

Kavalactone Kawain Impedes Urothelial Tumorigenesis in UPII-Mutant Ha-Ras Mice via Inhibition of mTOR Signaling and Alteration of Cancer Metabolism

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
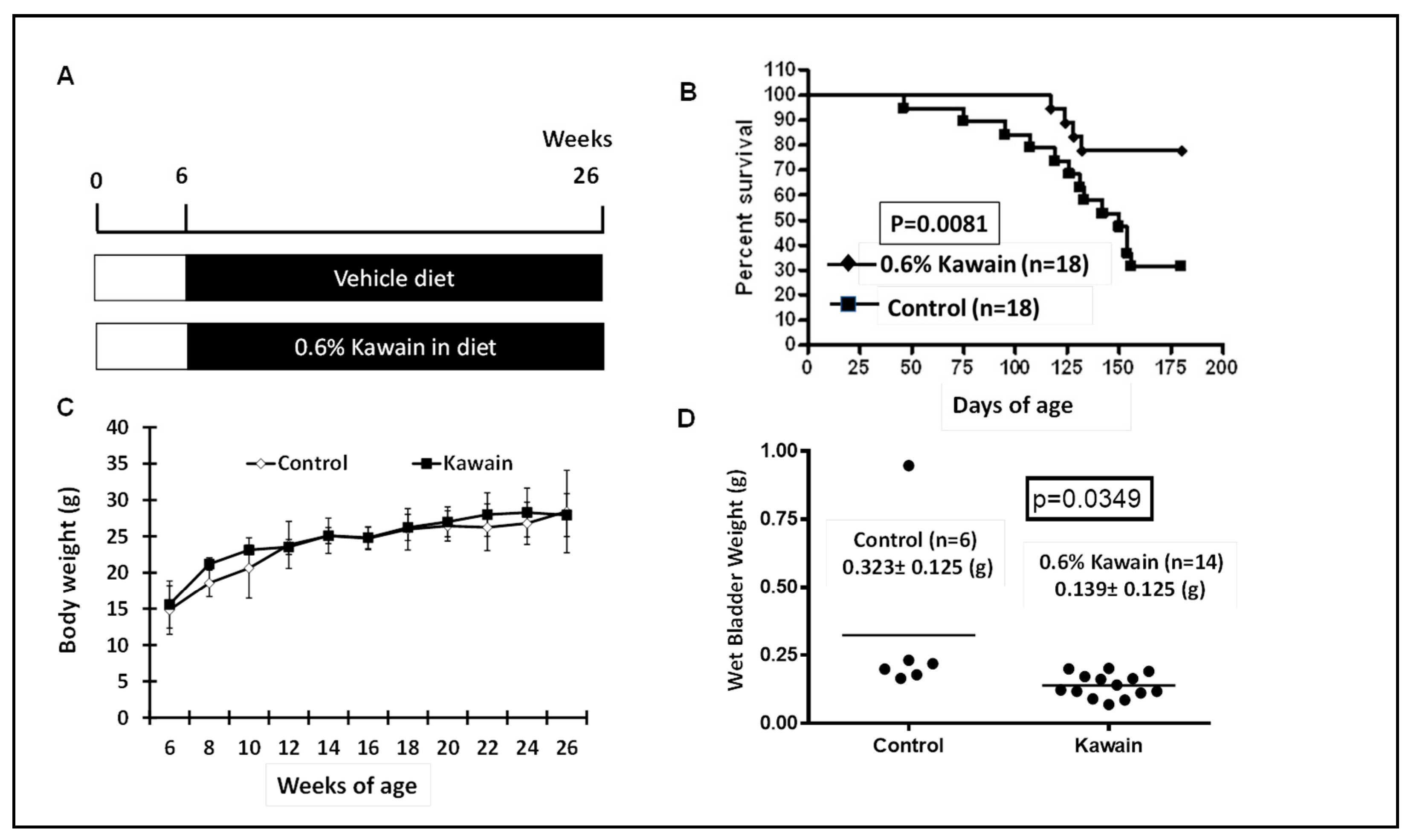
2.1. Kawain-Formulated Diet Reduces Tumor Burden and