

Rac1: A Regulator of Cell Migration and a Potential Target for Cancer Therapy

Abstract
:1. Introduction
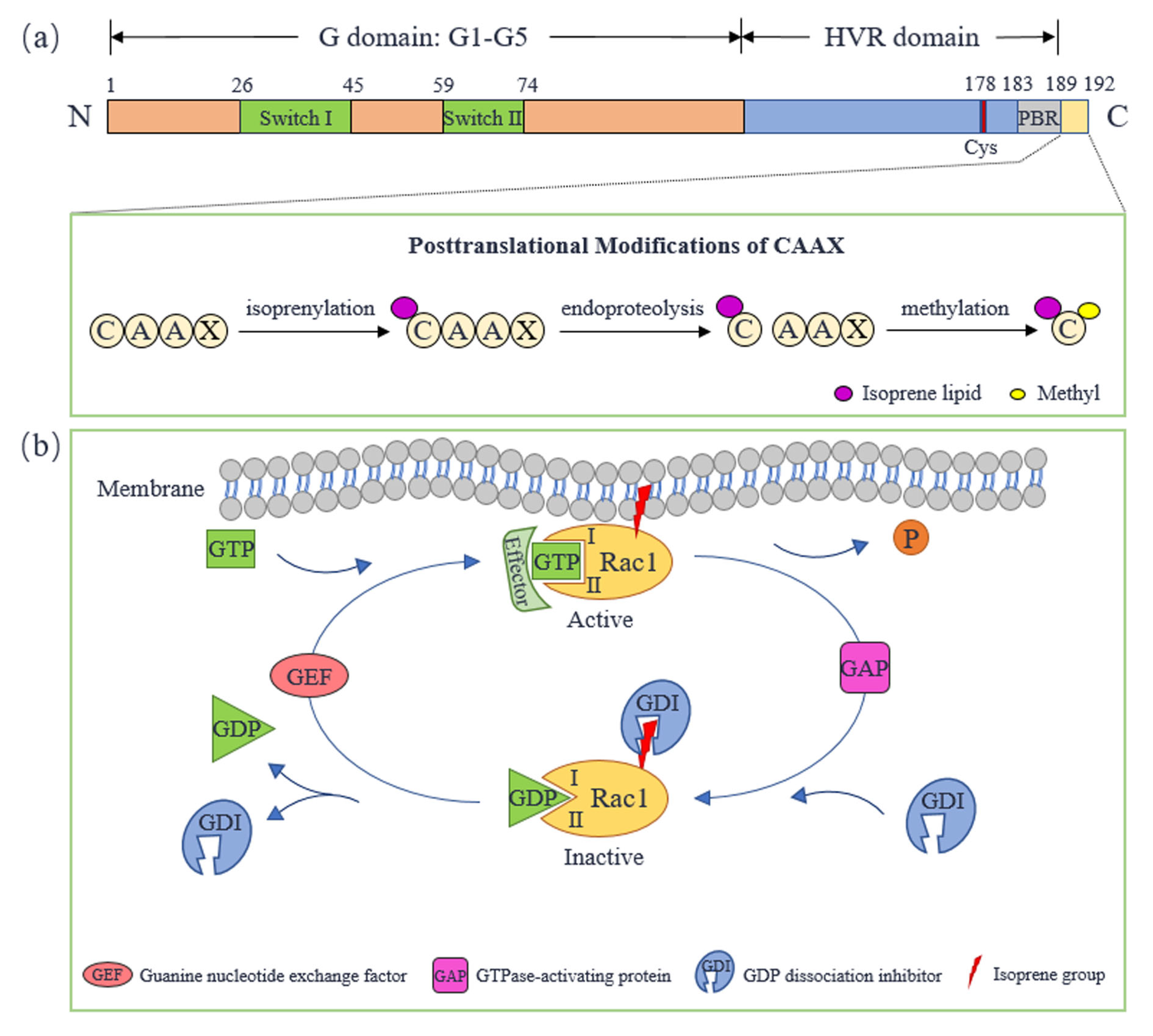
2. Molecular Structure and Activity Regulation of Rac1
3. Rac1 Regulates Cell Migration and Tumor Invasion and Metastasis
3.1. Rac1 Regulates Invasion and Metastasis of Tumor Cells
3.2. Rac1 Regulates Cell Migration in Angiogenesis
3.3. Rac1 Regulates Cell Migration in Immune Response
3.4. Rac1 Regulates Cell Migration in Other Physiological and Pathological Processes
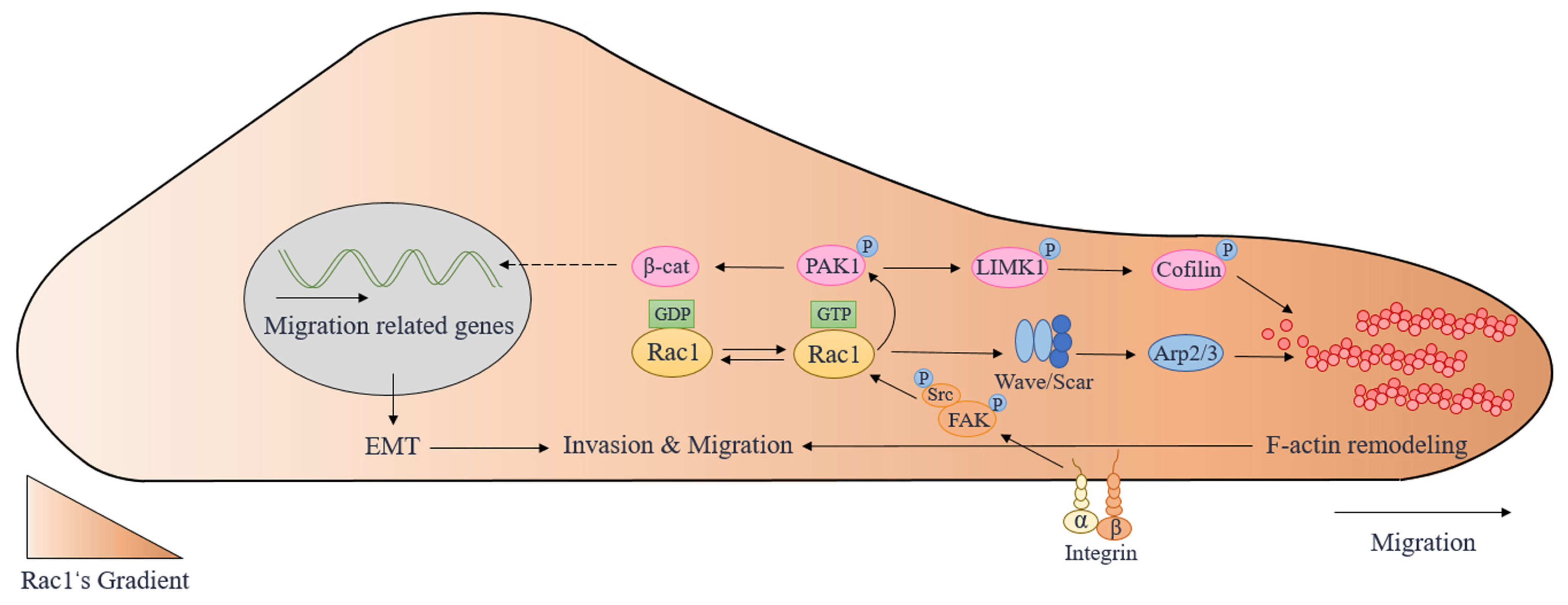
4. Molecular Mechanisms of Cell Migration Regulated by Rac1
4.1. Rac1 Regulates Cell Front–Rear Polarity
4.2. Rac1-Scar/Wave-Arp2/3 Signaling Pathway
4.3. Rac1/PAK1 Signaling Pathway
4.4. Src/FAK/Rac1 Signaling Pathway
5. Inhibition of Rac1 Activity in Cancer Therapy
6. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Trepat, X.; Chen, Z.; Jacobson, K. Cell Migration. Compr. Physiol. 2012, 2, 2369–2392. [Google Scholar] [CrossRef] [Green Version]
- Stock, J.; Pauli, A. Self-Organized Cell Migration Across Scales-From Single Cell Movement to Tissue Formation. Development 2021, 148, dev191767. [Google Scholar] [CrossRef] [PubMed]
- SenGupta, S.; Parent, C.A.; Bear, J.E. The Principles of Directed Cell Migration. Nat. Rev. Mol. Cell Biol. 2021, 22, 529–547. [Google Scholar] [CrossRef] [PubMed]
- Shellard, A.; Mayor, R. All Roads Lead to Directional Cell Migration. Trends Cell Biol. 2020, 30, 852–868. [Google Scholar] [CrossRef] [PubMed]
- Ecker, N.; Kruse, K. Excitable Actin Dynamics and Amoeboid Cell Migration. PLoS ONE 2021, 16, e246311. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Oyang, L.; Rao, S.; Han, Y.; Luo, X.; Yi, P.; Lin, J.; Xia, L.; Hu, J.; Tan, S.; et al. Rac1, a Potential Target for Tumor Therapy. Front. Oncol. 2021, 11, 674426. [Google Scholar] [CrossRef] [PubMed]
- Bailly, C.; Beignet, J.; Loirand, G.; Sauzeau, V. Rac1 as a Therapeutic Anticancer Target: Promises and Limitations. Biochem. Pharmacol. 2022, 203, 115180. [Google Scholar] [CrossRef]
- Polakis, P.G.; Weber, R.F.; Nevins, B.; Didsbury, J.R.; Evans, T.; Snyderman, R. Identification of the Ral and Rac1 Gene Products, Low Molecular Mass GTP-binding Proteins From Human Platelets. J. Biol. Chem. 1989, 264, 16383–16389. [Google Scholar] [CrossRef]
- Didsbury, J.; Weber, R.F.; Bokoch, G.M.; Evans, T.; Snyderman, R. Rac, a Novel Ras-Related Family of Proteins that are Botulinum Toxin Substrates. J. Biol. Chem. 1989, 264, 16378–16382. [Google Scholar] [CrossRef]
- Matos, P.; Skaug, J.; Marques, B.; Beck, S.; Verissimo, F.; Gespach, C.; Boavida, M.G.; Scherer, S.W.; Jordan, P. Small GTPase Rac1: Structure, Localization, and Expression of the Human Gene. Biochem. Biophys. Res. Commun. 2000, 277, 741–751. [Google Scholar] [CrossRef]
- Mosaddeghzadeh, N.; Ahmadian, M.R. The RHO Family GTPases: Mechanisms of Regulation and Signaling. Cells 2021, 10, 1831. [Google Scholar] [CrossRef] [PubMed]
- Wittinghofer, A.; Vetter, I.R. Structure-Function Relationships of the G Domain, a Canonical Switch Motif. Annu. Rev. Biochem. 2011, 80, 943–971. [Google Scholar] [CrossRef]
- Vetter, I.R.; Wittinghofer, A. The Guanine Nucleotide-Binding Switch in Three Dimensions. Science 2001, 294, 1299–1304. [Google Scholar] [CrossRef] [Green Version]
- De, P.; Aske, J.C.; Dey, N. RAC1 Takes the Lead in Solid Tumors. Cells 2019, 8, 382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, P.J.; Mitin, N.; Keller, P.J.; Chenette, E.J.; Madigan, J.P.; Currin, R.O.; Cox, A.D.; Wilson, O.; Kirschmeier, P.; Der, C.J. Rho Family GTPase Modification and Dependence On CAAX Motif-Signaled Posttranslational Modification. J. Biol. Chem. 2008, 283, 25150–25163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinsella, B.T.; Erdman, R.A.; Maltese, W.A. Carboxyl-Terminal Isoprenylation of Ras-Related GTP-binding Proteins Encoded by Rac1, Rac2, and ralA. J. Biol. Chem. 1991, 266, 9786–9794. [Google Scholar] [CrossRef]
