
Histamine and Tyramine in Chihuahua Cheeses during Shelf Life: Association with the Presence of tdc and hdc Genes

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Microbiological Quality of the Cheese Samples
2.2. Histamine and Tyramine Quantification
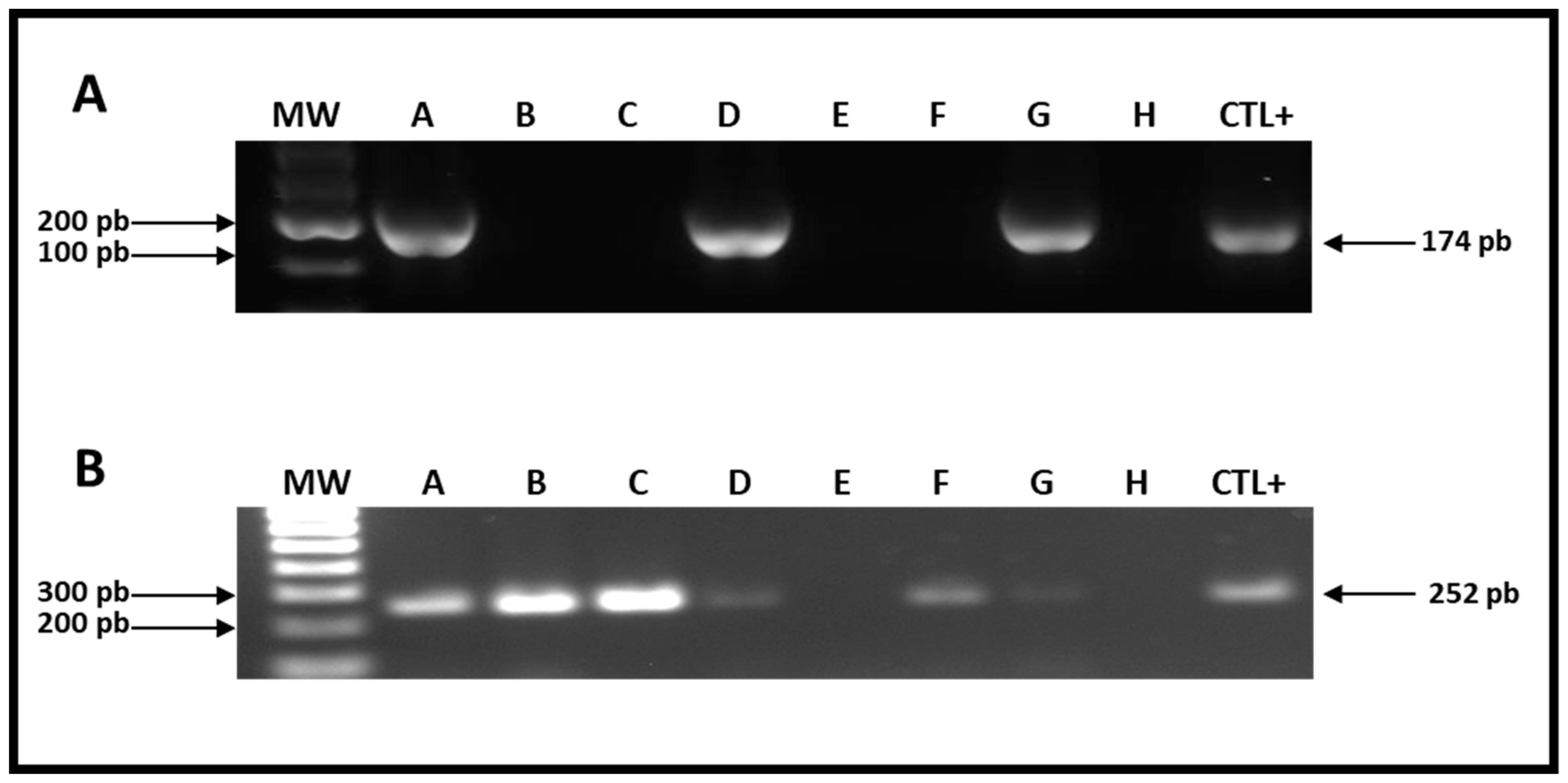
2.3. Detection of hdc and tdc Genes in Cheese Samples
3. Discussion
4. Materials and Methods
4.1. Sample Origin
4.2. Microbiological Analyses
4.3. Histamine and Tyramine Analysis
4.4. Molecular Biology Techniques Standardization
4.4.1. DNA Extraction
4.4.2. Amplification of Genes Encoding hdc and tdc
4.4.3. Design of Degenerated Oligonucleotides for the tdc Gene
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Renye, J.A.; Somkuti, G.A.; Van Hekken, D.L.; Guerrero Prieto, V.M. Characterization of Microflora in Mexican Chihuahua Cheese. J. Dairy Sci. 2011, 94, 3311–3315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Hekken, D.L.; Tunick, M.H.; Renye, J.A. The Character of Queso Chihuahua. ACS Symp. Ser. 2020, 1347, 142–160. [Google Scholar] [CrossRef]
- Sharma, R.; Garg, P.; Kumar, P.; Bhatia, S.K.; Kulshrestha, S. Microbial Fermentation and Its Role in Quality Improvement of Fermented Foods. Fermentation 2020, 6, 106. [Google Scholar] [CrossRef]
- Beresford, T.P.; Fitzsimons, N.A.; Brennan, N.L.; Cogan, T.M. Recent Advances in Cheese Microbiology. Int. Dairy J. 2001, 11, 259–274. [Google Scholar] [CrossRef]
- Özer, E.; Kesenkaş, H. The Effect of Using Different Starter Culture Combinations on Ripening Parameters, Microbiological and Sensory Properties of Mihaliç Cheese. J. Food Sci. Technol. 2019, 56, 1202. [Google Scholar] [CrossRef]
- Blaya, J.; Barzideh, Z.; LaPointe, G. Symposium Review: Interaction of Starter Cultures and Nonstarter Lactic Acid Bacteria in the Cheese Environment. J. Dairy Sci. 2018, 101, 3611–3629. [Google Scholar] [CrossRef]
- Dimou, A.; Tsimihodimos, V.; Bairaktari, E. The Critical Role of the Branched Chain Amino Acids (BCAAs) Catabolism-Regulating Enzymes, Branched-Chain Aminotransferase (BCAT) and Branched-Chain & alpha-Keto Acid Dehydrogenase (BCKD), in Human Pathophysiology. Int. J. Mol. Sci. 2022, 23, 4022. [Google Scholar]