Increases the Survival of UPII-Mutant Ha-Ras Transgenic Mice
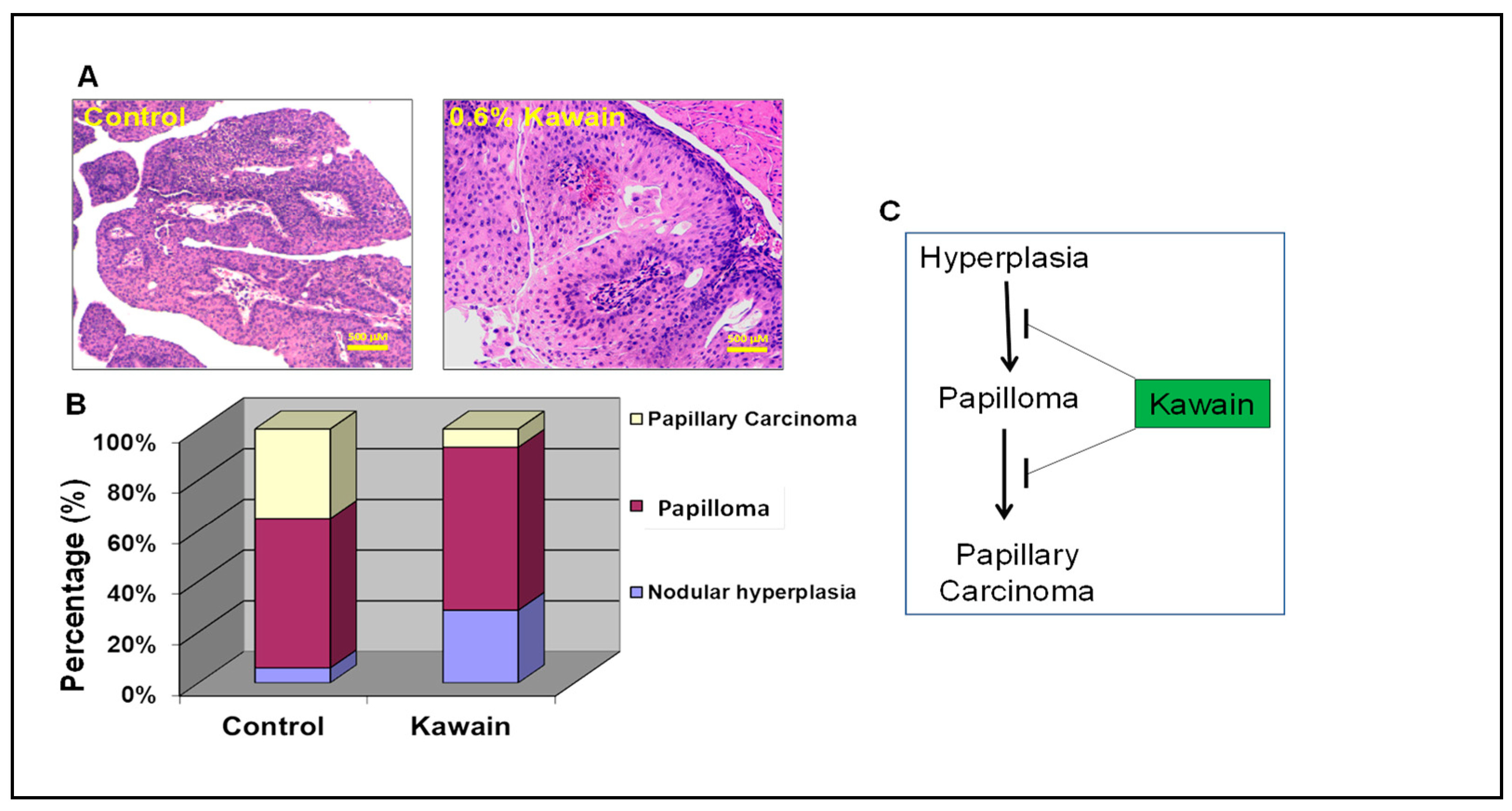
2.2. Kawain Diet Inhibits the Pathological Progression of Hyperplasia to Papillary Carcinoma
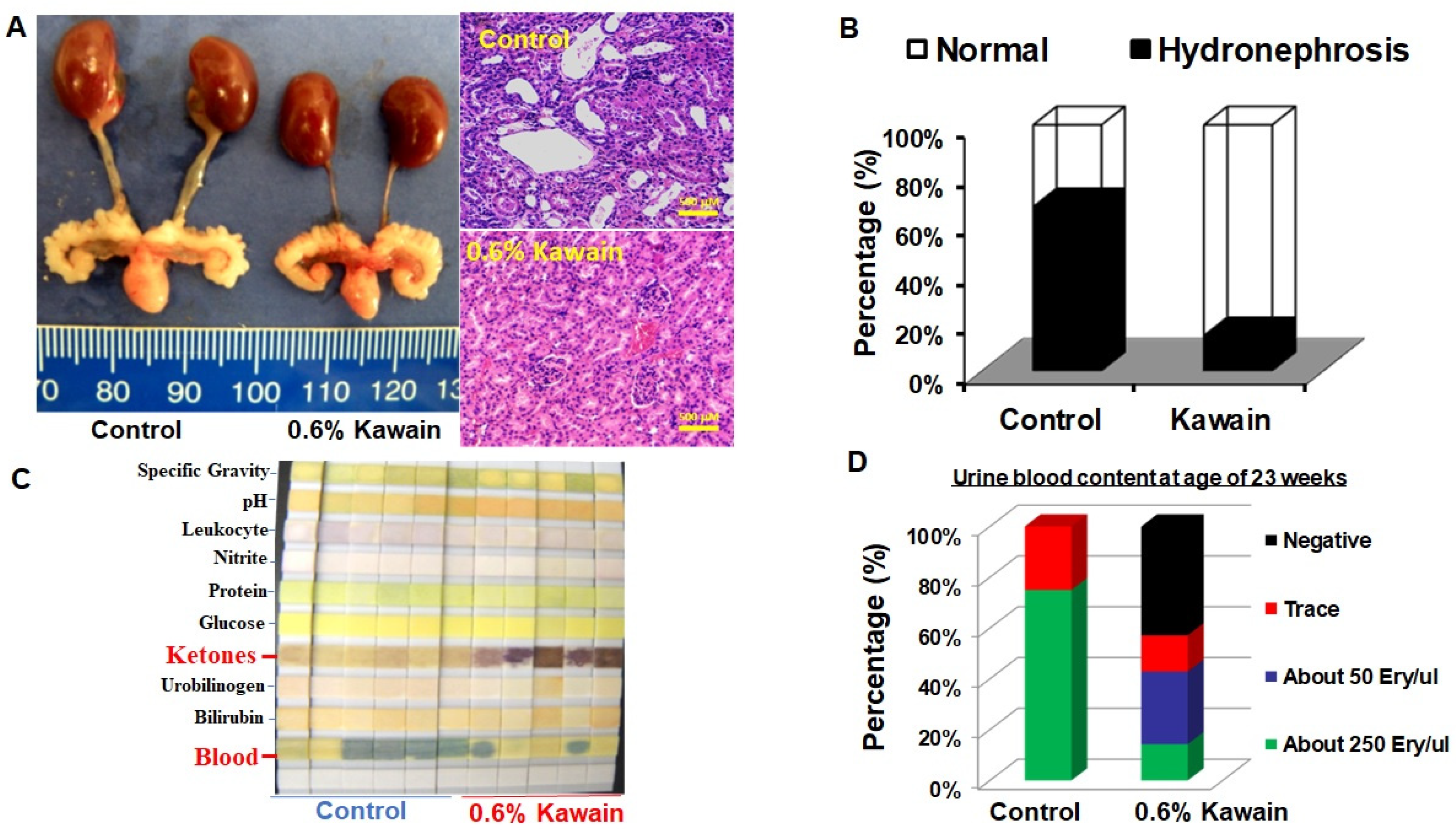
2.3. Kawain Diet Reduces the Incidence of Hydronephrosis and Hematouria
