
RNA-Seq Provides Insights into VEGF-Induced Signaling in Human Retinal Microvascular Endothelial Cells: Implications in Retinopathy of Prematurity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Quality Control of Biologic Samples in RNA-Seq Analysis
2.2. VEGF-Mediated Gene Regulation in HRMECs
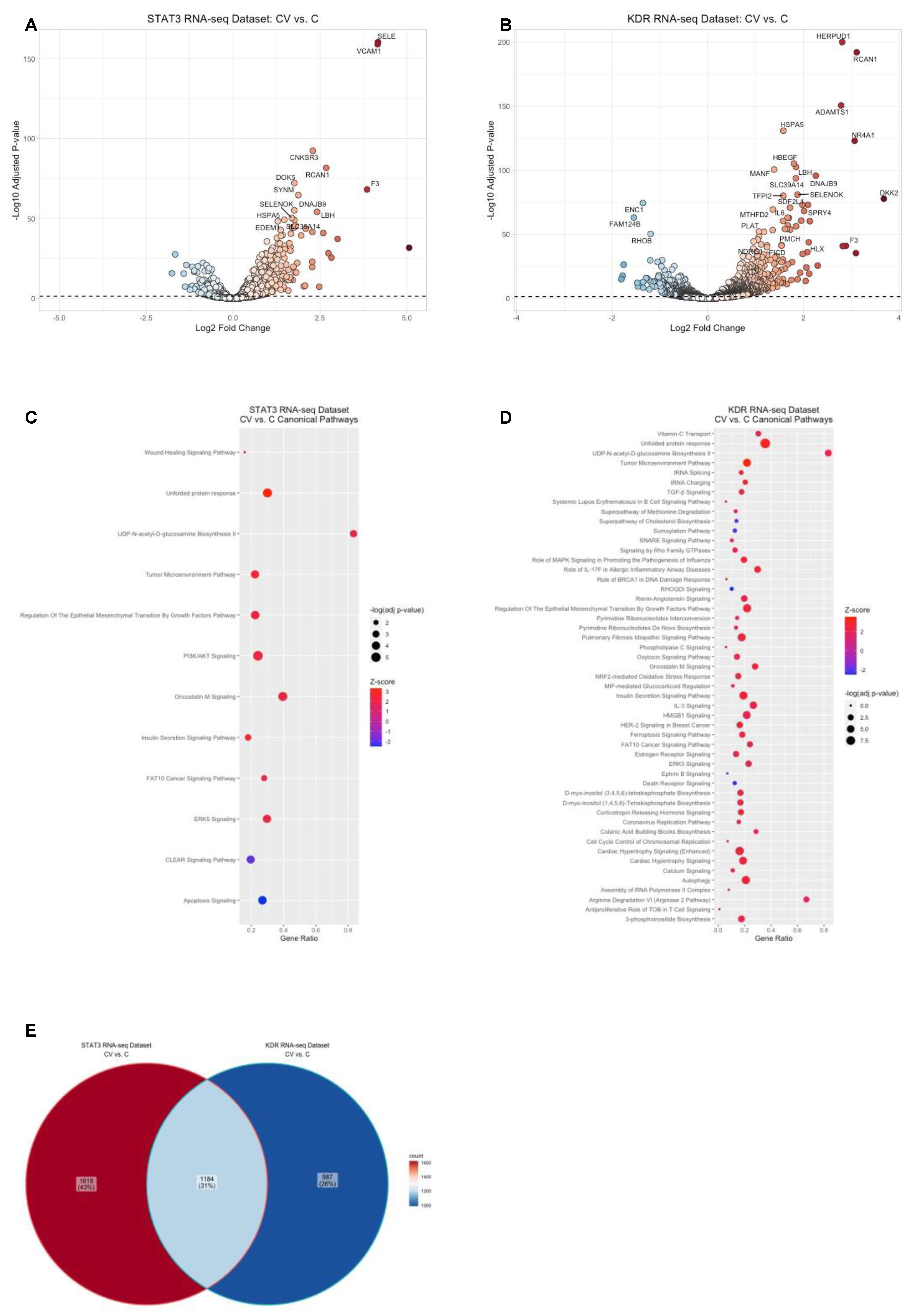
2.3. VEGFR2-Mediated Gene Regulation in HRMECs
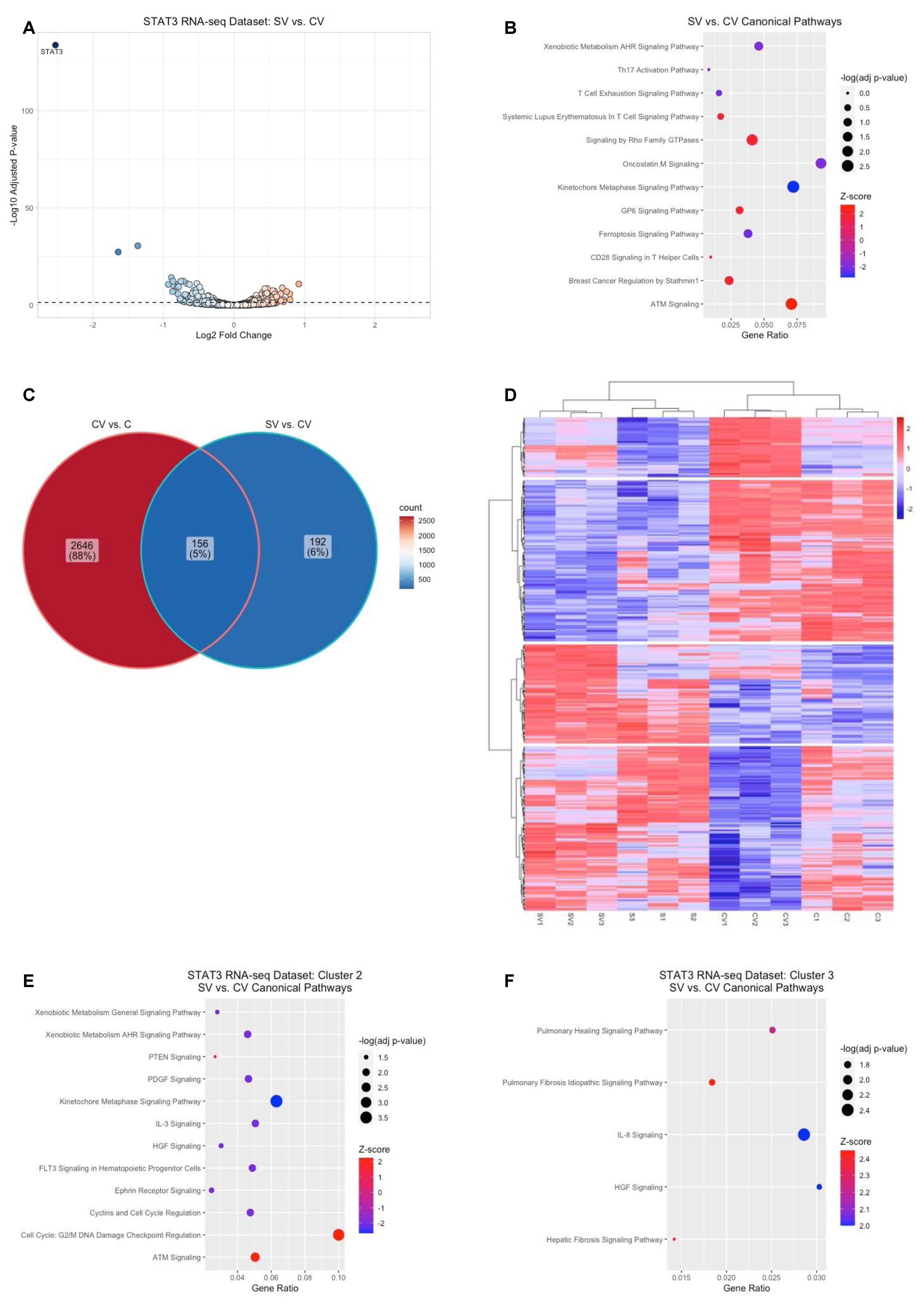
2.4. STAT3-Mediated Gene Regulation in HRMECs
2.5. Identification of Canonical Pathways Implicated in STAT3-Mediated or VEGFR2-Mediated Signaling by VEGF
3. Discussion
4. Materials and Methods
4.1. Cell Culture Conditions
4.2. Transfection
4.3. Treatments and RNA Isolation
4.4. RNA Quality Assessment
4.5. Library Preparation and High-Throughput RNA Sequencing
4.6. Genome Alignment and Differential Gene Expression Analysis
4.7. Pathway Enrichment Analysis
4.8. Real-Time PCR Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gilbert, C. Retinopathy of prematurity: A global perspective of the epidemics, population of babies at risk and implications for control. Early Hum. Dev. 2008, 84, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Hartnett, M.E.; Penn, J.S. Mechanisms and management of retinopathy of prematurity. N. Engl. J. Med. 2012, 367, 2515–2526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Early Treatment for Retinopathy of Prematurity Cooperative Group. Revised indications for the treatment of retinopathy of prematurity: Results of the early treatment for retinopathy of prematurity randomized trial. Arch. Ophthalmol. 2003, 121, 1684–1694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mintz-Hittner, H.A.; Kennedy, K.A.; Chuang, A.Z. Efficacy of Intravitreal Bevacizumab for Stage 3+ Retinopathy of Prematurity. N. Engl. J. Med. 2011, 364, 603–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J. Reactivation of retinopathy of prematurity after bevacizumab injection. Arch. Ophthalmol. 2012, 130, 1000–1006. [Google Scholar] [CrossRef]
- Ittiara, S.; Blair, M.P.; Shapiro, M.J.; Lichtenstein, S.J. Exudative retinopathy and detachment: A late reactivation of retinopathy of prematurity after intravitreal bevacizumab. J. Am. Assoc. Pediatr. Ophthalmol. Strabismus 2013, 17, 323–325. [Google Scholar] [CrossRef]
