
Schisandra henryi—A Rare Species with High Medicinal Potential


Abstract
:1. Introduction
2. Methodology
3. Morphology and Natural Habitats
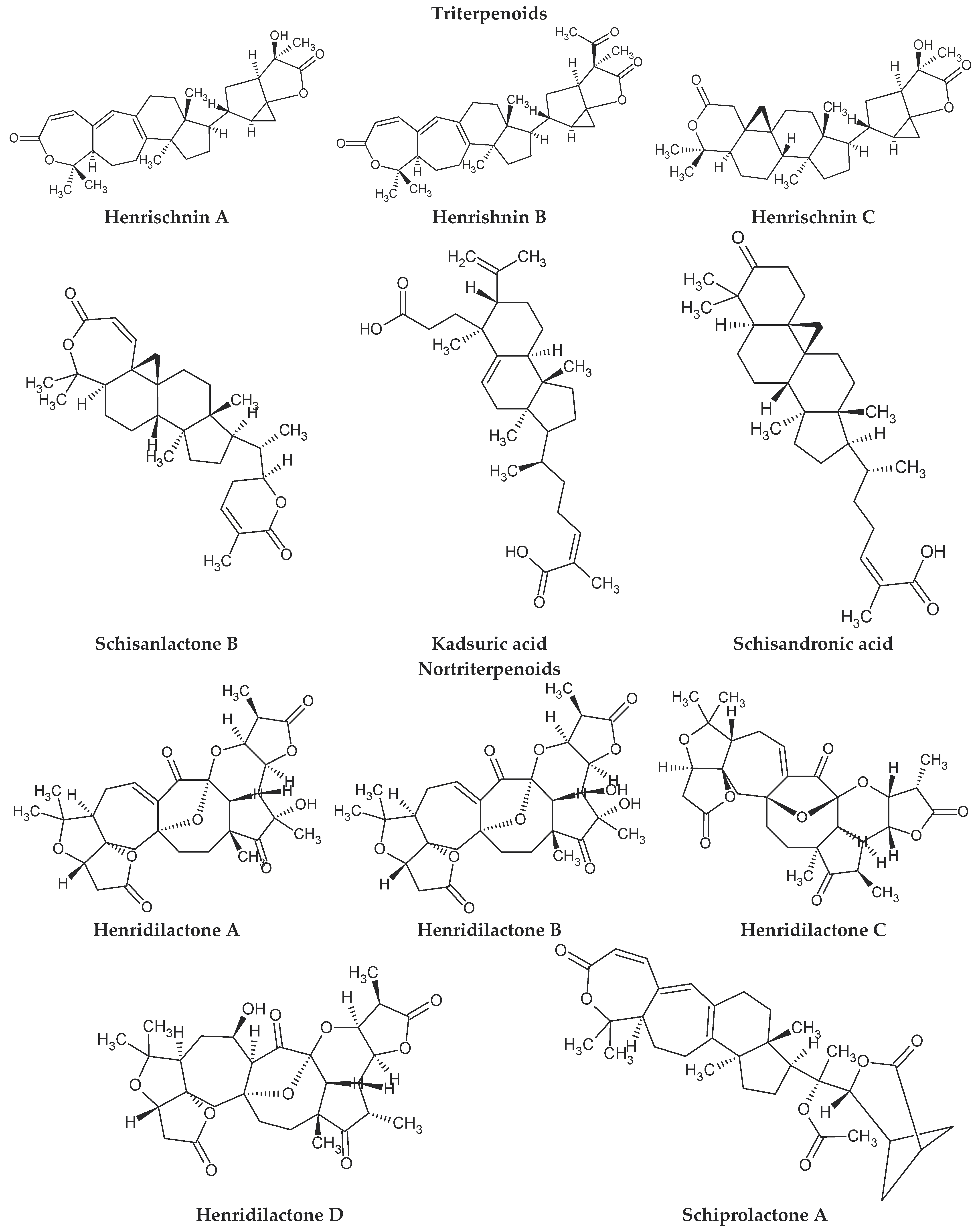
4. Chemical Composition
5. Reports on the Biological Activities
6. Biological Activity of Chosen Dibenzocyclooctadiene Lignans
6.1. Antioxidant Activity
6.2. Anti-Inflammatory Activity
6.3. Anticancer Activity
6.4. Antiviral Activity
6.5. Neuroprotective Activity
6.6. Hepatoprotective and Hepatoregenerative Activity
6.7. Cardioprotective Activity
6.8. Supportive Activity in the Treatment of Intestinal Dysfunction
6.9. Anti-Osteoporotic Activity
7. Plant Biotechnology Research
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Saunders, R.M.K. Monograph of Schisandra (Schisandraceae). Am. Soc. Plant Taxon. 2000, 58, 1–146. [Google Scholar] [CrossRef]
- Saunders, R.M.K. Species Plantarum Flora of the World Part 4. Schisandraceae. In Australian Biological Resources Study; Australian Biological Resources Study: Canberra, Australia, 2001; pp. 1–68. [Google Scholar]
- Chinese Pharmacopoeia Commission. Pharmacopoeia of the People’s Republic of China; China Chemical Industry Press: Beijing, China, 2005. [Google Scholar]
- Committee of the Japanese Pharmacopoeia Evaluation and Licensing Division Pharmaceuticals and Food Safety. Japanese Pharmacopoeia. Labour and Welfare; Bureau Ministry of Health: Tokyo, Japan, 2006. [Google Scholar]
- Central Pharmaceutical Affairs Council of Korea. Korean Pharmacopoeia; Central Pharmaceutical Affairs Council of Korea: Seoul, Korea, 2002. [Google Scholar]
- European Directorate for the Quality of Medicine. European Pharmacopoeia 10.0; European Directorate for the Quality of Medicine: Strasbourg, France, 2010. [Google Scholar]
- Upton, R.; Graff, A.; Jolliffe, G.; Länger, R.; Williamson, E. American Herbal Pharmacopoeia: Botanical Pharmacognosy—Microscopic Characterization of Botanical Medicines; CRC Press: Boca Raton, FL, USA, 2011; ISBN 1420073281. [Google Scholar]
- Shikov, A.N.; Pozharitskaya, O.N.; Makarov, V.G.; Wagner, H.; Verpoorte, R.; Heinrich, M. Medicinal Plants of the Russian Pharmacopoeia; Their History and Applications. J. Ethnopharmacol. 2014, 154, 481–536. [Google Scholar] [CrossRef] [PubMed]
- WHO Monographs on Selected Medicinal Plants. Volume 2; Department of Essential Drugs and Medicines Policy—World Health Organization: Geneva, Switzerland, 2004.