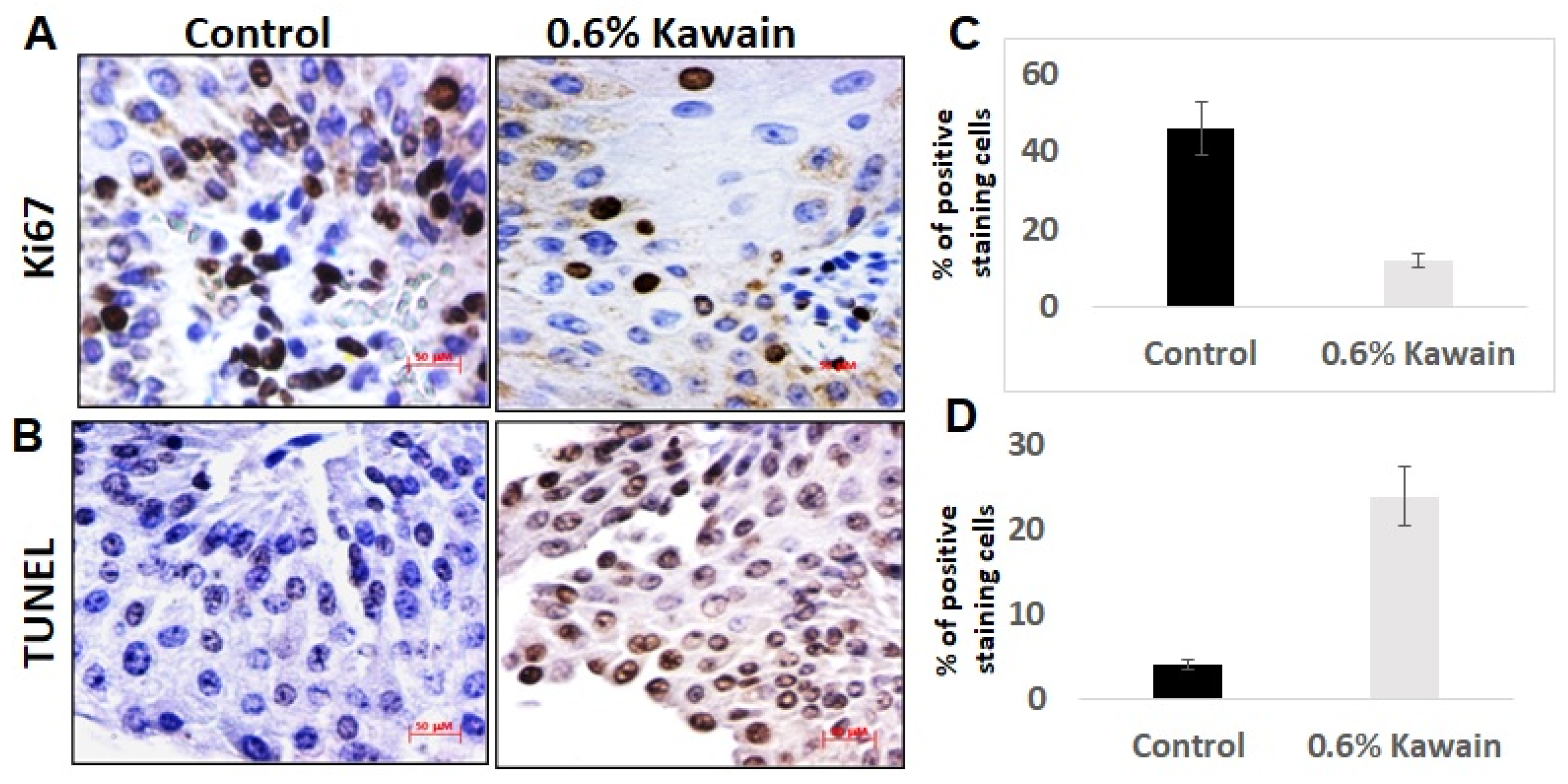
2.4. Kawain Diet Decreases Cell Proliferation and Increases Apoptosis In Vivo
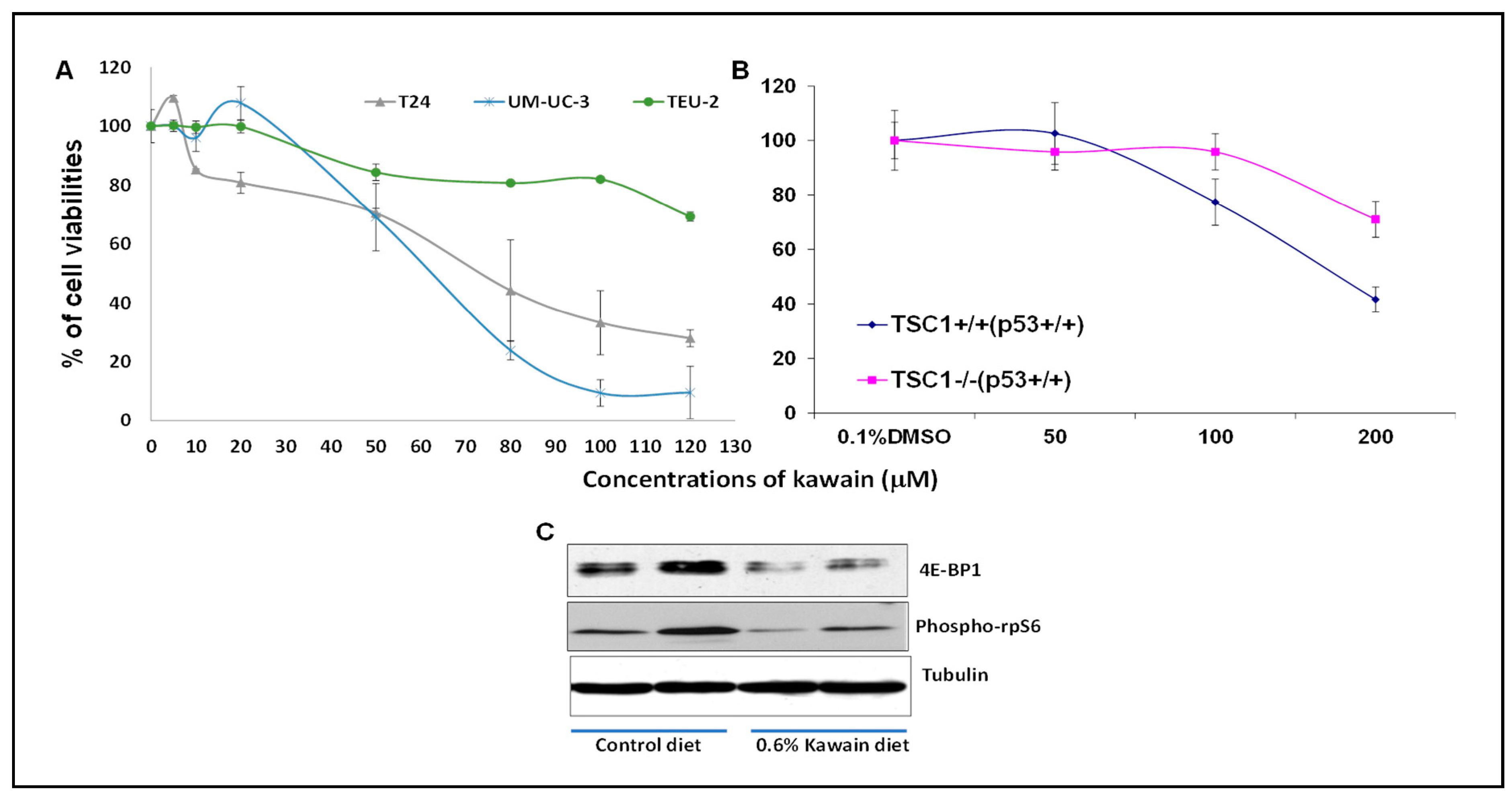
2.5. Kawain Selectively Inhibits the Growth of Bladder Cancer Cell Lines and TSC1 Positive Cells, and Kawain Diet Attenuates the mTOR Signaling In Vivo
2.6. Kawain Diet Changes the Metabolism of UPII-Ha-Ras Bladder Tumors
3. Materials and Methods
3.1. Chemopreventive Efficay of of Dietary Kawain on Mutant Ha-Ras-Driven Urothelial Carcinogenesis
3.2. Histology and Immunohistochemistry (IHC) Analyses
3.3. In Vitro Cell Proliferation Assay
3.4. Western Blotting Analysis
3.5. Metabolomic Profiling
3.6. Statistical Evaluation
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Key Statistics for Bladder Cancer. Available online: https://www.cancer.org/cancer/bladder-cancer/about/key-statistics.html (accessed on 27 December 2022).
