, 93, 4894–4901. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, S.; Park, H.-W.; Han, K.H. Antibodies Regulate Dual-Function Enzyme IYD to Induce Functional Synergy between Metabolism and Thermogenesis. Int. J. Mol. Sci. 2022, 23, 7834. https://doi.org/10.3390/ijms23147834
Kang S, Park H-W, Han KH. Antibodies Regulate Dual-Function Enzyme IYD to Induce Functional Synergy between Metabolism and Thermogenesis. International Journal of Molecular Sciences. 2022; 23(14):7834. https://doi.org/10.3390/ijms23147834
Chicago/Turabian StyleKang, Sunghyun, Hwan-Woo Park, and Kyung Ho Han. 2022. "Antibodies Regulate Dual-Function Enzyme IYD to Induce Functional Synergy between Metabolism and Thermogenesis" International Journal of Molecular Sciences 23, no. 14: 7834. https://doi.org/10.3390/ijms23147834