- Kam Ming, K.; Jun, Y.; Chuixin, Q. Schisandra Chinensis: An Herb of North Eastern China Origin; World Scientific Publishing: Singapore, 2015. [Google Scholar]
- Tian, Z. Characterization and Genetic Analysis of the Complete Chloroplast Genome of Schisandra chinensis (Magnoliaceae: Schisandra), an Herbal Medicine from China. Mitochondrial DNA B Resour. 2019, 4, 2428–2430. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.L.; Liu, B.Y.; Ma, K.W. Traditional Chinese Medicine. Lancet 2008, 372, 1938–1940. [Google Scholar] [CrossRef]
- Jin, J.; Bi, H.; Hu, J.; Zeng, H.; Zhong, G.; Zhao, L.; Huang, Z.; Huang, M. Effect of Wuzhi Tablet (Schisandra sphenanthera Extract) on the Pharmacokinetics of Paclitaxel in Rats. Phytother. Res. 2011, 25, 1250–1253. [Google Scholar] [CrossRef]
- Xue, X.P.; Qin, X.L.; Xu, C.; Zhong, G.P.; Wang, Y.; Huang, M.; Bi, H.C. Effect of Wuzhi Tablet (Schisandra sphenanthera Extract) on the Pharmacokinetics of Cyclosporin A in Rats. Phytother. Res. 2013, 27, 1255–1259. [Google Scholar] [CrossRef]
- Bi, H.; Li, F.; Krausz, K.W.; Qu, A.; Johnson, C.H.; Gonzalez, F.J. Targeted Metabolomics of Serum Acylcarnitines Evaluates Hepatoprotective Effect of Wuzhi Tablet (Schisandra sphenanthera Extract) against Acute Acetaminophen Toxicity. Evid. Based Complement. Altern. Med. 2013, 2013, 985257. [Google Scholar] [CrossRef]
- Lu, Y.; Chen, D.F. Analysis of Schisandra chinensis and Schisandra sphenanthera. J. Chromatogr. A 2009, 1216, 1980–1990. [Google Scholar] [CrossRef]
- Szopa, A.; Ekiert, H. Lignans in Schisandra chinensis in Vitro Cultures. Pharmazie 2011, 66, 633–634. [Google Scholar]
- Jiang, P.; Lu, Y.; Chen, D. Authentication of Schisandra chinensis and Schisandra sphenanthera in Chinese Patent Medicines. J. Pharm. Biomed. Anal. 2016, 131, 263–271. [Google Scholar] [CrossRef]
- Szopa, A.; Barnaś, M.; Ekiert, H. Phytochemical Studies and Biological Activity of Three Chinese Schisandra Species (Schisandra sphenanthera, Schisandra henryi and Schisandra rubriflora): Current Findings and Future Applications. Phytochem. Rev. 2019, 18, 109–128. [Google Scholar] [CrossRef]
- Szopa, A.; Dziurka, M.; Granica, S.; Klimek-Szczykutowicz, M.; Kubica, P.; Warzecha, A.; Jafernik, K.; Ekiert, H. Schisandra Rubriflora Plant Material and in Vitro Microshoot Cultures as Rich Sources of Natural Phenolic Antioxidants. Antioxidants 2020, 9, 488. [Google Scholar] [CrossRef]