- Kowluru, A. Tiam1/Vav2-Rac1 Axis: A Tug-of-war Between Islet Function and Dysfunction. Biochem. Pharmacol. 2017, 132, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Hernan, R.; Marcogliese, P.C.; Huang, Y.; Gertler, T.S.; Akcaboy, M.; Liu, S.; Chung, H.L.; Pan, X.; Sun, X.; et al. Loss-Of-Function Variants in TIAM1 are Associated with Developmental Delay, Intellectual Disability, and Seizures. Am. J. Hum. Genet. 2022, 109, 571–586. [Google Scholar] [CrossRef]
- Payapilly, A.; Guilbert, R.; Descamps, T.; White, G.; Magee, P.; Zhou, C.; Kerr, A.; Simpson, K.L.; Blackhall, F.; Dive, C.; et al. TIAM1-RAC1 Promote Small-Cell Lung Cancer Cell Survival through Antagonizing Nur77-Induced BCL2 Conformational Change. Cell Rep. 2021, 37, 109979. [Google Scholar] [CrossRef]
- Paskus, J.D.; Herring, B.E.; Roche, K.W. Kalirin and Trio: RhoGEFs in Synaptic Transmission, Plasticity, and Complex Brain Disorders. Trends Neurosci. 2020, 43, 505–518. [Google Scholar] [CrossRef]
- Wang, J.; Yuan, L.; Xu, X.; Zhang, Z.; Ma, Y.; Hong, L.; Ma, J. Rho-GEF Trio Regulates Osteosarcoma Progression and Osteogenic Differentiation through Rac1 and RhoA. Cell Death Dis. 2021, 12, 1148. [Google Scholar] [CrossRef]
- Um, K.; Niu, S.; Duman, J.G.; Cheng, J.X.; Tu, Y.K.; Schwechter, B.; Liu, F.; Hiles, L.; Narayanan, A.S.; Ash, R.T.; et al. Dynamic Control of Excitatory Synapse Development by a Rac1 GEF/GAP Regulatory Complex. Dev. Cell 2014, 29, 701–715. [Google Scholar] [CrossRef] [Green Version]
- Smith, K.R.; Rajgor, D.; Hanley, J.G. Differential Regulation of the Rac1 GTPase-activating Protein (GAP) BCR during Oxygen/Glucose Deprivation in Hippocampal and Cortical Neurons. J. Biol. Chem. 2017, 292, 20173–20183. [Google Scholar] [CrossRef] [Green Version]
- Bustelo, X.R.; Sauzeau, V.; Berenjeno, I.M. GTP-binding Proteins of the Rho/Rac Family: Regulation, Effectors and Functions in Vivo. Bioessays 2007, 29, 356–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tejada-Simon, M.V. Modulation of Actin Dynamics by Rac1 to Target Cognitive Function. J. Neurochem. 2015, 133, 767–779. [Google Scholar] [CrossRef]
- Dvorsky, R.; Ahmadian, M.R. Always Look On the Bright Site of Rho: Structural Implications for a Conserved Intermolecular Interface. EMBO Rep. 2004, 5, 1130–1136. [Google Scholar] [CrossRef] [Green Version]
- Machacek, M.; Hodgson, L.; Welch, C.; Elliott, H.; Pertz, O.; Nalbant, P.; Abell, A.; Johnson, G.L.; Hahn, K.M.; Danuser, G. Coordination of Rho GTPase Activities During Cell Protrusion. Nature 2009, 461, 99–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grubisha, M.J.; DeGiosio, R.A.; Wills, Z.P.; Sweet, R.A. Trio and Kalirin as Unique Enactors of Rho/Rac Spatiotemporal Precision. Cell. Signal. 2022, 98, 110416. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, M.; Horio, R.; Wang, L.; Takenami, T.; Moriya, J.; Suzuka, J.; Sugino, H.; Tanei, Z.; Tanino, M.; Tanaka, S. Novel Rapid Immunohistochemistry Using an Alternating Current Electric Field Identifies Rac and Cdc42 Activation in Human Colon Cancer FFPE Tissues. Sci. Rep. 2022, 12, 1733. [Google Scholar] [CrossRef]
- Wu, Y.I.; Frey, D.; Lungu, O.I.; Jaehrig, A.; Schlichting, I.; Kuhlman, B.; Hahn, K.M. A Genetically Encoded Photoactivatable Rac Controls the Motility of Living Cells. Nature 2009, 461, 104–108. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.; Li, P.; Liu, J.; Liao, J.; Li, H.; Chen, L.; Li, Z.; Guo, Q.; Belguise, K.; Yi, B.; et al. Two Rac1 Pools Integrate the Direction and Coordination of Collective Cell Migration. Nat. Commun. 2022, 13, 6014. [Google Scholar] [CrossRef]
- de Beco, S.; Vaidziulyte, K.; Manzi, J.; Dalier, F.; di Federico, F.; Cornilleau, G.; Dahan, M.; Coppey, M. Optogenetic Dissection of Rac1 and Cdc42 Gradient Shaping. Nat. Commun. 2018, 9, 4816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehidi, A.; Rossier, O.; Schaks, M.; Chazeau, A.; Biname, F.; Remorino, A.; Coppey, M.; Karatas, Z.; Sibarita, J.B.; Rottner, K.; et al. Transient Activations of Rac1 at the Lamellipodium Tip Trigger Membrane Protrusion. Curr. Biol. 2019, 29, 2852–2866. [Google Scholar] [CrossRef]
