
The Extraction and High Antiproliferative Effect of Anthocyanin from Gardenblue Blueberry
Abstract
:1. Introduction
2. Results
2.1. Main Phytochemicals Analysis
2.2. Component Analysis
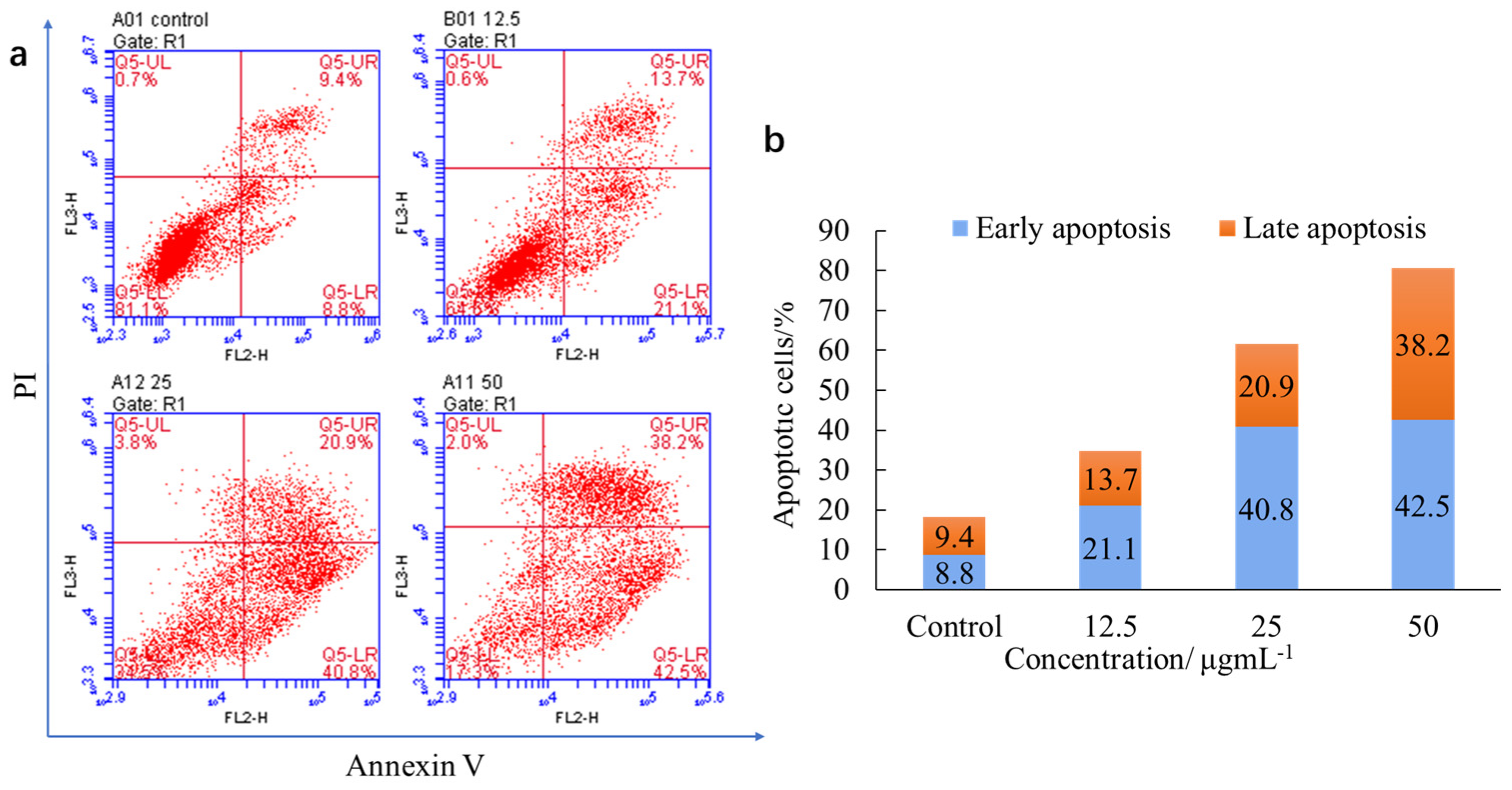
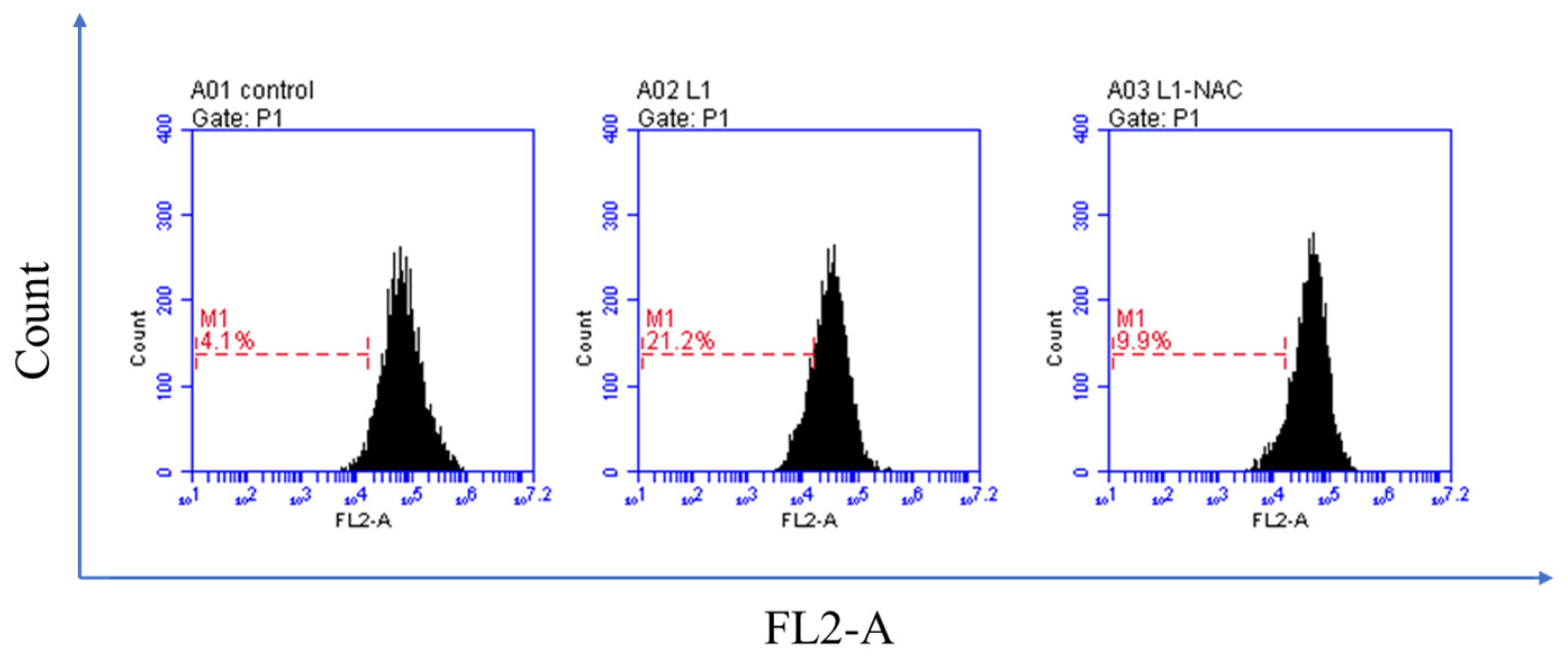
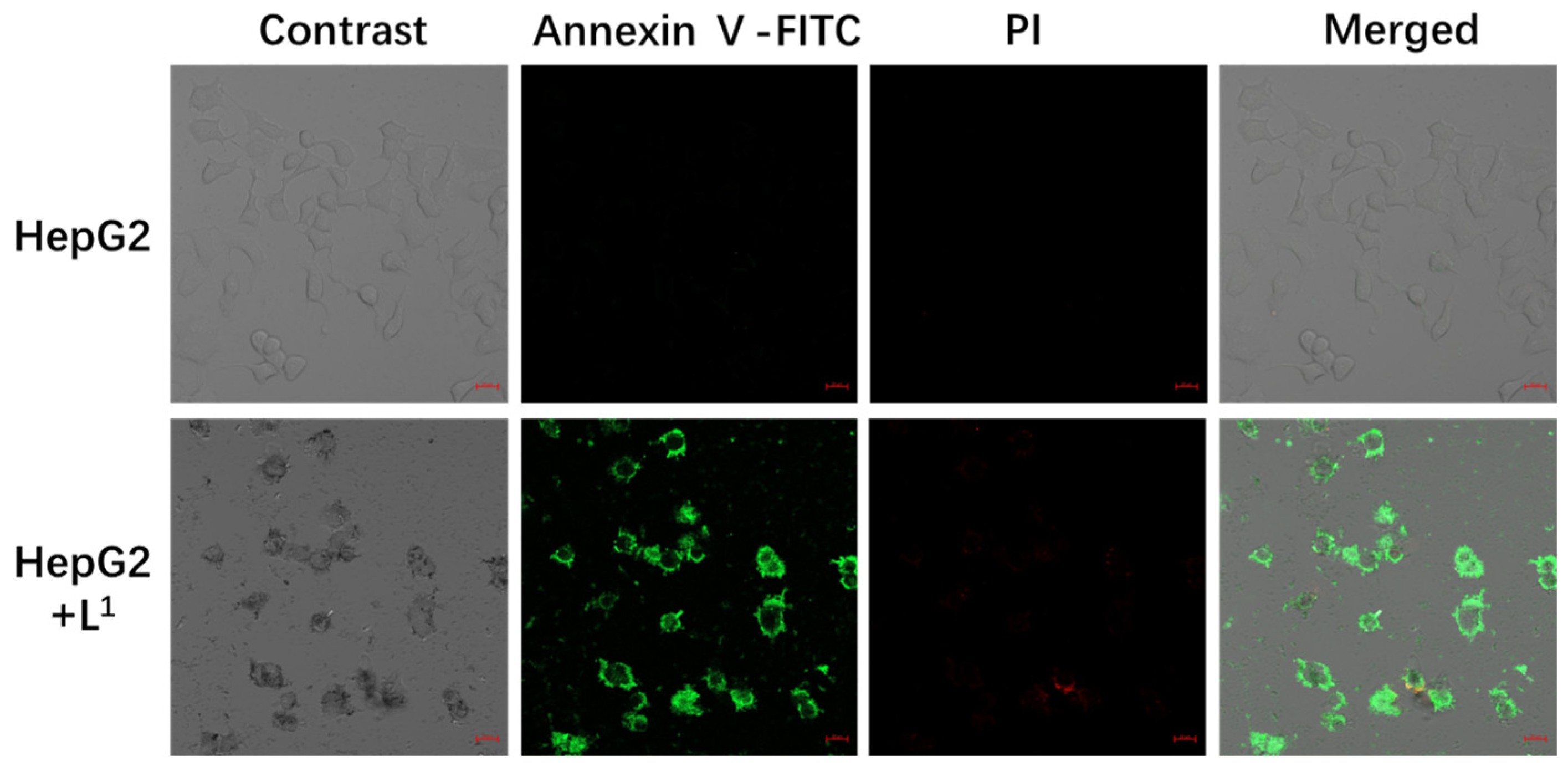
2.3. Biological Evaluation
2.3.1. Antiproliferative Activities
2.3.2. Induction of Apoptosis
2.3.3. Detection of Intracellular Reactive Oxygen Species (ROS)
2.3.4. Confocal Fluorescence Images
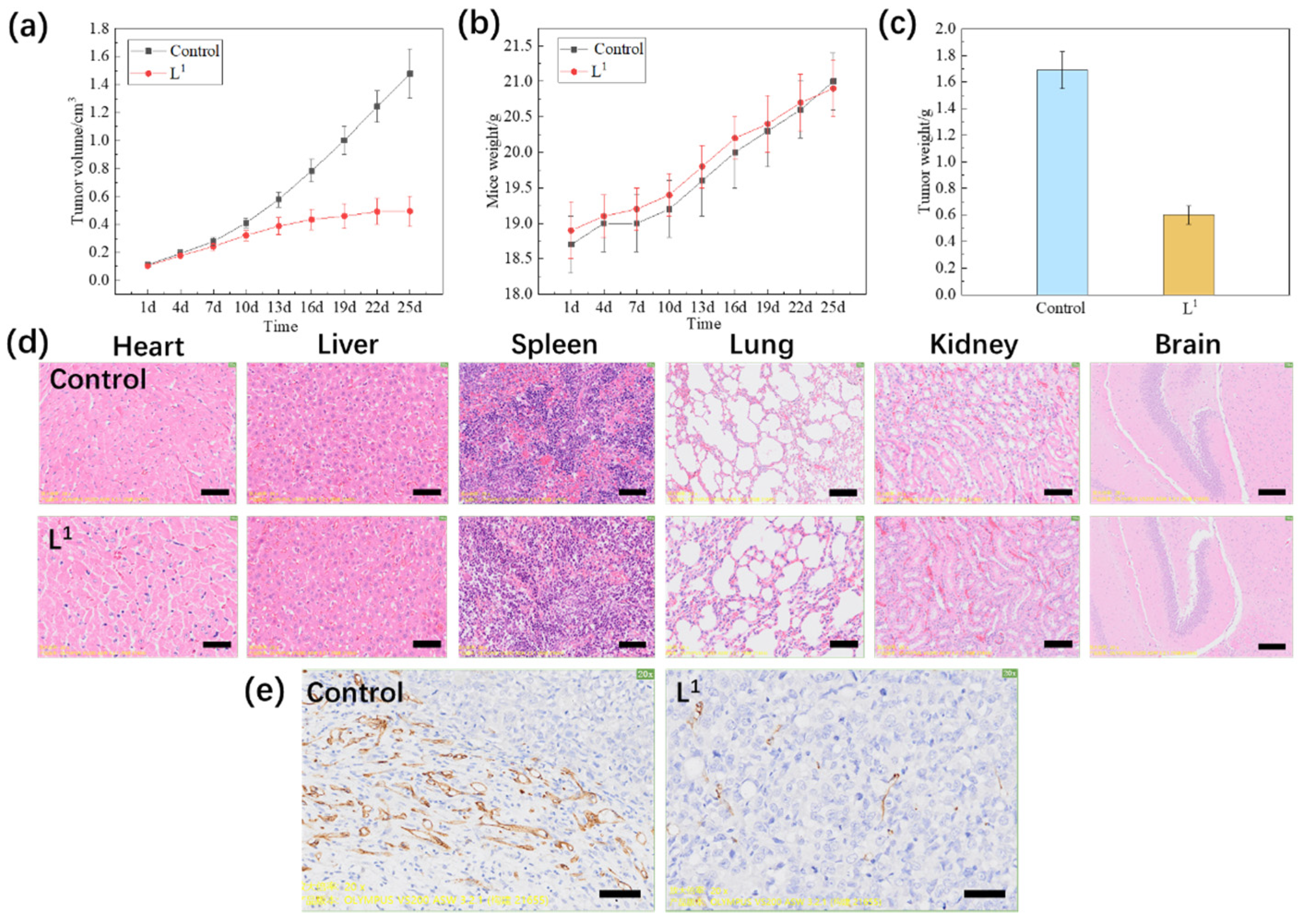
2.3.5. In Vivo Experiment
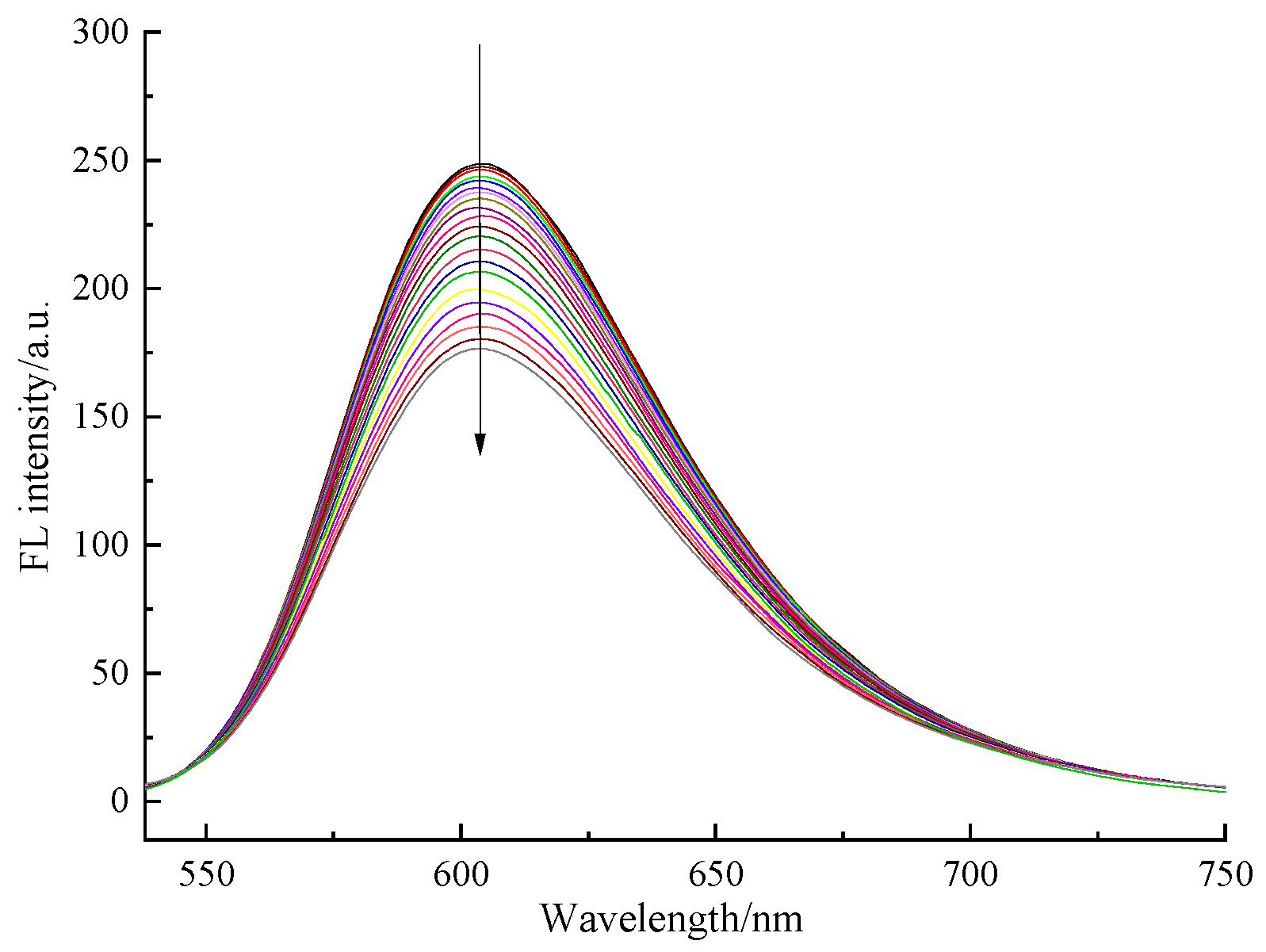
2.3.6. DNA Binding Modes
3. Discussion
4. Materials and Methods
4.1. General Materials
4.2. Soluble Solids Content
4.3. Titratable Acid Content Determination
4.4. Extraction and Purification
4.5. Component Analysis
4.6. Cell Culture, Antitumor Activities, and Cytotoxicity Assay
4.7. Induction of Apoptosis Assay
4.8. Detection of Intracellular Reactive Oxygen Species (ROS)
4.9. Confocal Fluorescence Images
4.10. In Vivo Experiment
4.11. DNA Binding Modes
4.12. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Su, X.; Zhang, J.; Wang, H.; Xu, J.; He, J.; Liu, L.; Zhang, T.; Chen, R.; Kang, J. Phenolic acid profiling, antioxidant, and anti-inflammatory activities, and miRNA regulation in the polyphenols of 16 blueberry samples from China. Molecules 2017, 22, 312. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Thakali, K.M.; Jensen, G.S.; Wu, X. Phenolic acids of the two major blueberry species in the US market and their antioxidant and anti-inflammatory activities. Plant. Foods. Hum. Nutr. 2015, 70, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.; Gill, C.; Stewart, D.; McDougall, G. Lingonberries (Vaccinium vitis-idaea L.) and blueberries (Vaccinium corymbo-sum) contain discrete epicatechin anthocyanin derivatives linked by ethyl bridges. J. Berry. Res. 2016, 6, 13–23. [Google Scholar] [CrossRef]
- Zhao, F.; Huang, Z.; Zhao, H.; Lyu, L.; Wu, W.; Li, W. Characteristics of four northern highbush blueberry varieties and per-formance in Nanjing. Mod. Agric. Sci. Technol. 2022, 1, 62–64. [Google Scholar]
- Yang, H.; Tian, T.; Wu, D.; Guo, D.; Lu, J. Prevention and treatment effects of edible berries for three deadly diseases: Car-diovascular disease, cancer and diabetes. Crit. Rev. Food. Sci. 2019, 59, 1903–1912. [Google Scholar] [CrossRef]
