
Piper tectoniifolium Kunth: A New Natural Source of the Bioactive Neolignan (−)-Grandisin

 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Results and Discussion
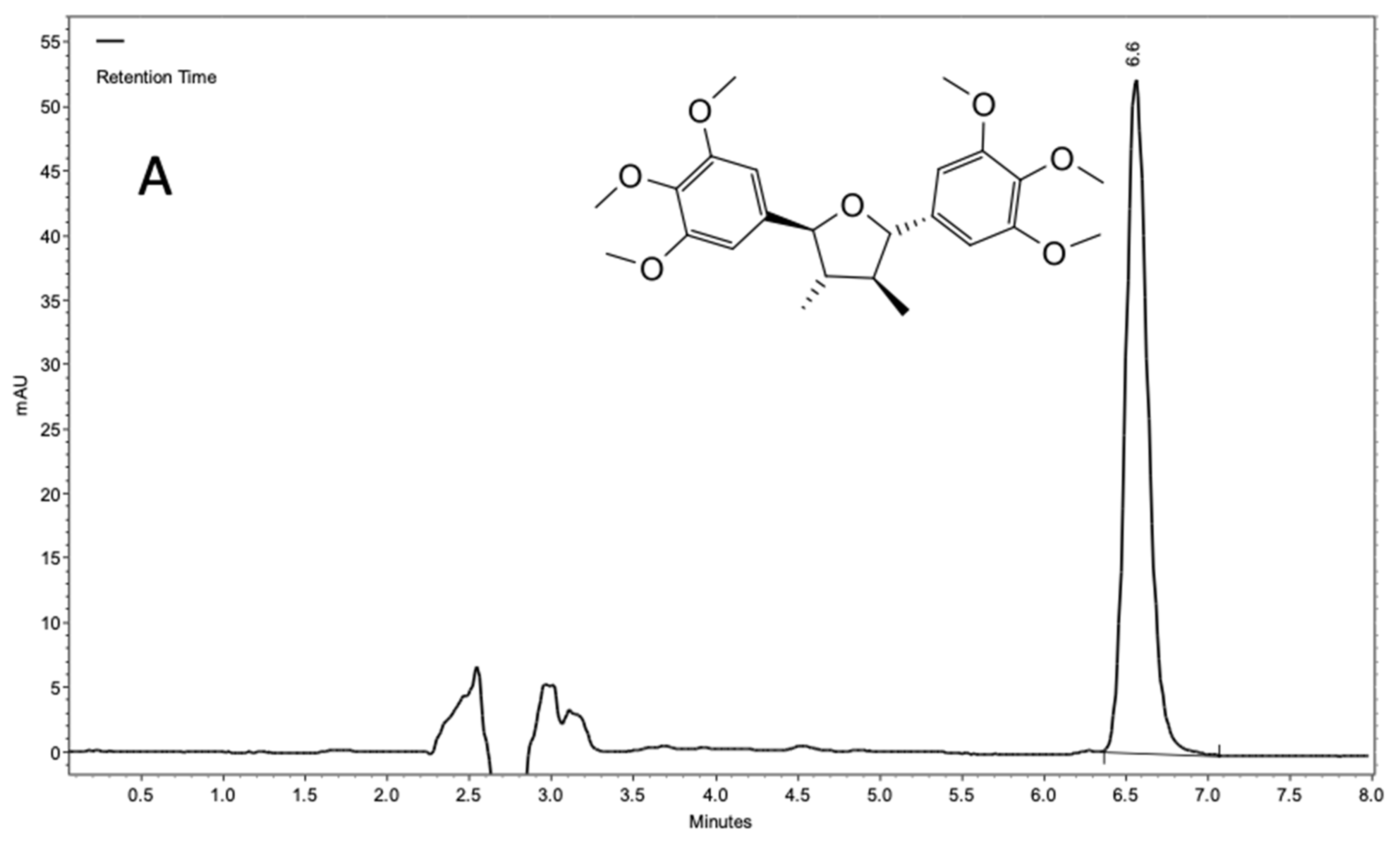
2.1. Chemical Characterization of Piper Tectoniifolium as a Source of (−)-Grandisin
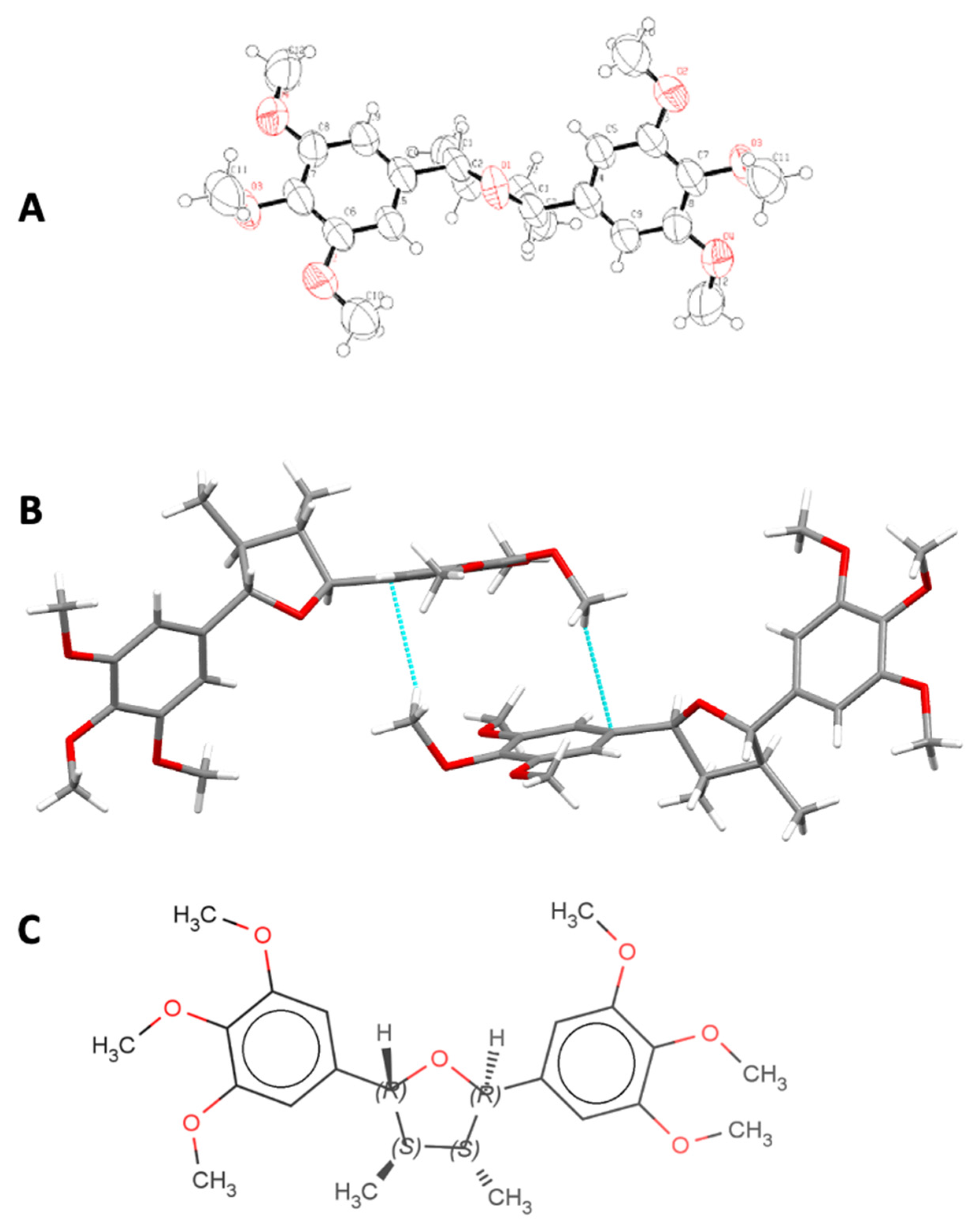
2.2. Crystalline and Molecular Structure of (−)-Grandisin Confirmed by X-ray Diffractometry
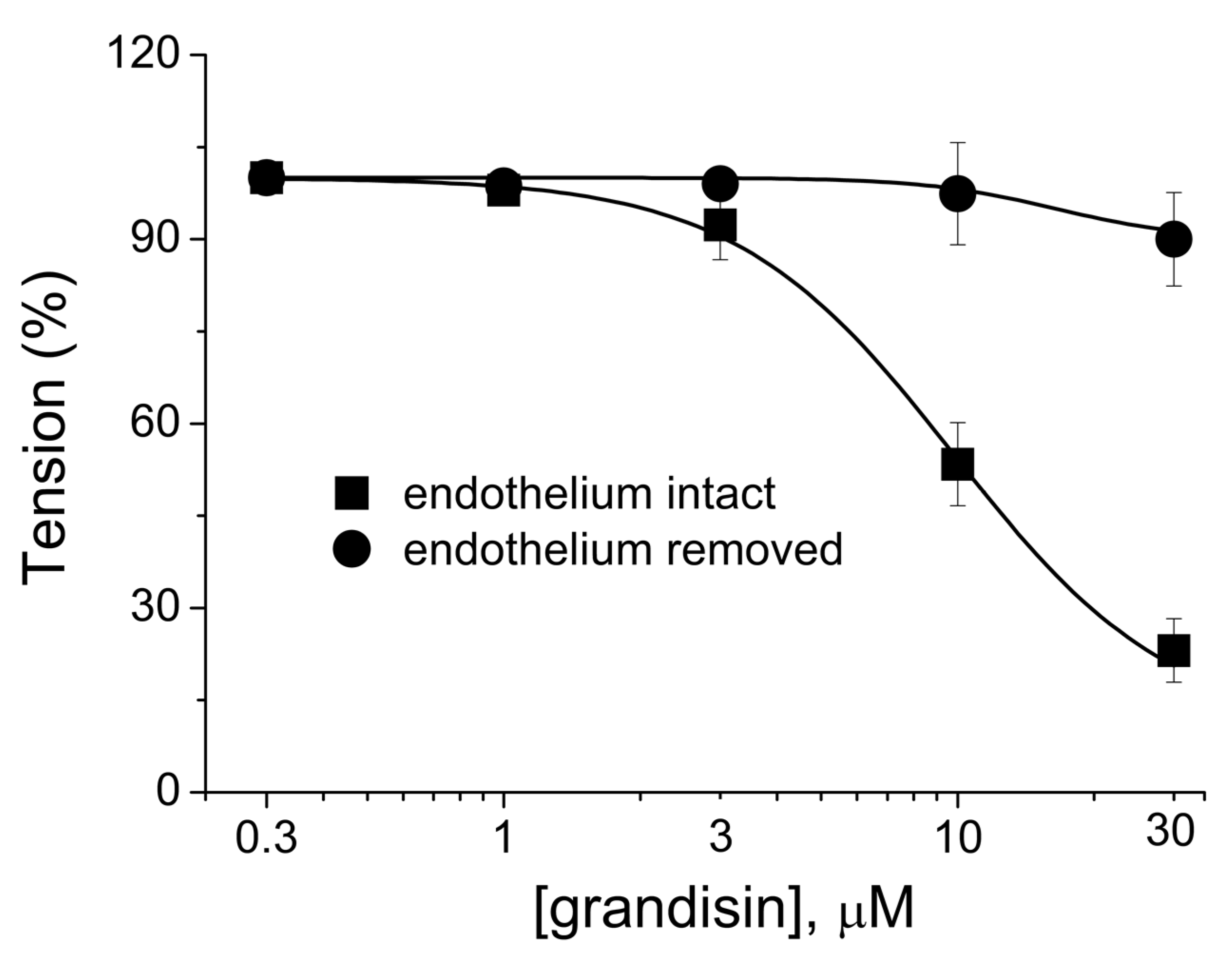
2.3. Pharmacological Evaluation
3. Materials and Methods
3.1. Chemicals
3.2. Botanical Material
3.3. Extract Preparation and Separation Procedures
3.3.1. Separation Procedures
(−)-Grandisin Isolation by Precipitation
(−)-Grandisin Isolation by Silica Gel Chromatographic Column
3.4. GC-FID Analysis
3.5. GC-MS Analysis
3.6. NMR Analysis
3.7. Quantification of the Major Isolated Lignan (−)-Grandisin from Extracts of P. tectoniifolium
3.8. X-ray Diffractometry
3.9. Pharmacological Evaluation
Ethics
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [PubMed]
- Dewick, P.M. Medicinal Natural Products—A Biosynthetic Approach, 3rd ed.; John Wiley & Sons: Chichester, UK, 2009; p. 550. [Google Scholar]
- Parmar, V.S.; Jain, S.C.; Bisht, K.S.; Jain, R.; Taneja, P.; Jha, A.; Tyagi, O.D.; Prasad, A.K.; Wengel, J.; Olsen, C.E.; et al. Phytochemistry of the genus Piper. Phytochemistry 1997, 46, 597–673. [Google Scholar] [CrossRef]
- Macedo, A.L.; Santos, T.C.C.; Valverde, A.L.; Moreira, D.L.; Vasconcelos, T.R.A. An Overview of Neolignans of the Genus Piper, L.: Isolation Methods and Biological Activities. Mini Rev. Med. Chem. 2017, 17, 693–720. [Google Scholar] [CrossRef]