- Liu, M.; Banerjee, R.; Rossa, C.J.; D’Silva, N.J. RAP1-RAC1 Signaling Has an Important Role in Adhesion and Migration in HNSCC. J. Dent. Res. 2020, 99, 959–968. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.C.; Liu, C.X.; Pan, W.D.; Shang, L.R.; Zheng, J.L.; Huang, B.Y.; Chen, J.Y.; Zheng, L.; Fang, J.H.; Zhuang, S.M. Dual and Opposing Roles of the Androgen Receptor in VETC-dependent and Invasion-Dependent Metastasis of Hepatocellular Carcinoma. J. Hepatol. 2021, 75, 900–911. [Google Scholar] [CrossRef]
- Swaminathan, K.; Campbell, A.; Papalazarou, V.; Jaber-Hijazi, F.; Nixon, C.; McGhee, E.; Strathdee, D.; Sansom, O.J.; Machesky, L.M. The RAC1 Target NCKAP1 Plays a Crucial Role in the Progression of Braf; Pten-Driven Melanoma in Mice. J. Investig. Dermatol. 2021, 141, 628–637. [Google Scholar] [CrossRef]
- Guo, Y.; Zhu, J.; Wang, X.; Li, R.; Jiang, K.; Chen, S.; Fan, J.; Xue, L.; Hao, D. Orai1 Promotes Osteosarcoma Metastasis by Activating the Ras-Rac1-WAVE2 Signaling Pathway. Med. Sci. Monit. 2019, 25, 9227–9236. [Google Scholar] [CrossRef]
- Pham, T.Q.; Robinson, K.; Xu, L.; Pavlova, M.N.; Skapek, S.X.; Chen, E.Y. HDAC6 Promotes Growth, Migration/Invasion, and Self-Renewal of Rhabdomyosarcoma. Oncogene 2021, 40, 578–591. [Google Scholar] [CrossRef]
- Kalli, M.; Li, R.; Mills, G.B.; Stylianopoulos, T.; Zervantonakis, I.K. Mechanical Stress Signaling in Pancreatic Cancer Cells Triggers p38 MAPK- and JNK-Dependent Cytoskeleton Remodeling and Promotes Cell Migration via Rac1/cdc42/Myosin II. Mol. Cancer Res. 2022, 20, 485–497. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Lerida, I.; Pellinen, T.; Sanchez, S.A.; Guadamillas, M.C.; Wang, Y.; Mirtti, T.; Calvo, E.; Del, P.M. Rac1 Nucleocytoplasmic Shuttling Drives Nuclear Shape Changes and Tumor Invasion. Dev. Cell 2015, 32, 318–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Gu, L.; Wu, N.; Song, J.; Yan, J.; Yang, S.; Feng, Y.; Wang, Z.; Wang, L.; Zhang, Y.; et al. Overexpression of DTL Enhances Cell Motility and Promotes Tumor Metastasis in Cervical Adenocarcinoma by Inducing RAC1-JNK-FOXO1 Axis. Cell Death Dis. 2021, 12, 929. [Google Scholar] [CrossRef] [PubMed]
- Du, G.S.; Qiu, Y.; Wang, W.S.; Peng, K.; Zhang, Z.C.; Li, X.S.; Xiao, W.D.; Yang, H. Knockdown On aPKC-iota Inhibits Epithelial-Mesenchymal Transition, Migration and Invasion of Colorectal Cancer Cells through Rac1-JNK Pathway. Exp. Mol. Pathol. 2019, 107, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Min, P.; Liu, L.; Zhang, L.; Zhang, Y.; Wang, Y.; Zhao, X.; Ma, Y.; Xie, H.; Zhu, C.; et al. NEDD9 Facilitates Hypoxia-Induced Gastric Cancer Cell Migration via MICAL1 Related Rac1 Activation. Front. Pharmacol. 2019, 10, 291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Bibi, M.; Min, P.; Deng, W.; Zhang, Y.; Du, J. SOX2 Promotes Hypoxia-Induced Breast Cancer Cell Migration by Inducing NEDD9 Expression and Subsequent Activation of Rac1/HIF-1alpha Signaling. Cell. Mol. Biol. Lett. 2019, 24, 55. [Google Scholar] [CrossRef] [Green Version]
- Pu, J.; Huang, Y.; Fang, Q.; Wang, J.; Li, W.; Xu, Z.; Wu, X.; Lu, Y.; Wei, H. Hypoxia-Induced Fascin-1 Upregulation is Regulated by Akt/Rac1 Axis and Enhances Malignant Properties of Liver Cancer Cells Via Mediating Actin Cytoskeleton Rearrangement and Hippo/YAP Activation. Cell Death Discov. 2021, 7, 385. [Google Scholar] [CrossRef]
- Liu, X.; Wang, Q.; Liu, B.; Zheng, X.; Li, P.; Zhao, T.; Jin, X.; Ye, F.; Zhang, P.; Chen, W.; et al. Genistein Inhibits Radiation-Induced Invasion and Migration of Glioblastoma Cells by Blocking the DNA-PKcs/Akt2/Rac1 Signaling Pathway. Radiother. Oncol. 2021, 155, 93–104. [Google Scholar] [CrossRef]
- Wang, M.; Niu, J.; Gao, L.; Gao, Y.; Gao, S. Zerumbone Inhibits Migration in ESCC Via Promoting Rac1 Ubiquitination. Biomed. Pharmacother. 2019, 109, 2447–2455. [Google Scholar] [CrossRef]
- Liu, L.; Cui, J.; Zhao, Y.; Liu, X.; Chen, L.; Xia, Y.; Wang, Y.; Chen, S.; Sun, S.; Shi, B.; et al. KDM6A-ARHGDIB Axis Blocks Metastasis of Bladder Cancer by Inhibiting Rac1. Mol. Cancer 2021, 20, 77. [Google Scholar] [CrossRef]
