
Effect of Psilocybin and Ketamine on Brain Neurotransmitters, Glutamate Receptors, DNA and Rat Behavior

 , and
, and 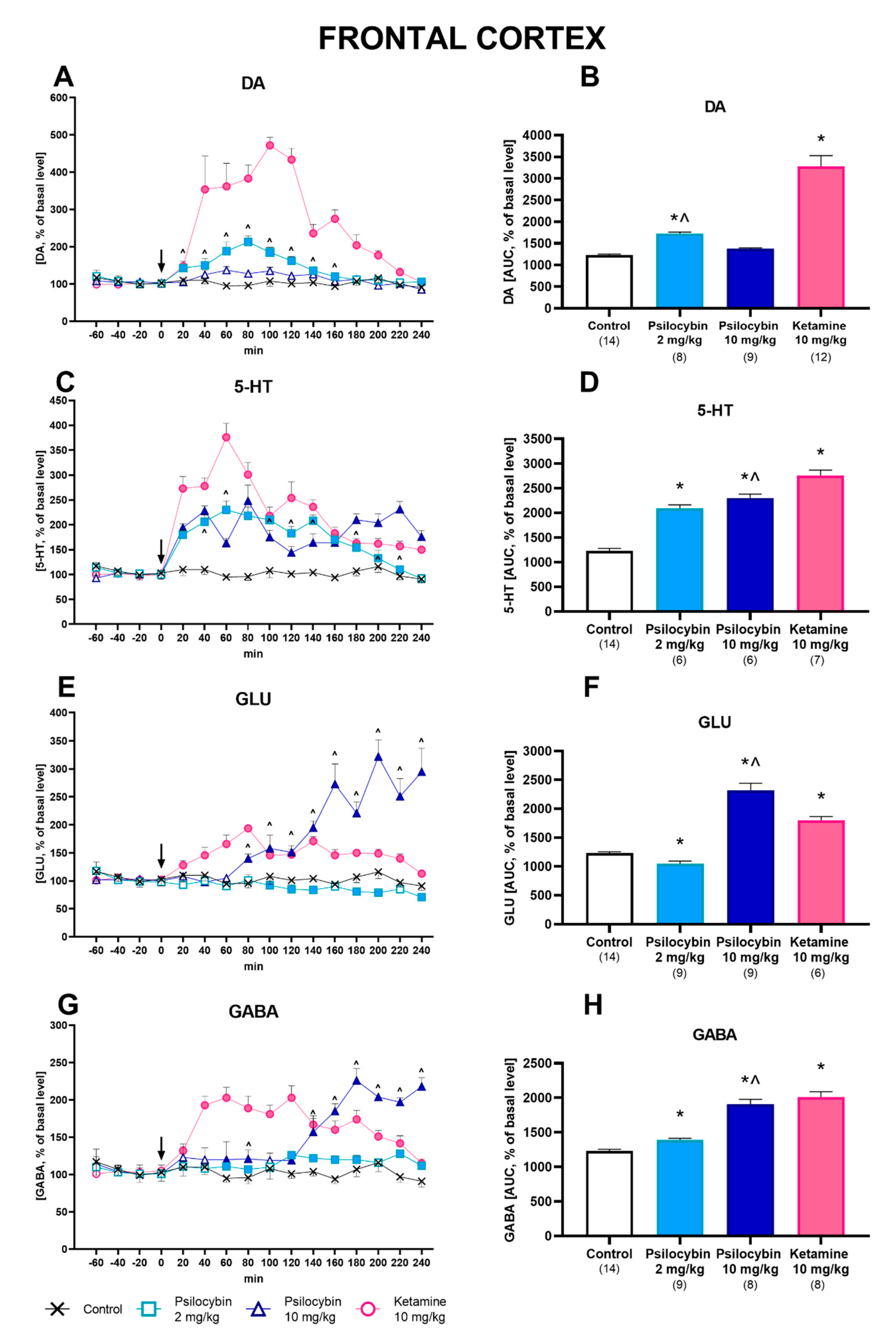{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effect of Psilocybin and Ketamine on Extracellular Levels of DA, 5-HT, Glutamate and GABA in the Rat Frontal Cortex
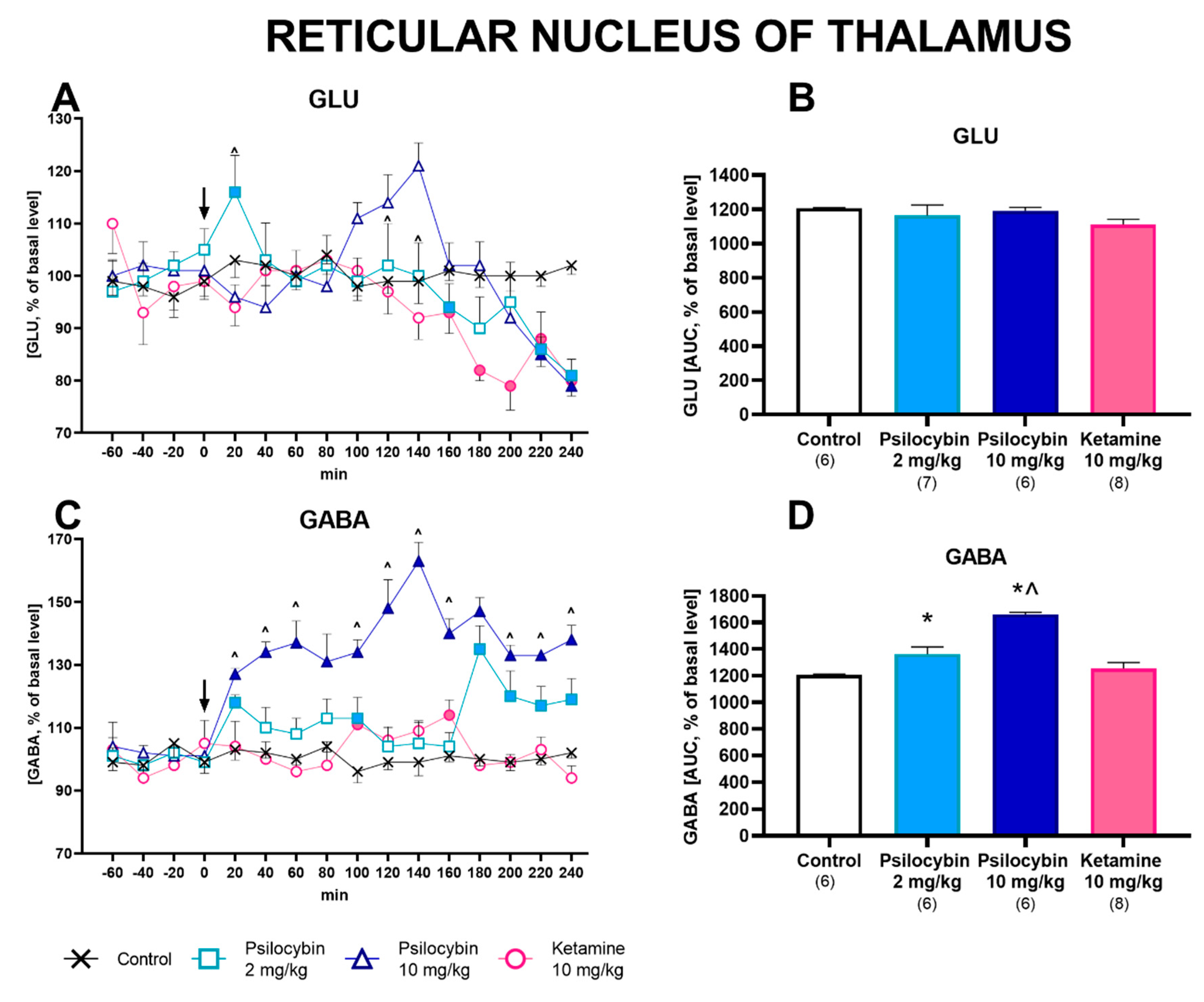
2.2. Effect of Psilocybin and Ketamine on Extracellular Levels of Glutamate and GABA in the Rat Reticular Nucleus of the Thalamus
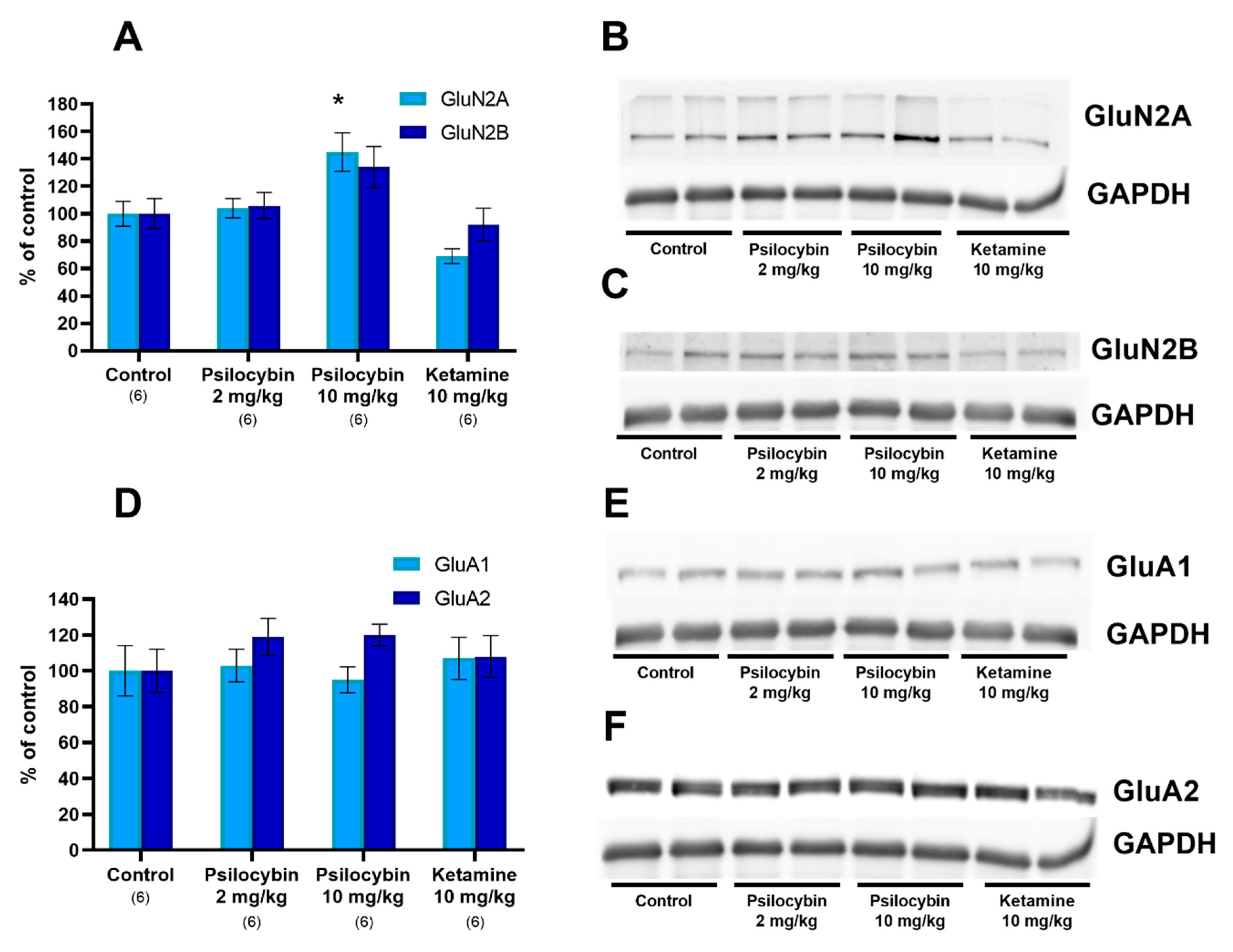
2.3. The NMDA and AMPA Receptor Subunit Level in the Rat Frontal Cortex
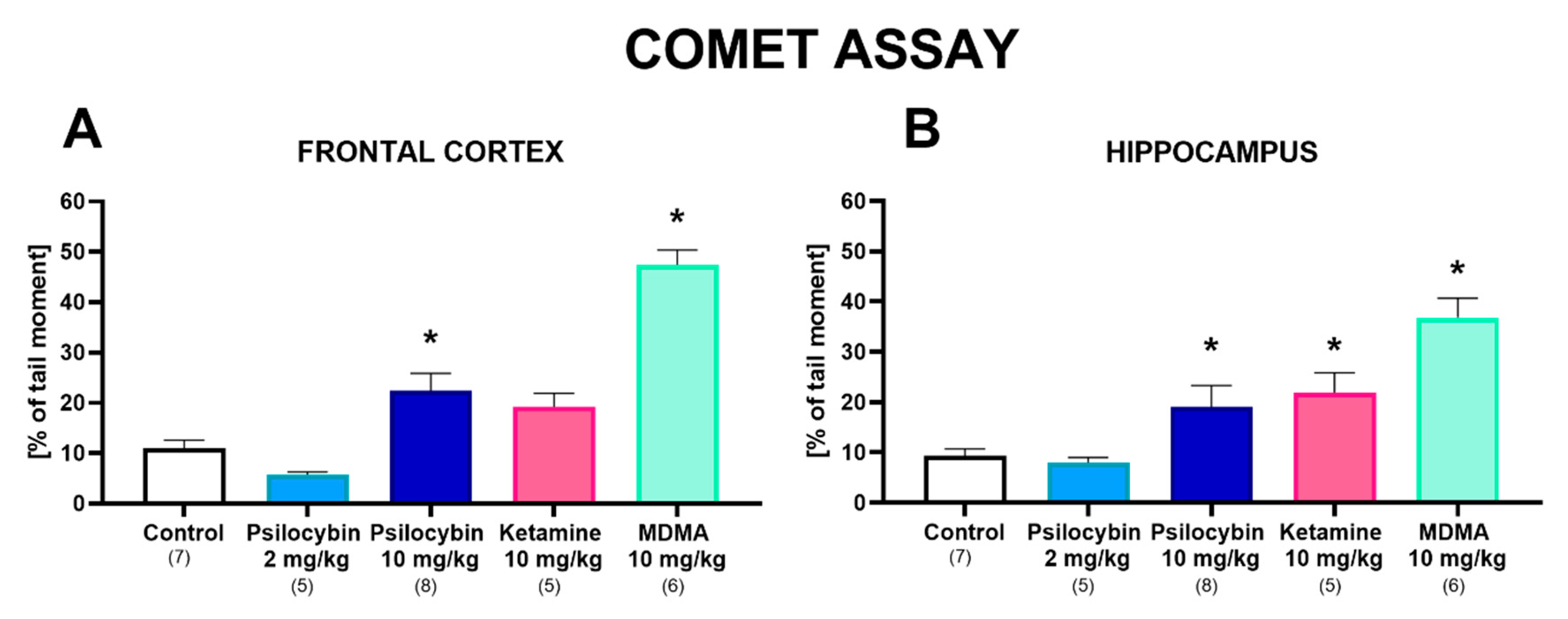
2.4. The Effect of Psilocybin and Ketamine on DNA Damage in the Rat Frontal Cortex and Hippocampus
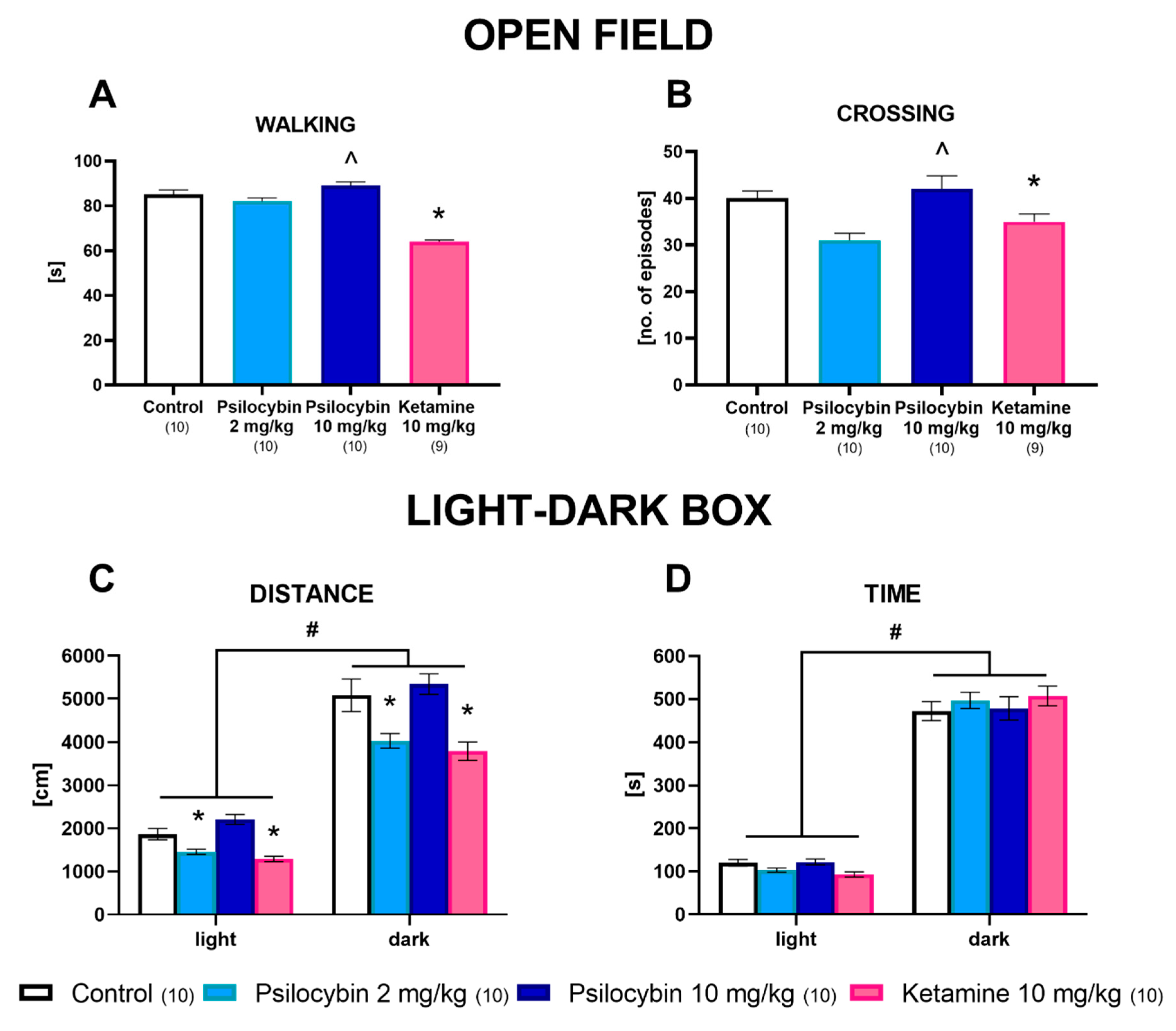
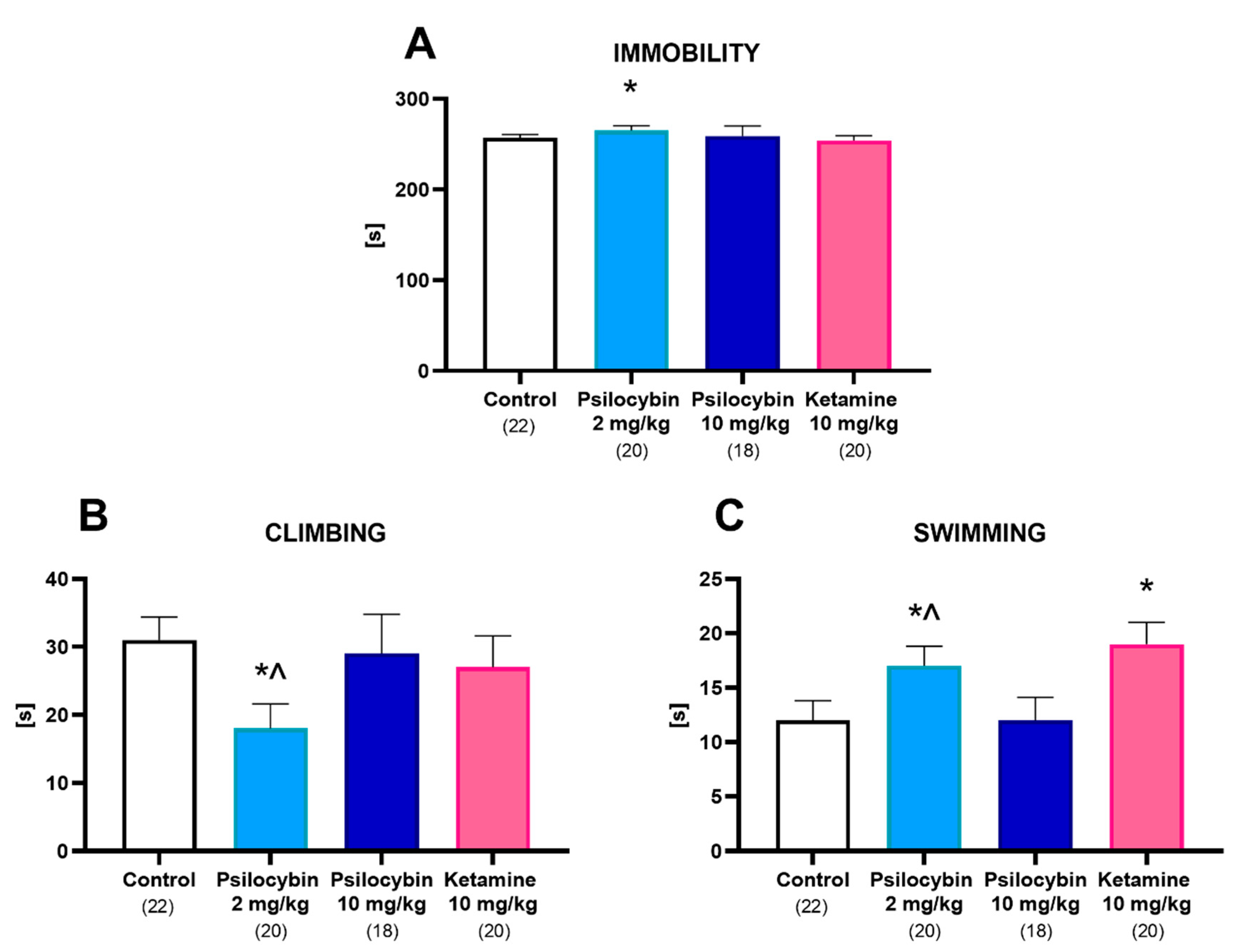
2.5. The Effect of Psilocybin and Ketamine on Rat Behavior in the Open Field, Light–Dark Box and Forced Swimming Tests 24 h after Administration
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Drugs and Reagents
4.3. Brain Microdialysis
4.4. Extracellular Concentration of DA, 5-HT, Glutamate and GABA
4.5. Alkaline Comet Assay
4.6. Western Blotting
4.7. Open Field (OF) Test
4.8. Light–Dark Box (LDB) Test
4.9. Forced Swim Test (FST) in Rats
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Whiteford, H.A.; Degenhardt, L.; Rehm, J.; Baxter, A.J.; Ferrari, A.J.; Erskine, H.E.; Charlson, F.J.; Norman, R.E.; Flaxman, A.D.; Johns, N.; et al. Global Burden of Disease Attributable to Mental and Substance Use Disorders: Findings from the Global Burden of Disease Study 2010. Lancet 2013, 382, 1575–1586. [Google Scholar] [CrossRef]
- Steffen, A.; Nübel, J.; Jacobi, F.; Bätzing, J.; Holstiege, J. Mental and Somatic Comorbidity of Depression: A Comprehensive Cross-Sectional Analysis of 202 Diagnosis Groups Using German Nationwide Ambulatory Claims Data. BMC Psychiatry 2020, 20, 142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obuobi-Donkor, G.; Nkire, N.; Agyapong, V.I.O. Prevalence of Major Depressive Disorder and Correlates of Thoughts of Death, Suicidal Behaviour, and Death by Suicide in the Geriatric Population—A General Review of Literature. Behav. Sci. 2021, 11, 142. [Google Scholar] [CrossRef] [PubMed]
- Duman, R.S.; Aghajanian, G.K.; Sanacora, G.; Krystal, J.H. Synaptic Plasticity and Depression: New Insights from Stress and Rapid-Acting Antidepressants. Nat. Med. 2016, 22, 238–249. [Google Scholar] [CrossRef] [Green Version]
- Andrade, C. Ketamine for Depression, 1: Clinical Summary of Issues Related to Efficacy, Adverse Effects, and Mechanism of Action. J. Clin. Psychiatry 2017, 78, e415–e419. [Google Scholar] [CrossRef]