- Polat Kose, L.; Gulcin, İ. Evaluation of the Antioxidant and Antiradical Properties of Some Phyto and Mammalian Lignans. Molecules 2021, 26, 7099. [Google Scholar] [CrossRef] [PubMed]
- Moreira, D.L.; Paiva, R.A.; Marques, A.M.; Borges, R.M.; Barreto, A.L.S.; Curvelo, J.A.R.; Cavalcanti, J.F.; Romanos, M.T.V.; de Araujo Soares, R.M.; Kaplan, M.A.C. Bioactive Neolignans from the Leaves of Piper rivinoides Kunth (Piperaceae). Rec. Nat. Prod. 2016, 10, 472–484. [Google Scholar]
- Raimundo, J.M.; Trindade, A.P.; Velozo, L.S.M.; Kaplan, M.A.C.; Sudo, T.S.; Zapata-Sudo, G. The lignan eudesmin extracted from Piper truncatum induced vascular relaxation via activation of endothelial histamine H1 receptors. Eur. J. Pharmacol. 2009, 606, 150–154. [Google Scholar] [CrossRef]
- Saad, J.M.; Soepadamo, E.; Fang, X.P.; McLaughlin, J.L.; Fanwick, P.E. (-)-Grandisin form Cryptocarya crassinervia. J. Nat. Prod. 1991, 54, 1681–1683. [Google Scholar] [CrossRef]
- Martins, R.C.C.; Lago, J.H.; Albuquerque, S.; Kato, M.J. Trypanocidal tetrahydrofuran lignans from inflorescences of Piper solmsianum. Phytochemicals 2003, 64, 667–670. [Google Scholar] [CrossRef]
- Carvalho, A.A.; Galdino, P.M.; Nascimento, M.V.M.; Kato, M.J.; Valadares, M.C.; Cunha, L.C.; Costa, E.A. Antinociceptive and antiinflammatory activities of grandisin extracted from Virola surinamensis. Phytother. Res. 2010, 24, 113–118. [Google Scholar] [CrossRef]