- Kalt, W.; Cassidy, A.; Howard, L.R.; Krikorian, R.; Stull, A.J.; Tremblay, F.; Zamora-Ros, R. Recent Research on the Health Benefits of Blueberries and Their Anthocyanins. Adv. Nutr. 2020, 11, 224–236. [Google Scholar] [CrossRef]
- Bornsek, S.M.; Ziberna, L.; Polak, T.; Vanzo, A.; Ulrih, N.P.; Abram, V.; Tramer, F.; Passamonti, S. Bilberry and blueberry an-thocyanins act as powerful intracellular antioxidants in mammalian cells. Food. Chem. 2012, 134, 1878–1884. [Google Scholar] [CrossRef]
- Joshi, S.S.; Howell, A.B.; D’Souza, D.H. Reduction of enteric viruses by blueberry juice and blueberry proanthocyanidins. Food Environ. Virol. 2016, 8, 235–243. [Google Scholar] [CrossRef]
- Huang, W.; Yan, Z.; Li, D.; Ma, Y.; Zhou, J.; Sui, Z. Antioxidant and anti-Inflammatory effects of blueberry anthocyanins on high glucose-induced human retinal capillary endothelial cells. Oxid. Med. Cell. Longev. 2018, 2018, 1862462. [Google Scholar] [CrossRef]
- Youdim, K.A.; Mcdonald, J.; Kalt, W.; Joseph, J.A. Potential role of dietary flavonoids in reducing microvascular endothelium vulnerability to oxidative and inflammatory insults (small star, filled). J. Nutr. Biochem. 2002, 13, 282–288. [Google Scholar] [CrossRef]
- Johnson, M.H.; Wallig, M.; Luna Vital, D.A.; de Mejia, E.G. Alcohol-free fermented blueberry-blackberry beverage phenolic extract attenuates diet-induced obesity and blood glucose in C57BL/6J mice. J. Nutr. Biochem. 2016, 31, 45–59. [Google Scholar] [CrossRef] [PubMed]
- Ştefănuţ, M.N.; Căta, A.; Pop, R.; Tănasie, C.; Boc, D.; Ienaşcu, I.; Ordodi, V. Anti-hyperglycemic effect of bilberry, blackberry and mulberry ultrasonic extracts on diabetic rats. Plant. Foods. Hum. Nutr. 2013, 68, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Lee, S.G.; Park, Y.-K.; Ku, C.S.; Pham, T.X.; Wegner, C.J.; Yang, Y.; Koo, S.I.; Chun, O.K.; Lee, J.-Y. Blueberry, blackberry, and blackcurrant differentially affect plasma lipids and pro-inflammatory markers in diet-induced obesity mice. Nutr. Res. Pract. 2016, 10, 494–500. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Daza, M.