- Xiao, W.L.; Gong, Y.Q.; Wang, R.R.; Weng, Z.Y.; Luo, X.; Li, X.N.; Yang, G.Y.; He, F.; Pu, J.X.; Yang, L.M.; et al. Bioactive Nortriterpenoids from Schisandra grandiflora. J. Nat. Prod. 2009, 72, 1678–1681. [Google Scholar] [CrossRef]
- Poornima, B.; Kumar, D.A.; Siva, B.; Venkanna, A.; Vadaparthi, P.R.R.; Kumar, K.; Tiwari, A.K.; Babu, K.S. Advanced Glycation End-Products Inhibitors Isolated from Schisandra grandiflora. Nat. Prod. Res. 2016, 30, 493–496. [Google Scholar] [CrossRef]
- Shi, W.; Liu, H.W.; Guo, X.; Hou, L.; Gao, J.M. Triterpenoids from the Stems of Schisandra grandiflora and Their Biological Activity. J. Asian Nat. Prod. Res. 2016, 18, 711–718. [Google Scholar] [CrossRef]
- Poornima, B.; Siva, B.; Shankaraiah, G.; Venkanna, A.; Nayak, V.L.; Ramakrishna, S.; Venkat Rao, C.; Babu, K.S. Novel Sesquiterpenes from Schisandra grandiflora: Isolation, Cytotoxic Activity and Synthesis of Their Triazole Derivatives Using “Clickg” Reaction. Eur. J. Med. Chem. 2015, 92, 449–458. [Google Scholar] [CrossRef]
- Lei, C.; Huang, S.X.; Chen, J.J.; Yang, L.B.; Xiao, W.L.; Chang, Y.; Lu, Y.; Huang, H.; Pu, J.X.; Sun, H.D. Propindilactones E-J, Schiartane Nortriterpenoids from Schisandra propinqua Var. propinqua. J. Nat. Prod. 2008, 71, 1228–1232. [Google Scholar] [CrossRef]
- Lei, C.; Huang, S.X.; Chen, J.J.; Pu, J.X.; Yang, L.B.; Zhao, Y.; Liu, J.P.; Gao, X.M.; Xiao, W.L.; Sun, H.D. Lignans from Schisandra propinqua Var. propinqua. Chem. Pharm. Bull. 2007, 55, 1281–1283. [Google Scholar] [CrossRef]
- Ding, W.P.; Hu, K.; Liu, M.; Li, X.R.; Chen, R.; Li, X.N.; Du, X.; Puno, P.T.; Sun, H.D. Five New Schinortriterpenoids from Schisandra propinqua Var. propinqua. Fitoterapia 2018, 127, 193–200. [Google Scholar] [CrossRef]
- Lei, C.; Huang, S.X.; Xiao, W.L.; Ma, Y.B.; Chen, J.J.; Pu, J.X.; Sun, H.D. Bisnortriterpenoids Possessing an 18-Nor-Schiartane Skeleton from Schisandra propinqua Var. propinqua. Planta Med. 2010, 76, 1611–1615. [Google Scholar] [CrossRef]
- Liu, J.S.; Huang, M.F.; Ayer, W.A.; Nakashima, T.T. Structure of Enshicine from Schisandra henryi. Phytochemistry 1984, 23, 1143–1145. [Google Scholar] [CrossRef]
- Iu, H.L.; Li-jia, X.U.; Eng, Y.P.; Ang, X.Y.; Iao, P.X. Two New Lignans from Schisandra henryi. Chem. Pharm. Bull. 2009, 57, 405–407. [Google Scholar] [CrossRef]
- Schisandraceae. Flora China 2008, 7, 39–43.
