
Amino Acid Substitutions at P1 Position Change the Inhibitory Activity and Specificity of Protease Inhibitors BmSPI38 and BmSPI39 from Bombyx mori

 ,
,
Abstract
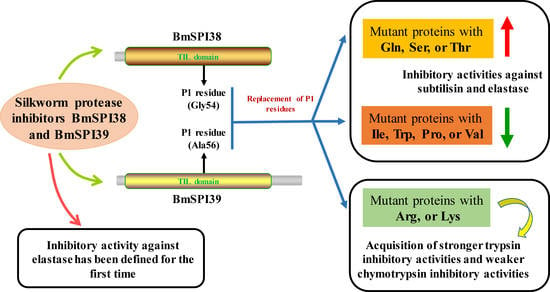
:
1. Introduction
2. Results
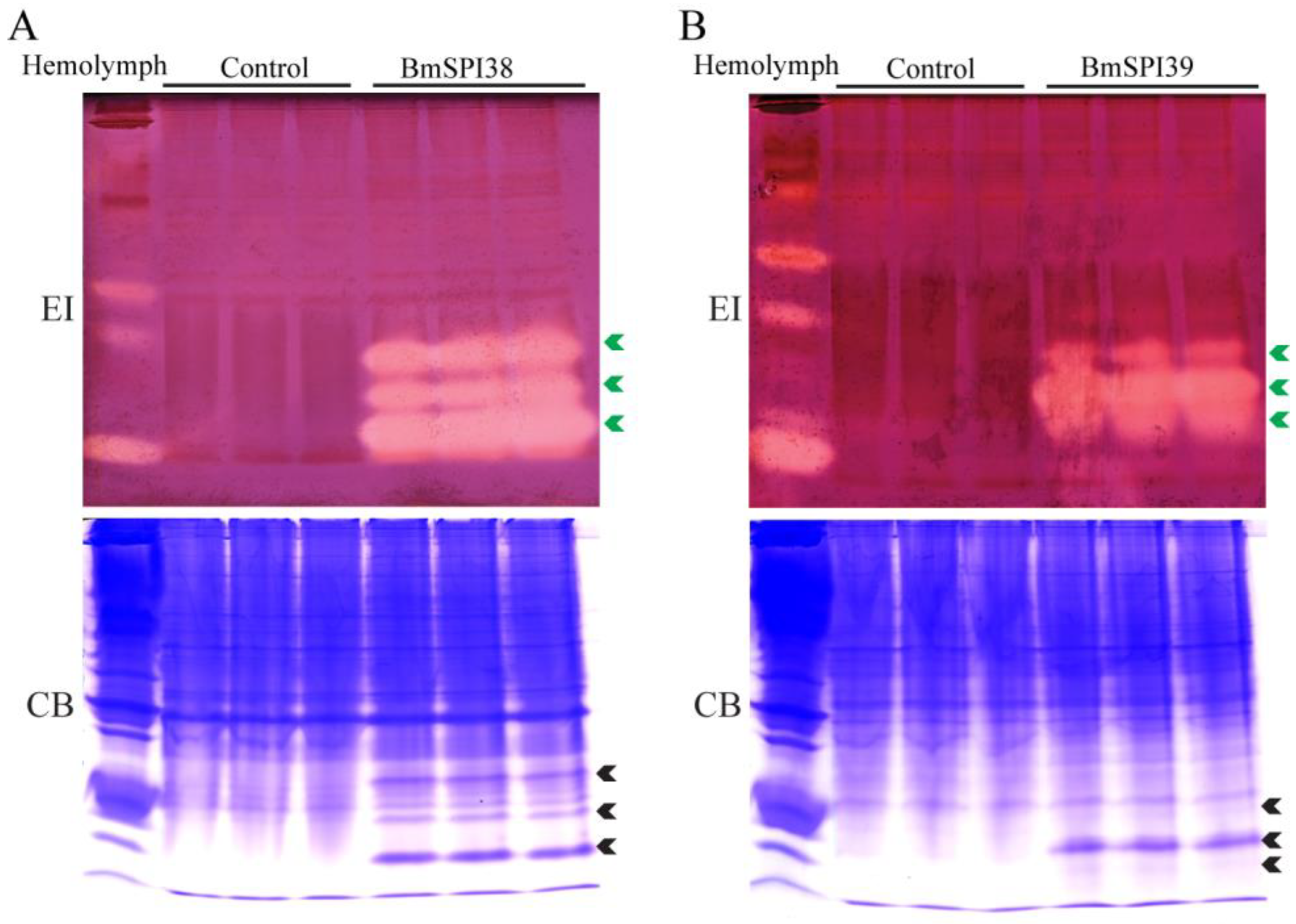
2.1. BmSPI38 and BmSPI39 Have Inhibitory Activity against Elastase
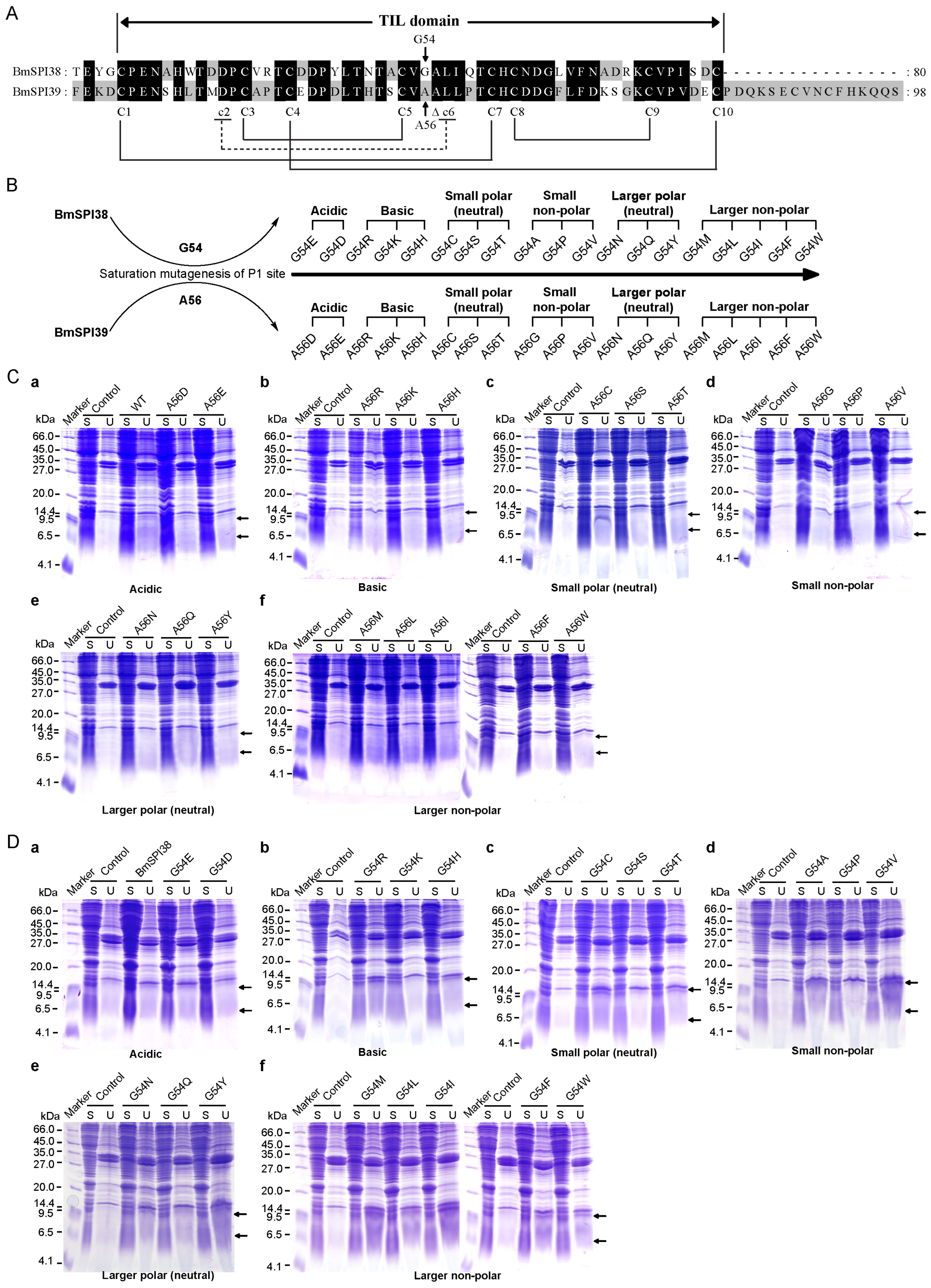
2.2. Design, Expression Vector Construction, and Prokaryotic Expression of the P1 Mutants of BmSPI38 and BmSPI39
2.3. Inhibition Activity of the P1 Mutants of BmSPI38 against the Serine Protease
2.4. Inhibition Activity of the P1 Mutants of BmSPI39 against the Serine Protease
2.5. The Replacement of P1 Residue with Lysine or Arginine Enables BmSPI38 and BmSPI39 to Obtain Trypsin Inhibitory Activity
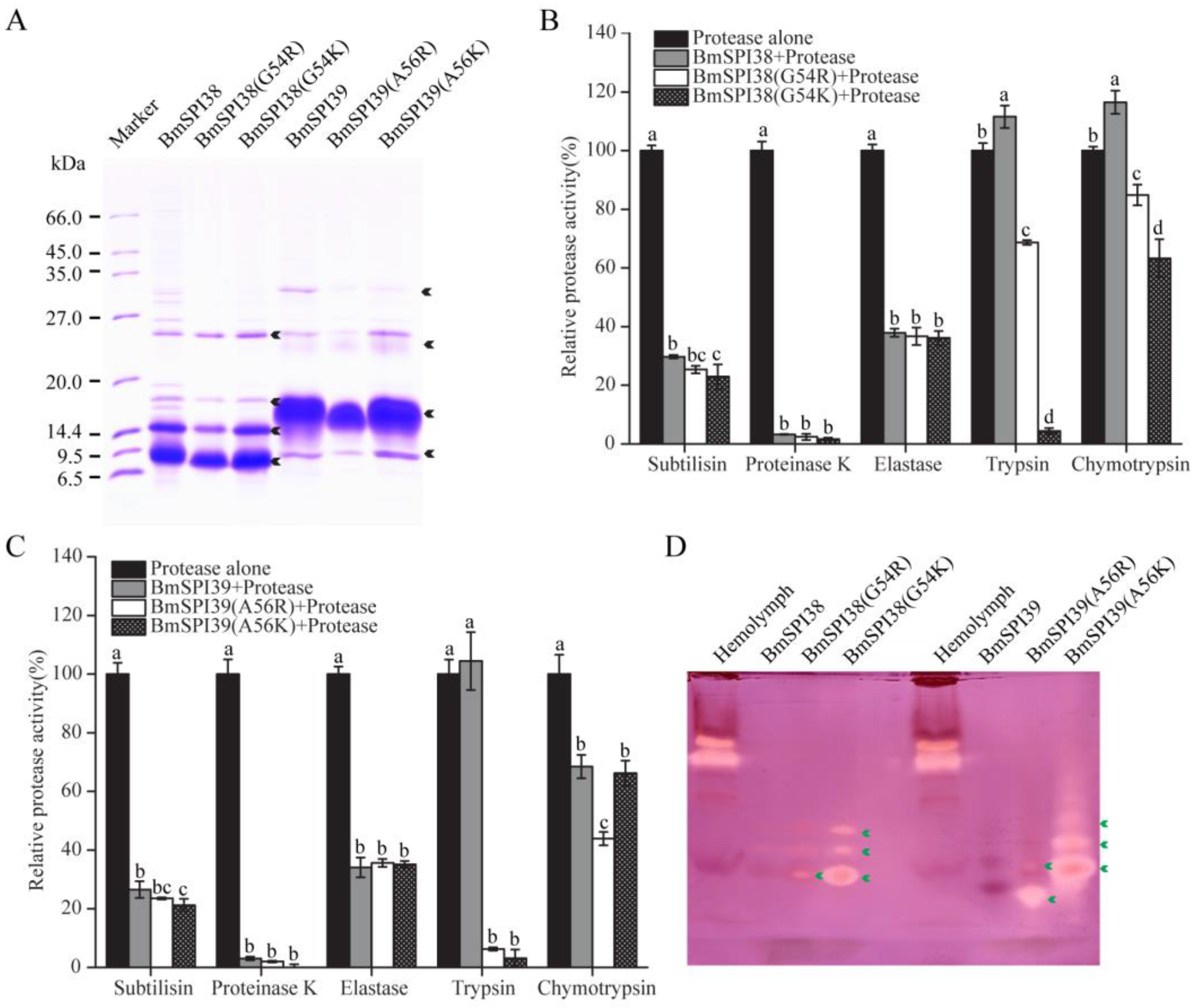
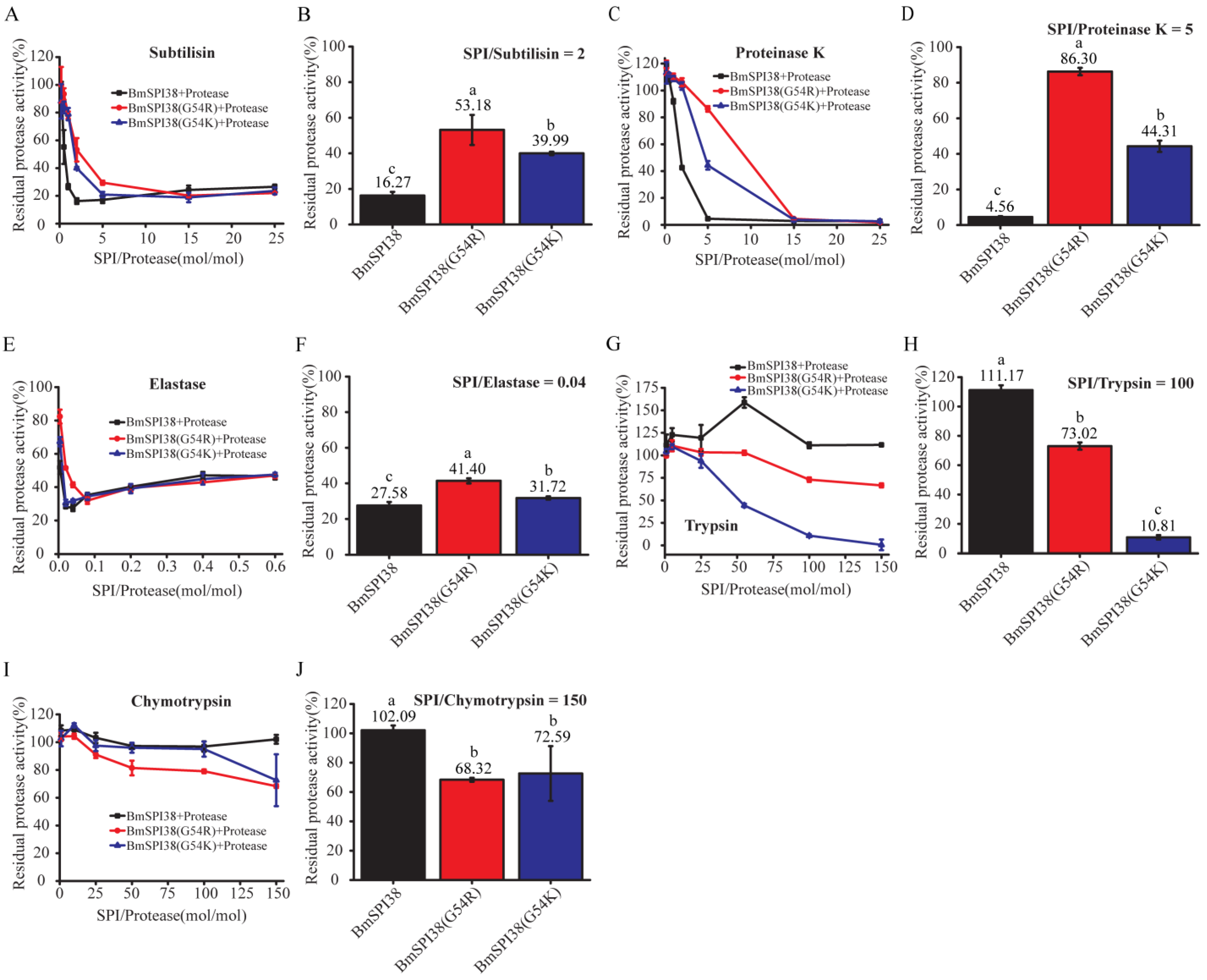
2.6. Comparison of the Inhibitory Ability of Mutant BmSPI38 Proteins to Different Serine Proteases
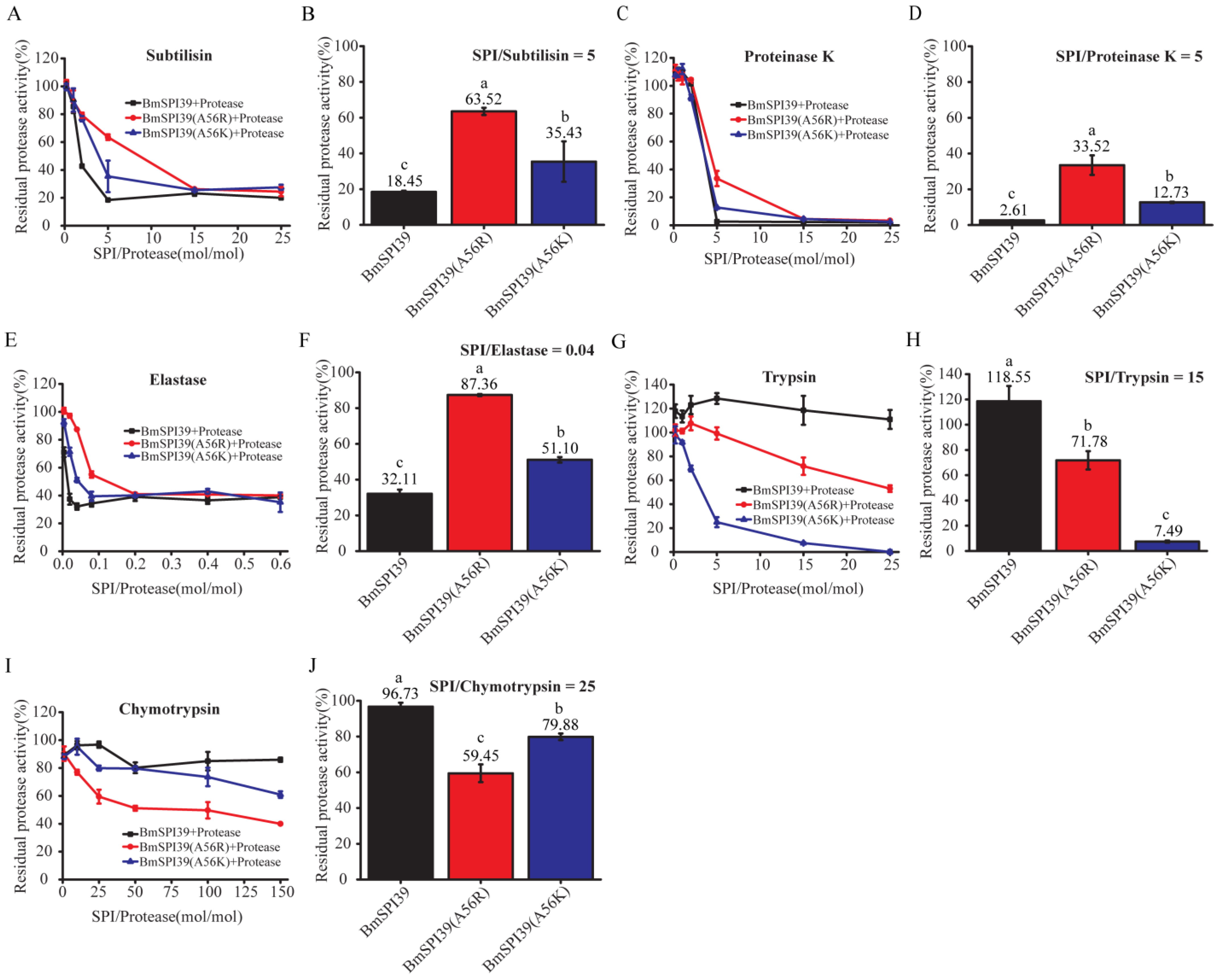
2.7. Comparison of the Inhibitory Ability of Mutant BmSPI39 Proteins to Different Serine Proteases
2.8. P1 Mutants of BmSPI38 and BmSPI39 Have Extremely High Acid–Base and Thermal Stability
3. Discussion
4. Materials and Methods
4.1. Escherichia coli Strains, Plasmids, and Reagents
4.2. Expression Vector Construct of the P1 Mutants
4.3. Expression and Purification of the Mutants
4.4. In-Gel Activity Staining of Protease Inhibitor
4.5. Protease Inhibition Assays
4.6. Acid–Base and Thermal Stability Analysis
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Hibbetts, K.; Hines, B.; Williams, D. An overview of proteinase inhibitors. J. Vet. Intern. Med. 1999, 13, 302–308. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, Y.; Li, Y. Key active sites of proteases and protease inhibitors: A review. Chin. J. Biotechnol. 2021, 37, 561–579. [Google Scholar]
- Xia, Q.; Zhou, Z.; Lu, C.; Cheng, D.; Dai, F.; Li, B.; Zhao, P.; Zha, X.; Cheng, T.; Chai, C.; et al. A draft sequence for the genome of the domesticated silkworm (Bombyx mori). Science 2004, 306, 1937–1940. [Google Scholar]
- Xia, Q.; Cheng, D.; Duan, J.; Wang, G.; Cheng, T.; Zha, X.; Liu, C.; Zhao, P.; Dai, F.; Zhang, Z.; et al. Microarray-based gene expression profiles in multiple tissues of the domesticated silkworm, Bombyx mori. Genome Biol. 2007, 8, R162. [Google Scholar] [CrossRef] [Green Version]
- Duan, J.; Li, R.; Cheng, D.; Fan, W.; Zha, X.; Cheng, T.; Wu, Y.; Wang, J.; Mita, K.; Xiang, Z.; et al. SilkDB v2.0: A platform for silkworm (Bombyx mori) genome biology. Nucleic Acids Res. 2010, 38, D453–D456. [Google Scholar] [CrossRef] [Green Version]
- Xia, Q.; Li, S.; Feng, Q. Advances in silkworm studies accelerated by the genome sequencing of Bombyx mori. Annu. Rev. Entomol. 2014, 59, 513–536. [Google Scholar] [CrossRef]
- Kanost, M.R. Serine proteinase inhibitors in arthropod immunity. Dev. Comp. Immunol. 1999, 23, 291–301. [Google Scholar] [CrossRef]