- Carhart-Harris, R.; Giribaldi, B.; Watts, R.; Baker-Jones, M.; Murphy-Beiner, A.; Murphy, R.; Martell, J.; Blemings, A.; Erritzoe, D.; Nutt, D.J. Trial of Psilocybin versus Escitalopram for Depression. N. Engl. J. Med. 2021, 384, 1402–1411. [Google Scholar] [CrossRef]
- Maeng, S.; Zarate, C.A. The Role of Glutamate in Mood Disorders: Results from the Ketamine in Major Depression Study and the Presumed Cellular Mechanism Underlying Its Antidepressant Effects. Curr. Psychiatry Rep. 2007, 9, 467–474. [Google Scholar] [CrossRef]
- Mathews, D.C.; Henter, I.D.; Zarate, C.A. Targeting the Glutamatergic System to Treat Major Depressive Disorder: Rationale and Progress to Date. Drugs 2012, 72, 1313–1333. [Google Scholar] [CrossRef]
- Murrough, J.W.; Perez, A.M.; Pillemer, S.; Stern, J.; Parides, M.K.; Aan Het Rot, M.; Collins, K.A.; Mathew, S.J.; Charney, D.S.; Iosifescu, D.V. Rapid and Longer-Term Antidepressant Effects of Repeated Ketamine Infusions in Treatment-Resistant Major Depression. Biol. Psychiatry 2013, 74, 250–256. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, A.; Kabbaj, M. Sex Differences in Effects of Ketamine on Behavior, Spine Density, and Synaptic Proteins in Socially Isolated Rats. Biol. Psychiatry 2016, 80, 448–456. [Google Scholar] [CrossRef] [Green Version]
- Dong, C.; Zhang, J.C.; Yao, W.; Ren, Q.; Ma, M.; Yang, C.; Chaki, S.; Hashimoto, K. Rapid and Sustained Antidepressant Action of the MGlu2/3 Receptor Antagonist MGS0039 in the Social Defeat Stress Model: Comparison with Ketamine. Int. J. Neuropsychopharmacol. 2017, 20, 228–236. [Google Scholar] [CrossRef] [Green Version]
- Schultes, R.E.; Hofmann, A.; Rätsch, C. Plants of the Gods: Their Sacred, Healing, and Hallucinogenic Powers; Healing Arts Press: Fairfield, CT, USA, 1998; p. 208. [Google Scholar] [CrossRef]
- Nichols, D.E. Structure-Activity Relationships of Serotonin 5-HT2A Agonists. Wiley Interdiscip. Rev. Membr. Transp. Signal. 2012, 1, 559–579. [Google Scholar] [CrossRef]
- Halberstadt, A.L.; Geyer, M.A. Multiple Receptors Contribute to the Behavioral Effects of Indoleamine Hallucinogens. Neuropharmacology 2011, 61, 364–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierce, P.A.; Peroutka, S.J. Hallucinogenic Drug Interactions with Neurotransmitter Receptor Binding Sites in Human Cortex. Psychopharmacology 1989, 97, 118–122. [Google Scholar] [CrossRef]
- Titeler, M.; Lyon, R.A.; Glennon, R.A. Radioligand Binding Evidence Implicates the Brain 5-HT2 Receptor as a Site of Action for LSD and Phenylisopropylamine Hallucinogens. Psychopharmacology 1988, 94, 213–216. [Google Scholar] [CrossRef]