- Snyder, L.L.; Garcia-Gonzalez, J.M.; Shapiro, M.J.; Blair, M.P. Very Late Reactivation of Retinopathy of Prematurity after Monotherapy with Intravitreal Bevacizumab. Ophthalmic Surg. Lasers Imaging Retina 2016, 47, 280–283. [Google Scholar] [CrossRef] [Green Version]
- Saint-Geniez, M.; Maharaj, A.S.; Walshe, T.E.; Tucker, B.A.; Sekiyama, E.; Kurihara, T.; Darland, D.C.; Young, M.J.; D’Amore, P.A. Endogenous VEGF is required for visual function: Evidence for a survival role on muller cells and photoreceptors. PLoS ONE 2008, 3, e3554. [Google Scholar] [CrossRef] [Green Version]
- Becker, S.; Wang, H.; Simmons, A.B.; Suwanmanee, T.; Stoddard, G.J.; Kafri, T.; Hartnett, M.E. Targeted Knockdown of Overexpressed VEGFA or VEGF164 in Muller cells maintains retinal function by triggering different signaling mechanisms. Sci. Rep. 2018, 8, 2003. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Wang, H.; Culp, D.; Yang, Z.; Fotheringham, L.; Flannery, J.; Hammond, S.; Kafri, T.; Hartnett, M.E. Targeting Muller cell-derived VEGF164 to reduce intravitreal neovascularization in the rat model of retinopathy of prematurity. Investig. Ophthalmol. Vis. Sci. 2014, 55, 824–831. [Google Scholar] [CrossRef] [Green Version]
- Ferrara, N.; Gerber, H.P.; LeCouter, J. The biology of VEGF and its receptors. Nat. Med. 2003, 9, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Simons, M.; Gordon, E.; Claesson-Welsh, L. Mechanisms and regulation of endothelial VEGF receptor signalling. Nat. Rev. Mol. Cell Biol. 2016, 17, 611–625. [Google Scholar] [CrossRef] [PubMed]
- Penn, J.S.; Henry, M.M.; Tolman, B.L. Exposure to alternating hypoxia and hyperoxia causes severe proliferative retinopathy in the newborn rat. Pediatr. Res. 1994, 36, 724–731. [Google Scholar] [CrossRef] [Green Version]
- Simmons, A.B.; Bretz, C.A.; Wang, H.; Kunz, E.; Hajj, K.; Kennedy, C.; Yang, Z.; Suwanmanee, T.; Kafri, T.; Hartnett, M.E. Gene therapy knockdown of VEGFR2 in retinal endothelial cells to treat retinopathy. Angiogenesis 2018, 21, 751–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Byfield, G.; Jiang, Y.; Smith, G.W.; McCloskey, M.; Hartnett, M.E. VEGF-mediated STAT3 activation inhibits retinal vascularization by down-regulating local erythropoietin expression. Am. J. Pathol. 2012, 180, 1243–1253. [Google Scholar] [CrossRef] [Green Version]