- Jordan, B.; Meeks, J.J. T1 bladder cancer: Current considerations for diagnosis and management. Nat. Rev. Urol. 2019, 16, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Botteman, M.F.; Pashos, C.L.; Redaelli, A.; Laskin, B.; Hauser, R. The health economics of bladder cancer: A comprehensive review of the published literature. Pharmacoeconomics 2003, 21, 1315–1330. [Google Scholar] [PubMed]
- Wu, X.-R. Urothelial tumorigenesis: A tale of divergent pathways. Nat. Rev. Cancer 2005, 5, 713–725. [Google Scholar] [CrossRef] [PubMed]
- Meeks, J.J.; Lerner, S.P. Molecular Landscape of Non-Muscle Invasive Bladder Cancer. Cancer Cell 2017, 32, 550–551. [Google Scholar] [CrossRef] [PubMed]
- Mo, L.; Zheng, X.; Huang, H.-Y.; Shapiro, E.; Lepor, H.; Cordon-Cardo, C.; Sun, T.-T.; Wu, X.-R. Hyperactivation of Ha-ras oncogene, but not Ink4a/Arf deficiency, triggers bladder tumorigenesis. J. Clin. Investig. 2007, 117, 314–325. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Yokoyama, N.N.; Blair, C.A.; Li, X.; Avizonis, D.; Wu, X.-R.; Uchio, E.; Youssef, R.; McClelland, M.; Pollak, M.; et al. High Sensitivity of an Ha-RAS Transgenic Model of Superficial Bladder Cancer to Metformin Is Associated with ∼240-Fold Higher Drug Concentration in Urine than Serum. Mol. Cancer Ther. 2016, 15, 430–438. [Google Scholar] [CrossRef]
- Liu, Z.; Song, L.; Xie, J.; Simoneau, A.R.; Uchio, E.; Zi, X. Chemoprevention of Urothelial Cell Carcinoma Tumorigenesis by Dietary Flavokawain A in UPII-Mutant Ha-ras Transgenic Mice. Pharmaceutics 2022, 14, 496. [Google Scholar] [CrossRef]
- Huan, J.; Grivas, P.; Birch, J.; Hansel, D.E. Emerging Roles for Mammalian Target of Rapamycin (mTOR) Complexes in Bladder Cancer Progression and Therapy. Cancers 2022, 14, 1555. [Google Scholar] [CrossRef]
- Mossmann, D.; Park, S.; Hall, M.N. mTOR signalling and cellular metabolism are mutual determinants in cancer. Nat. Rev. Cancer 2018, 18, 744–757. [Google Scholar] [CrossRef]
- Wang, Y.; Narayanapillai, S.C.; Tessier, K.M.; Strayer, L.G.; Upadhyaya, P.; Hu, Q.; Kingston, R.; Salloum, R.G.; Lu, J.; Hecht, S.S.; et al. The Impact of One-week Dietary Supplementation with Kava on Biomarkers of Tobacco Use and Nitrosamine-based Carcinogenesis Risk among Active Smokers. Cancer Prev. Res. 2020, 13, 483–492. [Google Scholar] [CrossRef] [Green Version]
- Steiner, G.G. The correlation between cancer incidence and kava consumption. Hawaii Med. J. 2000, 59, 420–422. [Google Scholar] [PubMed]
- Einbond, L.; Negrin, A.; Kulakowski, D.; Wu, H.-A.; Antonetti, V.; Jalees, F.; Law, W.; Roller, M.; Redenti, S.; Kennelly, E.; et al. Traditional preparations of kava (Piper methysticum) inhibit the growth of human colon cancer cells in vitro. Phytomedicine 2017, 24, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Song, L.; Xu, S.; Tippin, M.; Meng, S.; Xie, J.; Uchio, E.; Zi, X. Kava root extracts hinder prostate cancer development and tumorigenesis by involvement of dual inhibition of MAO-A and LSD1. J. Transl. Genet. Genom. 2021, 5, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Leitzman, P.; Narayanapillai, S.C.; Balbo, S.; Zhou, B.; Upadhyaya, P.; Shaik, A.A.; O’Sullivan, M.G.; Hecht, S.S.; Lu, J.; Xing, C. Kava Blocks 4-(Methylnitrosamino)-1-(3-pyridyl)-1-Butanone–Induced Lung Tumorigenesis in Association with Reducing O6-methylguanine DNA Adduct in A/J Mice. Cancer Prev. Res. 2014, 7, 86–96. [Google Scholar] [CrossRef]