- Glennon, R.A.; Titeler, M.; McKenney, J.D. Evidence for 5-HT2 Involvement in the Mechanism of Action of Hallucinogenic Agents. Life Sci. 1984, 35, 2505–2511. [Google Scholar] [CrossRef]
- Sipes, T.A.; Geyer, M.A. DOI Disruption of Prepulse Inhibition of Startle in the Rat Is Mediated by 5-HT(2A) and Not by 5-HT(2C) Receptors. Behav. Pharmacol. 1995, 6, 839–842. [Google Scholar] [CrossRef]
- Wing, L.L.; Tapson, G.S.; Geyer, M.A. 5HT-2 Mediation of Acute Behavioral Effects of Hallucinogens in Rats. Psychopharmacology 1990, 100, 417–425. [Google Scholar] [CrossRef]
- Scruggs, J.L.; Schmidt, D.; Deutch, A.Y. The Hallucinogen 1-[2,5-Dimethoxy-4-Iodophenyl]-2-Aminopropane (DOI) Increases Cortical Extracellular Glutamate Levels in Rats. Neurosci. Lett. 2003, 346, 137–140. [Google Scholar] [CrossRef]
- Muschamp, J.W.; Regina, M.J.; Hull, E.M.; Winter, J.C.; Rabin, R.A. Lysergic Acid Diethylamide and [-]-2,5-Dimethoxy-4-Methylamphetamine Increase Extracellular Glutamate in Rat Prefrontal Cortex. Brain Res. 2004, 1023, 134–140. [Google Scholar] [CrossRef]
- Vollenweider, F.F. Brain Mechanisms of Hallucinogens and Entactogens. Dialogues Clin. Neurosci. 2001, 3, 353–365. [Google Scholar] [CrossRef]
- Nichols, D.E. Psychedelics. Pharmacol. Rev. 2016, 68, 264–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carhart-Harris, R.L.; Bolstridge, M.; Day, C.M.J.; Rucker, J.; Watts, R.; Erritzoe, D.E.; Kaelen, M.; Giribaldi, B.; Bloomfield, M.; Pilling, S.; et al. Psilocybin with Psychological Support for Treatment-Resistant Depression: Six-Month Follow-Up. Psychopharmacology 2018, 235, 399–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ly, C.; Greb, A.C.; Cameron, L.P.; Wong, J.M.; Barragan, E.V.; Wilson, P.C.; Burbach, K.F.; Soltanzadeh Zarandi, S.; Sood, A.; Paddy, M.R.; et al. Psychedelics Promote Structural and Functional Neural Plasticity. Cell Rep. 2018, 23, 3170–3182. [Google Scholar] [CrossRef] [PubMed]
- Hibicke, M.; Landry, A.N.; Kramer, H.M.; Talman, Z.K.; Nichols, C.D. Psychedelics, but Not Ketamine, Produce Persistent Antidepressant-like Effects in a Rodent Experimental System for the Study of Depression. ACS Appl. Mater. Interfaces 2020, 11, 864–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumeister, D.; Barnes, G.; Giaroli, G.; Tracy, D. Classical Hallucinogens as Antidepressants? A Review of Pharmacodynamics and Putative Clinical Roles. Ther. Adv. Psychopharmacol. 2014, 4, 156–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, E.-J.; Jeong, J.H.; Nguyen, B.-T.; Sharma, N.; Nah, S.-Y.; Chung, Y.H.; Lee, Y.; Byun, J.K.; Nabeshima, T.; Ko, S.K.; et al. Ginsenoside Re Protects against Serotonergic Behaviors Evoked by 2,5-Dimethoxy-4-iodo-amphetamine in Mice via Inhibition of PKCδ-Mediated Mitochondrial Dysfunction. Int. J. Mol. Sci. 2021, 22, 7219. [Google Scholar] [CrossRef]
