

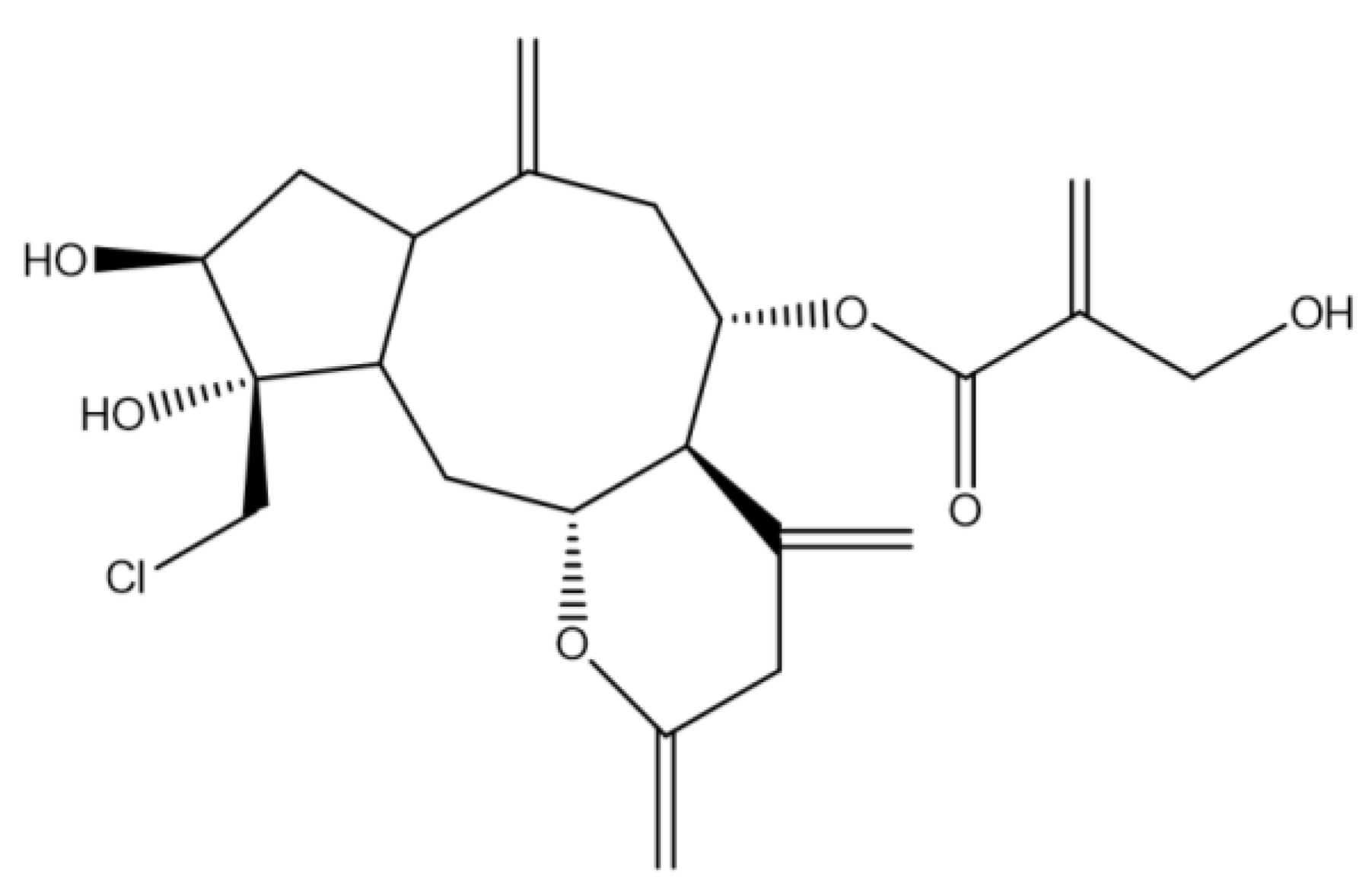
Assessment of the Anticancer Effect of Chlorojanerin Isolated from Centaurothamnus maximus on A549 Lung Cancer Cells
 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Results
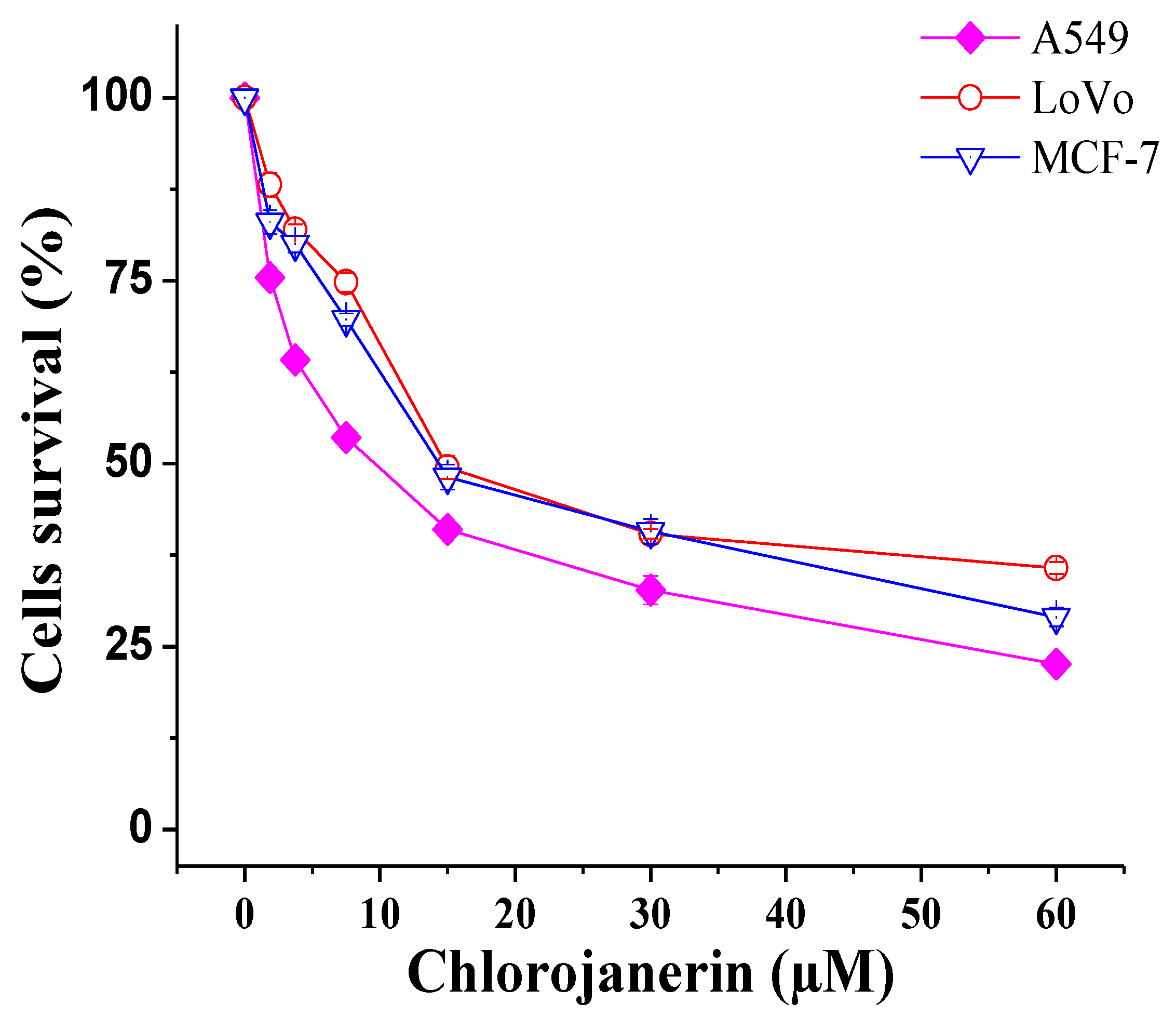
2.1. Antiproliferative Effects of Chlorojanerin on Cancer Cell Lines
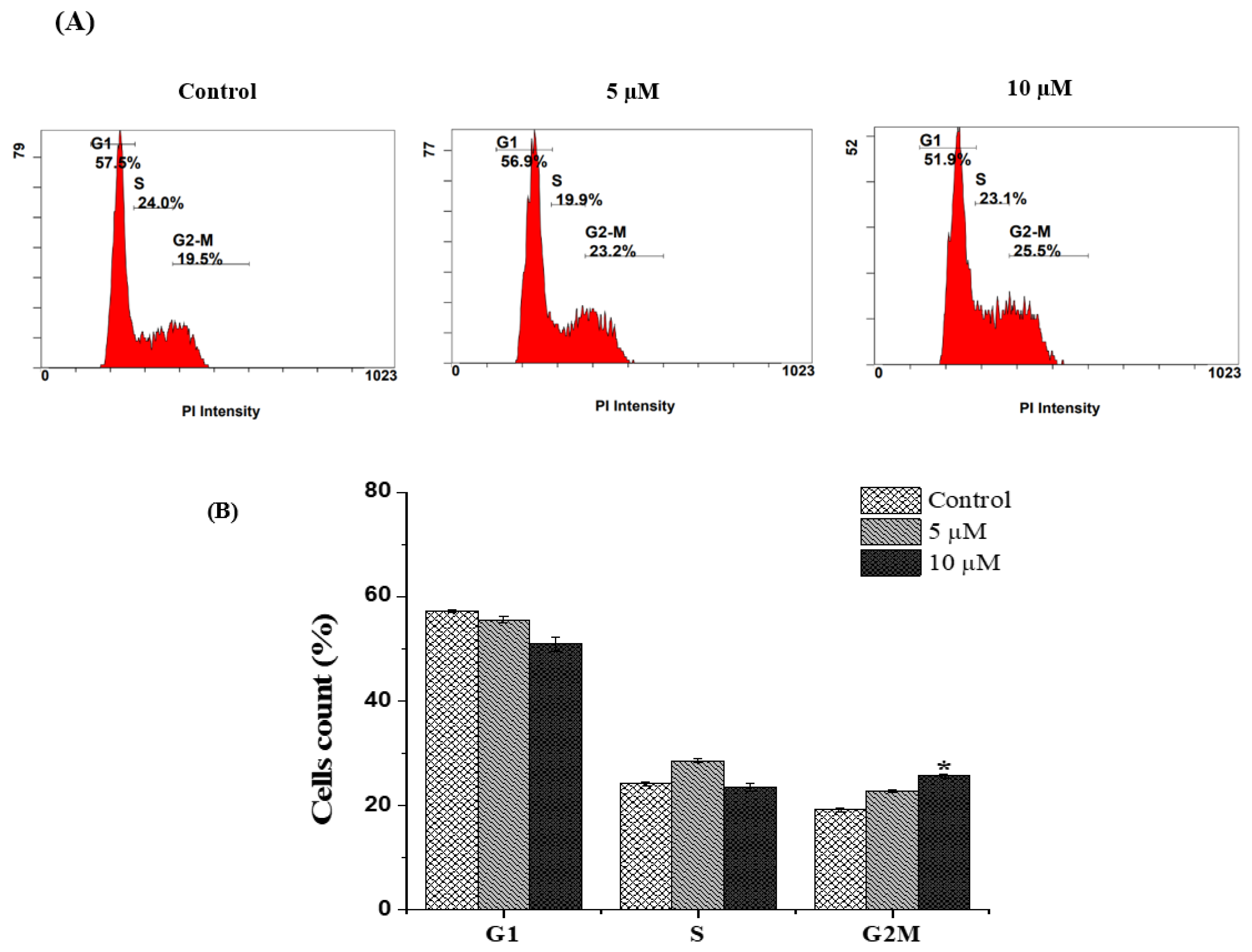
