

Extended-Synaptotagmin 1 Enhances Liver Cancer Progression Mediated by the Unconventional Secretion of Cytosolic Proteins
 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
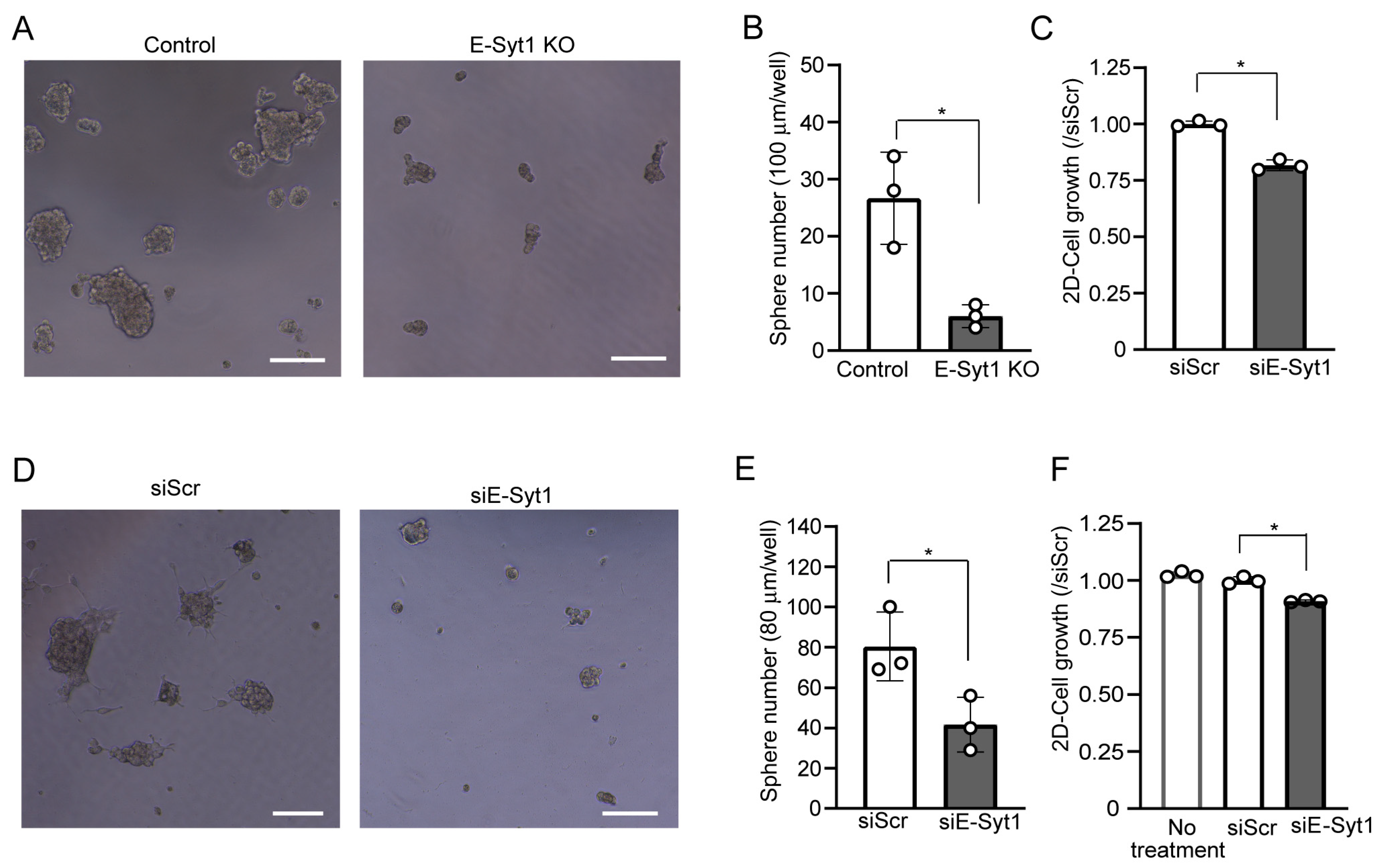
2.1. E-Syt1 Affects Growth of Liver Cancer Cells
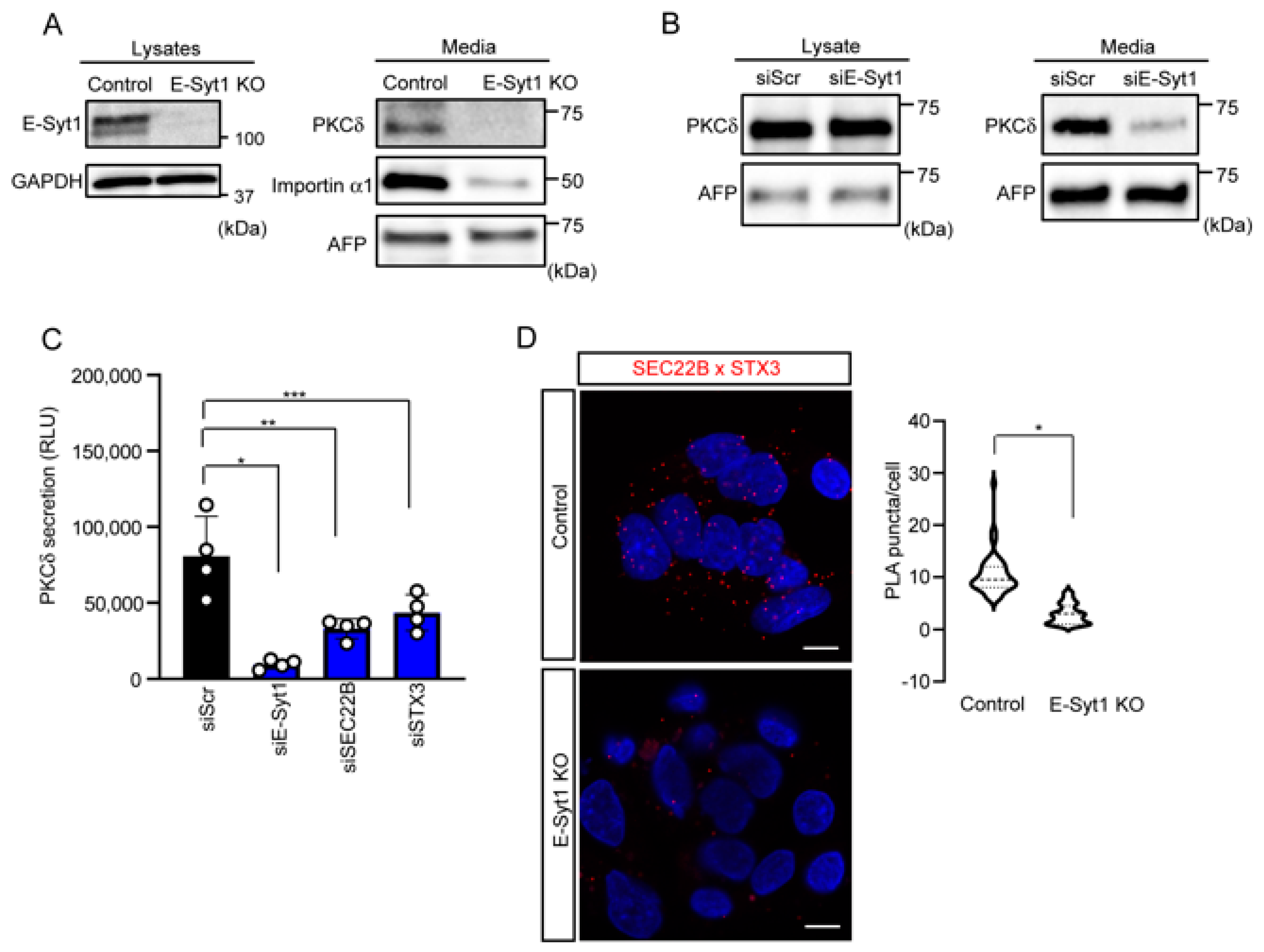
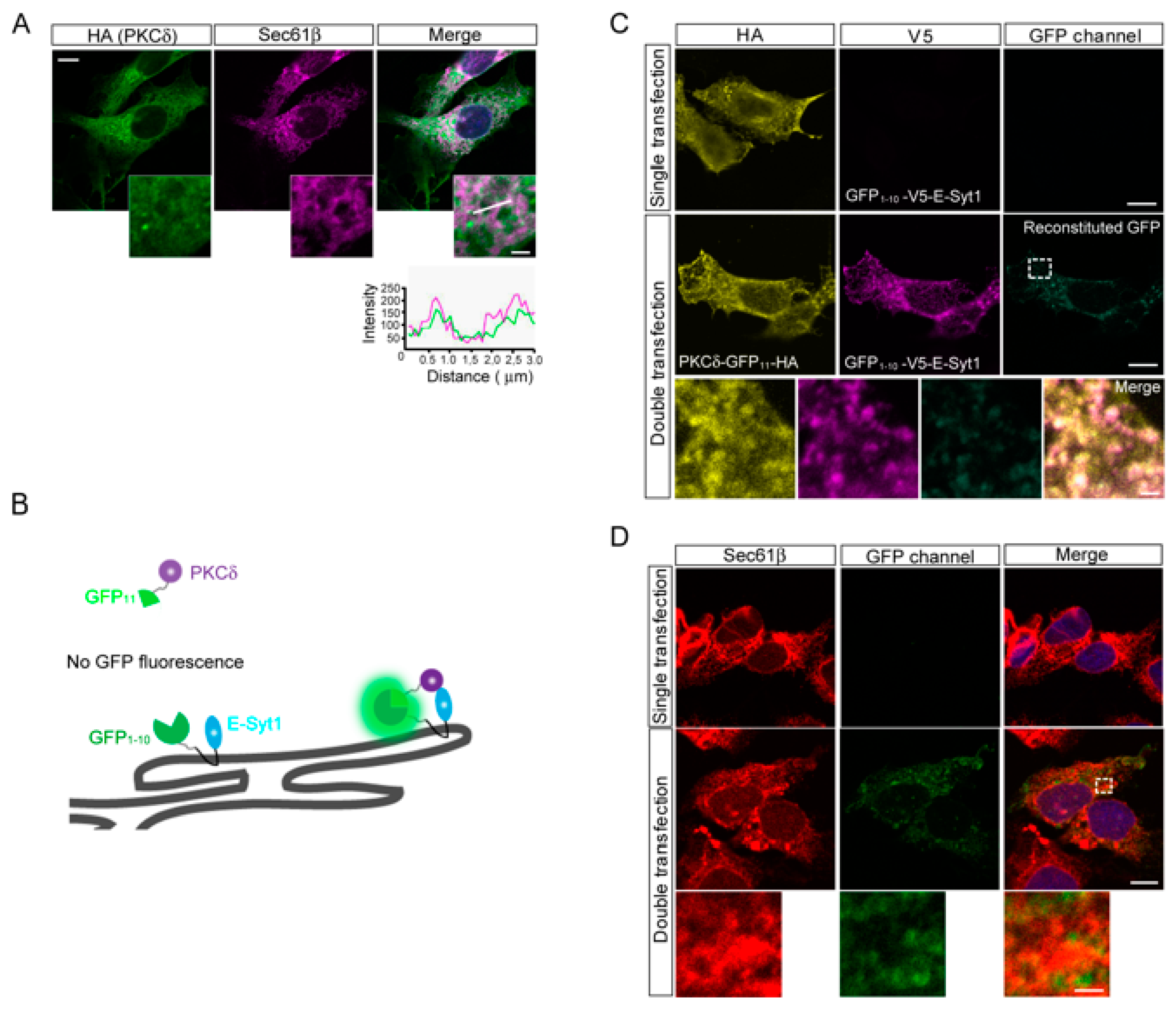
2.2. E-Syt1 Contributes to PKCδ Secretion in Liver Cancer Cell Lines
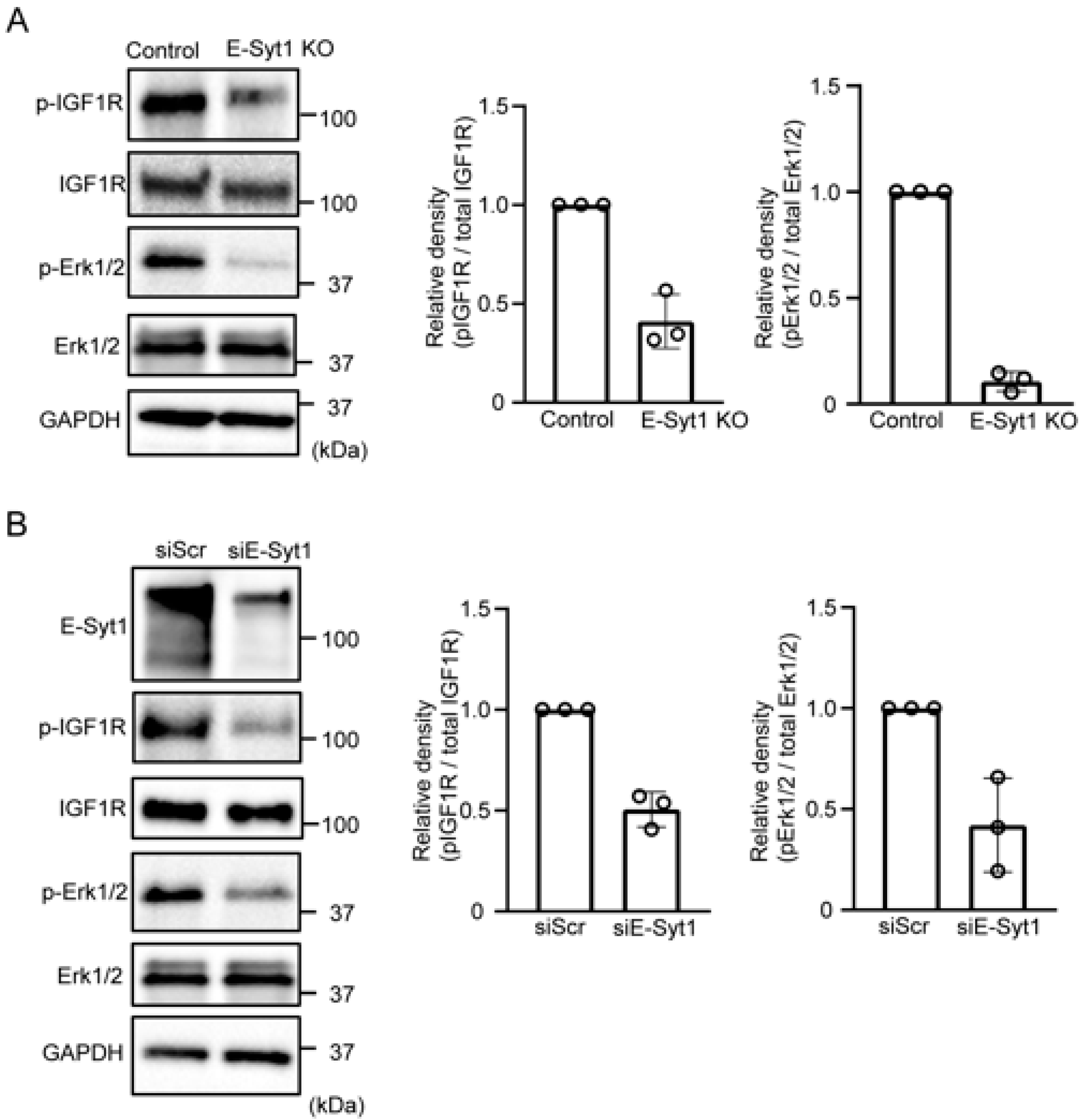
2.3. E-Syt1 Is Involved in the Activation of PKCδ-IGF1R Signaling
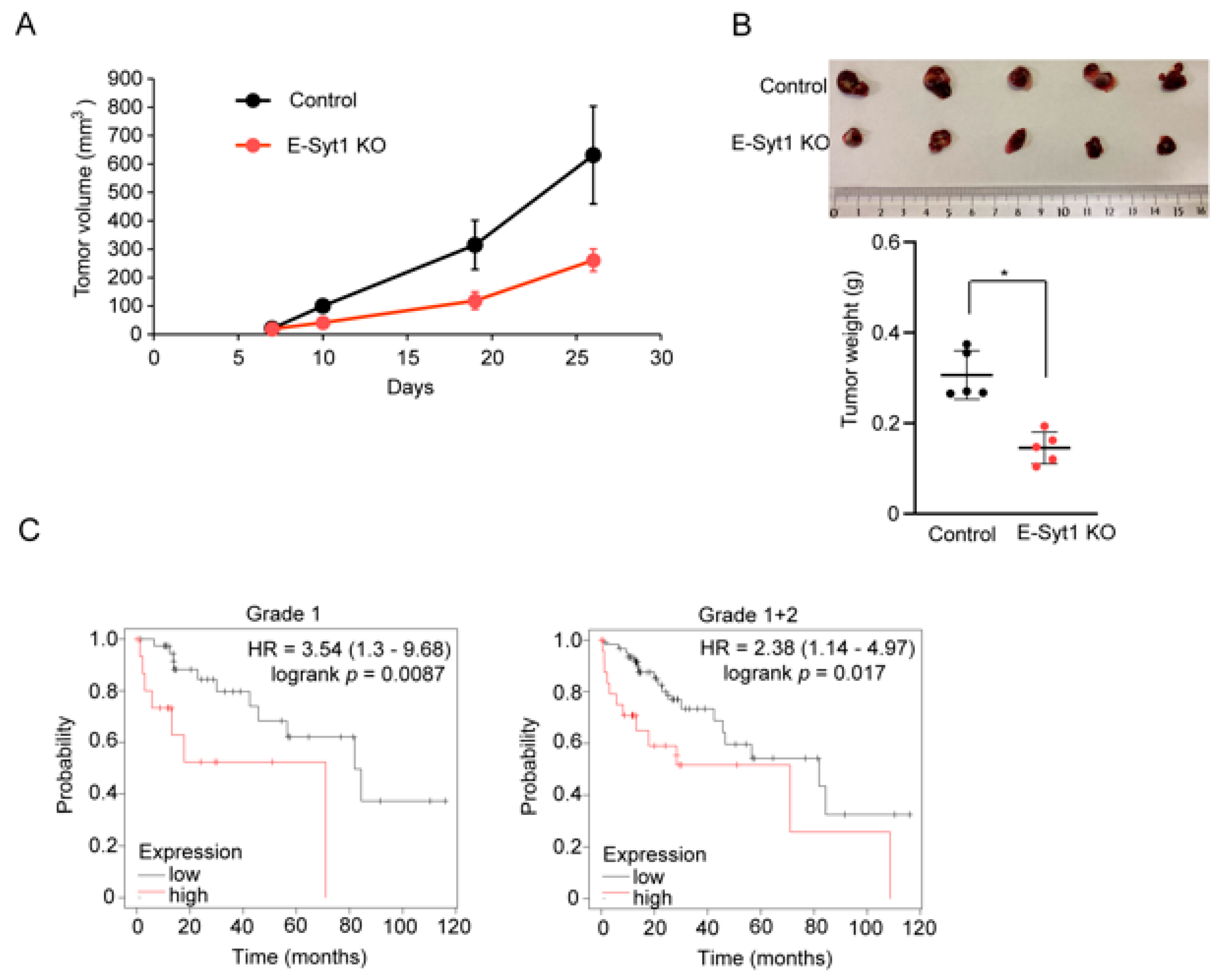
2.4. E-Syt1 Is Involved in Liver Cancer Tumorigenesis
3. Materials and Methods
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Abbreviations
| E-Syt1 | Extended-synaptotagmin 1 |
| PKCδ | protein kinase C delta |
| HCC | hepatocellular carcinoma |
| IGF1R | insulin-like growth factor 1 receptor |
| Erk1/2 | extracellular-signal-related kinase ½ |
References
- Takasugi, M.; Yoshida, Y.; Hara, E.; Ohtani, N. The role of cellular senescense and SASP in tumor microenvironment. FEBS J. 2023, 290, 1348–1361. [Google Scholar] [CrossRef] [PubMed]