-C.; Daoust, L.; Boutkrabt, L.; Pilon, G.; Varin, T.; Dudonné, S.; Levy, É.; Marette, A.; Roy, D.; Desjardins, Y. Wild blueberry proanthocyanidins shape distinct gut microbiota profile and influence glucose homeostasis and intestinal phenotypes in high-fat high-sucrose fed mice. Sci. Rep. 2020, 10, 2217. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Gao, Y.; Guo, X.; Zhang, M.; Gong, L. Blackberry and blueberry anthocyanin supplementation counteract high-fat-diet-induced obesity by alleviating oxidative stress and inflammation and accelerating energy expenditure. Oxid. Med. Cell. Longev. 2018, 2018, 4051232. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Johnson, S.L.; Liu, W.; DaSilva, N.A.; Meschwitz, S.; Dain, J.A.; Seeram, N.P. Evaluation of polyphenol anthocyanin-enriched extracts of blackberry, black raspberry, blueberry, cranberry, red raspberry, and strawberry for free radical scavenging, reactive carbonyl species trapping, anti-glycation, anti-β-amyloid aggregation, and microglial neuroprotective effects. Int. J. Mol. Sci. 2018, 19, 461. [Google Scholar]
- Wood, E.; Hein, S.; Heiss, C.; Williams, C.; Rodriguez-Mateos, A. Blueberries and cardiovascular disease prevention. Food. Funct. 2019, 10, 7621–7633. [Google Scholar] [CrossRef]
- Silva, S.; Costa, E.; Calhau, C.; Morais, R.; Pintado, M. Anthocyanin extraction from plant tissues: A review. Crit. Rev. Food. Sci. 2017, 57, 3072–3083. [Google Scholar] [CrossRef]
- Zhao, X.-J.; Yang, W.-J.; Yang, R.-L.; Wu, T.-T.; Wang, Z.-Y.; Xu, N.-N.; He, J.-M. Research progress on biotransformation modi-fication of anthocyanins. Biotechnol. Bull. 2019, 35, 205–211. [Google Scholar]
- Song, Y.-Y.; Wang, Y.; Zhu, Q.-L.; Nie, Z.-K.; Yang, R.-L.; Zhao, X.-J. Research progress on sources and extraction methods of plant-based anthocyanin. Food. Res. Dev. 2022, 43, 199–208. [Google Scholar]
- Koide, T.; Kamei, H.; Hashimoto, Y.; Kojima, T.; Hasegawa, M. Antitumor effect of hydrolyzed anthocyanin from grape rinds and red rice. Cancer. Biother. Radio. 1996, 11, 273–277. [Google Scholar] [CrossRef] [PubMed]
- Koide, T.; Hashimoto, Y.; Kamei, H.; Kojima, T.; Hasegawa, M.; Terabe, K. Antitumor effect of anthocyanin fractions extracted from red soybeans and red beans in vitro and in vivo. Cancer. Biother. Radio. 1997, 12, 277–280. [Google Scholar] [CrossRef] [PubMed]