- Luttgenau, S.M.; Emming, C.; Wagner, T.; Harms, J.; Guske, J.; Weber, K.; Neugebauer, U.; Schroter, R.; Panichkina, O.; Petho, Z.; et al. Pals1 Prevents Rac1-dependent Colorectal Cancer Cell Metastasis by Inhibiting Arf6. Mol. Cancer 2021, 20, 74. [Google Scholar] [CrossRef]
- Rong, Z.; Cheng, B.; Zhong, L.; Ye, X.; Li, X.; Jia, L.; Li, Y.; Shue, F.; Wang, N.; Cheng, Y.; et al. Activation of FAK/Rac1/Cdc42-GTPase Signaling Ameliorates Impaired Microglial Migration Response to Abeta(42) in Triggering Receptor Expressed On Myeloid Cells 2 Loss-Of-Function Murine Models. FASEB J. 2020, 34, 10984–10997. [Google Scholar] [CrossRef]
- Li, Y.; Niu, M.; Zhao, A.; Kang, W.; Chen, Z.; Luo, N.; Zhou, L.; Zhu, X.; Lu, L.; Liu, J. CXCL12 is Involved in Alpha-Synuclein-Triggered Neuroinflammation of Parkinson’s Disease. J. Neuroinflamm. 2019, 16, 263. [Google Scholar] [CrossRef] [Green Version]
- Fionda, C.; Stabile, H.; Molfetta, R.; Kosta, A.; Peruzzi, G.; Ruggeri, S.; Zingoni, A.; Capuano, C.; Soriani, A.; Paolini, R.; et al. Cereblon Regulates NK Cell Cytotoxicity and Migration Via Rac1 Activation. Eur. J. Immunol. 2021, 51, 2607–2617. [Google Scholar] [CrossRef] [PubMed]
- Ramadass, M.; Johnson, J.L.; Marki, A.; Zhang, J.; Wolf, D.; Kiosses, W.B.; Pestonjamasp, K.; Ley, K.; Catz, S.D. The Trafficking Protein JFC1 Regulates Rac1-GTP Localization at the Uropod Controlling Neutrophil Chemotaxis and in Vivo Migration. J. Leukoc. Biol. 2019, 105, 1209–1224. [Google Scholar] [CrossRef]
- Kanauchi, Y.; Yamamoto, T.; Yoshida, M.; Zhang, Y.; Lee, J.; Hayashi, S.; Kadowaki, M. Cholinergic Anti-Inflammatory Pathway Ameliorates Murine Experimental Th2-type Colitis by Suppressing the Migration of Plasmacytoid Dendritic Cells. Sci. Rep. 2022, 12, 54. [Google Scholar] [CrossRef]
- Bu, F.; Min, J.W.; Munshi, Y.; Lai, Y.J.; Qi, L.; Urayama, A.; McCullough, L.D.; Li, J. Activation of Endothelial Ras-Related C3 Botulinum Toxin Substrate 1 (Rac1) Improves Post-Stroke Recovery and Angiogenesis Via Activating Pak1 in Mice. Exp. Neurol. 2019, 322, 113059. [Google Scholar] [CrossRef] [PubMed]
- Barbera, S.; Lugano, R.; Pedalina, A.; Mongiat, M.; Santucci, A.; Tosi, G.M.; Dimberg, A.; Galvagni, F.; Orlandini, M. The C-type Lectin CD93 Controls Endothelial Cell Migration Via Activation of the Rho Family of Small GTPases. Matrix Biol. 2021, 99, 1–17. [Google Scholar] [CrossRef]
- Wang, H.; Ramshekar, A.; Kunz, E.; Sacks, D.B.; Hartnett, M.E. IQGAP1 Causes Choroidal Neovascularization by Sustaining VEGFR2-mediated Rac1 Activation. Angiogenesis 2020, 23, 685–698. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ramshekar, A.; Kunz, E.; Hartnett, M.E. 7-Ketocholesterol Induces Endothelial-Mesenchymal Transition and Promotes Fibrosis: Implications in Neovascular Age-Related Macular Degeneration and Treatment. Angiogenesis 2021, 24, 583–595. [Google Scholar] [CrossRef]
- Wang, X.; Yao, Y.; Gao, J. Sevoflurane Inhibits Growth Factor-Induced Angiogenesis through Suppressing Rac1/paxillin/FAK and Ras/Akt/mTOR. Future Oncol. 2020, 16, 1619–1627. [Google Scholar] [CrossRef]
- Tang, H.; Wang, X.; Zhang, M.; Yan, Y.; Huang, S.; Ji, J.; Xu, J.; Zhang, Y.; Cai, Y.; Yang, B.; et al. MicroRNA-200b/c-3p Regulate Epithelial Plasticity and Inhibit Cutaneous Wound Healing by Modulating TGF-beta-mediated RAC1 Signaling. Cell Death Dis. 2020, 11, 931. [Google Scholar] [CrossRef]
- Sawaya, A.P.; Jozic, I.; Stone, R.C.; Pastar, I.; Egger, A.N.; Stojadinovic, O.; Glinos, G.D.; Kirsner, R.S.; Tomic-Canic, M. Mevastatin Promotes Healing by Targeting Caveolin-1 to Restore EGFR Signaling. JCI Insight 2019, 4, e129320. [Google Scholar] [CrossRef] [Green Version]
- Gu, K.; Liu, G.; Wu, C.; Jia, G.; Zhao, H.; Chen, X.; Tian, G.; Cai, J.; Zhang, R.; Wang, J. Tryptophan Improves Porcine Intestinal Epithelial Cell Restitution through the CaSR/Rac1/PLC-gamma1 Signaling Pathway. Food Funct. 2021, 12, 8787–8799. [Google Scholar] [CrossRef] [PubMed]