- Rabouille, C. Pathways of Unconventional Protein Secretion. Trends Cell Biol. 2017, 27, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; Jia, J.; Kumar, S.; Choi, S.W.; Gu, Y.; Mudd, M.; Dupont, N.; Jiang, S.; Peters, R.; Farzam, F.; et al. Dedicated SNAREs and specialized TRIM cargo receptors mediate secretory autophagy. EMBO J. 2017, 36, 42–60. [Google Scholar] [CrossRef]
- Yamada, K.; Oikawa, T.; Kizawa, R.; Motohashi, S.; Yoshida, S.; Kumamoto, T.; Saeki, C.; Nakagawa, C.; Shimoyama, Y.; Aoki, K.; et al. Unconventional Secretion of PKCδ Exerts Tumorigenic Function via Stimulation of ERK1/2 Signaling in Liver Cancer. Cancer Res. 2021, 81, 414–425. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Kizawa, R.; Yoshida, A.; Koizumi, R.; Motohashi, S.; Shimoyama, Y.; Hannya, Y.; Yoshida, S.; Oikawa, T.; Shimoda, M.; et al. Extracellular PKCδ signals to EGF receptor for tumor proliferation in liver cancer cells. Cancer Sci. 2022, 113, 2378–2385. [Google Scholar] [CrossRef]
- Yamada, K.; Miyamoto, Y.; Tsujii, A.; Moriyama, T.; Ikuno, Y.; Shiromizu, T.; Serada, S.; Fujimoto, M.; Tomonaga, T.; Naka, T.; et al. Cell surface localization of importin α1/KPNA2 affects cancer cell proliferation by regulating FGF1 signalling. Sci. Rep. 2016, 6, 21410. [Google Scholar] [CrossRef]