- Fullaondo, A.; Garcia-Sanchez, S.; Sanz-Parra, A.; Recio, E.; Lee, S.Y.; Gubb, D. Spn1 regulates the GNBP3-dependent Toll signaling pathway in Drosophila melanogaster. Mol. Cell. Biol. 2011, 31, 2960–2972. [Google Scholar] [CrossRef] [Green Version]
- Cerenius, L.; Kawabata, S.; Lee, B.L.; Nonaka, M.; Soderhall, K. Proteolytic cascades and their involvement in invertebrate immunity. Trends Biochem. Sci. 2010, 35, 575–583. [Google Scholar] [CrossRef]
- Zhao, P.; Dong, Z.; Duan, J.; Wang, G.; Wang, L.; Li, Y.; Xiang, Z.; Xia, Q. Genome-wide identification and immune response analysis of serine protease inhibitor genes in the silkworm, Bombyx mori. PLoS ONE 2012, 7, e31168. [Google Scholar] [CrossRef]
- Li, Y.; Zhao, P.; Liu, S.; Dong, Z.; Chen, J.; Xiang, Z.; Xia, Q. A novel protease inhibitor in Bombyx mori is involved in defense against Beauveria bassiana. Insect Biochem. Mol. Biol. 2012, 42, 766–775. [Google Scholar] [CrossRef]
- Li, Y.; Zhao, P.; Liu, H.; Guo, X.; He, H.; Zhu, R.; Xiang, Z.; Xia, Q. TIL-type protease inhibitors may be used as targeted resistance factors to enhance silkworm defenses against invasive fungi. Insect Biochem. Mol. Biol. 2015, 57, 11–19. [Google Scholar] [CrossRef]
- Li, Y.S.; Liu, H.W.; Zhu, R.; Xia, Q.Y.; Zhao, P. Protease inhibitors in Bombyx mori silk might participate in protecting the pupating larva from microbial infection. Insect Sci. 2016, 23, 835–842. [Google Scholar] [CrossRef]
- Guo, X.; Dong, Z.; Zhang, Y.; Li, Y.; Liu, H.; Xia, Q.; Zhao, P. Proteins in the Cocoon of Silkworm Inhibit the Growth of Beauveria bassiana. PLoS ONE 2016, 11, e0151764. [Google Scholar] [CrossRef]
- Huang, K.; Strynadka, N.C.; Bernard, V.D.; Peanasky, R.J.; James, M.N. The molecular structure of the complex of Ascaris chymotrypsin/elastase inhibitor with porcine elastase. Structure 1994, 2, 679–689. [Google Scholar] [CrossRef] [Green Version]
- Cierpicki, T.; Bania, J.; Otlewski, J. NMR solution structure of Apis mellifera chymotrypsin/cathepsin G inhibitor-1 (AMCI-1): Structural similarity with Ascaris protease inhibitors. Protein Sci. 2000, 9, 976–984. [Google Scholar] [CrossRef] [Green Version]
- Rosengren, K.J.; Daly, N.L.; Scanlon, M.J.; Craik, D.J. Solution structure of BSTI: A new trypsin inhibitor from skin secretions of Bombina bombina. Biochemistry 2001, 40, 4601–4609. [Google Scholar] [CrossRef]
- Bania, J.; Stachowiak, D.; Polanowski, A. Primary structure and properties of the cathepsin G/chymotrypsin inhibitor from the larval hemolymph of Apis mellifera. Eur. J. Biochem. 1999, 262, 680–687. [Google Scholar] [CrossRef]
- Mignogna, G.; Pascarella, S.; Wechselberger, C.; Hinterleitner, C.; Mollay, C.; Amiconi, G.; Barra, D.; Kreil, G. BSTI, a trypsin inhibitor from skin secretions of Bombina bombina related to protease inhibitors of nematodes. Protein Sci. 1996, 5, 357–362. [Google Scholar] [CrossRef] [Green Version]
- Babin, D.R.; Peanasky, R.J.; Goos, S.M. The isoinhibitors of chymotrypsin/elastase from Ascaris lumbricoides: The primary structure. Arch. Biochem. Biophys. 1984, 232, 143–161. [Google Scholar] [CrossRef]