- Kamei, H.; Kojima, T.; Hasegawa, M.; Koide, T.; Umeda, T.; Yukawa, T.; Terabe, K. Suppression of tumor cell growth by an-thocyanins in vitro. Cancer. Investig. 1995, 13, 590–594. [Google Scholar] [CrossRef]
- Shi, N.; Chen, X.; Chen, T. Anthocyanins in colorectal cancer prevention review. Antioxidants 2021, 10, 1600. [Google Scholar] [CrossRef]
- Kubow, S.; Iskandar, M.M.; Melgar-Bermudez, E.; Sleno, L.; Sabally, K.; Azadi, B.; How, E.; Prakash, S.; Burgos, G.; Felde, T.Z. Effects of simulated human gastrointestinal digestion of two purple-fleshed potato cultivars on anthocyanin composition and cytotoxicity in colonic cancer and non-tumorigenic cells. Nutrients 2017, 9, 953. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.; Liu, Y.; Xu, C.; Liu, J. Antiproliferative and proapoptotic activities of anthocyanin and anthocyanidin extracts from blueberry fruits on B16–F10 melanoma cells. Food. Nutr. Res. 2017, 61, 1325308–1325313. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, D.; Liu, Y.; Wang, D.; Liu, J.; Ji, B. The protective effects of berry-derived anthocyanins against visible light-induced damage in human retinal pigment epithelial cells. J. Sci. Food. Agric. 2015, 95, 936–944. [Google Scholar] [CrossRef]
- Yang, W.; Guo, Y.; Liu, M.; Chen, X.; Xiao, X.; Wang, S.; Gong, P.; Ma, Y.; Chen, F. Structure and function of blueberry anthocyanins: A review of recent advances. J. Funct. Foods. 2022, 88, 104864. [Google Scholar] [CrossRef]
- Surangi Dharmawansa, K.V.; Hoskin, D.W.; Vasantha Ru-pasinghe, H.P. Chemopreventive effect of dietary anthocyanins against gastrointestinal cancers: A review of recent advances and perspectives. Int. J. Mol. Sci. 2020, 21, 6555. [Google Scholar] [CrossRef]
- Zhao, F.; Wang, W.; Lu, W.; Xu, L.; Yang, S.; Cai, X.-M.; Zhou, M.; Lei, M.; Ma, M.; Xu, H.-J.; et al. High anticancer potency on tumor cells of dehydroabietylamine Schiff-base derivatives and a copper(II) complex. Eur. J. Med. Chem. 2018, 146, 451–459. [Google Scholar] [CrossRef]
- Ghosh, S. Cisplatin: The first metal based anticancer drug. Bioorg. Chem. 2019, 88, 102925. [Google Scholar] [CrossRef] [PubMed]
- Lian, Y.-R.; Gan, H.; Meng, Z.-Y.; Gu, R.-L.; Zhu, X.-X.; Wu, Z.-N.; Yang, W.-J.; Kang, M.; Dou, G.-F. Analysis of anthocyanin components and antioxidant properties of a new species of blueberry. Food R&D 2022, 43, 30–36. [Google Scholar]