- Jafari, A.; Isa, A.; Chen, L.; Ditzel, N.; Zaher, W.; Harkness, L.; Johnsen, H.E.; Abdallah, B.M.; Clausen, C.; Kassem, M. TAFA2 Induces Skeletal (Stromal) Stem Cell Migration through Activation of Rac1-P38 Signaling. Stem Cells 2019, 37, 407–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Jiang, X.; Zhang, C.; Zhong, J.; Fang, X.; Li, H.; Xie, F.; Huang, X.; Zhang, X.; Hu, Q.; et al. Actin Alpha 2 (ACTA2) Downregulation Inhibits Neural Stem Cell Migration through Rho GTPase Activation. Stem Cells Int. 2020, 2020, 4764012. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Li, B.; Wang, J.; Lu, Y.; Chen, A.; Sun, K.; Yu, Y.; Chen, S. GAA Deficiency Promotes Angiogenesis through Upregulation of Rac1 Induced by Autophagy Disorder. Biochim. Biophys. Acta Mol. Cell Res. 2021, 1868, 118969. [Google Scholar] [CrossRef]
- Uemura, A.; Fukushima, Y. Rho GTPases in Retinal Vascular Diseases. Int. J. Mol. Sci. 2021, 22, 3684. [Google Scholar] [CrossRef]
- Ramshekar, A.; Wang, H.; Hartnett, M.E. Regulation of Rac1 Activation in Choroidal Endothelial Cells: Insights into Mechanisms in Age-Related Macular Degeneration. Cells 2021, 10, 2414. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Chen, J.; Shi, J.; Zhou, W.; Wang, L.; Fang, W.; Zhong, Y.; Chen, X.; Chen, Y.; Sabri, A.; et al. M1-Like Macrophage-Derived Exosomes Suppress Angiogenesis and Exacerbate Cardiac Dysfunction in a Myocardial Infarction Microenvironment. Basic Res. Cardiol. 2020, 115, 22. [Google Scholar] [CrossRef]
- Khan, S.; Shukla, S.; Farhan, M.; Sinha, S.; Lakra, A.D.; Penta, D.; Kannan, A.; Meeran, S.M. Centchroman Prevents Metastatic Colonization of Breast Cancer Cells and Disrupts Angiogenesis Via Inhibition of RAC1/PAK1/beta-catenin Signaling Axis. Life Sci. 2020, 256, 117976. [Google Scholar] [CrossRef]
- Mamula, D.; Korthals, M.; Hradsky, J.; Gottfried, A.; Fischer, K.D.; Tedford, K. Arhgef6 (alpha-PIX) Cytoskeletal Regulator Signals to GTPases and Cofilin to Couple T Cell Migration Speed and Persistence. J. Leukoc. Biol. 2021, 110, 839–852. [Google Scholar] [CrossRef]
- Arts, J.J.; Mahlandt, E.K.; Gronloh, M.L.; Schimmel, L.; Noordstra, I.; Gordon, E.; van Steen, A.C.; Tol, S.; Walzog, B.; van Rijssel, J.; et al. Endothelial Junctional Membrane Protrusions Serve as Hotspots for Neutrophil Transmigration. Elife 2021, 10, e66074. [Google Scholar] [CrossRef]
- Glemain, A.; Neel, M.; Neel, A.; Andre-Gregoire, G.; Gavard, J.; Martinet, B.; Le Bloas, R.; Riquin, K.; Hamidou, M.; Fakhouri, F.; et al. Neutrophil-Derived Extracellular Vesicles Induce Endothelial Inflammation and Damage through the Transfer of miRNAs. J. Autoimmun. 2022, 129, 102826. [Google Scholar] [CrossRef]
- Movassagh, H.; Shan, L.; Koussih, L.; Alamri, A.; Ariaee, N.; Kung, S.; Gounni, A.S. Semaphorin 3E Deficiency Dysregulates Dendritic Cell Functions: In Vitro and in Vivo Evidence. PLoS ONE 2021, 16, e252868. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Wang, Z.; Zhang, H.; Chen, J.; Wang, X.; Cui, L.; Xie, C.; Li, M.; Wang, F.; Zhou, P.; et al. Mevalonate Blockade in Cancer Cells Triggers CLEC9A(+) Dendritic Cell-Mediated Antitumor Immunity. Cancer Res. 2021, 81, 4514–4528. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Liang, H.; Liu, X.; Wu, J.; Yang, C.; Wong, T.M.; Kwan, K.; Cheung, K.; Wu, S.; Yeung, K. Regulation of Macrophage Polarization through Surface Topography Design to Facilitate Implant-To-Bone Osteointegration. Sci. Adv. 2021, 7, eabf6654. [Google Scholar] [CrossRef]
- Scarpa, E.; Mayor, R. Collective Cell Migration in Development. J. Cell Biol. 2016, 212, 143–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahimi-Balaei, M.; Bergen, H.; Kong, J.; Marzban, H. Neuronal Migration During Development of the Cerebellum. Front. Cell. Neurosci. 2018, 12, 484. [Google Scholar] [CrossRef]
- Kanamori, M.; Oikawa, K.; Tanemura, K.; Hara, K. Mammalian Germ Cell Migration During Development, Growth, and Homeostasis. Reprod. Med. Biol. 2019, 18, 247–255. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Liu, D.; Wei, F.; Li, Y.; Wang, X.; Li, L.; Wang, G.; Zhang, S.; Zhang, L. Stress-Sensitive Protein Rac1 and its Involvement in Neurodevelopmental Disorders. Neural Plast. 2020, 2020, 8894372. [Google Scholar] [CrossRef]
- Gahankari, A.; Dong, C.; Bartoletti, G.; Galazo, M.; He, F. Deregulated Rac1 Activity in Neural Crest Controls Cell Proliferation, Migration and Differentiation During Midbrain Development. Front. Cell Dev. Biol. 2021, 9, 704769. [Google Scholar] [CrossRef]