- Ma, Y.; Han, G.Q.; Li, C.L.; Cheng, J.R.; Arison, B.L.; Hwang, S.B. Neolignans from Piper polysyphorum C.DC. Yao Xue Xue Bao 1991, 26, 345–350. [Google Scholar]
- Atanasov, A.G.; Waltenberger, B.; Pferschy-Wenzig, E.M.; Linder, T.; Wawrosch, C.; Uhrin, P.; Temml, V.; Wang, L.; Schwaiger, S.; Heiss, E.H.; et al. Discovery and resupply of pharmacologically active plant-derived natural products: A review. Biotechnol. Adv. 2015, 33, 1582–1614. [Google Scholar] [CrossRef] [Green Version]
- Valadares, M.C.; Carvalho, I.C.; Oliveira, J.L.; Vieira, M.S.; Carvalho, F.S.; Andrade, L.V.; Lima, E.M.; Kato, M.J. Cytotoxicity and antiangiogenic activity of grandisin. J. Pharm. Pharmacol. 2009, 61, 1709–1714. [Google Scholar] [CrossRef] [PubMed]
- Barth, T.; Habenschus, M.D.; Lima, M.F.; Ferreira, L.S.; Lopes, N.P.; Moraes, O.A.R. In vitro metabolism of the lignan (-)-grandisin, an anticancer drug candidate, by human liver microsomes. Drug Test. Anal. 2015, 7, 780–786. [Google Scholar] [CrossRef] [PubMed]
- Cortez, A.P.; Menezes, E.G.P.; Benfica, P.L.; Santos, A.P.; Cleres, L.M.; Ribeiro, H.O.; Lima, E.M.; Kato, M.J.; Valadares, M.C. Grandisin induces apoptosis in leukemic K562 cells. Braz. J. Pharm. Sci. 2017, 53, e15210. [Google Scholar] [CrossRef] [Green Version]
- Cabral, M.M.; Alencar, J.A.; Guimarães, E.A.; Kato, M.J. Larvicidal activity of grandisin against Aedes aegypti. J. Am. Mosq. Control Assoc. 2009, 25, 103–105. [Google Scholar] [CrossRef]
- Pan, W.H.; Xu, X.Y.; Shi, N.; Tsang, S.W.; Zhang, H.J. Antimalarial activity of plant metabolites. Int. J. Mol. Sci. 2018, 19, 1382. [Google Scholar] [CrossRef] [Green Version]