- Wang, X.; Song, X.; Zhuo, W.; Fu, Y.; Shi, H.; Liang, Y.; Tong, M.; Chang, G.; Luo, Y. The regulatory mechanism of Hsp90alpha secretion and its function in tumor malignancy. Proc. Natl. Acad. Sci. USA 2009, 106, 21288–21293. [Google Scholar] [CrossRef]
- Oikawa, T.; Yamada, K.; Tsubota, A.; Saeki, C.; Tago, N.; Nakagawa, C.; Ueda, K.; Kamioka, H.; Taniai, T.; Haruki, K.; et al. Protein Kinase C Delta Is a Novel Biomarker for Hepatocellular Carcinoma. Gastro Hep Adv. 2023, 2, 83–95. [Google Scholar] [CrossRef]
- Yamada, K.; Motohashi, S.; Oikawa, T.; Tago, N.; Koizumi, R.; Ono, M.; Tachibana, T.; Yoshida, A.; Yoshida, S.; Shimoda, M.; et al. Extended-synaptotagmin 1 engages in unconventional protein secretion mediated via SEC22B(+) vesicle pathway in liver cancer. Proc. Natl. Acad. Sci. USA 2022, 119, e2202730119. [Google Scholar] [CrossRef]
- Chen, Y.A.; Scheller, R.H. SNARE-mediated membrane fusion. Nat. Rev. Mol. Cell Biol. 2001, 2, 98–106. [Google Scholar] [CrossRef]
- New, J.; Thomas, S.M. Autophagy-dependent secretion: Mechanism, factors secreted, and disease implications. Autophagy 2019, 15, 1682–1693. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.; Huang, P.-C.; Chao, H.