- Li, Y.; Liu, H.; Zhu, R.; Xia, Q.; Zhao, P. Loss of second and sixth conserved cysteine residues from trypsin inhibitor-like cysteine-rich domain-type protease inhibitors in Bombyx mori may induce activity against microbial proteases. Peptides 2016, 86, 13–23. [Google Scholar] [CrossRef]
- Khamrui, S.; Dasgupta, J.; Dattagupta, J.K.; Sen, U. Single mutation at P1 of a chymotrypsin inhibitor changes it to a trypsin inhibitor: X-ray structural (2.15 A) and biochemical basis. Biochim. Biophys. Acta 2005, 1752, 65–72. [Google Scholar] [CrossRef]
- Legowska, A.; Debowski, D.; Lesner, A.; Wysocka, M.; Rolka, K. Introduction of non-natural amino acid residues into the substrate-specific P1 position of trypsin inhibitor SFTI-1 yields potent chymotrypsin and cathepsin G inhibitors. Bioorg. Med. Chem. 2009, 17, 3302–3307. [Google Scholar] [CrossRef]
- Rojas, L.; Cabrera-Munoz, A.; Gil Pradas, D.; Gonzalez, J.B.; Alonso-Del-Rivero, M.; Gonzalez-Gonzalez, Y. Arginine substitution by alanine at the P1 position increases the selectivity of CmPI-II, a non-classical Kazal inhibitor. Biochem. Biophys. Rep. 2021, 26, 101008. [Google Scholar] [CrossRef]
- Kojima, S.; Fushimi, N.; Ikeda, A.; Kumagai, I.; Miura, K. Secretory production of chicken ovomucoid domain 3 by Escherichia coli and alteration of inhibitory specificity toward proteases by substitution of the P1 site residue. Gene 1994, 143, 239–243. [Google Scholar]
- Ding, L.; Hao, J.; Luo, X.; Chen, Z. Engineering varied serine protease inhibitors by converting P1 site of BF9, a weakly active Kunitz-type animal toxin. Int. J. Biol. Macromol. 2018, 120, 1190–1197. [Google Scholar] [CrossRef]
- Hu, C.; Cui, J.; Ren, F.; Peng, C. Enzyme hydrolysis of silk fibroin and the anti-diabetic activity of the hydrolysates. Int. J. Food Eng. 2008, 4, 13. [Google Scholar] [CrossRef]
- Jeong, J.; Hur, W. Even-numbered peptides from a papain hydrolysate of silk fibroin. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2010, 878, 836–840. [Google Scholar] [CrossRef]
- Xu, Y.; He, G. Properties of elastase from Bacillus sp. EL31410 hydrolyzing soluble silk fibroin. Food Ferment. Ind. 2008, 34, 28–31. [Google Scholar]
- Cheng, W.; Chen, J.; Zhang, Y. Comparison of hydrolysis of silk fibroin by different proteases. J. Silk 2003, 1, 30–32. [Google Scholar]
- Spix, B.; Butz, E.S.; Chen, C.C.; Rosato, A.S.; Tang, R.; Jeridi, A.; Kudrina, V.; Plesch, E.; Wartenberg, P.; Arlt, E.; et al. Lung emphysema and impaired macrophage elastase clearance in mucolipin 3 deficient mice. Nat. Commun. 2022, 13, 318. [Google Scholar] [CrossRef]
- Suki, B.; Bartolak-Suki, E.; Rocco, P.R.M. Elastase-Induced Lung Emphysema Models in Mice. Methods Mol. Biol. 2017, 1639, 67–75. [Google Scholar]
- Wang, Z.; Chen, F.; Zhai, R.; Zhang, L.; Su, L.; Lin, X.; Thompson, T.; Christiani, D.C. Plasma neutrophil elastase and elafin imbalance is associated with acute respiratory distress syndrome (ARDS) development. PLoS ONE 2009, 4, e4380. [Google Scholar] [CrossRef] [Green Version]