- Costa, E.C.; Cassamale, T.B.; Carvalho, D.; Bosquiroli, L.S.S.; Ojeda, M.; Ximenes, T.V.; Matos, M.F.; Kadri, M.C.; Baroni, A.; Arruda, C.C. Antileishmanial Activity and Structure-Activity Relationship of Triazolic Compounds Derived from the Neolignans Grandisin, Veraguensin, and Machilin, G. Molecules 2016, 21, 802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habenschus, M.D.; Moreira, F.L.; Lopes, N.P.; Oliveira, A.R.M. In Vitro Inhibition of Human CYP450s 1A2, 2C9, 3A4/5, 2D6 and 2E1 by grandisin. Planta Med. 2017, 83, 727–736. [Google Scholar] [CrossRef]
- Felipe, D.F.; Dias Filho, B.P.; Nakamura, C.V.; Franco, S.L.; Cortez, D.A.G. Analysis of neolignans compounds of Piper regnellii (Miq.) C. DC. var. pallescens (C. DC.) Yunck by HPLC. J. Pharm. Biomed. Anal. 2006, 41, 1371–1375. [Google Scholar] [CrossRef]
- Messiano, G.B.; Silva Santos, R.A.; Ferreira, L.S.; Simões, R.A.; Jabor, V.A.P.; Kato, M.J.; Lopes, N.P.; Pupo, M.T.; de Oliveira, A.R.M. In vitro metabolism study of the promising anticancer agent the lignan (-)-grandisin. J. Pharm. Biomed. Anal. 2013, 72, 240–244. [Google Scholar] [CrossRef]
- Stecanella, L.A.; Taveira, S.F.; Marreto, R.N.; Valadares, M.C.; de Sousa Vieira, M.; Kato, M.J.; Lima, E.M. Development and characterization of PLGA nanocapsules of grandisin isolated from Virola surinamensis: In vitro release and cytotoxicity studies. Rev. Bras. Farmacogn. 2013, 23, 153–159. [Google Scholar] [CrossRef] [Green Version]