2.2. Chlorojanerin Induces G2/M Cell Cycle Arrest in A549 Lung Cancer Cells
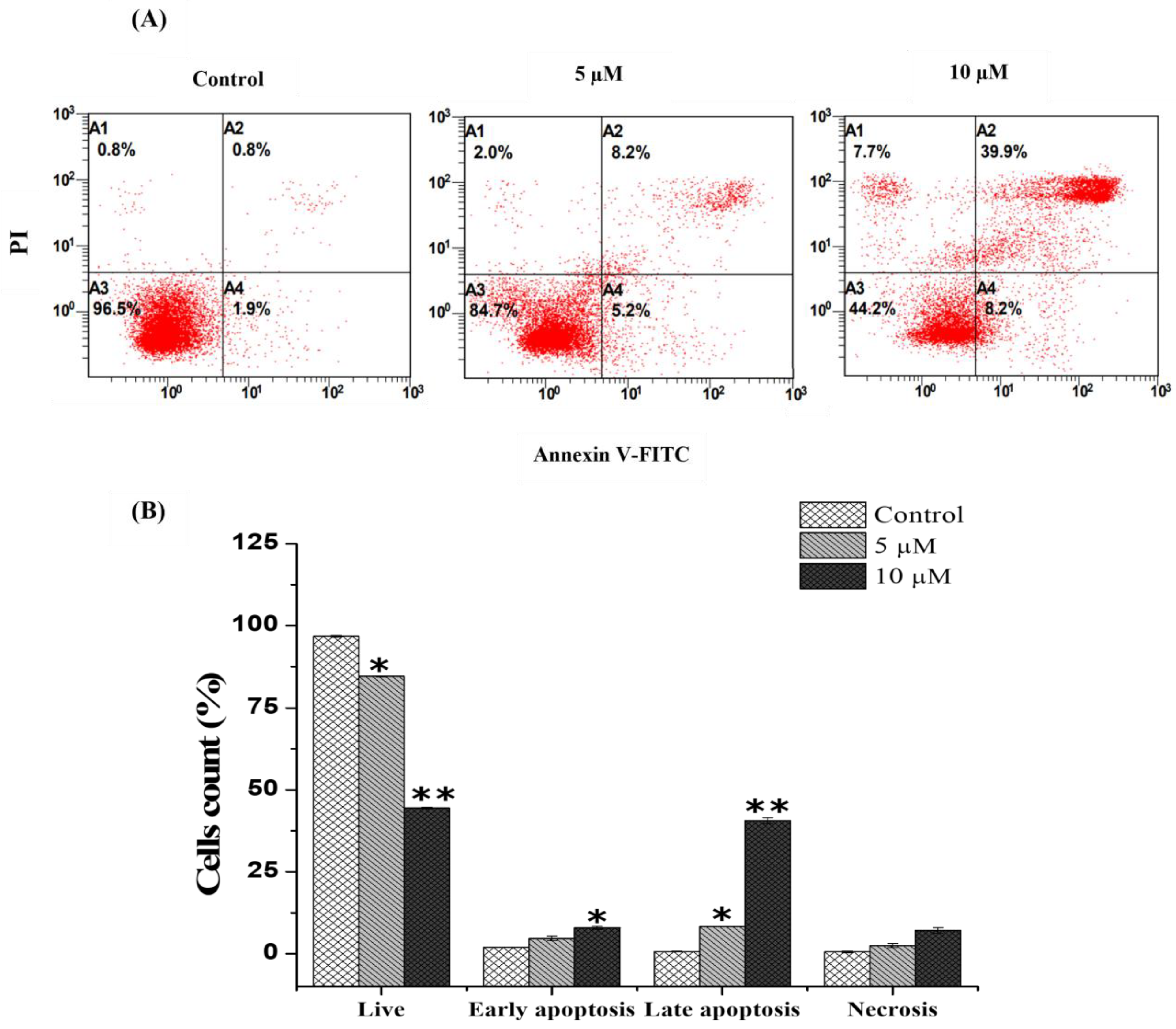
2.3. Chlorojanerin Promotes the Apoptosis of A549 Cells
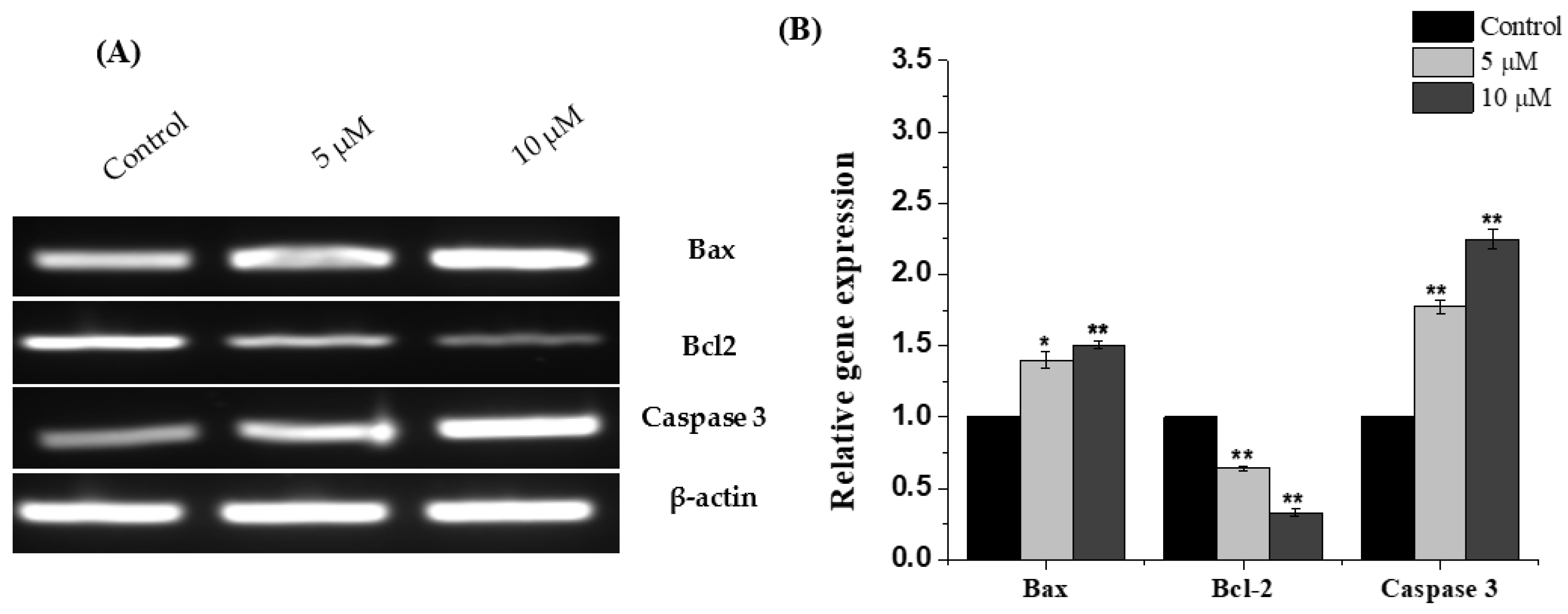
2.4. Chlorojanerin Modulated the Expression of Apoptosis Related Genes
2.5. Chlorojanerin Modulated the Expression of Apoptosis Related Proteins
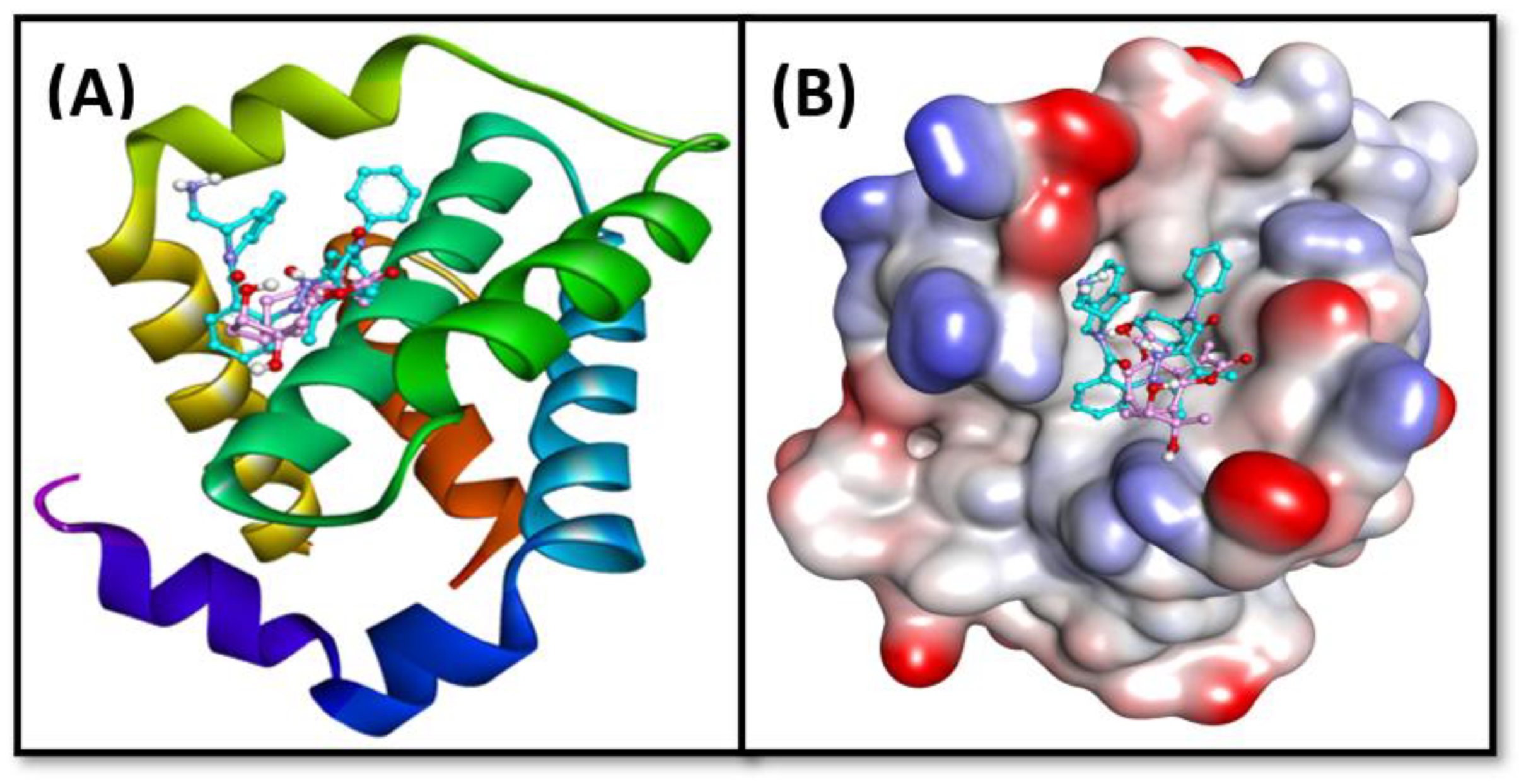