- John, D.S.; Aschenbach, J.; Kruger, B.; Sendler, M.; Weiss, F.U.; Mayerle, J.; Lerch, M.M.; Aghdassi, A.A. Deficiency of cathepsin C ameliorates severity of acute pancreatitis by reduction of neutrophil elastase activation and cleavage of E-cadherin. J. Biol. Chem. 2019, 294, 697–707. [Google Scholar] [CrossRef] [Green Version]
- Trzybulska, D.; Olewicz-Gawlik, A.; Graniczna, K.; Kisiel, K.; Moskal, M.; Cieslak, D.; Sikora, J.; Hrycaj, P. Quantitative analysis of elastase and cathepsin G mRNA levels in peripheral blood CD14(+) cells from patients with rheumatoid arthritis. Cell. Immunol. 2014, 292, 40–44. [Google Scholar] [CrossRef]
- Bazzichi, L.; Ciompi, M.L.; Betti, L.; Rossi, A.; Melchiorre, D.; Fiorini, M.; Giannaccini, G.; Lucacchini, A. Impaired glutathione reductase activity and levels of collagenase and elastase in synovial fluid in rheumatoid arthritis. Clin. Exp. Rheumatol. 2002, 20, 761–766. [Google Scholar]
- Fujita, J.; Nelson, N.L.; Daughton, D.M.; Dobry, C.A.; Spurzem, J.R.; Irino, S.; Rennard, S.I. Evaluation of elastase and antielastase balance in patients with chronic bronchitis and pulmonary emphysema. Am. Rev. Respir. Dis. 1990, 142, 57–62. [Google Scholar] [CrossRef]
- Kumasaka, R.; Nakamura, N.; Fujita, T.; Murakami, R.; Shimada, M.; Osawa, H.; Yamabe, H.; Okumura, K. Beneficial effect of neutrophil elastase inhibitor on anti-Thy1.1 nephritis in rats. Nephrology 2008, 13, 27–32. [Google Scholar] [CrossRef]
- Henriksen, P.A.; Sallenave, J.M. Human neutrophil elastase: Mediator and therapeutic target in atherosclerosis. Int. J. Biochem. Cell. Biol. 2008, 40, 1095–1100. [Google Scholar] [CrossRef]
- Marino, F.; Tozzi, M.; Schembri, L.; Ferraro, S.; Tarallo, A.; Scanzano, A.; Legnaro, M.; Castelli, P.; Cosentino, M. Production of IL-8, VEGF and Elastase by Circulating and Intraplaque Neutrophils in Patients with Carotid Atherosclerosis. PLoS ONE 2015, 10, e0124565. [Google Scholar] [CrossRef] [Green Version]
- Mirea, A.M.; Toonen, E.J.M.; van den Munckhof, I.; Munsterman, I.D.; Tjwa, E.; Jaeger, M.; Oosting, M.; Schraa, K.; Rutten, J.H.W.; van der Graaf, M.; et al. Increased proteinase 3 and neutrophil elastase plasma concentrations are associated with non-alcoholic fatty liver disease (NAFLD) and type 2 diabetes. Mol. Med. 2019, 25, 16. [Google Scholar] [CrossRef] [Green Version]
- Shu, L.; Zhong, L.; Xiao, Y.; Wu, X.; Liu, Y.; Jiang, X.; Tang, T.; Hoo, R.; Zhou, Z.; Xu, A. Neutrophil elastase triggers the development of autoimmune diabetes by exacerbating innate immune responses in pancreatic islets of non-obese diabetic mice. Clin. Sci. 2020, 134, 1679–1696. [Google Scholar] [CrossRef]
- Moroy, G.; Alix, A.J.; Sapi, J.; Hornebeck, W.; Bourguet, E. Neutrophil elastase as a target in lung cancer. Anti-Cancer Agents Med. Chem. 2012, 12, 565–579. [Google Scholar] [CrossRef]