- Jafernik, K.; Szopa, A.; Barnaś, M.; Dziurka, M.; Ekiert, H. Schisandra henryi C. B. Clarke in Vitro Cultures: A Promising Tool for the Production of Lignans and Phenolic Compounds. Plant Cell Tissue Organ Cult. 2020, 143, 45–60. [Google Scholar] [CrossRef]
- Yuan, L.C.; Luo, Y.B.; Thien, L.B.; Fan, J.H.; Xu, H.L.; Chen, Z.D. Pollination of Schisandra henryi (Schisandraceae) by Female, Pollen-Eating Megommata Species (Cecidomyiidae, Diptera) in South-Central China. Ann. Bot. 2007, 99, 451–460. [Google Scholar] [CrossRef]
- Lian-Niang, L.; Hong, X. Henricine, a New Tetrahydrofuran Lignan from Schisandra henryi. Planta Med. 1986, 52, 493–494. [Google Scholar] [CrossRef]
- Umezawa, T. Diversity in Lignan Biosynthesis. Phytochem. Rev. 2003, 2, 371–390. [Google Scholar] [CrossRef]
- Kochetkov, N.; Khorlin, A.; Chizhov, O.; Al, E. Schizandrin—Lignan of Unusual Structure. Tetrahedron Lett. 1961, 2, 730–734. [Google Scholar] [CrossRef]
- Whiting, D.A. Lignans, Neolignans, and Related Compounds. Nat. Prod. Rep. 1990, 7, 349–364. [Google Scholar] [CrossRef]
- Liu, G. Bicyclol: A Novel Drug for Treating Chronic Viral Hepatitis B and C. Med. Chem. 2009, 5, 29–43. [Google Scholar] [CrossRef]
- Wang, Y.; Nie, H.; Zhao, X.; Qin, Y.; Gong, X. Bicyclol Induces Cell Cycle Arrest and Autophagy in HepG2 Human Hepatocellular Carcinoma Cells through the PI3K/AKT and Ras/Raf/MEK/ERK Pathways. BMC Cancer 2016, 16, 742. [Google Scholar] [CrossRef] [PubMed]
- Yi, G.; Dong, Y.; Zongile, Z. Bicyclol Medical Composition and Preparation Method Thereof. CN103242286A, 14 August 2013. [Google Scholar]
- Hu, W.; Li, Y.; Zhang, C.Z. Enantioseparation of Racemic Anti-Hepatitis New Drug Bicyclol with Crystallization. Chin. Chem. Lett. 2005, 16, 1471–1473. [Google Scholar]
- Yang, X.Y.; Zhuo, Q.; Wu, T.X.; Liu, G.J. Bicyclol for Chronic Hepatitis C. Cochrane Database Syst. Rev. 2007, 1. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Yu, L.; Wei, H.; Liu, G. A Novel Antihepatitis Drug, Bicyclol, Prevents Liver Carcinogenesis in Diethylnitrosamine-Initiated and Phenobarbital-Promoted Mice Tumor Model. J. Biomed. Biotechnol. 2012. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.N.; Chen, H.; Li, Y. Effect of Bicyclol on Cisplatin-Induced Hepatotoxicity in the Hepatocarcinoma 22 Tumour-Bearing Mice. Basic Clin. Pharmacol. Toxicol. 2009, 104, 300–305. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-S.; Tao, Y.; Huang, M.-F. Studies on the Constituents of Schisandra Henryi. V. The Structures of Wulignan A1, A2, Epiwulignan A1 and Epischisandrone. J. Chin. Chem. Soc. 1988, 46, 483–488. [Google Scholar]
- Yue, J.-m.; Chen, Y.-z.; Huang, S.-m.; Cheng, J.-l.; Cui, Y.-x. Ganschisandrine, a Lignan from Schisandra sphenanthera. Phytochemistry 1989, 28, 1774–1776. [Google Scholar] [CrossRef]
- Yamada, Y.; Hsu, C.; Iguchi, K.; Suzuki, S.; Hsu, H.Y.; Chen, Y.P. Structure of kadsuric acid. A new seco-triterpenoid from Kadsura japonica Dunal. Chem. Lett. 1976, 5, 1307–1310. [Google Scholar] [CrossRef]