-M.; Jeng, Y.-M.; Hsu, H.-C.; Pan, H.-W.; Hwu, W.-L.; Lee, Y.-M. Glypican-3 induces oncogenicity by preventing IGF-1R degradation, a process that can be blocked by Grb10. Oncotarget 2017, 8, 80429–80442. [Google Scholar] [CrossRef] [PubMed]
- Manford, A.G.; Stefan, C.J.; Yuan, H.L.; MacGurn, J.A.; Emr, S.D. ER-to-plasma membrane tethering proteins regulate cell signaling and ER morphology. Dev. Cell 2012, 23, 1129–1140. [Google Scholar] [CrossRef]
- Min, S.-W.; Chang, W.-P.; Südhof, T.C. E-Syts, a family of membranous Ca2+-sensor proteins with multiple C2 domains. Proc. Natl. Acad. Sci. USA 2007, 104, 3823–3828. [Google Scholar] [CrossRef] [PubMed]
- Giordano, F.; Saheki, Y.; Idevall-Hagren, O.; Colombo, S.F.; Pirruccello, M.; Milosevic, I.; Gracheva, E.O.; Bagriantsev, S.N.; Borgese, N.; De Camilli, P. PI(4,5)P(2)-dependent and Ca(2+)-regulated ER-PM interactions mediated by the extended synaptotagmins. Cell 2013, 153, 1494–1509. [Google Scholar] [CrossRef]
- Idevall-Hagren, O.; Lü, A.; Xie, B.; De Camilli, P. Triggered Ca2+ influx is required for extended synaptotagmin 1-induced ER-plasma membrane tethering. EMBO J. 2015, 34, 2291–2305. [Google Scholar] [CrossRef]