- Ramos, C.S.; Linnert, H.V.; Moraes, M.M.; Amaral, J.H.; Yamaguchi, L.F.; Kato, M.J. Configuration and stability of naturally occurring all-cis-tetrahydrofuran lignans from Piper solmsianum. RSC Adv. 2017, 7, 46932. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, M.T.M.; Rezende, K.C.S.; Evora, P.R.B.; Bastos, J.K.; Cunha, W.R.; Andrade, M.L.; Celotto, A.C. The lignan (-)-cubebin inhibits vascular contraction and induces relaxation via nitric oxide activation in isolated rat aorta. Phytother. Res. 2013, 27, 1784–1789. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, R.; Menengat, T.; Andrade, G.; Cotrim, B.; Ponte, C.; Santos, W.C.; Resende, G. Microwave Assisted Synthesis of 4-Phenylquinazolin-2(1H)-one Derivatives that Inhibit Vasopressor Tonus in Rat Thoracic Aorta. Molecules 2020, 25, 1467. [Google Scholar] [CrossRef] [Green Version]
- INMETRO (2016). DOQ-CGCRE-008—Orientação sobre Validação de Métodos Analíticos. In Revisão 02.—Instituto Nacional de Metrologia, Qualidade e Tecnologia; INMETRO: Rio de Janeiro, Brazil, 2011. Available online: http://www.inmetro.gov.br/credenciamento/organismos/doc_organismos.asp?torganismo=calibensaios (accessed on 28 December 2021).
- Ferraz, A.P.; Seara, F.A.C.; Baptista, E.F.; Barenco, T.S.; Sottani, T.B.; Souza, N.S.; Domingos, A.E.; Barbosa, R.A.; Takiya, C.M.; Couto, M.T.; et al. BKCa Channel Activation Attenuates the Pathophysiological Progression of Monocrotaline-Induced Pulmonary Arterial Hypertension in Wistar Rats. Cardiovasc. Drugs Ther. 2021, 35, 719–732. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample * | (−)-Grandisin Average % | SD | RSD% | mg/g | SD |