- Xue, Y.B.; Yang, J.H.; Li, X.N.; Du, X.; Pu, J.X.; Xiao, W.L.; Su, J.; Zhao, W.; Li, Y.; Sun, H.D. Henrischinins A–C: Three New Triterpenoids from Schisandra henryi. Org. Lett. 2011, 13, 1564–1567. [Google Scholar] [CrossRef]
- Li, R.; Shen, Y.; Xiang, W.; Sun, H. Four Novel Nortriterpenoids Isolated from Schisandra henryi Var. yunnanensis. Eur. J. Org. Chem. 2004, 2004, 807–811. [Google Scholar] [CrossRef]
- He, T.B.; Yan, B.C.; Hu, K.; Li, X.N.; Sun, H.D.; Puno, P.T. Neuroprotective Schinortriterpenoids with Diverse Scaffolds from Schisandra henryi. Bioorg. Chem. 2020, 105, 104353. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-S.; Huang, M.-F.; Gao, Y.-L. Studies on the Constituents of Schisandra henryi Clarke. H. The Structures of Schisanhenrin and Schisanhenric Acid. Acta Chim. Sin. 1980, 38, 363–370. [Google Scholar]
- Chen, Y.G.; Wu, Z.C.; Lv, Y.P.; Gui, S.H.; Wen, J.; Liao, X.R.; Yuan, L.M.; Halaweish, F. Triterpenoids from Schisandra henryi with Cytotoxic Effect on Leukemia and Hela Cells in Vitro. Arch. Pharm. Res. 2003, 26, 912–916. [Google Scholar] [CrossRef] [PubMed]
- Xiao, W.-L.; Wang, R.-R.; Zhao, W.; Tian, R.-R.; Shang, S.-Z.; Yang, L.-M.; Yang, J.-H.; Pu, J.-X.; Zheng, Y.-T.; Sun, H.-D. Anti-HIV-1 activity of lignans from the fruits of Schisandra rubriflora. Arch. Pharm. Res. 2010, 33, 697–701. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Kim, J.; Jeong, K.; Han, S.; Jeong, T.; Yum, M. Antioxidant Activity and Inhibition of MMP-1 Expression of Schizandrae Fructus (Schizandra Chinensis) Extract. Korean J. Pharmacogn. 2013, 1, 47–52. [Google Scholar]
- Yan, Z.; Guo, J.; Tian, F.; Mao, X.; Li, Y.; Li, C. Active Compounds from Schisandra chinensis Exhibiting Tyrosinase Activity and Melanin Content Inhibition in B16 Melanoma Cells. Biotechnol. Bioprocess Eng. 2015, 4, 814–823. [Google Scholar] [CrossRef]
- Chiu, P.Y.; Lam, P.Y.; Yan, C.W.; Ko, K.M. Schisandrin B Protects against Solar Irradiation-Induced Oxidative Injury in BJ Human Fibroblasts. Fitoterapia 2011, 82, 682–691. [Google Scholar] [CrossRef]
- Oh, S.Y.; Kim, Y.H.; Bae, D.S.; Um, B.H.; Pan, C.H.; Kim, C.Y.; Lee, H.J.; Lee, J.K. Anti-Inflammatory Effects of Gomisin N, Gomisin J, and Schisandrin C Isolated from the Fruit of Schisandra chinensis. Biosci. Biotechnol. Biochem. 2010, 74, 285–291. [Google Scholar] [CrossRef]
- Casarin, E.; Dall’Acqua, S.; Šmejkal, K.; Šlapetová, T.; Innocenti, G.; Carrara, M. Molecular Mechanisms of Antiproliferative Effects Induced by Schisandra-Derived Dibenzocyclooctadiene Lignans (+)-Deoxyschisandrin and (-)-Gomisin N in Human Tumour Cell Lines. Fitoterapia 2014, 98, 241–247. [Google Scholar] [CrossRef]
- Wang, Z.; Yu, K.; Hu, Y.; Su, F.; Gao, Z.; Hu, T.; Yang, Y.; Cao, X.; Qian, F. Schisantherin A Induces Cell Apoptosis through ROS/JNK Signaling Pathway in Human Gastric Cancer Cells. Biochem. Pharmacol. 2020, 173, 113673. [Google Scholar] [CrossRef]