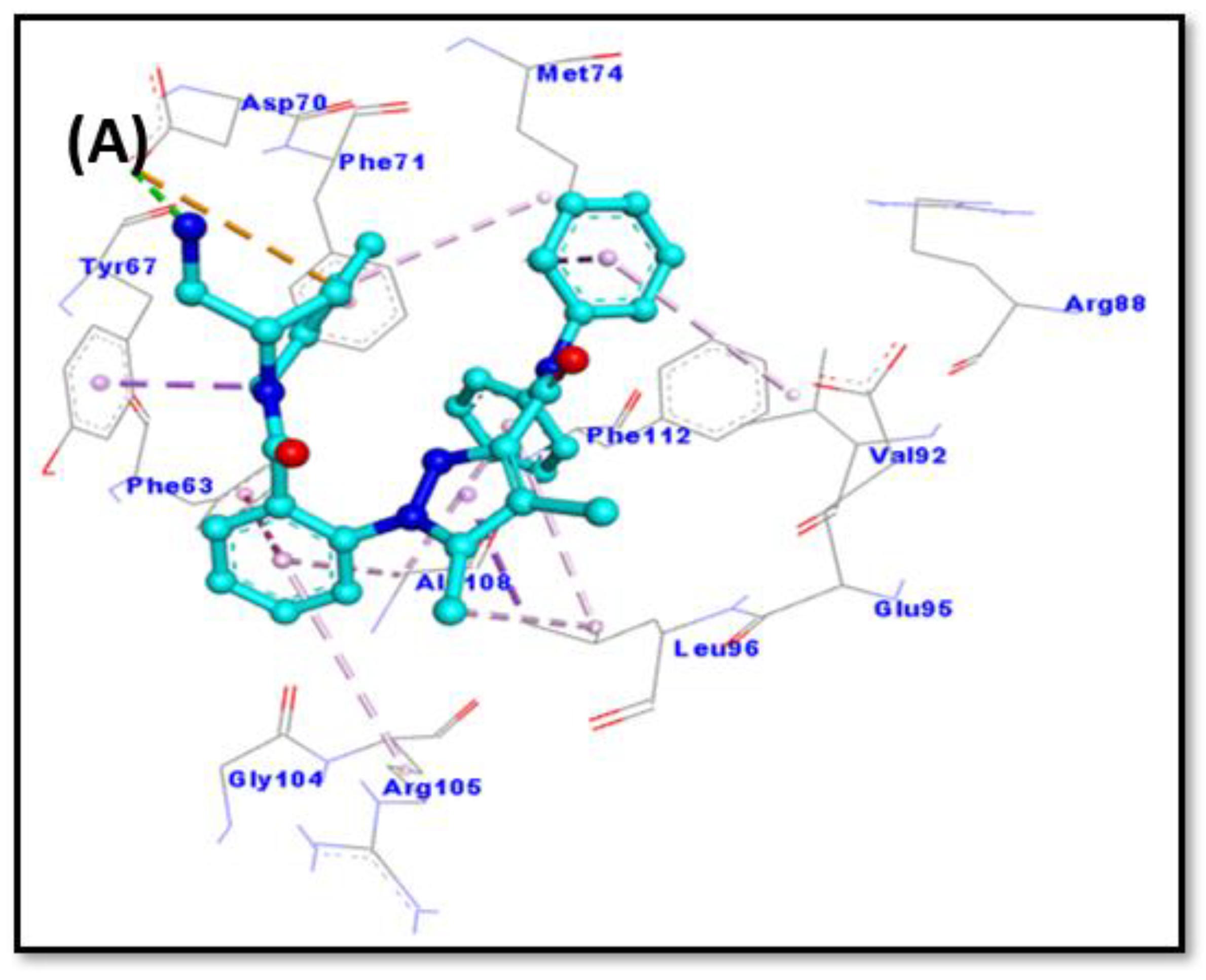
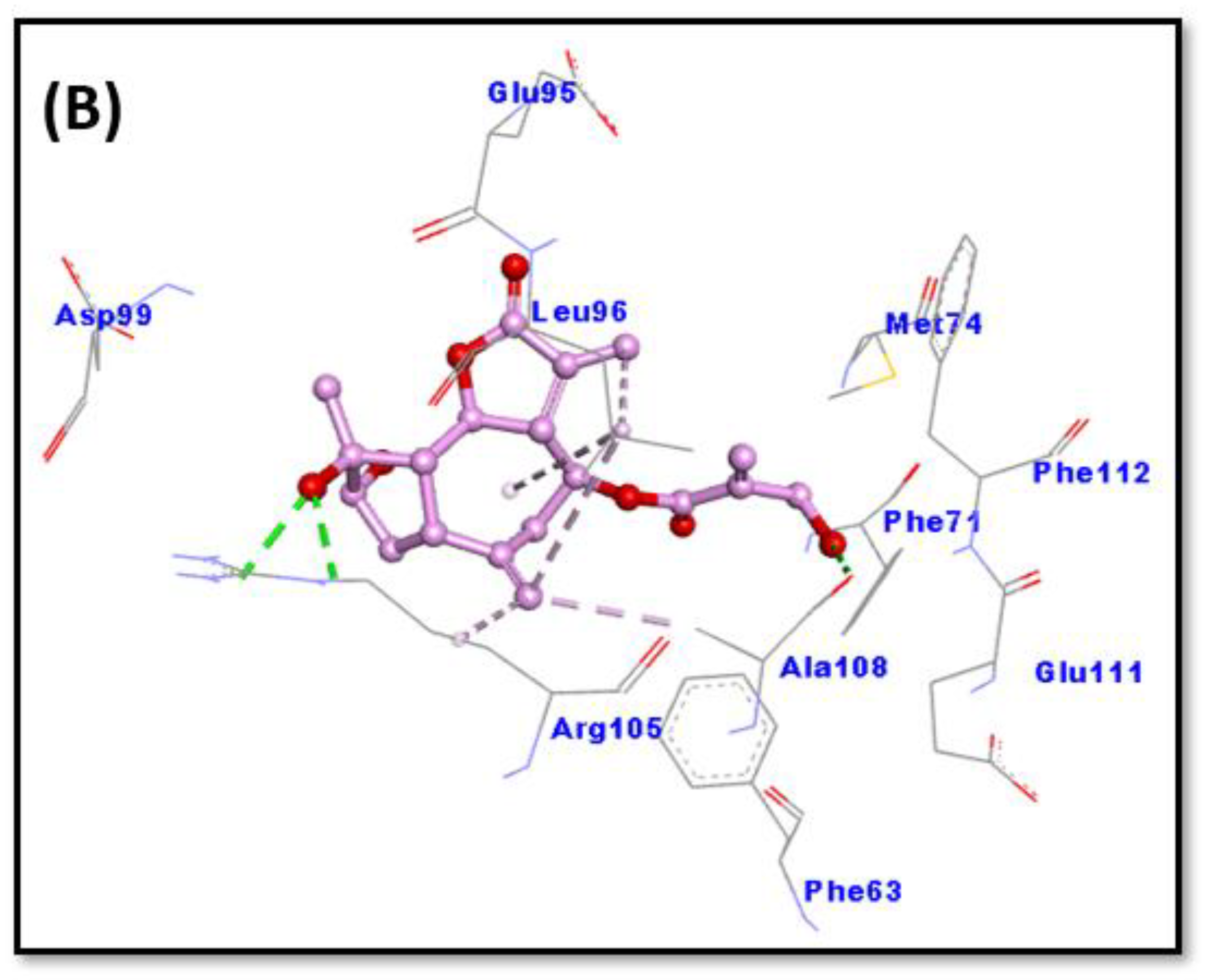
2.6. Molecular Docking
3. Discussion
4. Materials and Methods
4.1. Plant Collection, Extraction and Fractionation
4.2. Compound Isolation and Identification
4.3. Cell Culture Conditions
4.4. Cell Viability Assay
4.5. Cell Cycle Analysis
4.6. Detection of Apoptosis by Flow Cytometry (Annexin V-FITC/PI Staining)