- Wojtas, A.; Herian, M.; Skawski, M.; Sobocińska, M.; González-Marín, A.; Noworyta-Sokołowska, K.; Gołembiowska, K. Neurochemical and Behavioral Effects of a New Hallucinogenic Compound 25B-NBOMe in Rats. Neurotox. Res. 2021, 39, 305–326. [Google Scholar] [CrossRef]
- Savalia, N.K.; Shao, L.X.; Kwan, A.C. A Dendrite-Focused Framework for Understanding the Actions of Ketamine and Psychedelics. Trends Neurosci. 2021, 44, 260–275. [Google Scholar] [CrossRef]
- Moghaddam, B.; Adams, B.; Verma, A.; Daly, D. Activation of Glutamatergic Neurotransmission by Ketamine: A Novel Step in the Pathway from NMDA Receptor Blockade to Dopaminergic and Cognitive Disruptions Associated with the Prefrontal Cortex. J. Neurosci. 1997, 17, 2921–2927. [Google Scholar] [CrossRef]
- Wohleb, E.S.; Wu, M.; Gerhard, D.M.; Taylor, S.R.; Picciotto, M.R.; Alreja, M.; Duman, R.S. GABA Interneurons Mediate the Rapid Antidepressant-like Effects of Scopolamine. J. Clin. Investig. 2016, 126, 2482–2494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerhard, D.M.; Pothula, S.; Liu, R.J.; Wu, M.; Li, X.Y.; Girgenti, M.J.; Taylor, S.R.; Duman, C.H.; Delpire, E.; Picciotto, M.; et al. GABA Interneurons Are the Cellular Trigger for Ketamine’s Rapid Antidepressant Actions. J. Clin. Investig. 2020, 130, 1336–1349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pham, T.H.; Defaix, C.; Nguyen, T.M.L.; Mendez-David, I.; Tritschler, L.; David, D.J.; Gardier, A.M. Cortical and Raphe GABAA, AMPA Receptors and Glial GLT-1 Glutamate Transporter Contribute to the Sustained Antidepressant Activity of Ketamine. Pharmacol. Biochem. Behav. 2020, 192, 172913. [Google Scholar] [CrossRef]
- Ingram, R.; Kang, H.; Lightman, S.; Jane, D.E.; Bortolotto, Z.A.; Collingridge, G.L.; Lodge, D.; Volianskis, A. Some Distorted Thoughts about Ketamine as a Psychedelic and a Novel Hypothesis Based on NMDA Receptor-Mediated Synaptic Plasticity. Neuropharmacology 2018, 142, 30–40. [Google Scholar] [CrossRef] [Green Version]
- Duman, R.S.; Sanacora, G.; Krystal, J.H. Altered Connectivity in Depression: GABA and Glutamate Neurotransmitter Deficits and Reversal by Novel Treatments. Neuron 2019, 102, 75–90. [Google Scholar] [CrossRef]
- Yamamoto, S.; Ohba, H.; Nishiyama, S.; Harada, N.; Kakiuchi, T.; Tsukada, H.; Domino, E.F. Subanesthetic Doses of Ketamine Transiently Decrease Serotonin Transporter Activity: A PET Study in Conscious Monkeys. Neuropsychopharmacology 2013, 38, 2666–2674. [Google Scholar] [CrossRef] [Green Version]