- Sa, F.; Zhang, L.Q.; Chong, C.M.; Guo, B.J.; Li, S.; Zhang, Z.J.; Zheng, Y.; Hoi, P.M.; Lee, S.S.S.M.Y. Discovery of Novel Anti-Parkinsonian Effect of Schisantherin A in in Vitro and in Vivo. Neurosci. Lett. 2015, 593, 7–12. [Google Scholar] [CrossRef]
- Caichompoo, W.; Zhang, Q.-Y.; Hou, T.-T.; Gao, H.-J.; Qin, L.-P.; Zhou, X.-J. Optimization of Extraction and Purification of Active Fractions from Schisandra chinensis (Turcz.) and Its Osteoblastic Proliferation Stimulating Activity. Phytother. Res. 2009, 23, 289–292. [Google Scholar] [CrossRef]
- Lam, P.; Yan, C.; Chiu, P.; Leung, H.; Ko, K. Schisandrin B Protects against Solar Irradiation-Induced Oxidative Stress in Rat Skin Tissue. Fitoterapia 2011, 82, 393–400. [Google Scholar] [CrossRef]
- Xu, L.; Grandi, N.; Del Vecchio, C.; Al, E. From the Traditional Chinese Medicine Plant Schisandra chinensis New Scaffolds Effective on HIV-1 Reverse Transcriptase Resistant to Non-Nucleoside Inhibitors. J. Microbiol. 2015, 53, 288–293. [Google Scholar] [CrossRef]
- Chang, R.; Li, Y.; Yang, X.; Yue, Y.; Dou, L.; Wang, Y.; Zhang, W.; Li, X. Protective Role of Deoxyschizandrin and Schisantherin A against Myocardial Ischemia-Reperfusion Injury in Rats. PLoS ONE 2013, 8, 2–11. [Google Scholar] [CrossRef]
- Xu, Z.; Zhang, M.; Dou, D.; Kang, T.; Li, F. Effects of Deoxyschisandrin on Visceral Sensitivity of Mice with Inflammatory Bowel Disease. Evid. -Based Complement. Altern. Med. 2019, 29, 86–97. [Google Scholar] [CrossRef]
- Ci, X.; Ren, R.; UXu, K.; Li, H.; Yu, Q.; Song, Y.; Wang, D.; Li, R.; Deng, X. Schisantherin A Exhibits Anti-Inflammatory Properties by down-Regulating NF-KappaB and MAPK Signaling Pathways in Lipopolysaccharide-Treated RAW 264.7 Cells. Inflammation 2010, 33, 126–136. [Google Scholar] [CrossRef]
- Liao, S.; Zhou, K.; Li, D.; Xie, X.; Jun, F.; Wang, J. Schisantherin A Suppresses Interleukin-1β-Induced Inflammation in Human Chondrocytes via Inhibition of NF-ΚB and MAPKs Activation. Eur. J. Pharmacol. 2016, 780, 65–70. [Google Scholar] [CrossRef]
- Li, C.; Chen, T.; Zhou, H.; Zhang, C.; Feng, Y.; Tang, F.; Hoi, M.P.M.; He, C.; Zheng, Y.; Lee, S.M.Y. Schisantherin a Attenuates Neuroinflammation in Activated Microglia: Role of Nrf2 Activation through ERK Phosphorylation. Cell. Physiol. Biochem. 2018, 47, 1769–1784. [Google Scholar] [CrossRef]
- Zhou, E.; Li, Y.; Wei, Z.; Fu, Y.; Lei, H.; Zhang, N.; Yang, Z.; Xie, G. Schisantherin A Protects Lipopolysaccharide-Induced Acute Respiratory Distress Syndrome in Mice through Inhibiting NF-??B and MAPKs Signaling Pathways. Int. Immunopharmacol. 2014, 22, 133–140. [Google Scholar] [CrossRef]