- Ganesan, B.; Weimer, B.C. Amino Acid Catabolism and Its Relationship to Cheese Flavor Outcomes. Cheese Chem. Phys. Microbiol. Fourth Ed. 2017, 1, 483–516. [Google Scholar] [CrossRef]
- Linares, D.M.; del Río, B.; Ladero, V.; Martínez, N.; Fernández, M.; Martín, M.C.; Alvarez, M.A. Factors Influencing Biogenic Amines Accumulation in Dairy Products. Front. Microbiol. 2012, 3, 180. [Google Scholar] [CrossRef] [Green Version]
- Aliakbarlu, J.; Alizadeh, M.; Razavi-Rohani, S.M.; Vahabzade, Z.; Saei, S.S.; Agh, N. Effects of Processing Factors on Biogenic Amines Production in Iranian White Brine Cheese. Res. J. Biol. Sci. 2009, 4, 23–28. [Google Scholar]
- Fernández, M.; Linares, D.M.; Del Río, B.; Ladero, V.; Alvarez, M.A. HPLC Quantification of Biogenic Amines in Cheeses: Correlation with PCR-Detection of Tyramine-Producing Microorganisms. J. Dairy Res. 2007, 74, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Pinho, O.; Pintado, A.I.E.; Gomes, A.M.P.; Pintado, M.M.E.; Malcata, F.X.; Ferreira, I.M.P.L.V. Interrelationships among Microbiological, Physicochemical, and Biochemical Properties of Terrincho Cheese, with Emphasis on Biogenic Amines. J. Food Prot. 2004, 67, 2779–2785. [Google Scholar] [CrossRef] [PubMed]
- Standarová, E.; Vorlová, L.; Kordiovská, P.; Janštová, B.; Dračková, M.; Borkovcová, I. Biogenic Amine Production in Olomouc Curd Cheese (Olomoucké Tvarůžky) at Various Storage Conditions. Acta Vet. Brno 2010, 79, 147–156. [Google Scholar] [CrossRef] [Green Version]
- Lavizzari, T.; Veciana-Nogués, M.T.; Weingart, O.; Bover-Cid, S.; Mariné-Font, A.; Vidal-Carou, M.C. Occurrence of Biogenic Amines and Polyamines in Spinach and Changes during Storage under Refrigeration. J. Agric. Food Chem. 2007, 55, 9514–9519. [Google Scholar] [CrossRef]
- Mariné i Font, A. Les Amines Biògenes En Els Aliments: Història i Recerca En El Marc de Les Ciències de L’alimentació; Institut d’Estudis Catalans: Barcelona, Spain, 2005; ISBN 8472837882.