- Byfield, G.; Budd, S.; Hartnett, M.E. The role of supplemental oxygen and JAK/STAT signaling in intravitreous neovascularization in a ROP rat model. Investig. Ophthalmol. Vis. Sci. 2009, 50, 3360–3365. [Google Scholar] [CrossRef] [Green Version]
- Sheng, Q.; Vickers, K.; Zhao, S.; Wang, J.; Samuels, D.C.; Koues, O.; Shyr, Y.; Guo, Y. Multi-perspective quality control of Illumina RNA sequencing data analysis. Brief Funct. Genom. 2017, 16, 194–204. [Google Scholar] [CrossRef] [Green Version]
- Ramshekar, A.; Hartnett, M.E. Vascular Endothelial Growth Factor Signaling in Models of Oxygen-Induced Retinopathy: Insights into Mechanisms of Pathology in Retinopathy of Prematurity. Front. Pediatr. 2021, 9, 796143. [Google Scholar] [CrossRef]
- Kusaka, S.; Shima, C.; Wada, K.; Arahori, H.; Shimojyo, H.; Sato, T.; Fujikado, T. Efficacy of intravitreal injection of bevacizumab for severe retinopathy of prematurity: A pilot study. Br. J. Ophthalmol. 2008, 92, 1450–1455. [Google Scholar] [CrossRef]
- Arambulo, O.; Dib, G.; Iturralde, J.; Duran, F.; Brito, M.; Fortes Filho, J.B. Intravitreal ranibizumab as a primary or a combined treatment for severe retinopathy of prematurity. Clin. Ophthalmol. 2015, 9, 2027–2032. [Google Scholar] [CrossRef] [Green Version]
- Castellanos, M.A.; Schwartz, S.; Garcia-Aguirre, G.; Quiroz-Mercado, H. Short-term outcome after intravitreal ranibizumab injections for the treatment of retinopathy of prematurity. Br. J. Ophthalmol. 2013, 97, 816–819. [Google Scholar] [CrossRef] [PubMed]
- Autrata, R.; Krejcirova, I.; Senkova, K.; Holousova, M.; Dolezel, Z.; Borek, I. Intravitreal pegaptanib combined with diode laser therapy for stage 3+ retinopathy of prematurity in zone I and posterior zone II. Eur. J. Ophthalmol. 2012, 22, 687–694. [Google Scholar] [CrossRef] [PubMed]