- Chen, H.; Niu, X. Effects of microwave extraction on anthocyanins components and antioxidant activity of blueberry. Food Sci. Tech. 2021, 46, 167–175. [Google Scholar]
- Wang, Y.; Lin, J.; Tian, J.; Si, X.; Jiao, X.; Zhang, W.; Gong, E.; Li, B. Blueberry malvidin-3-galactoside suppresses hepatocellular carcinoma by regulating apoptosis, proliferation, and metastasis pathways in vivo and in vitro. J. Agric. Food Chem. 2019, 67, 625–636. [Google Scholar] [CrossRef]
- Gu, Z.; Lu, W.; Xue, H.; Zhang, J.; Yang, S.; Xu, L. Syntheses and high selective cytotoxicity of dehydroabietylamine C-ring nitration derivatives. Fitoterapia 2022, 161, 105232. [Google Scholar] [CrossRef]
- Zhao, F.; Zhao, H.; Wu, W.; Wang, W.; Li, W. Research on anthocyanins from Rubus “Shuofeng” as potential antiproliferative and apoptosis-inducing agents. Foods 2023, 12, 1216. [Google Scholar] [CrossRef]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of apoptosis signalling pathways by reactive oxygen species. Biochim. Biophys. Acta 2016, 1863, 2977–2992. [Google Scholar] [CrossRef]
- Halliwell, B.; Whiteman, M. Measuring reactive species and oxidative damage in vivo and in cell culture: How should you do it and what do the results mean? Br. J. Pharmacol. 2004, 142, 231–255. [Google Scholar] [CrossRef] [PubMed]
- Rowe, L.A.; Degtyareva, N.; Doetsch, P.W. DNA damage-induced reactive oxygen species (ROS) stress response in Saccharomyces cerevisiae. Free. Radic. Biol. Med. 2008, 45, 1167–1177. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.-H.; Ye, C.; Bai, E.-H.; Zhang, L.-L.; Huo, S.-J.; Yu, H.-H.; Xiang, S.-Y.; Yu, S.-Q. Co-delivery nanoparticles of doxorubicin and chloroquine for improving the anti-cancer effect in vitro. Nanotechnology 2019, 30, 085101–085126. [Google Scholar] [CrossRef]
- Li, Q.; Chu, D.T.W.; Claiborne, A.; Cooper, C.S.; Lee, C.M.; Raye, K.; Berst, K.B.; Donner, P.; Wang, W.; Hasvold, L.; et al. Synthesis and structure-activity rela-tionships of 2-pyridones: A novel series of potent DNA gyrase inhibitors as antibacterial agents. J. Med. Chem. 1996, 39, 3070–3088. [Google Scholar] [CrossRef] [PubMed]