4.7. Reverse Transcription Polymerase Chain Reaction (RT-PCR)
4.8. Western Blot Analysis
4.9. Molecular Docking Method
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, J.; Abbasi, B.A.; Mahmood, T.; Kanwal, S.; Ali, B.; Shah, S.A.; Khalil, A.T. Plant-derived anticancer agents: A green anticancer approach. Asian Pac. J. Trop. Biomed. 2017, 7, 1129–1150. [Google Scholar] [CrossRef]
- Greenwell, M.; Rahman, P.K. Medicinal Plants: Their Use in Anticancer Treatment. Int. J. Pharm. Sci. Res. 2015, 6, 4103–4112. [Google Scholar] [CrossRef] [PubMed]
- Dawud, M.A.-E.; Suad, A.-R. The flora of holy Mecca district, Saudi Arabia. Int. J. Biodivers. Conserv. 2015, 7, 173–189. [Google Scholar] [CrossRef] [Green Version]
- Al-Zahrani, A.A. Saudi anti-human cancer plants database (SACPD): A collection of plants with anti-human cancer activities. Oncol. Rev. 2018, 12, 349. [Google Scholar] [CrossRef] [Green Version]
- Wood, J.R.I.; Haig-Thomas, H. Handbook of the Yemen Flora; Royal Botanic Gardens, Kew: Richmond, UK, 1997. [Google Scholar]
- Collenette, S. Wildflowers of Saudi Arabia; National Commission for Wildlife Conservation and Development (NCWCD): London, UK, 1999.