|---|---|---|---|---|---|
| n-Hexane leaf extract | 52.78 | 0.52 | 0.98 | 527.80 | 5.20 |
| Methanol leaf extract | 1.77 | 0.02 | 1.35 | 17.70 | 0.20 |
| Methanol branch extract | 7.84 | 0.03 | 0.45 | 78.40 | 0.30 |
| Methanol inflorescence extract | 2.08 | 0.00 | 0.20 | 20.80 | 0.04 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marques, A.M.; da Rocha Queiroz, A.S.; Guimarães, E.F.; Mafud, A.C.; de Sousa Carvalho, P.; Mascarenhas, Y.P.; da Silva Barenco, T.; Souza, P.D.N.; Provance, D.W.; do Nascimento, J.H.M.; et al. Piper tectoniifolium Kunth: A New Natural Source of the Bioactive Neolignan (−)-Grandisin. Molecules 2022, 27, 1151. https://doi.org/10.3390/molecules27041151
Marques AM, da Rocha Queiroz AS, Guimarães EF, Mafud AC, de Sousa Carvalho P, Mascarenhas YP, da Silva Barenco T, Souza PDN, Provance DW, do Nascimento JHM, et al. Piper tectoniifolium Kunth: A New Natural Source of the Bioactive Neolignan (−)-Grandisin. Molecules. 2022; 27(4):1151. https://doi.org/10.3390/molecules27041151
Chicago/Turabian StyleMarques, André M., Alexandre Siqueira da Rocha Queiroz, Elsie F. Guimarães, Ana Carolina Mafud, Paulo de Sousa Carvalho, Yvonne Primerano Mascarenhas, Thais da Silva Barenco, Pâmella Dourila N. Souza, David William Provance, José Hamilton M. do Nascimento, and et al. 2022. "Piper tectoniifolium Kunth: A New Natural Source of the Bioactive Neolignan (−)-Grandisin" Molecules 27, no. 4: 1151. https://doi.org/10.3390/molecules27041151