- Gao, E.-J.; Wang, K.-H.; Zhu, M.-C.; Liu, L. Hairpin-shaped tetranuclear palladium (II) complex: Synthesis, crystal structure, DNA binding and cytotoxicity activity studies. Eur. J. Med. Chem. 2010, 45, 2784–2790. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Sun, X.; Lu, W.; Xu, L.; Shi, J.; Yang, S.; Zhou, M.; Su, F.; Lin, F.; Cao, F. Synthesis of novel, DNA binding heterocyclic dehydroabietylamine derivatives as potential antiproliferative and apoptosis-inducing agents. Drug. Deliv. 2020, 27, 216–227. [Google Scholar] [CrossRef]
- Banc, R.; Rusu, M.E.; Filip, L.; Popa, D.-S. The impact of ellagitannins and their metabolites through gut microbiome on the gut health and brain wellness within the gut-brain axis. Foods 2023, 12, 270. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
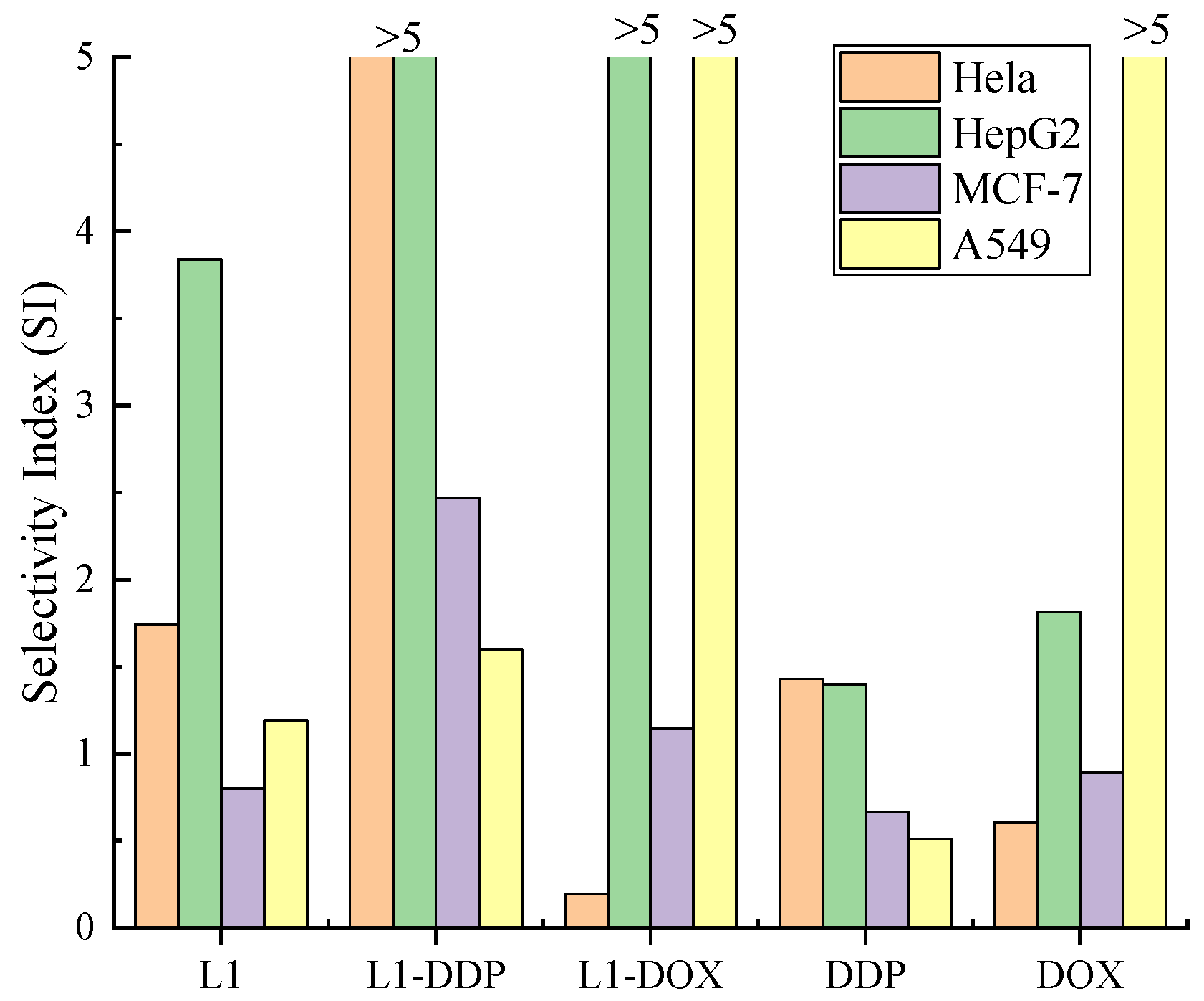{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Soluble Solid (%) | Total Acid (Citrate, %) | Solidity-Acid Ratio | Total Anthocyanins (mg/100 g) |
|---|---|---|---|---|
| Sharpblue | 10.40 | 0.79 | 18.87 | 104 ± 1 |
| O’Neal | 10.70 | 0.49 | 25.70 | 100 ± 3 |
| Sunshine | 12.63 | 0.48 | 22.40 | 115 ± 3 |
| Magnolia | 11.30 | 0.67 | 21.13 | 103 ± 3 |
| Blue rain | 12.40 | 0.53 | 19.84 | 117.0 ± 0.6 |
| Anna | 13.50 | 0.49 | 32.38 | 72 ± 4 |
| Biloxi | 11.93 | 0.86 | 20.20 | 138 ± 2 |
| Misty | 10.33 | 0.36 | 21.83 | 101 ± 3 |
| Star | 11.00 | 0.71 | 16.66 | 77 ± 1 |
| Summit | 9.40 | 0.87 | 13.64 | 110 ± 4 |
| Cooper | 11.00 | 0.62 | 17.45 | 96.3 ± 0.9 |
| Ozarkblue | 9.53 | 0.94 | 10.14 | 111.5 ± 0.9 |
| Springhigh | 11.63 | 0.77 | 15.10 | 151 ± 2 |
| Primadonna | 10.80 | 0.46 | 25.00 | 129 ± 2 |
| Millennia | 11.03 | 0.70 | 18.75 | 118 ± 1 |
| Emerald | 10.20 | 0.61 | 20.19 | 155 ± 3 |
| Jewel | 10.00 | 0.41 | 27.76 | 128 ± 1 |
| Windsor | 12.40 | 0.51 | 24.31 | 181 ± 2 |
| Millennia | 10.70 | 0.70 | 15.29 | 110 ± 2 |
| Eureka | 9.90 | 0.59 | 16.78 | 165 ± 2 |
| Farthing | 12.43 | 0.69 | 18.01 | 117 ± 2 |
| Zhaixuan ‘4’ | 12.70 | 0.56 | 27.11 | 230 ± 2 |
| Zhaixuan ‘7’ | 12.08 | 0.69 | 24.29 | 168 ± 1 |
| Zhongzhi ‘1’ | 9.80 | 0.42 | 24.24 | 130 ± 3 |
| Zhongzhi ‘2’ | 10.73 | 0.77 | 16.10 | 111.8 ± 0.6 |
| Zhongzhi ‘3’ | 11.40 | 0.51 | 24.04 | 117 ± 1 |
| Zhongzhi ‘4’ | 9.40 | 0.59 | 22.12 | 178 ± 2 |
| Chandler | 13.03 | 0.55 | 22.75 | 112 ± 3 |