- Ghazal, E. Taxonomic studies on the family asteraceae (Compositae) of Hajjah governorate, West of Yemen. J. Med. Plants Stud. 2019, 7, 90–100. [Google Scholar]
- Basahi, M.; Thomas, J.; Al-Turkiº, T.; Alfarhan, A.; Al-Atar, A.; Sivadasan, M. Current Status and Conservation Efforts of CEVT4. Uroth, 4 Mvus Maximus Wagenitz & Dittrich. Plant Arch. 2010, 10, 807–814. [Google Scholar]
- Muhammad, I.; Takamatsu, S.; Mossa, J.S.; El-Feraly, F.S.; Walker, L.A.; Clark, A.M. Cytotoxic sesquiterpene lactones from Centaurothamnus maximus and Vicoa pentanema. Phytother. Res. 2003, 17, 168–173. [Google Scholar] [CrossRef]
- Hussien, T.A.; Mohamed, T.A.; Elshamy, A.I.; Moustafa, M.F.; El-Seedi, H.R.; Pare, P.W.; Hegazy, M.-E.F. Guaianolide sesquiterpene lactones from Centaurothamnus maximus. Molecules 2021, 26, 2055. [Google Scholar] [CrossRef]
- Öksüz, S.; Serin, S.; Topçu, G. Sesquiterpene lactones from Centaurea hermannii. Phytochemistry 1994, 35, 435–438. [Google Scholar] [CrossRef]
- Medjroubi, K.; Benayache, F.; Bermejo, J. Sesquiterpene lactones from Centaurea musimomum. Antiplasmodial and cytotoxic activities. Fitoterapia 2005, 76, 744–746. [Google Scholar] [CrossRef]
- Gürbüz, I.; Yesilada, E. Evaluation of the anti-ulcerogenic effect of sesquiterpene lactones from Centaurea solstitialis L. ssp. solstitialis by using various in vivo and biochemical techniques. J. Ethnopharmacol. 2007, 112, 284–291. [Google Scholar] [CrossRef]
- Saklani, A.; Hegde, B.; Mishra, P.; Singh, R.; Mendon, M.; Chakrabarty, D.; Kamath, D.V.; Lobo, A.; Mishra, P.D.; Dagia, N.M.; et al. NF-κB dependent anti-inflammatory activity of chlorojanerin isolated from Saussurea heteromalla. Phytomedicine 2012, 19, 988–997. [Google Scholar] [CrossRef] [PubMed]
- Özçelik, B.; Gürbüz, I.; Karaoglu, T.; Yeşilada, E. Antiviral and antimicrobial activities of three sesquiterpene lactones from Centaurea solstitialis L. ssp. solstitialis. Microbiol. Res. 2009, 164, 545–552. [Google Scholar] [CrossRef] [PubMed]
- Tastan, P.; Hajdú, Z.; Kúsz, N.; Zupkó, I.; Sinka, I.; Kivcak, B.; Hohmann, J. Sesquiterpene Lactones and Flavonoids from Psephellus pyrrhoblepharus with Antiproliferative Activity on Human Gynecological Cancer Cell Lines. Molecules 2019, 24, 3165. [Google Scholar] [CrossRef] [Green Version]
- Al-Shabib, N.A.; Khan, J.M.; Malik, A.; Alsenaidy, M.A.; Rehman, M.T.; AlAjmi, M.F.; Alsenaidy, A.M.; Husain, F.M.; Khan, R.H. Molecular insight into binding behavior of polyphenol (rutin) with beta lactoglobulin: Spectroscopic, molecular docking and MD simulation studies. J. Mol. Liq. 2018, 269, 511–520. [Google Scholar] [CrossRef]
- Zhang, S.; Won, Y.K.; Ong, C.N.; Shen, H.M. Anti-cancer potential of sesquiterpene lactones: Bioactivity and molecular mechanisms. Curr. Med. Chem. Anticancer Agents 2005, 5, 239–249. [Google Scholar] [CrossRef]
- Kobayashi, T.; Ito, A.; Honda, H. Magnetic Nanoparticle-Mediated Hyperthermia and Induction of Anti-Tumor Immune Responses. In Hyperthermic Oncology from Bench to Bedside; Kokura, S., Yoshikawa, T., Ohnishi, T., Eds.; Springer: Singapore, 2016; pp. 137–150. [Google Scholar]
- Ahmed, M.Z.; Nasr, F.A.; Qamar, W.; Noman, O.M.; Khan, J.M.; Al-Mishari, A.A.; Alqahtani, A.S. Janerin Induces Cell Cycle Arrest at the G2/M Phase and Promotes Apoptosis Involving the MAPK Pathway in THP-1, Leukemic Cell Line. Molecules 2021, 26, 7555. [Google Scholar] [CrossRef]
- Suski, J.M.; Braun, M.; Strmiska, V.; Sicinski, P. Targeting cell-cycle machinery in cancer. Cancer Cell 2021, 39, 759–778. [Google Scholar] [CrossRef]
- Wang, Y.; Ji, P.; Liu, J.; Broaddus, R.R.; Xue, F.; Zhang, W. Centrosome-associated regulators of the G(2)/M checkpoint as targets for cancer therapy. Mol. Cancer 2009, 8, 1476–4598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- van Engeland, M.; Nieland, L.J.; Ramaekers, F.C.; Schutte, B.; Reutelingsperger, C.P. Annexin V-affinity assay: A review on an apoptosis detection system based on phosphatidylserine exposure. Cytometry 1998, 31, 1–9. [Google Scholar] [CrossRef]