- Sato, T.; Takahashi, S.; Mizumoto, T.; Harao, M.; Akizuki, M.; Takasugi, M.; Fukutomi, T.; Yamashita, J. Neutrophil elastase and cancer. Surg. Oncol. 2006, 15, 217–222. [Google Scholar] [CrossRef]
- Tsuji, N.; Moriwaki, S.; Suzuki, Y.; Takema, Y.; Imokawa, G. The role of elastases secreted by fibroblasts in wrinkle formation: Implication through selective inhibition of elastase activity. Photochem. Photobiol. 2001, 74, 283–290. [Google Scholar] [CrossRef]
- Imokawa, G. Mechanism of UVB-induced wrinkling of the skin: Paracrine cytokine linkage between keratinocytes and fibroblasts leading to the stimulation of elastase. J. Investig. Dermatol. Symp. Proc. 2009, 40, 36–43. [Google Scholar]
- Shinguh, Y.; Yamazaki, A.; Inamura, N.; Fujie, K.; Okamoto, M.; Nakahara, K.; Notsu, Y.; Okuhara, M.; Ono, T. Biochemical and pharmacological characterization of FR134043, a novel elastase inhibitor. Eur. J. Pharmacol. 1998, 345, 299–308. [Google Scholar] [CrossRef]
- Patel, A.; Gandhi, K.; Shah, S.; Patel, D.; Chhatbar, S.; Shah, D.; Patel, S.; Patel, H.; Bambharoliya, T. In silico Study and Solvent-free one-pot Synthesis of Tetrahydropyrimidine derivatives by Mechanochemistry Approach for Targeting Human Neutrophil Elastase against Lung Cancer. Curr. Comput.-Aided Drug Des. 2022, 18, 293–306. [Google Scholar] [CrossRef]
- Sahebnasagh, A.; Saghafi, F.; Safdari, M.; Khataminia, M.; Sadremomtaz, A.; Talaei, Z.; Rezai Ghaleno, H.; Bagheri, M.; Habtemariam, S.; Avan, R. Neutrophil elastase inhibitor (sivelestat) may be a promising therapeutic option for management of acute lung injury/acute respiratory distress syndrome or disseminated intravascular coagulation in COVID-19. J. Clin. Pharm. Ther. 2020, 45, 1515–1519. [Google Scholar] [CrossRef]
- Kumar, J.P.; Mandal, B.B. Inhibitory role of silk cocoon extract against elastase, hyaluronidase and UV radiation-induced matrix metalloproteinase expression in human dermal fibroblasts and keratinocytes. Photochem. Photobiol. Sci. 2019, 18, 1259–1274. [Google Scholar] [CrossRef]
- Saric, M.; Scheibel, T. Engineering of silk proteins for materials applications. Curr. Opin. Biotechnol. 2019, 60, 213–220. [Google Scholar] [CrossRef]
- Kundu, B.; Kurland, N.E.; Yadavalli, V.K.; Kundu, S.C. Isolation and processing of silk proteins for biomedical applications. Int. J. Biol. Macromol. 2014, 70, 70–77. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhao, P.; Dong, Z.; Wang, D.; Guo, P.; Guo, X.; Song, Q.; Zhang, W.; Xia, Q. Comparative proteome analysis of multi-layer cocoon of the silkworm, Bombyx mori. PLoS ONE 2015, 10, e0123403. [Google Scholar] [CrossRef]
- Dong, Z.; Zhao, P.; Wang, C.; Zhang, Y.; Chen, J.; Wang, X.; Lin, Y.; Xia, Q. Comparative proteomics reveal diverse functions and dynamic changes of Bombyx mori silk proteins spun from different development stages. J. Proteome Res. 2013, 12, 5213–5222. [Google Scholar] [CrossRef]