| Legacy | 11.07 | 0.47 | 23.74 | 148.8 ± 0.2 |
| Elliott | 9.93 | 0.55 | 18.05 | 168 ± 1 |
| Duke | 9.30 | 0.60 | 19.11 | 127.7 ± 0.7 |
| Bluecrop | 10.03 | 0.48 | 20.90 | 111 ± 4 |
| Reka | 9.00 | 0.54 | 16.67 | 132.5 ± 0.9 |
| Darrow | 12.43 | 0.70 | 16.25 | 149 ± 2 |
| Jersey | 11.70 | 0.64 | 22.27 | 134 ± 1 |
| Berkeley | 11.47 | 0.80 | 16.80 | 145.5 ± 0.8 |
| Brigitta | 11.10 | 0.68 | 16.71 | 113 ± 5 |
| St·Blue | 11.05 | 0.52 | 21.25 | 151 ± 2. |
| Sweetheart | 8.83 | 1.00 | 13.59 | 106 ± 2 |
| Ruble | 11.27 | 0.43 | 26.21 | 192.70 ± 0.08 |
| Aurora | 8.10 | 0.70 | 16.73 | 78 ± 2 |
| Camellia | 11.77 | 0.58 | 20.29 | 83 ± 1 |
| Biggold | 11.93 | 0.54 | 22.09 | 127 ± 5 |
| Spartan | 10.40 | 0.56 | 18.57 | 113 ± 2 |
| Denise blue | 8.87 | 0.70 | 12.67 | 70 ± 3 |
| Powderblue | 13.67 | 0.72 | 20.21 | 119 ± 3 |
| Gardenblue | 12.33 | 0.63 | 23.68 | 259 ± 2 |
| Centurion | 11.20 | 0.70 | 16.35 | 221 ± 2 |
| Tifblue | 13.93 | 0.60 | 24.07 | 153.8 ± 0.5 |
| Brightwell | 13.60 | 0.56 | 25.10 | 113 ± 3 |
| Climax | 13.03 | 0.76 | 19.90 | 143 ± 2 |
| Premier | 11.70 | 0.53 | 25.17 | 162 ± 4 |
| Woodard | 10.17 | 0.75 | 16.68 | 152 ± 1 |
| Choice | 11.03 | 0.65 | 18.54 | 101 ± 2 |
| Baldwin | 11.00 | 0.78 | 16.25 | 116 ± 1 |
| Prolific | 9.80 | 0.45 | 22.17 | 121 ± 2 |
| Briteblue | 11.97 | 0.58 | 21.44 | 163.1 ± 0.5 |
| Beckyblue | 13.73 | 0.57 | 27.74 | 126 ± 1 |
| Vernon | 15.63 | 0.56 | 24.63 | 228.9 ± 0.5 |
| Onslow | 13.17 | 0.55 | 24.20 | 182 ± 1 |
| Homebell | 14.77 | 0.57 | 24.97 | 185 ± 3 |
| Bonita | 11.00 | 0.44 | 24.02 | 186 ± 2 |
| Titan | 9.07 | 0.73 | 12.42 | 86 ± 4 |
| Coastal | 11.57 | 0.74 | 15.15 | 137 ± 2 |
| Caroline blue | 11.50 | 1.44 | 7.99 | 157 ± 2 |
| Pink Lemonade | 13.57 | 0.66 | 23.78 | 3.4 ± 0.7 |
| Average | 11.33 | 0.61 | 20.11 | 133.50 |
| Gardenblue | Phenols | Anthocyanin | Ellagic Acid | Flavonoid |
|---|---|---|---|---|
| Fresh fruit | 4.62 ± 0.25 | 2.59 ± 0.12 | 1.28 ± 0.05 | 0.32 ± 0.02 |
| Dried powder | 809.65 ± 4.98 | 459.18 ± 4.04 | 216.79 ± 2.30 | 67.14 ± 1.85 |
| No. | RT/min | MS/[M]+ | MS2 | Δm | Tentative Identification |
|---|---|---|---|---|---|
| 1 | 13.95 | 465.1 | 303.0 | 162 | Del-hexose |
| 2 | 15.01 | 465.1 | 303.0 | 162 | Del-hexose |
| 3 | 15.35 | 465.1 | 303.0 | 162 | Del-gal |