- Jun, H.J.; Johnson, H.; Bronson, R.T.; de Feraudy, S.; White, F.; Charest, A. The oncogenic lung cancer fusion kinase CD74-ROS activates a novel invasiveness pathway through E-Syt1 phosphorylation. Cancer Res. 2012, 72, 3764–3774. [Google Scholar] [CrossRef]
- Petkovic, M.; Jemaiel, A.; Daste, F.; Specht, C.G.; Izeddin, I.; Vorkel, D.; Verbavatz, J.-M.; Darzacq, X.; Triller, A.; Pfenninger, K.H.; et al. The SNARE Sec22b has a non-fusogenic function in plasma membrane expansion. Nat. Cell Biol. 2014, 16, 434–444. [Google Scholar] [CrossRef]
- Kamiyama, D.; Sekine, S.; Barsi-Rhyne, B.; Hu, J.; Chen, B.; Gilbert, L.A.; Ishikawa, H.; Leonetti, M.D.; Marshall, W.F.; Weissman, J.S.; et al. Versatile protein tagging in cells with split fluorescent protein. Nat. Commun. 2016, 7, 11046. [Google Scholar] [CrossRef]
- Tashiro, S.; Kakimoto, Y.; Shinmyo, M.; Fujimoto, S.; Tamura, Y. Improved Split-GFP Systems for Visualizing Organelle Contact Sites in Yeast and Human Cells. Front. Cell Dev. Biol. 2020, 8, 571388. [Google Scholar] [CrossRef]
- Andrei, C.; Dazzi, C.; Lotti, L.; Torrisi, M.R.; Chimini, G.; Rubartelli, A. The secretory route of the leaderless protein interleukin 1beta involves exocytosis of endolysosome-related vesicles. Mol. Biol. Cell 1999, 10, 1463–1475. [Google Scholar] [CrossRef] [PubMed]