- Zhao, B.; Xu, P.; Jiang, L.; Paaske, B.; Kromann-Hansen, T.; Jensen, J.K.; Sorensen, H.P.; Liu, Z.; Nielsen, J.T.; Christensen, A.; et al. A cyclic peptidic serine protease inhibitor: Increasing affinity by increasing peptide flexibility. PLoS ONE 2014, 9, e115872. [Google Scholar] [CrossRef] [Green Version]
- Lin, Z.; Jiang, L.; Yuan, C.; Jensen, J.K.; Zhang, X.; Luo, Z.; Furie, B.C.; Furie, B.; Andreasen, P.A.; Huang, M. Structural basis for recognition of urokinase-type plasminogen activator by plasminogen activator inhibitor-1. J. Biol. Chem. 2011, 286, 7027–7032. [Google Scholar] [CrossRef] [Green Version]
- Hosseini, M.; Jiang, L.; Sorensen, H.P.; Jensen, J.K.; Christensen, A.; Fogh, S.; Yuan, C.; Andersen, L.M.; Huang, M.; Andreasen, P.A.; et al. Elucidation of the contribution of active site and exosite interactions to affinity and specificity of peptidylic serine protease inhibitors using non-natural arginine analogs. Mol. Pharmacol. 2011, 80, 585–597. [Google Scholar] [CrossRef]
- Eguchi, M.; Itoh, M.; Chou, L.-Y.; Nishino, K. Purification and characterization of a fungal protease specific protein inhibitor. Comp. Biochem. Physiol. 1993, 104B, 537–543. [Google Scholar]
- Taniguchi, M.; Atiwetin, P.; Hirai, T.; Itoh, M.; Harada, S.; Hara, S.; Kamei, K. Interaction of subtilisin BPN’ and recombinant fungal protease inhibitor F from silkworm with substituted P1 site residues. Biosci. Biotechnol. Biochem. 2006, 70, 1262–1264. [Google Scholar] [CrossRef]
- Kojima, S.; Nishiyama, Y.; Kumagai, I.; Miura, K. Inhibition of subtilisin BPN’ by reaction site P1 mutants of Streptomyces subtilisin inhibitor. J. Biochem. 1991, 109, 377–382. [Google Scholar] [CrossRef]
- Perona, J.J.; Craik, C.S. Structural basis of substrate specificity in the serine proteases. Protein Sci. 1995, 4, 337–360. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.J.; Dong, K.C.; Eigenbrot, C.; van Lookeren Campagne, M.; Kirchhofer, D. Structures of neutrophil serine protease 4 reveal an unusual mechanism of substrate recognition by a trypsin-fold protease. Structure 2014, 22, 1333–1340. [Google Scholar] [CrossRef] [Green Version]
- Kojima, S.; Obata, S.; Kumagai, I.; Miura, K. Alteration of the specificity of the Streptomyces subtilisin inhibitor by gene engineering. Bio/Technology 1990, 8, 449–452. [Google Scholar] [CrossRef]
- Beck, M.H.; Strand, M.R. A novel polydnavirus protein inhibits the insect prophenoloxidase activation pathway. Proc. Natl. Acad Sci. USA 2007, 104, 19267–19272. [Google Scholar] [CrossRef] [Green Version]
- Gu, Q.J.; Zhou, S.M.; Zhou, Y.N.; Huang, J.H.; Shi, M.; Chen, X.X. A trypsin inhibitor-like protein secreted by Cotesia vestalis teratocytes inhibits hemolymph prophenoloxidase activation of Plutella xylostella. J. Insect Physiol. 2019, 116, 41–48. [Google Scholar] [CrossRef]
- Luo, Z.; Yang, J.; Zhang, J.; Meng, G.; Lu, Q.; Yang, X.; Zhao, P.; Li, Y. Physicochemical Properties and Elimination of the Activity of Anti-Nutritional Serine Protease Inhibitors from Mulberry Leaves. Molecules 2022, 27, 1820. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mutants | Templates | Desired Mutations | DNA Polymerases | Primer Sequences a |