| 4 | 16.36 | 465.1 | 303.0 | 162 | Del-glc |
| 5 | 18.33 | 449.1 | 287.1 | 162 | Cya-gal |
| 6 | 18.40 | 435.1 | 303.0 | 132 | Del-ara/xyl |
| 7 | 19.71 | 449.1 | 287.1 | 162 | Cya-glc |
| 8 | 19.91 | 479.1 | 317.1 | 162 | Pet-gal |
| 9 | 21.17 | 479.1 | 317.1 | 162 | Pet-glc |
| 10 | 21.85 | 419.1 | 287.1 | 132 | Cya-ara/xyl |
| 11 | 22.57 | 435.1 | 303.0 | 132 | Del-ara/xyl |
| 12 | 23.36 | 463.1 | 301.1 | 162 | Peo-gal |
| 13 | 23.63 | 449.1 | 317.1 | 132 | Pet-ara/xyl |
| 14 | 24.85 | 493.1 | 331.1 | 162 | Mal-gal |
| 15 | 25.60 | 463.1 | 301.1 | 162 | Peo-glc |
| 16 | 27.08 | 493.1 | 331.1 | 162 | Mal-glc |
| 17 | 28.57 | 433.1 | 301.1 | 132 | Peo-ara/xyl |
| 18 | 29.56 | 463.1 | 331.1 | 132 | Mal-ara/xyl |
| 19 | 31.35 | 433.1 | 301.1 | 132 | Peo-ara/xyl |
| 20 | 31.60 | 463.1 | 331.1 | 132 | Mal-ara/xyl |
| 21 | 31.93 | 287.1 | 137.0 | — | Cya |
| 22 | 32.34 | 317.1 | 245.0 | — | Pet |
| 23 | 33.02 | 611.2 | 303.1 | 308 | Del-glc-rha |
| 24 | 33.63 | 271.1 | — | — | Pel |
| 25 | 34.24 | 465.1 | 303.0 | 162 | Del-hexose |
| Compd. | Hela | HepG2 | MCF-7 | A549 | HUVEC |
|---|---|---|---|---|---|
| L1 | 52.0 ± 0.9 | 23.6 ± 0.9 | 113 ± 3 | 76 ± 3 | 91 ± 3 |
| L1-DDP | 2.6 ± 0.3 **b | 10.47 ± 0.54 *c | 30 ± 2 ***a | 47 ± 3 **b | 75 ± 2 *c |
| L1-DOX | 40.89 ± 0.9 *c | 0.02 ± 0.01 **b | 7.0 ± 0.9*c | 0.55 ± 0.05 *c | 8.0 ± 0.2 **b |
| DDP | 18 ± 1 *c | 18.9 ± 0.7 *c | 40 ± 1 ***a | 52 ± 4 ***a | 26 ± 1 **b |
| DOX | 1.0 ± 0.1 ***a | 0.3 ± 0.1 ***a | 0.6 ± 0.1 *c | 0.10 ± 0.01 **b | 0.58 ± 0.07 **b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, F.; Wang, J.; Wang, W.; Lyu, L.; Wu, W.; Li, W. The Extraction and High Antiproliferative Effect of Anthocyanin from Gardenblue Blueberry. Molecules 2023, 28, 2850. https://doi.org/10.3390/molecules28062850
Zhao F, Wang J, Wang W, Lyu L, Wu W, Li W. The Extraction and High Antiproliferative Effect of Anthocyanin from Gardenblue Blueberry. Molecules. 2023; 28(6):2850. https://doi.org/10.3390/molecules28062850
Chicago/Turabian StyleZhao, Fengyi, Jialuan Wang, Weifan Wang, Lianfei Lyu, Wenlong Wu, and Weilin Li. 2023. "The Extraction and High Antiproliferative Effect of Anthocyanin from Gardenblue Blueberry" Molecules 28, no. 6: 2850. https://doi.org/10.3390/molecules28062850