- Jarysta, A.; Riou, L.; Firlej, V.; Lapoujade, C.; Kortulewski, T.; Barroca, V.; Gille, A.S.; Dumont, F.; Jacques, S.; Letourneur, F.; et al. Abnormal Migration Behavior Linked to Rac1 Signaling Contributes to Primordial Germ Cell Exhaustion in Fanconi Anemia Pathway-Deficient Fancg-/- Embryos. Hum. Mol. Genet. 2021, 31, 97–110. [Google Scholar] [CrossRef]
- Mizoguchi, T.; Ikeda, S.; Watanabe, S.; Sugawara, M.; Itoh, M. Mib1 Contributes to Persistent Directional Cell Migration by Regulating the Ctnnd1-Rac1 Pathway. Proc. Natl. Acad. Sci. USA 2017, 114, E9280–E9289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matera, D.L.; Lee, A.T.; Hiraki, H.L.; Baker, B.M. The Role of Rho GTPases During Fibroblast Spreading, Migration, and Myofibroblast Differentiation in 3D Synthetic Fibrous Matrices. Cell. Mol. Bioeng. 2021, 14, 381–396. [Google Scholar] [CrossRef] [PubMed]
- Bibi, S.; Ahmad, F.; Alam, M.R.; Ansar, M.; Yeou, K.S.; Wahedi, H.M. Lapachol-Induced Upregulation of Sirt1/Sirt3 is linked with Improved Skin Wound Healing in Alloxan-Induced Diabetic Mice. Iran. J. Pharm. Res. 2021, 20, 419–430. [Google Scholar] [CrossRef]
- Kann, A.P.; Hung, M.; Wang, W.; Nguyen, J.; Gilbert, P.M.; Wu, Z.; Krauss, R.S. An Injury-Responsive Rac-to-Rho GTPase Switch Drives Activation of Muscle Stem Cells through Rapid Cytoskeletal Remodeling. Cell Stem Cell 2022, 29, 933–947. [Google Scholar] [CrossRef]
- Mayor, R.; Etienne-Manneville, S. The Front and Rear of Collective Cell Migration. Nat. Rev. Mol. Cell Biol. 2016, 17, 97–109. [Google Scholar] [CrossRef] [Green Version]
- Jain, S.; Cachoux, V.; Narayana, G.; de Beco, S.; D’Alessandro, J.; Cellerin, V.; Chen, T.; Heuze, M.L.; Marcq, P.; Mege, R.M.; et al. The Role of Single Cell Mechanical Behavior and Polarity in Driving Collective Cell Migration. Nat. Phys. 2020, 16, 802–809. [Google Scholar] [CrossRef]
- Cheng, H.W.; Hsiao, C.T.; Chen, Y.Q.; Huang, C.M.; Chan, S.I.; Chiou, A.; Kuo, J.C. Centrosome Guides Spatial Activation of Rac to Control Cell Polarization and Directed Cell Migration. Life Sci. Alliance 2019, 2, e201800135. [Google Scholar] [CrossRef]
- Becsky, D.; Gyulai-Nagy, S.; Balind, A.; Horvath, P.; Dux, L.; Keller-Pinter, A. Myoblast Migration and Directional Persistence Affected by Syndecan-4-Mediated Tiam-1 Expression and Distribution. Int. J. Mol. Sci. 2020, 21, 823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller, P.M.; Rademacher, J.; Bagshaw, R.D.; Wortmann, C.; Barth, C.; van Unen, J.; Alp, K.M.; Giudice, G.; Eccles, R.L.; Heinrich, L.E.; et al. Systems Analysis of RhoGEF and RhoGAP Regulatory Proteins Reveals Spatially Organized RAC1 Signalling From Integrin Adhesions. Nat. Cell Biol. 2020, 22, 498–511. [Google Scholar] [CrossRef]
- Singh, S.P.; Paschke, P.; Tweedy, L.; Insall, R.H. AKT and SGK Kinases Regulate Cell Migration by Altering Scar/WAVE Complex Activation and Arp2/3 Complex Recruitment. Front. Mol. Biosci. 2022, 9, 965921. [Google Scholar] [CrossRef]
- Singh, S.P.; Thomason, P.A.; Insall, R.H. Extracellular Signalling Modulates Scar/WAVE Complex Activity through Abi Phosphorylation. Cells 2021, 10, 3485. [Google Scholar] [CrossRef] [PubMed]
- Qiu, F.; Long, H.; Zhang, L.; Liu, J.; Yang, Z.; Huang, X. Dermcidin Enhances the Migration, Invasion, and Metastasis of Hepatocellular Carcinoma Cells in Vitro and in Vivo. J. Clin. Transl. Hepatol. 2022, 10, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Law, A.L.; Jalal, S.; Pallett, T.; Mosis, F.; Guni, A.; Brayford, S.; Yolland, L.; Marcotti, S.; Levitt, J.A.; Poland, S.P.; et al. Nance-Horan Syndrome-Like 1 Protein Negatively Regulates Scar/WAVE-Arp2/3 Activity and Inhibits Lamellipodia Stability and Cell Migration. Nat. Commun. 2021, 12, 5687. [Google Scholar] [CrossRef]
- Yelland, T.; Le, A.H.; Nikolaou, S.; Insall, R.; Machesky, L.; Ismail, S. Structural Basis of CYRI-B Direct Competition with Scar/WAVE Complex for Rac1. Structure 2021, 29, 226–237. [Google Scholar] [CrossRef]