- Campbell, K.J.; Tait, S.W.G. Targeting BCL-2 regulated apoptosis in cancer. Open Biol. 2018, 8, 180002. [Google Scholar] [CrossRef] [Green Version]
- Porter, A.G.; Jänicke, R.U. Emerging roles of caspase-3 in apoptosis. Cell Death Differ. 1999, 6, 99–104. [Google Scholar] [CrossRef]
- Sathishkumar, N.; Sathiyamoorthy, S.; Ramya, M.; Yang, D.U.; Lee, H.N.; Yang, D.C. Molecular docking studies of anti-apoptotic BCL-2, BCL-XL, and MCL-1 proteins with ginsenosides from Panax ginseng. J. Enzyme Inhib. Med. Chem. 2012, 27, 685–692. [Google Scholar] [CrossRef]
- D’Aguanno, S.; Del Bufalo, D. Inhibition of anti-apoptotic Bcl-2 proteins in preclinical and clinical studies: Current overview in cancer. Cells 2020, 9, 1287. [Google Scholar] [CrossRef]
- Porter, J.; Payne, A.; de Candole, B.; Ford, D.; Hutchinson, B.; Trevitt, G.; Turner, J.; Edwards, C.; Watkins, C.; Whitcombe, I. Tetrahydroisoquinoline amide substituted phenyl pyrazoles as selective Bcl-2 inhibitors. Bioorganic Med. Chem. Lett. 2009, 19, 230–233. [Google Scholar] [CrossRef] [PubMed]
- Rehman, M.T.; Shamsi, H.; Khan, A.U. Insight into the binding mechanism of imipenem to human serum albumin by spectroscopic and computational approaches. Mol. Pharm. 2014, 11, 1785–1797. [Google Scholar] [CrossRef]
- Rabbani, N.; Tabrez, S.; Islam, B.u.; Rehman, M.T.; Alsenaidy, A.M.; AlAjmi, M.F.; Khan, R.A.; Alsenaidy, M.A.; Khan, M.S. Characterization of colchicine binding with normal and glycated albumin: In vitro and molecular docking analysis. J. Biomol. Struct. Dyn. 2018, 36, 3453–3462. [Google Scholar] [CrossRef]
- Rehman, M.T.; Ahmed, S.; Khan, A.U. Interaction of meropenem with ‘N’and ‘B’ isoforms of human serum albumin: A spectroscopic and molecular docking study. J. Biomol. Struct. Dyn. 2016, 34, 1849–1864. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Cells and IC50 (µM) | ||
|---|---|---|---|
| A549 | LoVo | MCF-7 | |
| Chlorojanerin | 10.0 ± 0.4 | 15.9 ± 0.8 | 15.5 ± 0.7 |
| Doxorubicin | 1.58 ± 0.02 | 2.35 ± 0.05 | 1.74 ± 0.07 |
| Donor Atoms | Acceptor Atom | Distance (Å) | Type of Interaction | Binding Free Energy, ΔG (kcal mol−1) | Binding Affinity, Kd (M−1) |
|---|---|---|---|---|---|
| Bcl-2 and DRO * complex | |||||
| LIG:N ASP70:OD2 MET74:CE LEU96:CD1 LIG:C PHE63 PHE71 LIG:C LIG LIG LIG LIG LIG LIG | ASP70:OD2 LIG LIG LIG TYR67 LIG LIG LEU96 ARG105 ALA108 MET74 LEU96 ALA108 VAL92 | 2.9553 4.8135 3.5674 3.8681 3.5199 5.2880 4.9682 4.9539 5.2941 4.4414 5.4984 5.1674 4.4935 5.0873 | Conventional Hydrogen Bond Electrostatic (π-Anion) Hydrophobic (π-σ) Hydrophobic (π-σ) Hydrophobic (π-σ) Hydrophobic (π-π T-shaped) Hydrophobic (π-π T-shaped) Hydrophobic (Alkyl) Hydrophobic (π-Alkyl) Hydrophobic (π-Alkyl) Hydrophobic (π-Alkyl) Hydrophobic (π-Alkyl) Hydrophobic (π-Alkyl) Hydrophobic (π-Alkyl) | −10.2 | 3.03 × 107 |
| Bcl-2 and chlorojanerin complex | |||||
| ARG105:HE ARG105:HH21 LIG:O LEU96 ALA108 LIG:C LIG:C LIG:C | LIG:O LIG:O ALA108:O LIG LIG:C LEU96 ARG105 LEU96 | 2.5827 2.5827 2.7004 5.2020 3.6803 5.0962 4.1620 4.4392 | Conventional Hydrogen Bond Conventional Hydrogen Bond Conventional Hydrogen Bond Hydrophobic (Alkyl) Hydrophobic (Alkyl) Hydrophobic (Alkyl) Hydrophobic (Alkyl) Hydrophobic (π-Alkyl) | −6.5 | 5.85 × 104 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Noman, O.; Nasr, F.A.; Ahmed, M.Z.; Rehman, M.T.; Qamar, W.; Alqahtani, A.S.; Guenther, S. Assessment of the Anticancer Effect of Chlorojanerin Isolated from Centaurothamnus maximus on A549 Lung Cancer Cells. Molecules 2023, 28, 3061. https://doi.org/10.3390/molecules28073061
Noman O, Nasr FA, Ahmed MZ, Rehman MT, Qamar W, Alqahtani AS, Guenther S. Assessment of the Anticancer Effect of Chlorojanerin Isolated from Centaurothamnus maximus on A549 Lung Cancer Cells. Molecules. 2023; 28(7):3061. https://doi.org/10.3390/molecules28073061
Chicago/Turabian StyleNoman, Omar, Fahd A. Nasr, Mohammad Z. Ahmed, Md Tabish Rehman, Wajhul Qamar, Ali S. Alqahtani, and Sebastian Guenther. 2023. "Assessment of the Anticancer Effect of Chlorojanerin Isolated from Centaurothamnus maximus on A549 Lung Cancer Cells" Molecules 28, no. 7: 3061. https://doi.org/10.3390/molecules28073061