- Arena, M.E.; Manca de Nadra, M.C. Biogenic Amine Production by Lactobacillus. J. Appl. Microbiol. 2001, 90, 158–162. [Google Scholar] [CrossRef]
- Roig-Sagues, A.X.; Hernandez-Herrero, M.; Lopez-Sabater, E.I.; Rodriguez-Jerez, J.J.; Mora-Ventura, M.T. Histidine Decarboxylase Activity of Bacteria Isolated from Raw and Ripened Salchichon, a Spanish Cured Sausage. J. Food Prot. 1996, 59, 516–520. [Google Scholar] [CrossRef] [PubMed]
- Maintz, L.; Novak, N. Histamine and Histamine Intolerance. Am. J. Clin. Nutr. 2007, 85, 1185–1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbieri, F.; Montanari, C.; Gardini, F.; Tabanelli, G. Biogenic Amine Production by Lactic Acid Bacteria: A Review. Foods 2019, 8, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCabe-Sellers, B.J.; Staggs, C.G.; Bogle, M.L. Tyramine in Foods and Monoamine Oxidase Inhibitor Drugs: A Crossroad Where Medicine, Nutrition, Pharmacy, and Food Industry Converge. J. Food Compos. Anal. 2006, 19, S58–S65. [Google Scholar] [CrossRef]
- Rao, T.S.S.; Yeragani, V.K. Hypertensive Crisis and Cheese. Indian J. Psychiatry 2009, 51, 65–66. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations/World Health Organization. Public Health Risks of Histamine and Other Biogenic Amines from Fish and Fishery Products; Food and Agriculture Organization/World Health Organization: Rome, Italy, 2012; ISBN 9789251078495.
- Karovičová, J.; Kohajdová, Z. Biogenic Amines in Food. Chem. Pap. 2005, 59, 70–79. [Google Scholar]
- Van Hekken, D.L.; Tunick, M.H.; Tomasula, P.M.; Corral, F.J.M.; Gardea, A.A. Mexican Queso Chihuahua: Rheology of Fresh Cheese. Int. J. Dairy Technol. 2007, 60, 5–12. [Google Scholar] [CrossRef]
- Tunick, M.H.; Van Hekken, D.L.; Molina-Corral, F.J.; Tomasula, P.M.; Call, J.; Luchansky, J.; Gardea, A.A. Queso Chihuahua: Manufacturing Procedures, Composition, Protein Profiles, and Microbiology. Int. J. Dairy Technol. 2008, 61, 62–69. [Google Scholar] [CrossRef]
- Ventura, M.; Reniero, R.; Zink, R. Specific Identification and Targeted Characterization of Bifidobacterium lactis from Different Environmental Isolates by a Combined Multiplex-PCR Approach. Appl. Environ. Microbiol. 2001, 67, 2760–2765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández, M.; Del Río, B.; Linares, D.M.; Martín, M.C.; Alvarez, M.A. Real-Time Polymerase Chain Reaction for Quantitative Detection of Histamine-Producing Bacteria: Use in Cheese Production. J. Dairy Sci. 2006, 89, 3763–3769. [Google Scholar] [CrossRef] [Green Version]
- Izquierdo, P.; Allara, M.; Torres, G.; García, A.; Barboza, Y.; Piñero, M.I. Histamina En Quesos Madurados: Manchego, Parmesano y de Año. Rev. Cient. 2003, 13, 431–435. [Google Scholar]
- Burdychova, R.; Komprda, T. Biogenic Amine-Forming Microbial Communities in Cheese. FEMS Microbiol. Lett. 2007, 276, 149–155. [Google Scholar] [CrossRef]