- Chen, Y.G.; Wu, Z.C.; Gui, S.H.; Lv, Y.P.; Liao, X.R.; Halaweish, F. Lignans from Schisandra hernyi with DNA Cleaving Activity and Cytotoxic Effect on Leukemia and Hela Cells in Vitro. Fitoterapia 2005, 76, 370–373. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhao, X.; Xu, X.; Mao, X.; Zhi, L.; Li, H.; Guo, L.; Kaishun, B.; Jia, Y. Schisantherin A Recovers Aβ-Induced Neurodegeneration with Cognitive Decline in Mice. Physiol. Behav. 2014, 132, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.Q.; Sa, F.; Chong, C.M.; Ying, W.; Zhou, Y.Z.; Chang, R.C.C.; Chan, S.W.; Hoi, P.M.; Lee, S.M.Y. Schisantherin A Protects against 6-OHDA-Induced Dopaminergic Neuron Damage in Zebrafish and Cytotoxicity in SH-SY5Y Cells through the ROS/NO and AKT/GSK3β Pathways. J. Ethnopharmacol. 2015, 170, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Zheng, N.; Liu, F.; Lu, H.; Zhan, Y.; Zhang, M.; Guo, W.; Ding, G. Schisantherin A Protects against Liver Ischemia-Reperfusion Injury via Inhibition of Mitogen-Activated Protein Kinase Pathway. Int. Immunopharmacol. 2017, 47, 28–37. [Google Scholar] [CrossRef]
- Wang, H.; Che, J.; Cui, K.; Zhuang, W.; Li, H.; Sun, J.; Chen, J.; Wang, C. Schisantherin A Ameliorates Liver Fibrosis through TGF-Β1mediated Activation of TAK1/MAPK and NF-ΚB Pathways in Vitro and in Vivo. Phytomedicine 2021, 88. [Google Scholar] [CrossRef]
- Kopustinskiene, D.M.; Bernatoniene, J. Antioxidant Effects of Schisandra chinensis Fruits and Their Active Constituents. Antioxidants 2021, 10, 620. [Google Scholar] [CrossRef]
- Lu, H.; Liu, G.T. Anti-Oxidant Activity of Dibenzocyclooctene Lignans Isolated from Schisandraceae. Planta Med. 1992, 58, 311–313. [Google Scholar] [CrossRef]
- Maharjan, S.; Park, B.K.; Lee, S.I.; Lim, Y.; Lee, K.; Kwon, H.J. Gomisin G Inhibits the Growth of Triple-Negative Breast Cancer Cells by Suppressing AKT Phosphorylation and Decreasing Cyclin D1. Biomol. Ther. 2018, 26, 322–327, Erratum in Biomol. Ther. 2018, 26, 520. [Google Scholar] [CrossRef]
- Maharjan, S.; Park, B.K.; Lee, S.I.; Lim, Y.; Lee, K.; Lee, Y.; Kwon, H.J. Gomisin g Suppresses the Growth of Colon Cancer Cells by Attenuation of Akt Phosphorylation and Arrest of Cell Cycle Progression. Biomol. Ther. 2019, 27, 210–215. [Google Scholar] [CrossRef]
- Chen, D.; Zhang, S.; Xie, L.J.X.; Xie, J.; Chen, K.; Kashiwada, Y.; Zhou, B.; Wang, P.; Cosentino, L.; Lee, K. Anti-AIDS Agents—XXVI. Structure-Activity Correlations of Gomisin-G-Related Anti-HIV Lignans from Kadsura Interior and of Related Synthetic Analogues. Bioorg. Med. Chem. 1997, 5, 1715–1723. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound Extracted from Raw Material | Extraction Condition | Analysis Method | References |
|---|---|---|---|
| Enshicine from fruit |
| column chromatography on silica gel (gradient mode: benzene, benzene–ethyl acetate (10:1), benzene–ethyl acetate (4:1), and ethyl acetate) | [29] |
| Triterpenoids, lignans from leaves |
| UHPLC-MS/MS with triple quadrupole mass filter (QQQ) (analytical column: Kinetex C18 150 × 4.6 mm, 2.6 µm, gradient mode: 50% methanol in water (A), 100% methanol (B) with 1% formic acid) | [32] |
| Phenolic acids and flavonoids from leaves |