- Steringer, J.P.; Müller, H.-M.; Nickel, W. Unconventional secretion of fibroblast growth factor 2a novel type of protein translocation across membranes? J. Mol. Biol. 2015, 427 Pt A, 1202–1210. [Google Scholar] [CrossRef]
- Ejlerskov, P.; Rasmussen, I.; Nielsen, T.T.; Bergström, A.-L.; Tohyama, Y.; Jensen, P.H.; Vilhardt, F. Tubulin polymerization-promoting protein (TPPP/p25α) promotes unconventional secretion of α-synuclein through exophagy by impairing autophagosome-lysosome fusion. J. Biol. Chem. 2013, 288, 17313–17335. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, T.; Zentgraf, H.; Zehe, C.; Brügger, B.; Bernhagen, J.; Nickel, W. Unconventional secretion of fibroblast growth factor 2 is mediated by direct translocation across the plasma membrane of mammalian cells. J. Biol. Chem. 2004, 279, 6244–6251. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Kenny, S.J.; Ge, L.; Xu, K.; Schekman, R. Translocation of interleukin-1β into a vesicle intermediate in autophagy-mediated secretion. eLife 2015, 4, e11205. [Google Scholar] [CrossRef] [PubMed]
- Duran, J.M.; Anjard, C.; Stefan, C.; Loomis, W.F.; Malhotra, V. Unconventional secretion of Acb1 is mediated by autophagosomes. J. Cell Biol. 2010, 188, 527–536. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamada, K.; Hannya, Y.; Oikawa, T.; Yoshida, A.; Katagiri, K.; Yoshida, S.; Koizumi, R.; Tago, N.; Shimoyama, Y.; Kawamura, A.; et al. Extended-Synaptotagmin 1 Enhances Liver Cancer Progression Mediated by the Unconventional Secretion of Cytosolic Proteins. Molecules 2023, 28, 4033. https://doi.org/10.3390/molecules28104033
Yamada K, Hannya Y, Oikawa T, Yoshida A, Katagiri K, Yoshida S, Koizumi R, Tago N, Shimoyama Y, Kawamura A, et al. Extended-Synaptotagmin 1 Enhances Liver Cancer Progression Mediated by the Unconventional Secretion of Cytosolic Proteins. Molecules. 2023; 28(10):4033. https://doi.org/10.3390/molecules28104033
Chicago/Turabian StyleYamada, Kohji, Yoshito Hannya, Tsunekazu Oikawa, Ayano Yoshida, Kuniko Katagiri, Saishu Yoshida, Rei Koizumi, Naoko Tago, Yuya Shimoyama, Akira Kawamura, and et al. 2023. "Extended-Synaptotagmin 1 Enhances Liver Cancer Progression Mediated by the Unconventional Secretion of Cytosolic Proteins" Molecules 28, no. 10: 4033. https://doi.org/10.3390/molecules28104033