- Tittarelli, F.; Perpetuini, G.; Di Gianvito, P.; Tofalo, R. Biogenic Amines Producing and Degrading Bacteria: A Snapshot from Raw Ewes’ Cheese. LWT 2019, 101, 1–9. [Google Scholar] [CrossRef]
- Novella-Rodríguez, S.; Veciana-Nogués, M.T.; Roig-Sagués, A.X.; Trujillo-Mesa, A.J.; Vidal-Carou, M.C. Evaluation of Biogenic Amines and Microbial Counts throughout the Ripening of Goat Cheeses from Pasteurized and Raw Milk. J. Dairy Res. 2004, 71, 245–252. [Google Scholar] [CrossRef]
- Diaz-Cinco, M.E.; Fraijo, O.; Grajeda, P.; Lozano-Taylor, J.; De Mejía, E.G. Microbial and Chemical Analysis of Chihuahua Cheese and Relationship to Histamine and Tyramine. J. Food Sci. 1992, 57, 355–356. [Google Scholar] [CrossRef]
- Standarová, E.; Borkovcová, I.; Dušková, M.; Přidalová, H.; Dračková, M. Effect of Some Factors on the Biogenic Amines and Polyamines Content in Blue-Veined Cheese Niva. Spec. Issue Czech J. Food Sci 2009, 27, S410–S413. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, E.M.A.; Amer, A.A.M. Comparison of Biogenic Amines Levels in Different Processed Cheese Varieties with Regulatory Specifications. World J. Dairy Food Sci. 2010, 5, 127–133. [Google Scholar]
- Roig-Sagués, A.X.; Molina, A.P.; Hernández-Herrero, M. Histamine and Tyramine-Forming Microorganisms in Spanish Traditional Cheeses. Eur. Food Res. Technol. 2002, 215, 96–100. [Google Scholar] [CrossRef]
- McCabe, B.J. Dietary Tyramine and Other Pressor Amines in MAOI Regimens: A Review. J. Am. Diet. Assoc. 1986, 86, 1059–1064. [Google Scholar] [CrossRef] [PubMed]
- Secretaría de Salud. NORMA Oficial Mexicana NOM-242-SSA1-2009, Productos y Servicios. Productos de la Pesca Frescos, Refrigerados, Congelados y Procesados. Especificaciones Sanitarias y Métodos de Prueba. Diario Oficial de la Federación. 2009. Available online: https://www.dof.gob.mx/normasOficiales/4295/salud2a/salud2a.htm (accessed on 24 February 2023).
- FDA (Food and Drug Administration). Hazards and Controls Guidance, 3rd ed.; Center of Food Safety and Nutrition: Washington, DC, USA, 2001.
- FDA (Food and Drug Administration). Fish and Fishery Products Hazards and Controls Guidance, 4th ed.; FDA: Silver Spring, MD, USA, 2020; pp. 113–152.
- Secretaría de Salud. PROYECTO de Norma Oficial Mexicana NOM-121-SSA1-1994, Bienes y Servicios. Quesos: Frescos, Ma-durados y Procesados. Especificaciones Sanitarias. Diario Oficial de la Federación. 1994. Available online: https://dof.gob.mx/nota_detalle.php?codigo=4729075&fecha=15/08/1994#gsc.tab=0 (accessed on 24 February 2023).
- Secretaría de Salud. NORMA Mexicana NMX-F-209-1985. Alimentos-Lácteos-Queso Tipo Chihuahua. Dirección General de Normas. 1985. Available online: http://cide.uach.mx/pdf/NORMAS%20MEXICANAS%20NMX/PRODUCTOS%20ALIMENTICIOS/ALIMENTOS.LACTEOS.QUESO%20TIPO%20CHIHUAHUA.pdf (accessed on 24 February 2023).