| HPLC-DAD (analytical column: Purospher RP-18, mobile phase: methanol and 0.5% acetic acid (A), methanol (B)) | [32] |
| Triterpenoids from leaves and stems |
| column chromatography on silica gel, (chloroform–acetone (1:0 to 0:1), semi-preparative HPLC (analytical column: Agilent 1100 HPLC; Zorbax SB-C-18, Agilent, 9.4 mm 25 cm, gradient mode: methanol–water (65:35) | [48] |
| Nortriterpenoids form stems and leaves |
| RP-HPLC (55% methanol/water) column chromatography on silica gel (gradient systems: chloroform-Me2CO 1:0 0:1), repeated column chromatography (silica gel, petroleum ether/Me2CO, 9:1 and petroleum ether/ethyl acetate 4:1), RP-HPLC (gradient mode: 55% methanol/water) | [49] |
| Triterpenoids from stems |
| column chromatography on silica gel (petroleum ether–ethyl acetate 4:1), repeated column chromatography on silica gel | [52] |
| Schinortriterpenoids from stems and leaves |
| column chromatography with silica gel (chloroform/acetone 1:0, 9:1, 7:3, 3:2, 1:1 and 0:1), semi-preparative HPLC (analytical column; RP-18, Sephadex LH-20-methanol/water) | [50] |
| Lignan | Chemical Structure of Compound | Maximal Content [mg/100 g DM ± SD] | Action | Mode of Action | Reference | |
|---|---|---|---|---|---|---|
| Microshoot Cultures | Leaves of the Parent Plant | |||||
| Schisandrin (schizandrin, schizandrol A, schisandrol A) |  | 61.24 ± 0.23 | 8.62 ± 0.95 | Antioxidant |
| [54] |
| Anti-osteoporotic |
| [60] | ||||
| Schisandrin C (wuweizisu C) |  | 28.61 ± 0.23 | 1.06 ± 0.38 | Antioxidant |
| [57,62] |
| Anti-inflammatory |
| [57] | ||||
| Deoxyschisandrin (schisandrin A, schizandrin A, deoxyschizandrin) |  | 3.63 ± 0.27 | 1.70 ± 0.55 | Anticancer |
| [54,57,60,63] |
| Antiviral |
| [63,64] | ||||
| Cardioprotective |
| |||||
| Supportive treatment in intestinal dysfunction |
| [65] | ||||
| Anti-osteoporotic |
| [61] | ||||
| Schisantherin A (gomisin C, gomisin) |  | 143.74 ± 0.43 | 4.75 ± 0.54 | Anti-inflammatory |
| [66,67,68,69] |
| Anticancer |
| [59,70] | ||||
| Neuroprotective |
| [60,71,72] | ||||
| Hepatoprotective |
| [73,74] | ||||
| Cardioprotective |
| [64] | ||||
| Schisantherin B (Gomisin B, Schisandrer, Wuweizi ester B) |  | 622.59 ± 0.57 | 48.99 ± 4.73 | Antioxidant |
| [75,76] |
| Gomisin G |  | 18.20 ± 0.18 | 1.62 ± 0.51 | Anticancer |
| [70,77,78] |
| Antiviral |
| [79] | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jafernik, K.; Ekiert, H.; Szopa, A. Schisandra henryi—A Rare Species with High Medicinal Potential. Molecules 2023, 28, 4333. https://doi.org/10.3390/molecules28114333
Jafernik K, Ekiert H, Szopa A. Schisandra henryi—A Rare Species with High Medicinal Potential. Molecules. 2023; 28(11):4333. https://doi.org/10.3390/molecules28114333
Chicago/Turabian StyleJafernik, Karolina, Halina Ekiert, and Agnieszka Szopa. 2023. "Schisandra henryi—A Rare Species with High Medicinal Potential" Molecules 28, no. 11: 4333. https://doi.org/10.3390/molecules28114333