- Fukumoto, K.; Iijima, M.; Chaki, S. The Antidepressant Effects of an MGlu2/3 Receptor Antagonist and Ketamine Require AMPA Receptor Stimulation in the MPFC and Subsequent Activation of the 5-HT Neurons in the DRN. Neuropsychopharmacology 2016, 41, 1046–1056. [Google Scholar] [CrossRef]
- Ago, Y.; Tanabe, W.; Higuchi, M.; Tsukada, S.; Tanaka, T.; Yamaguchi, T.; Igarashi, H.; Yokoyama, R.; Seiriki, K.; Kasai, A.; et al. (R)-Ketamine Induces a Greater Increase in Prefrontal 5-HT Release Than (S)-Ketamine and Ketamine Metabolites via an AMPA Receptor-Independent Mechanism. Int. J. Neuropsychopharmacol. 2019, 22, 665–674. [Google Scholar] [CrossRef]
- Vertes, R.P. Differential Projections of the Infralimbic and Prelimbic Cortex in the Rat. Synapse 2004, 51, 32–58. [Google Scholar] [CrossRef]
- Wu, M.; Minkowicz, S.; Dumrongprechachan, V.; Hamilton, P.; Xiao, L.; Kozorovitskiy, Y. Attenuated Dopamine Signaling after Aversive Learning Is Restored by Ketamine to Rescue Escape Actions. elife 2021, 10, e64041. [Google Scholar] [CrossRef]
- Kokkinou, M.; Ashok, A.H.; Howes, O.D. The Effects of Ketamine on Dopaminergic Function: Meta-Analysis and Review of the Implications for Neuropsychiatric Disorders. Mol. Psychiatry 2018, 23, 59–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Passie, T.; Seifert, J.; Schneider, U.; Emrich, H.M. The Pharmacology of Psilocybin. Addict. Biol. 2002, 7, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Willins, D.L.; Deutch, A.Y.; Roth, B.L. Serotonin 5-HT(2A) Receptors Are Expressed on Pyramidal Cells and Interneurons in the Rat Cortex. Synapse 1997, 27, 79–82. [Google Scholar] [CrossRef]
- Barre, A.; Berthoux, C.; De Bundel, D.; Valjent, E.; Bockaert, J.; Marin, P.; Bécamel, C. Presynaptic Serotonin 2A Receptors Modulate Thalamocortical Plasticity and Associative Learning. Proc. Natl. Acad. Sci. USA 2016, 113, E1382–E1391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mason, N.L.; Kuypers, K.P.C.; Müller, F.; Reckweg, J.; Tse, D.H.Y.; Toennes, S.W.; Hutten, N.R.P.W.; Jansen, J.F.A.; Stiers, P.; Feilding, A.; et al. Me, Myself, Bye: Regional Alterations in Glutamate and the Experience of Ego Dissolution with Psilocybin. Neuropsychopharmacology 2020, 45, 2003–2011. [Google Scholar] [CrossRef]
- De Deurwaerdère, P.; Di Giovanni, G. Serotonergic modulation of the activity of mesencephalic dopaminergic systems: Therapeutic implications. Prog. Neurobiol. 2017, 151, 175–236. [Google Scholar] [CrossRef]
- Celada, P.; Victoria Puig, M.; Casanovas, J.M.; Guillazo, G.; Artigas, F. Control of Dorsal Raphe Serotonergic Neurons by the Medial Prefrontal Cortex: Involvement of Serotonin-1A, GABAA, and Glutamate Receptors. J. Neurosci. 2001, 21, 9917–9929. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Jolas, T.; Aghajanian, G. Serotonin 5-HT2 Receptors Activate Local GABA Inhibitory Inputs to Serotonergic Neurons of the Dorsal Raphe Nucleus. Brain Res. 2000, 873, 34–45. [Google Scholar] [CrossRef]