- Salman, A.G.; Said, A.M. Structural, visual and refractive outcomes of intravitreal aflibercept injection in high-risk prethreshold type 1 retinopathy of prematurity. Ophthalmic Res. 2015, 53, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Writing Committee for the Pediatric Eye Disease Investigator Group; Hartnett, M.E.; Wallace, D.K.; Dean, T.W.; Li, Z.; Boente, C.S.; Dosunmu, E.O.; Freedman, S.F.; Golden, R.P.; Kong, L.; et al. Plasma Levels of Bevacizumab and Vascular Endothelial Growth Factor after Low-Dose Bevacizumab Treatment for Retinopathy of Prematurity in Infants. JAMA Ophthalmol. 2022, 140, 337–344. [Google Scholar] [CrossRef]
- Crespo-Garcia, S.; Tsuruda, P.R.; Dejda, A.; Ryan, R.D.; Fournier, F.; Chaney, S.Y.; Pilon, F.; Dogan, T.; Cagnone, G.; Patel, P.; et al. Pathological angiogenesis in retinopathy engages cellular senescence and is amenable to therapeutic elimination via BCL-xL inhibition. Cell Metab. 2021, 33, 818–832. [Google Scholar] [CrossRef]
- Zasada, M.; Madetko-Talowska, A.; Revhaug, C.; Rognlien, A.G.W.; Baumbusch, L.O.; Ksiazek, T.; Szewczyk, K.; Grabowska, A.; Bik-Multanowski, M.; Jozef Pietrzyk, J.; et al. Transcriptome analysis reveals dysregulation of genes involved in oxidative phosphorylation in a murine model of retinopathy of prematurity. Pediatr. Res. 2020, 88, 391–397. [Google Scholar] [CrossRef]
- Li Calzi, S.; Shaw, L.C.; Moldovan, L.; Shelley, W.C.; Qi, X.; Racette, L.; Quigley, J.L.; Fortmann, S.D.; Boulton, M.E.; Yoder, M.C.; et al. Progenitor cell combination normalizes retinal vascular development in the oxygen-induced retinopathy (OIR) model. JCI Insight 2019, 4, e129224. [Google Scholar] [CrossRef] [Green Version]
- Rojo Arias, J.E.; Jaszai, J. Gene expression profile of the murine ischemic retina and its response to Aflibercept (VEGF-Trap). Sci. Rep. 2021, 11, 15313. [Google Scholar] [CrossRef]
- Smith, L.E.; Wesolowski, E.; McLellan, A.; Kostyk, S.K.; D’Amato, R.; Sullivan, R.; D’Amore, P.A. Oxygen-induced retinopathy in the mouse. Investig. Ophthalmol. Vis. Sci. 1994, 35, 101–111. [Google Scholar]
- Bullard, L.E.; Qi, X.; Penn, J.S. Role for Extracellular Signal-Responsive Kinase-1 and -2 in Retinal Angiogenesis. Investig. Ophthalmol. Vis. Sci. 2003, 44, 1722–1731. [Google Scholar] [CrossRef] [Green Version]