- Li, Y.; Liu, S.; Liu, H.; Cui, Y.; Deng, Y. Dragon’s Blood Regulates Rac1-WAVE2-Arp2/3 Signaling Pathway to Protect Rat Intestinal Epithelial Barrier Dysfunction Induced by Simulated Microgravity. Int. J. Mol. Sci. 2021, 22, 2722. [Google Scholar] [CrossRef]
- Chiang, H.Y.; Chu, P.H.; Chen, S.C.; Lee, T.H. MFG-E8 Regulates Vascular Smooth Muscle Cell Migration through Dose-Dependent Mediation of Actin Polymerization. J. Am. Heart Assoc. 2021, 10, e20870. [Google Scholar] [CrossRef]
- Yao, D.; Li, C.; Rajoka, M.; He, Z.; Huang, J.; Wang, J.; Zhang, J. P21-Activated Kinase 1: Emerging Biological Functions and Potential Therapeutic Targets in Cancer. Theranostics 2020, 10, 9741–9766. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Zhang, L.; Cui, W.; Du, J.; Li, Z.; Pang, Y.; Liu, Q.; Shang, H.; Meng, L.; Li, W.; et al. Epigenetically Upregulated GEFT-derived Invasion and Metastasis of Rhabdomyosarcoma Via Epithelial Mesenchymal Transition Promoted by the Rac1/Cdc42-PAK Signalling Pathway. EBioMedicine 2019, 50, 122–134. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Wang, Q.; Peng, S.; Yao, K.; Chen, J.; Tao, Y.; Gao, Z.; Wang, F.; Li, H.; Cai, W.; et al. The Metastatic Promoter DEPDC1B Induces Epithelial-Mesenchymal Transition and Promotes Prostate Cancer Cell Proliferation Via Rac1-PAK1 Signaling. Clin. Transl. Med. 2020, 10, e191. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Shi, W.; Hu, W.; Ma, D.; Yan, D.; Yu, K.; Zhang, G.; Cao, Y.; Wu, J.; Jiang, C.; et al. DEP Domain-Containing Protein 1B (DEPDC1B) Promotes Migration and Invasion in Pancreatic Cancer through the Rac1/PAK1-LIMK1-Cofilin1 Signaling Pathway. Onco Targets Ther. 2020, 13, 1481–1496. [Google Scholar] [CrossRef] [Green Version]
- Xu, T.; He, B.S.; Pan, B.; Pan, Y.Q.; Sun, H.L.; Liu, X.X.; Xu, X.N.; Chen, X.X.; Zeng, K.X.; Xu, M.; et al. MiR-142-3p Functions as a Tumor Suppressor by Targeting RAC1/PAK1 Pathway in Breast Cancer. J. Cell. Physiol. 2020, 235, 4928–4940. [Google Scholar] [CrossRef]
- Wufuer, R.; Ma, H.X.; Luo, M.Y.; Xu, K.Y.; Kang, L. Downregulation of Rac1/PAK1/LIMK1/cofilin Signaling Pathway in Colon Cancer SW620 Cells Treated with Chlorin E6 Photodynamic Therapy. Photodiagnosis Photodyn. Ther. 2021, 33, 102143. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Ren, M.; Li, Y.; Liu, Y.; Chen, C.; Su, J.; Su, B.; Xia, H.; Liu, F.; Jiang, H.; et al. Knockdown of RhoGDI2 Represses Human Gastric Cancer Cell Proliferation, Invasion and Drug Resistance Via the Rac1/Pak1/LIMK1 Pathway. Cancer Lett. 2020, 492, 136–146. [Google Scholar] [CrossRef]
- Tang, C.; Wu, X.; Ren, Q.; Yao, M.; Xu, S.; Yan, Z. Hedgehog Signaling is Controlled by Rac1 Activity. Theranostics 2022, 12, 1303–1320. [Google Scholar] [CrossRef]
- Li, X.; Zhu, J.; Liu, Y.; Duan, C.; Chang, R.; Zhang, C. MicroRNA-331-3p Inhibits Epithelial-Mesenchymal Transition by Targeting ErbB2 and VAV2 through the Rac1/PAK1/beta-catenin Axis in Non-Small-Cell Lung Cancer. Cancer Sci. 2019, 110, 1883–1896. [Google Scholar] [CrossRef] [Green Version]
- Lv, P.C.; Jiang, A.Q.; Zhang, W.M.; Zhu, H.L. FAK Inhibitors in Cancer, a Patent Review. Expert. Opin. Ther. Pat. 2018, 28, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Mitra, S.K.; Schlaepfer, D.D. Integrin-Regulated FAK-Src Signaling in Normal and Cancer Cells. Curr. Opin. Cell Biol. 2006, 18, 516–523. [Google Scholar] [CrossRef]
- Mo, C.F.; Li, J.; Yang, S.X.; Guo, H.J.; Liu, Y.; Luo, X.Y.; Wang, Y.T.; Li, M.H.; Li, J.Y.; Zou, Q. IQGAP1 Promotes Anoikis Resistance and Metastasis through Rac1-dependent ROS Accumulation and Activation of Src/FAK Signalling in Hepatocellular Carcinoma. Br. J. Cancer 2020, 123, 1154–1163. [Google Scholar] [CrossRef]
- Xie, Y.; Liu, C.; Qin, Y.; Chen, J.; Fang, J. Knockdown of IRE1a Suppresses Metastatic Potential of Colon Cancer Cells through Inhibiting FN1-Src/FAK-GTPases Signaling. Int. J. Biochem. Cell Biol. 2019, 114, 105572. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.Q.; Song, H.Y.; Zhou, Z.Y.; Ma, J.; Luo, Z.Y.; Zhou, Y.; Wang, J.Y.; Liu, S.; Han, X.H. Osthole Inhibits the Migration and Invasion of Highly Metastatic Breast Cancer Cells by Suppressing ITGalpha3/ITGbeta5 Signaling. Acta Pharmacol. Sin. 2022, 43, 1544–1555. [Google Scholar] [CrossRef] [PubMed]