- Herian, M.; Wojtas, A.; Kamińska, K.; Świt, P.; Wach, A.; Gołembiowska, K. Hallucinogen-Like Action of the Novel Designer Drug 25I-NBOMe and Its Effect on Cortical Neurotransmitters in Rats. Neurotox Res. 2019, 36, 91–100. [Google Scholar] [CrossRef] [Green Version]
- Sakashita, Y.; Abe, K.; Katagiri, N.; Kambe, T.; Saitoh, T.; Utsunomiya, I.; Horiguchi, Y.; Taguchi, K. Effect of Psilocin on Extracellular Dopamine and Serotonin Levels in the Mesoaccumbens and Mesocortical Pathway in Awake Rats. Biol. Pharm. Bull. 2015, 38, 134–138. [Google Scholar] [CrossRef] [Green Version]
- Troyano-Rodriguez, E.; Lladó-Pelfort, L.; Santana, N.; Teruel-Martí, V.; Celada, P.; Artigas, F. Phencyclidine Inhibits the Activity of Thalamic Reticular Gamma-Aminobutyric Acidergic Neurons in Rat Brain. Biol. Psychiatry 2014, 76, 937–945. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, J.J.; Noristani, H.N.; Hoover, W.B.; Linley, S.B.; Vertes, R.P. Serotonergic Projections and Serotonin Receptor Expression in the Reticular Nucleus of the Thalamus in the Rat. Synapse 2011, 65, 919–928. [Google Scholar] [CrossRef] [PubMed]
- Nichols, D.E. Hallucinogens. Pharmacol. Ther. 2004, 101, 131–181. [Google Scholar] [CrossRef] [PubMed]
- Vollenweider, F.X.; Smallridge, J.W. Classic Psychedelic Drugs: Update on Biological Mechanisms. Pharmacopsychiatry 2022, 55, 121–138. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Liu, R.J.; Dwyer, J.M.; Banasr, M.; Lee, B.; Son, H.; Li, X.Y.; Aghajanian, G.; Duman, R.S. Glutamate N-Methyl-D-Aspartate Receptor Antagonists Rapidly Reverse Behavioral and Synaptic Deficits Caused by Chronic Stress Exposure. Biol. Psychiatry 2011, 69, 754–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Preskorn, S.H.; Baker, B.; Kolluri, S.; Menniti, F.S.; Krams, M.; Landen, J.W. An Innovative Design to Establish Proof of Concept of the Antidepressant Effects of the NR2B Subunit Selective N-Methyl-d-Aspartate Antagonist, CP-101,606, in Patients with Treatment-Refractory Major Depressive Disorder. J. Clin. Psychopharmacol. 2008, 28, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Pothula, S.; Kato, T.; Liu, R.J.; Wu, M.; Gerhard, D.; Shinohara, R.; Sliby, A.N.; Chowdhury, G.M.I.; Behar, K.L.; Sanacora, G.; et al. Cell-Type Specific Modulation of NMDA Receptors Triggers Antidepressant Actions. Mol. Psychiatry 2021, 26, 5097–5111. [Google Scholar] [CrossRef]