- Li, X.; Liu, Z.; Xu, X.; Blair, C.A.; Sun, Z.; Xie, J.; Lilly, M.B.; Zi, X. Kava Components Down-Regulate Expression of AR and AR Splice Variants and Reduce Growth in Patient-Derived Prostate Cancer Xenografts in Mice. PLoS ONE 2012, 7, e31213. [Google Scholar] [CrossRef]
- Wang, J.; Qi, Q.; Zhou, W.; Feng, Z.; Huang, B.; Chen, A.; Zhang, D.; Li, W.; Zhang, Q.; Jiang, Z.; et al. Inhibition of glioma growth by flavokawain B is mediated through endoplasmic reticulum stress induced autophagy. Autophagy 2018, 14, 2007–2022. [Google Scholar] [CrossRef]
- Zi, X.; Simoneau, A.R. Flavokawain A, a Novel Chalcone from Kava Extract, Induces Apoptosis in Bladder Cancer Cells by Involvement of Bax Protein-Dependent and Mitochondria-Dependent Apoptotic Pathway and Suppresses Tumor Growth in Mice. Cancer Res. 2005, 65, 3479–3486. [Google Scholar] [CrossRef]
- Narayanapillai, S.C.; Balbo, S.; Leitzman, P.; Grill, A.E.; Upadhyaya, P.; Shaik, A.A.; Zhou, B.; O’Sullivan, M.G.; Peterson, L.A.; Lu, J.; et al. Dihydromethysticin from kava blocks tobacco carcinogen 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone-induced lung tumorigenesis and differentially reduces DNA damage in A/J mice. Carcinogenesis 2014, 35, 2365–2372. [Google Scholar] [CrossRef]
- Tang, Y.; Fields, C. A UHPLC-UV Method Development and Validation for Determining Kavalactones and Flavokavains in Piper methysticum (Kava). Molecules 2019, 24, 1245. [Google Scholar] [CrossRef]
- Anderson, J.C.; Mattar, S.G.; Greenway, F.L.; Lindquist, R.J. Measuring ketone bodies for the monitoring of pathologic and therapeutic ketosis. Obes. Sci. Pr. 2021, 7, 646–656. [Google Scholar] [CrossRef]
- Jandial, D.D.; Krill, L.S.; Chen, L.; Wu, C.; Ke, Y.; Xie, J.; Hoang, B.H.; Zi, X. Induction of G2M Arrest by Flavokawain A, a Kava Chalcone, Increases the Responsiveness of HER2-Overexpressing Breast Cancer Cells to Herceptin. Molecules 2017, 22, 462. [Google Scholar] [CrossRef] [PubMed]
- Aporosa, A.; Atkins, M.; Brunton, R. Kava drinking in traditional settings: Towards understanding effects on cognitive function. Hum. Psychopharmacol. Clin. Exp. 2020, 35, e2725. [Google Scholar] [CrossRef]
- Qiu, T.; Chen, Y.; Meng, L.; Xu, T.; Zhang, H. Identification of a metabolism-related gene signature predicting overall survival for bladder cancer. Genomics 2022, 114, 110402. [Google Scholar] [CrossRef] [PubMed]
- Afonso, J.; Santos, L.L.; Morais, A.; Amaro, T.; Longatto-Filho, A.; Baltazar, M.D.F.M. Metabolic coupling in urothelial bladder cancer compartments and its correlation to tumor aggressiveness. Cell Cycle 2016, 15, 368–380. [Google Scholar] [CrossRef]
- Xia, Y.; Wang, X.; Liu, Y.; Shapiro, E.; Lepor, H.; Tang, M.-S.; Sun, T.-T.; Wu, X.-R. PKM2 Is Essential for Bladder Cancer Growth and Maintenance. Cancer Res 2022, 82, 571–585. [Google Scholar] [CrossRef]
- Xia, Y.; Liu, Y.; Yang, C.; Simeone, D.M.; Sun, T.-T.; DeGraff, D.J.; Tang, M.-S.; Zhang, Y.; Wu, X.-R. Dominant role of CDKN2B/p15INK4B of 9p21.3 tumor suppressor hub in inhibition of cell-cycle and glycolysis. Nat. Commun. 2021, 12, 2047. [Google Scholar] [CrossRef]
- Sun, C.Y.; Li, J.R.; Wang, Y.Y.; Lin, S.Y.; Ou, Y.C.; Lin, C.J.; Wang, J.D.; Liao, S.L.; Chen, C.J. Indoxyl sulfate caused behavioral abnormality and neurodegeneration in mice with unilateral nephrectomy. Aging 2021, 13, 6681–6701. [Google Scholar]
- Grzelczyk, J.; Budryn, G.; Peña-García, J.; Szwajgier, D.; Gałązka-Czarnecka, I.; Oracz, J.; Pérez-Sánchez, H. Evaluation of the inhibition of monoamine oxidase A by bioactive coffee compounds protecting serotonin degradation. Food Chem. 2021, 348, 129108. [Google Scholar] [CrossRef]