|---|---|---|---|---|
| BmSPI38(G54E) | BmSPI38 | G54E | FastPfu | F: 5′-CGCTTGCGTCGAAGCCCTCATTCAAACC-3′ R: 5′-TGAATGAGGGCTTCGACGCAAGCGGTG-3′ |
| BmSPI38(G54D) | BmSPI38 | G54D | FastPfu | F: 5′-CCGCTTGCGTCGATGCCCTCATTCAAACC-3′ R: 5′- GTTGAATGAGGGCATCGACGCAAGCGGTG-3′ |
| BmSPI38(G54R) | BmSPI38(G54S) | S54R | FastPfu | F: 5′-ACCGCTTGCGTCCGCGCCCTCATTC-3′ R: 5′-AATGAGGGCGCGGACGCAAGCGGTG-3′ |
| BmSPI38(G54K) | BmSPI38 | G54K | FastPfu | F: 5′-ACCGCTTGCGTCAAAGCCCTCATTCAAACC-3′ R: 5′-GTTTGAATGAGGGCTTTGACGCAAGCGGTG-3′ |
| BmSPI38(G54H) | BmSPI38(G54D) | D54H | FastPfu | F: 5′-ACCGCTTGCGTCCATGCCCTCATTCAAAC-3′ R: 5′-TGAATGAGGGCATGGACGCAAGCGGTG-3′ |
| BmSPI38(G54M) | BmSPI38 | G54M | FastPfu | F: 5′-CCGCTTGCGTCATGGCCCTCATTCAAACC-3′ R: 5′-GTTTGAATGAGGGCCATGACGCAAGCGGTG-3′ |
| BmSPI38(G54L) | BmSPI38 | G54L | FastPfu | F: 5′-CACCGCTTGCGTCCTGGCCCTCATTC-3′ R: 5′-GAATGAGGGCCAGGACGCAAGCGGTG-3′ |
| BmSPI38(G54I) | BmSPI38 | G54I | FastPfu | F: 5′-CCGCTTGCGTCATTGCCCTCATTCAAACC-3′ R: 5′-GTTTGAATGAGGGCAATGACGCAAGCGGTG-3′ |
| BmSPI38(G54F) | BmSPI38(G54I) | I54F | FastPfu | F: 5′-CACCGCTTGCGTCTTTGCCCTCATTCAAAC-3′ R: 5′-TTTGAATGAGGGCAAAGACGCAAGCGGTG-3′ |
| BmSPI38(G54W) | BmSPI38(G54C) | C54W | FastPfu Fly | F: 5′-CCGCTTGCGTCTGGGCCCTCATTCAAACC-3′ R: 5′-TTTGAATGAGGGCCCAGACGCAAGCGGTG-3′ |
| BmSPI38(G54N) | BmSPI38(G54Y) | Y54N | FastPfu Fly | F: 5′-CCGCTTGCGTCAACGCCCTCATTCAAAC-3′ R: 5′-TGAATGAGGGCGTTGACGCAAGCGGTG-3′ |
| BmSPI38(G54Q) | BmSPI38(G54P) | P54Q | EasyPfu | F: 5′-ACCGCTTGCGTCCAGGCCCTCATTCAAAC-3′ R: 5′-TGAATGAGGGCCTGGACGCAAGCGGTG-3′ |
| BmSPI38(G54Y) | BmSPI38 | G54Y | FastPfu | F: 5′-CACCGCTTGCGTCTACGCCCTCATTCAAAC-3′ R: 5′-GTTTGAATGAGGGCGTAGACGCAAGCGGTG-3′ |
| BmSPI38(G54C) | BmSPI38 | G54C | FastPfu | F: 5′-CACCGCTTGCGTCTGCGCCCTCATTC-3′ R: 5′-GAATGAGGGCGCAGACGCAAGCGGTG-3′ |
| BmSPI38(G54S) | BmSPI38 | G54S | FastPfu | F: 5′-CCGCTTGCGTCAGCGCCCTCATTCAAAC-3′ R: 5′-GAATGAGGGCGCTGACGCAAGCGGTG-3′ |
| BmSPI38(G54T) | BmSPI38 | G54T | FastPfu | F: 5′-CCGCTTGCGTCACCGCCCTCATTCAAAC-3′ R: 5′-TTGAATGAGGGCGGTGACGCAAGCGG-3′ |
| BmSPI38(G54A) | BmSPI38(G54P) | P54A | EasyPfu | F: 5′-CCGCTTGCGTCGCGGCCCTCATTCAAAC-3′ R: 5′-TGAATGAGGGCCGCGACGCAAGCGGTG-3′ |
| BmSPI38(G54P) | BmSPI38 | G54P | FastPfu | F: 5′-CGCTTGCGTCCCGGCCCTCATTCAAAC-3′ R: 5′-AATGAGGGCCGGGACGCAAGCGGTG-3′ |
| BmSPI38(G54V) | BmSPI38(G54L) | L54V | FastPfu Fly | F: 5′-ACCGCTTGCGTCGTGGCCCTCATTCAAAC-3′ R: 5′-TGAATGAGGGCCACGACGCAAGCGGTG-3′ |
| Mutants | Templates | Desired Mutations | DNA Polymerases | Primer Sequences a |
|---|---|---|---|---|
| BmSPI39(A56E) | BmSPI39(A56D) | D56E | FastPfu | F: 5′-CCAGCTGCGTAGAAGCATTGCTCCCAACATG-3′ R: 5′-TGGGAGCAATGCTTCTACGCAGCTGGTGTG-3′ |
| BmSPI39(A56D) | BmSPI39 | A56D | FastPfu Fly | F: 5′-CACCAGCTGCGTAGATGCATTGCTCCCAAC-3′ R: 5′-GTTGGGAGCAATGCATCTACGCAGCTGGTG-3′ |
| BmSPI39(A56R) | BmSPI39 | A56R | FastPfu | F: 5′-CAGCTGCGTACGCGCATTGCTCCCAAC-3′ R: 5′-GGGAGCAATGCGCGTACGCAGCTGGTG-3′ |