- Pryazhnikov, E.; Mugantseva, E.; Casarotto, P.; Kolikova, J.; Fred, S.M.; Toptunov, D.; Afzalov, R.; Hotulainen, P.; Voikar, V.; Terry-Lorenzo, R.; et al. Longitudinal Two-Photon Imaging in Somatosensory Cortex of Behaving Mice Reveals Dendritic Spine Formation Enhancement by Subchronic Administration of Low-Dose Ketamine. Sci. Rep. 2018, 8, 6464. [Google Scholar] [CrossRef] [Green Version]
- Martin, D.A.; Marona-Lewicka, D.; Nichols, D.E.; Nichols, C.D. Chronic LSD Alters Gene Expression Profiles in the MPFC Relevant to Schizophrenia. Neuropharmacology 2014, 83, 1–8. [Google Scholar] [CrossRef] [Green Version]
- De la Fuente Revenga, M.; Zhu, B.; Guevara, C.A.; Naler, L.B.; Saunders, J.M.; Zhou, Z.; Toneatti, R.; Sierra, S.; Wolstenholme, J.T.; Beardsley, P.M.; et al. Prolonged Epigenomic and Synaptic Plasticity Alterations Following Single Exposure to a Psychedelic in Mice. Cell Rep. 2021, 37, 109836. [Google Scholar] [CrossRef]
- Hanks, J.B.; González-Maeso, J. Animal Models of Serotonergic Psychedelics. ACS Chem. Neurosci. 2013, 4, 33–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jefsen, O.; Højgaard, K.; Christiansen, S.L.; Elfving, B.; Nutt, D.J.; Wegener, G.; Müller, H.K. Psilocybin Lacks Antidepressant-like Effect in the Flinders Sensitive Line Rat. Acta Neuropsychiatr. 2019, 31, 213–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viktorov, M.; Wilkinson, M.P.; Elston, V.C.E.; Stone, M.; Robinson, E.S.J. A Systematic Review of Studies Investigating the Acute Effects of N -Methyl-D-Aspartate Receptor Antagonists on Behavioural Despair in Normal Animals Suggests Poor Predictive Validity. Brain Neurosci. Adv. 2022, 6, 23982128221081645. [Google Scholar] [CrossRef] [PubMed]
- Cao, D.; Yu, J.; Wang, H.; Luo, Z.; Liu, X.; He, L.; Qi, J.; Fan, L.; Tang, L.; Chen, Z.; et al. Structure-based discovery of nonhallucinogenic psychedelic analogs. Science 2022, 375, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Shirota, O.; Hakamata, W.; Goda, Y. Concise Large-Scale Synthesis of Psilocin and Psilocybin, Principal Hallucinogenic Constituents of “Magic Mushroom”. J. Nat. Prod. 2003, 66, 885–887. [Google Scholar] [CrossRef] [Green Version]
- Popik, P.; Hogendorf, A.; Bugno, R.; Khoo, S.Y.S.; Zajdel, P.; Malikowska-Racia, N.; Nikiforuk, A.; Golebiowska, J. Effects of Ketamine Optical Isomers, Psilocybin, Psilocin and Norpsilocin on Time Estimation and Cognition in Rats. Psychopharmacology 2022, 239, 1689–1703. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates; Academic Press: San Diego, CA, USA, 1998. [Google Scholar]
- Herian, M.; Skawski, M.; Wojtas, A.; Sobocińska, M.K.; Noworyta, K.; Gołembiowska, K. Tolerance to Neurochemical and Behavioral Effects of the Hallucinogen 25I-NBOMe. Psychopharmacology 2021, 238, 2349–2364. [Google Scholar] [CrossRef]