- Blanc-Lapierre, A.; Rousseau, M.-C.; Weiss, D.; El-Zein, M.; Siemiatycki, J.; Parent, M. Lifetime report of perceived stress at work and cancer among men: A case-control study in Montreal, Canada. Prev. Med. 2017, 96, 28–35. [Google Scholar] [CrossRef]
- Liu, H.; Yang, J.; Zhang, Y.; Han, J.; Yang, Y.; Zhao, Z.; Dai, X.; Wang, H.; Ding, X.; Liu, Y.; et al. Psychologic Stress Drives Progression of Malignant Tumors via DRD2/HIF1α Signaling. Cancer Res. 2021, 81, 5353–5365. [Google Scholar]
- Falcinelli, M.; Thaker, P.H.; Lutgendorf, S.K.; Conzen, S.D.; Flaherty, R.L.; Flint, M.S. The Role of Psychologic Stress in Cancer Initiation: Clinical Relevance and Potential Molecular Mechanisms. Cancer Res. 2021, 81, 5131–5140. [Google Scholar] [CrossRef] [PubMed]
- Breiden, B.; Sandhoff, K. Acid Sphingomyelinase, a Lysosomal and Secretory Phospholipase C, Is Key for Cellular Phospholipid Catabolism. Int. J. Mol. Sci. 2021, 22, 9001. [Google Scholar] [PubMed]
- Ghio, A.J.; Karoly, E.D.; Bell, L.N.; Snow, S.J.; Madden, M.C.; Soukup, J.; Cascio, W.E.; Gilmour, M.I.; Kodavanti, U.P. Ozone Exposure Increases Circulating Stress Hormones and Lipid Metabolites in Humans. Am. J. Respir. Crit. Care Med. 2016, 193, 1382–1391. [Google Scholar] [CrossRef]
- Kokabi, K.; Gorelova, O.; Zorin, B.; Didi-Cohen, S.; Itkin, M.; Malitsky, S.; Solovchenko, A.; Boussiba, S.; Khozin-Goldberg, I. Lipidome Remodeling and Autophagic Respose in the Arachidonic-Acid-Rich Microalga Lobosphaera incisa Under Nitrogen and Phosphorous Deprivation. Front. Plant Sci. 2020, 11, 614846. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Ha, U.S.; Yu, K.; Wu, C.; Yokoyama, N.; Zi, X. Kavalactone yangonin induces autophagy and sensitizes bladder cancer cells to flavokawain A and docetaxel via inhibition of the mTOR pathway. J. Biomed. Res. 2017, 31, 408–418. [Google Scholar]
- Tang, X.; Amar, S. Kavain Involvement in LPS-Induced Signaling Pathways. J. Cell. Biochem. 2016, 117, 2272–2280. [Google Scholar] [CrossRef]
- Reid, R.C. Inhibitors of Secretory Phospholipase A2 Group IIA. Curr. Med. Chem. 2005, 12, 3011–3026. [Google Scholar] [CrossRef] [PubMed]
- Ai, S.; Lin, G.; Bai, Y.; Liu, X.; Piao, L. QSAR Classification-Based Virtual Screening Followed by Molecular Docking Identification of Potential COX-2 Inhibitors in a Natural Product Library. J. Comput. Biol. 2019, 26, 1296–1315. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.; Song, L.; Xie, J.; Wu, X.-R.; Gin, G.E.; Wang, B.; Uchio, E.; Zi, X. Kavalactone Kawain Impedes Urothelial Tumorigenesis in UPII-Mutant Ha-Ras Mice via Inhibition of mTOR Signaling and Alteration of Cancer Metabolism. Molecules 2023, 28, 1666. https://doi.org/10.3390/molecules28041666
Liu Z, Song L, Xie J, Wu X-R, Gin GE, Wang B, Uchio E, Zi X. Kavalactone Kawain Impedes Urothelial Tumorigenesis in UPII-Mutant Ha-Ras Mice via Inhibition of mTOR Signaling and Alteration of Cancer Metabolism. Molecules. 2023; 28(4):1666. https://doi.org/10.3390/molecules28041666
Chicago/Turabian StyleLiu, Zhongbo, Liankun Song, Jun Xie, Xue-Ru Wu, Greg E. Gin, Beverly Wang, Edward Uchio, and Xiaolin Zi. 2023. "Kavalactone Kawain Impedes Urothelial Tumorigenesis in UPII-Mutant Ha-Ras Mice via Inhibition of mTOR Signaling and Alteration of Cancer Metabolism" Molecules 28, no. 4: 1666. https://doi.org/10.3390/molecules28041666