| BmSPI39(A56K) | BmSPI39(A56L) | L56K | FastPfu Fly | F: 5′-CACCAGCTGCGTAAAGGCATTGCTCGGAAC-3′ R: 5′-TTGGGAGCAATGCCTTTACGCAGCTGGTG-3′ |
| BmSPI39(A56H) | BmSPI39 | A56H | FastPfu | F:5′-CACCAGCTGCGTACATGCATTGCTCCCAAC-3′ R: 5′-GGGAGCAATGCATGTACGCAGCTGGTGTG-3′ |
| BmSPI39(A56M) | BmSPI39(A56L) | L56M | FastPfu Fly | F: 5′-CACCAGCTGCGTAATGGCATTGCTCCCAAC-3′ R: 5′-TTGGGAGCAATGCCATTACGCAGCTGGTG-3′ |
| BmSPI39(A56L) | BmSPI39 | A56L | FastPfu | F: 5′-CACCAGCTGCGTACTGGCATTGCTCCCAAC-3 ′ R: 5′-GGGAGCAATGCCAGTACGCAGCTGGTGTG-3 ′ |
| BmSPI39(A56I) | BmSPI39 | A56I | FastPfu | F: 5′-CCAGCTGCGTAATTGCATTGCTCCCAACATG-3′ R: 5′-TGGGAGCAATGCAATTACGCAGCTGGTGTG-3′ |
| BmSPI39(A56F) | BmSPI39(A56I) | I56F | FastPfu | F: 5′-CACCAGCTGCGTATTTGCATTGCTCCCAAC-3′ R: 5′-TGGGAGCAATGCAAATACGCAGCTGGTGTG-3′ |
| BmSPI39(A56W) | BmSPI39 | A56W | FastPfu | F: 5′-CACCAGCTGCGTATGGGCATTGCTCCCAAC-3′ R: 5′-GGGAGCAATGCCCATACGCAGCTGGTGTG-3′ |
| BmSPI39(A56N) | BmSPI39(A56Y) | Y56N | EasyPfu | F: 5′-CACCAGCTGCGTAAACGCATTGCTCCCAAC-3′ R: 5′-TTGGGAGCAATGCGTTTACGCAGCTGGTG-3′ |
| BmSPI39(A56Q) | BmSPI39(A56L) | L56Q | EasyPfu | F:5′-CACCAGCTGCGTACAGGCATTGCTCCCAAC-3′ R: 5′-TTGGGAGCAATGCCTGTACGCAGCTGGTG-3′ |
| BmSPI39(A56Y) | BmSPI39 | A56Y | FastPfu | F: 5′-CCAGCTGCGTATACGCATTGCTCCCAACATG-3′ R: 5′-TGGGAGCAATGCGTATACGCAGCTGGTGTG-3′ |
| BmSPI39(A56C) | BmSPI39 | A56C | FastPfu | F: 5′-CCAGCTGCGTATGCGCATTGCTCCCAAC-3′ R: 5′-GGGAGCAATGCGCATACGCAGCTGGTG-3′ |
| BmSPI39(A56S) | BmSPI39 | A56S | FastPfu | F: 5′-CCAGCTGCGTAAGCGCATTGCTCCCAAC-3′ R: 5′-GGGAGCAATGCGCTTACGCAGCTGGTG-3′ |
| BmSPI39(A56T) | BmSPI39 | A56T | FastPfu Fly | F: 5′-CCAGCTGCGTAACCGCATTGCTCCCAAC-3′ R:5′-GGGAGCAATGCGGTTACGCAGCTGGTG-3′ |
| BmSPI39(A56G) | BmSPI39 | A56G | FastPfu | F: 5′-CCAGCTGCGTAGGCGCATTGCTCCCAAC-3′ R: 5′-GGGAGCAATGCGCCTACGCAGCTGGTG-3′ |
| BmSPI39(A56P) | BmSPI39 | A56P | FastPfu | F: 5′-CACCAGCTGCGTACCGGCATTGCTCCC-3′ R: 5′-GGGAGCAATGCCGGTACGCAGCTGGTG-3′ |
| BmSPI39(A56V) | BmSPI39(A56L) | L56V | EasyPfu | F: 5′-CACCAGCTGCGTAGTGGCATTGCTCCCAAC-3′ R: 5′-TTGGGAGCAATGCCACTACGCAGCTGGTG-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Wei, M.; Zhang, J.; Zhu, R.; Wang, Y.; Zhang, Z.; Chen, C.; Zhao, P. Amino Acid Substitutions at P1 Position Change the Inhibitory Activity and Specificity of Protease Inhibitors BmSPI38 and BmSPI39 from Bombyx mori. Molecules 2023, 28, 2073. https://doi.org/10.3390/molecules28052073
Li Y, Wei M, Zhang J, Zhu R, Wang Y, Zhang Z, Chen C, Zhao P. Amino Acid Substitutions at P1 Position Change the Inhibitory Activity and Specificity of Protease Inhibitors BmSPI38 and BmSPI39 from Bombyx mori. Molecules. 2023; 28(5):2073. https://doi.org/10.3390/molecules28052073
Chicago/Turabian StyleLi, Youshan, Meng Wei, Jie Zhang, Rui Zhu, Yuan Wang, Zhaofeng Zhang, Changqing Chen, and Ping Zhao. 2023. "Amino Acid Substitutions at P1 Position Change the Inhibitory Activity and Specificity of Protease Inhibitors BmSPI38 and BmSPI39 from Bombyx mori" Molecules 28, no. 5: 2073. https://doi.org/10.3390/molecules28052073