- Maćkowiak, M.; Latusz, J.; Głowacka, U.; Bator, E.; Bilecki, W. Adolescent Social Isolation Affects Parvalbumin Expression in the Medial Prefrontal Cortex in the MAM-E17 Model of Schizophrenia. Metab. Brain Dis. 2019, 34, 341–352. [Google Scholar] [CrossRef]
- Latusz, J.; Maćkowiak, M. Early-Life Blockade of NMDA Receptors Induces Epigenetic Abnormalities in the Adult Medial Prefrontal Cortex: Possible Involvement in Memory Impairment in Trace Fear Conditioning. Psychopharmacology 2020, 237, 231–248. [Google Scholar] [CrossRef] [Green Version]
- Rogóz, Z.; Skuza, G. Anxiolytic-like Effects of Olanzapine, Risperidone and Fluoxetine in the Elevated plus-Maze Test in Rats. Pharmacol. Rep. 2011, 63, 1547–1552. [Google Scholar] [CrossRef]
- Chocyk, A.; Majcher-Maślanka, I.; Przyborowska, A.; Maćkowiak, M.; Wedzony, K. Early-Life Stress Increases the Survival of Midbrain Neurons during Postnatal Development and Enhances Reward-Related and Anxiolytic-like Behaviors in a Sex-Dependent Fashion. Int. J. Dev. Neurosci. 2015, 44, 33–47. [Google Scholar] [CrossRef] [PubMed]
- Bilecki, W.; Latusz, J.; Gawlińska, K.; Chmelova, M.; Maćkowiak, M. Prenatal MAM Treatment Altered Fear Conditioning Following Social Isolation: Relevance to Schizophrenia. Behav. Brain Res. 2021, 406, 113231. [Google Scholar] [CrossRef] [PubMed]
- Detke, M.J.; Rickels, M.; Lucki, I. Active Behaviors in the Rat Forced Swimming Test Differentially Produced by Serotonergic and Noradrenergic Antidepressants. Psychopharmacology 1995, 121, 66–72. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wojtas, A.; Bysiek, A.; Wawrzczak-Bargiela, A.; Szych, Z.; Majcher-Maślanka, I.; Herian, M.; Maćkowiak, M.; Gołembiowska, K. Effect of Psilocybin and Ketamine on Brain Neurotransmitters, Glutamate Receptors, DNA and Rat Behavior. Int. J. Mol. Sci. 2022, 23, 6713. https://doi.org/10.3390/ijms23126713
Wojtas A, Bysiek A, Wawrzczak-Bargiela A, Szych Z, Majcher-Maślanka I, Herian M, Maćkowiak M, Gołembiowska K. Effect of Psilocybin and Ketamine on Brain Neurotransmitters, Glutamate Receptors, DNA and Rat Behavior. International Journal of Molecular Sciences. 2022; 23(12):6713. https://doi.org/10.3390/ijms23126713
Chicago/Turabian StyleWojtas, Adam, Agnieszka Bysiek, Agnieszka Wawrzczak-Bargiela, Zuzanna Szych, Iwona Majcher-Maślanka, Monika Herian, Marzena Maćkowiak, and Krystyna Gołembiowska. 2022. "Effect of Psilocybin and Ketamine on Brain Neurotransmitters, Glutamate Receptors, DNA and Rat Behavior" International Journal of Molecular Sciences 23, no. 12: 6713. https://doi.org/10.3390/ijms23126713