- Ridley, A.J.; Schwartz, M.A.; Burridge, K.; Firtel, R.A.; Ginsberg, M.H.; Borisy, G.; Parsons, J.T.; Horwitz, A.R. Cell Migration: Integrating Signals From Front to Back. Science 2003, 302, 1704–1709. [Google Scholar] [CrossRef] [Green Version]
- Hung, I.C.; Chen, T.M.; Lin, J.P.; Tai, Y.L.; Shen, T.L.; Lee, S.J. Wnt5b Integrates Fak1a to Mediate Gastrulation Cell Movements Via Rac1 and Cdc42. Open Biol. 2020, 10, 190273. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.; Sun, Z.M.; Zhu, D.C.; Guo, Q.; Xu, J.J.; Lin, J.H.; Chen, Z.X.; Wu, Y.S. Inhibition of Rac1 Activity by NSC23766 Prevents Cartilage Endplate Degeneration via Wnt/Beta-Catenin Pathway. J. Cell. Mol. Med. 2020, 24, 3582–3592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Cao, T.; Ding, W.; Liang, L.; Fan, G.C.; Qu, L.; Peng, T. Pharmacological Inhibition of Rac1 Attenuates Myocardial Abnormalities in Tail-Suspended Mice. J. Cardiovasc. Transl. Res. 2022, 15, 805–815. [Google Scholar] [CrossRef] [PubMed]
- Ciarlantini, M.S.; Barquero, A.; Bayo, J.; Wetzler, D.; Dodes, T.M.; Bucci, H.A.; Fiore, E.J.; Gandolfi, D.L.; Defelipe, L.; Turjanski, A.; et al. Development of an Improved Guanidine-Based Rac1 Inhibitor with in Vivo Activity Against Non-Small Cell Lung Cancer. ChemMedChem 2021, 16, 1011–1021. [Google Scholar] [CrossRef]
- Bianchi-Smiraglia, A.; Wolff, D.W.; Marston, D.J.; Deng, Z.; Han, Z.; Moparthy, S.; Wombacher, R.M.; Mussell, A.L.; Shen, S.; Chen, J.; et al. Regulation of Local GTP Availability Controls RAC1 Activity and Cell Invasion. Nat. Commun. 2021, 12, 6091. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Cell Types | Cell Lines | Signaling Pathways | Ref. | |
|---|---|---|---|---|
| Tumor cells | HNSC cells | UM-SCC-1, UM-SCC-17B | Rap1/α5β1-integrin/Rac1 | [34] |
| HCC cells | MHCC-97H, MHCC-LM9, VETC-2 | AR/Rac1 | [35] | |
| SNU-387, SNU-423, SNU-449, SNU-475, Hep3B, HepG2 | Akt/Rac1/Fascin-1/Hippo/YAP | [45] | ||
| CM cells | \ | Rac1/Scar/Wave | [36] | |
| OS cells | MG-63 | Ras/Rac1/Wave2 | [37] | |
| RMS cells | SMS-CTR, RD, Rh5, Rh30 | HDAC6/Rac1 | [38] | |
| PAAD cells | MIA, PaCa-2, PANC-1 | Myosin II/Rac1 | [39] | |
| Cervical cancer cells | Hela, SiHa, Caski | DTL/Rac1/JNK/FOXO1 | [41] | |
| COAD/READ cells | SW480, HCT116 | TGF-β1/aPKC-ι/Rac1/JNK | [42] | |
| HCT116 | Pals1/Arf6/Rac1 | [49] | ||
| Gastric cancer cells | BGC-823, SGC-7901 | NEDD9/MICAL1/Rac1 | [43] | |
| Breast cancer cells | MDA-MB-231, MDA-MB-46 | SOX2/NEDD9/Rac1/HIF-1α | [44] | |
| GBM cells | M059K, M059J, U87 | DNA-PKcs/Akt2/Rac1 | [46] | |
| Bladder cancer cells | RT4, 5637, T24 | KDM6A/ARHGDIB/Rac1 | [48] | |
| Immune cells | Microglia | \ | TREM2/FAK/Rac1 | [50] |
| BV-2 | CXCL12/CXCR4/FAK/Src/Rac1 | [51] | ||
| NK cells | NK-92 | CRBN/Rac1 | [52] | |
| Neutrophils | \ | JFC1/Rac1 | [53] | |
| DC cells | pDC | α7nAChR/JAK2/STAT3/casepase-3/Rac1 | [54] | |
| Other cells | ECs | HEBC-5i | Rac1/PAK1 | [55] |
| \ | CD93/Cbl/Crk/Rac1 | [56] | ||
| CEC | IQGAP1/Rac1 | [57] | ||
| 7KC/Rac1 | [58] | |||
| HUVEC | Rac1/paxillin/FAK | [59] | ||
| Keratinocytes | HaCaT2 | TGF-β1/miR-200b/c-3p/Rac1 | [60] | |
| \ | EGF/EGFR/Rac1 | [61] | ||
| Epithelial cells | IPEC-J2 | Trp/CaSR/Rac1/PLC-γ1 | [62] | |
| MSCs | hMSC | TAFA2/Rac1/p38 | [63] | |
| NSCs | NSC | α-SMA/Rac1/F-actin | [64] | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, N.; Xu, E.; Luo, Q.; Song, G. Rac1: A Regulator of Cell Migration and a Potential Target for Cancer Therapy. Molecules 2023, 28, 2976. https://doi.org/10.3390/molecules28072976
Ma N, Xu E, Luo Q, Song G. Rac1: A Regulator of Cell Migration and a Potential Target for Cancer Therapy. Molecules. 2023; 28(7):2976. https://doi.org/10.3390/molecules28072976
Chicago/Turabian StyleMa, Ning, Erqian Xu, Qing Luo, and Guanbin Song. 2023. "Rac1: A Regulator of Cell Migration and a Potential Target for Cancer Therapy" Molecules 28, no. 7: 2976. https://doi.org/10.3390/molecules28072976