- Hamilton-Miller, J.; Shah, S.; Winkler, J. Public Health Issues Arising from Microbiological and Labelling Quality of Foods and Supplements Containing Probiotic Microorganisms. Public Health Nutr. 1999, 2, 223–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elsanhoty, R.M.; Mahrous, H.; Ghanaimy, G.A. Chemical, Microbial Counts and Evaluation of Biogenic Amines during the Ripening of Egyptian Soft Domiati Cheese Made from Raw and Pasteurized Buffaloes Milk. Int. J. Dairy Sci. 2009, 4, 80–90. [Google Scholar] [CrossRef] [Green Version]
- AOAC. Histamine in seafood: Fluorometric method Sec. 35.1.32, Method 977.13. In Official Methods of Analysis of AOAC International; Cunniff, P.A., Ed.; AOAC International: Gaithersburg, MD, USA, 1995; pp. 6–17. [Google Scholar]
- Hoffman, C.; Winston, F. A Ten-Minute DNA Preparation from Yeast Efficiently Releases Autonomous Plasmids for Transformation of Escherichia coli. Gene 1987, 57, 267–272. [Google Scholar] [CrossRef]
- Randazzo, C.L.; Torriani, S.; Akkermans, A.D.L.; De Vos, W.M.; Vaughan, E.E. Diversity, Dynamics, and Activity of Bacterial Communities during Production of an Artisanal Sicilian Cheese as Evaluated by 16S RRNA Analysis. Appl. Environ. Microbiol. 2002, 68, 1882–1892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sambrock, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning, A Laboratory Manual, 2nd ed.; Volumes 1, 2 and 3; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989; ISBN 0-87969-309-6. [Google Scholar]


{kind=link}
| Cheeses | Aerobic Mesophilic Log CFU/g | Total Coliforms Log CFU/g | LAB Log UFC/g |
|---|---|---|---|
| A | 3.7 ± 0.03 a | 3.3 ± 0.01 c | 9.0 ± 0.02 a |
| B | 3.1 ± 0.02 b | 3.1 ± 0.02 a | 8.3 ± 0.04 c |
| C | 3.1 ± 0.01 b | 3.3 ± 0.02 c | 8.3 ± 0.04 c |
| D | 3.6 ± 0.01 a | 3.3 ± 0.01 c | 8.6 ± 0.04 c |
| E | 3.7 ± 0.02 a | 2.6 ± 0.01 b | 7.6 ± 0.03 e |
| F | 3.4 ± 0.03 c | 3.3 ± 0.01 cd | 8.8 ± 0.04 b |
| G | 3.8 ± 0.02 e | 3.4 ± 0.02 d | 7.8 ± 0.03 d |
| H | 3.8 ± 0.02 e | 3.3 ± 0.03 cd | 7.8 ± 0.03 d |
| Cheeses | HDC | TDC |
|---|---|---|
| A | L. pentosus | L. pentosus, L. plantarum |
| B | ND | ND |
| C | ND | L. pentosus |
| D | ND | L. lactis sub lactis |
| E | ND | ND |
| F | ND | L. rhamnosus |
| G | L. pentosus | L. rhamnosus |
| H | ND | ND |
| Cheese | Histamine (mg/kg) | Tyramine (mg/kg) | ||||
|---|---|---|---|---|---|---|
| Shelf Life | Shelf Life | |||||
| Start | Half | End | Start | Half | End | |
| A | ND | 45 ± 4 | 192 ± 20 | ND | 59 ± 15 | 115 ± 10 |
| B | ND | ND | ND | 122 ± 12 | 160 ± 18 | 209 ± 2 |
| C | ND | ND | ND | 34 ± 2 | 160 ± 17 | 166 ± 6 |
| D | ND | ND | 64 ± 11 | ND | 147 ± 4 | 194 ± 24 |
| E | ND | ND | ND | ND | ND | ND |
| F | ND | ND | ND | 42 ± 8 | 139 ± 9 | 150 ± 1 |
| G | ND | ND | 46 ± 6 | ND | ND | 205 ± 4 |
| H | ND | ND | ND | ND | ND | ND |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Campos-Góngora, E.; González-Martínez, M.T.; López-Hernández, A.A.; Arredondo-Mendoza, G.I.; Ortega-Villarreal, A.S.; González-Martínez, B.E. Histamine and Tyramine in Chihuahua Cheeses during Shelf Life: Association with the Presence of tdc and hdc Genes. Molecules 2023, 28, 3007. https://doi.org/10.3390/molecules28073007
Campos-Góngora E, González-Martínez MT, López-Hernández AA, Arredondo-Mendoza GI, Ortega-Villarreal AS, González-Martínez BE. Histamine and Tyramine in Chihuahua Cheeses during Shelf Life: Association with the Presence of tdc and hdc Genes. Molecules. 2023; 28(7):3007. https://doi.org/10.3390/molecules28073007
Chicago/Turabian StyleCampos-Góngora, Eduardo, María Teresa González-Martínez, Abad Arturo López-Hernández, Gerardo Ismael Arredondo-Mendoza, Ana Sofía Ortega-Villarreal, and Blanca Edelia González-Martínez. 2023. "Histamine and Tyramine in Chihuahua Cheeses during Shelf Life: Association with the Presence of tdc and hdc Genes" Molecules 28, no. 7: 3007. https://doi.org/10.3390/molecules28073007