- Zasada, M.; Madetko-Talowska, A.; Revhaug, C.; Rognlien, A.G.W.; Baumbusch, L.O.; Ksiazek, T.; Szewczyk, K.; Grabowska, A.; Bik-Multanowski, M.; Jozef Pietrzyk, J.; et al. Short- and long-term impact of hyperoxia on the blood and retinal cells’ transcriptome in a mouse model of oxygen-induced retinopathy. Pediatr. Res. 2020, 87, 485–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kusuhara, S.; Fukushima, Y.; Fukuhara, S.; Jakt, L.M.; Okada, M.; Shimizu, Y.; Hata, M.; Nishida, K.; Negi, A.; Hirashima, M.; et al. Arhgef15 promotes retinal angiogenesis by mediating VEGF-induced Cdc42 activation and potentiating RhoJ inactivation in endothelial cells. PLoS ONE 2012, 7, e45858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, X.; Ueno, M.; Yamashita, T.; Ikuno, Y. RhoA activation and effect of Rho-kinase inhibitor in the development of retinal neovascularization in a mouse model of oxygen-induced retinopathy. Curr. Eye Res. 2011, 36, 1028–1036. [Google Scholar] [CrossRef] [PubMed]
- Suarez, S.; McCollum, G.W.; Bretz, C.A.; Yang, R.; Capozzi, M.E.; Penn, J.S. Modulation of VEGF-induced retinal vascular permeability by peroxisome proliferator-activated receptor-beta/delta. Investig. Ophthalmol. Vis. Sci. 2014, 55, 8232–8240. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Kramer, A.; Green, J.; Pollard, J., Jr.; Tugendreich, S. Causal analysis approaches in Ingenuity Pathway Analysis. Bioinformatics 2014, 30, 523–530. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramshekar, A.; Bretz, C.A.; Hartnett, M.E. RNA-Seq Provides Insights into VEGF-Induced Signaling in Human Retinal Microvascular Endothelial Cells: Implications in Retinopathy of Prematurity. Int. J. Mol. Sci. 2022, 23, 7354. https://doi.org/10.3390/ijms23137354
Ramshekar A, Bretz CA, Hartnett ME. RNA-Seq Provides Insights into VEGF-Induced Signaling in Human Retinal Microvascular Endothelial Cells: Implications in Retinopathy of Prematurity. International Journal of Molecular Sciences. 2022; 23(13):7354. https://doi.org/10.3390/ijms23137354
Chicago/Turabian StyleRamshekar, Aniket, Colin A. Bretz, and M. Elizabeth Hartnett. 2022. "RNA-Seq Provides Insights into VEGF-Induced Signaling in Human Retinal Microvascular Endothelial Cells: Implications in Retinopathy of Prematurity" International Journal of Molecular Sciences 23, no. 13: 7354. https://doi.org/10.3390/ijms23137354