
Recent Advances in Natural Polyphenol Research


Department of Pharmacy, University of Naples Federico II, Via Domenico Montesano 49, 80131 Napoli, Italy
*
Authors to whom correspondence should be addressed.
Molecules 2022, 27(24), 8777; https://doi.org/10.3390/molecules27248777
Submission received: 12 November 2022
/
Revised: 8 December 2022
/
Accepted: 9 December 2022
/
Published: 11 December 2022
(This article belongs to the Special Issue Natural Polyphenols in Human Health)
Abstract
:Polyphenols are secondary metabolites produced by plants, which contribute to the plant’s defense against abiotic stress conditions (e.g., UV radiation and precipitation), the aggression of herbivores, and plant pathogens. Epidemiological studies suggest that long-term consumption of plant polyphenols protects against cardiovascular disease, cancer, osteoporosis, diabetes, and neurodegenerative diseases. Their structural diversity has fascinated and confronted analytical chemists on how to carry out unambiguous identification, exhaustive recovery from plants and organic waste, and define their nutritional and biological potential. The food, cosmetic, and pharmaceutical industries employ polyphenols from fruits and vegetables to produce additives, additional foods, and supplements. In some cases, nanocarriers have been used to protect polyphenols during food processing, to solve the issues related to low water solubility, to transport them to the site of action, and improve their bioavailability. This review summarizes the structure-bioactivity relationships, processing parameters that impact polyphenol stability and bioavailability, the research progress in nanocarrier delivery, and the most innovative methodologies for the exhaustive recovery of polyphenols from plant and agri-waste materials.
1. Introduction
Natural polyphenols are secondary metabolites of plants, vegetables, cereals, fruits, coffee, tea, and other plants. The exceptional functionality and biocompatibility of the polyphenols have stimulated the interest of researchers to use them as building blocks in functional foods, supplements, cosmetics, and drugs [1,2]. They have a phenolic ring, a basic monomer responsible for the protective action against oxidative injury [3]. Polyphenolic compounds can moderate oxidative stress and prevent or even inhibit oxidation by chelating iron and scavenging reactive radicals [4,5]. Dietary polyphenols can act as antioxidants, anti-inflammatory, and antiallergic compounds, decrease and prevent age-related diseases, can help against cardiovascular events (i.e., through their hypocholesterolemic, anti-thrombotic, antihypertensive, and anti-atherogenic), cancer, osteoporosis, diabetes, and neurodegenerative diseases [6]. The dietary polyphenols’ bioavailability depends on the chemical and physical characteristics of the natural matrix that contains them, the stability during the digestive process, the intestinal enzymes’ metabolization, and intestinal microbiota [7]. The gut microflora can modify the polyphenols’ bioactivity and bioavailability [8]. Their bioaccessibility can be affected by preservation and processing methods, the interaction with the matrix components, and the fluids and enzymes secreted during digestion [9]. Physical, chemical, and enzymatic treatments can alter their properties. The preservation and processing can determine damage to the native polyphenol molecules and the production of new “process-derived” compounds [10]. Nanodelivery technology can improve polyphenols’ absorption, bioavailability, functional quality, and performance [11,12,13,14,15].
This review summarizes the natural polyphenols classes, the extraction and methods performed to isolate them from natural sources and agro-waste, the factors that affect their bioavailability, and the application and development of nanodelivery systems.
2. Polyphenols in Nature
Polyphenols are involved in plant defense against pathogens and ultraviolet radiation [16]. The plants’ outer layers contain higher phenolics [17]. Insoluble phenolics are in cell walls, while soluble phenolics are in the plant cell vacuoles [18]. The degree of ripeness during harvest time, pedoclimatic conditions, infections, processing, and storage can affect the polyphenolic content [19]. The phenolic acids (e.g., derivatives of cinnamic acid and benzoic acid), flavonoids (e.g., flavonols, flavanones, flavones, flavanols, isoflavones, and anthocyanins), lignans, and stilbenes are the most naturally occurring classes of compounds (Figure 1). The shikimate pathway produces the phenolic acids. The phenylpropanoids pathway forms lignans, lignins, flavonoids, and stilbenes [20,21]. The biosynthesis of complex polyphenols is linked to primary metabolism: the flavonoids’ ring B and the chromane ring originate from the amino acid phenylalanine, obtained from the shikimate pathway, whereas ring A is formed from three malonyl-CoA units added through sequential decarboxylation condensation reactions [22]. In food, polyphenolic compounds can impact astringency, bitterness, flavor, color, and oxidative stability [19].
3. Polyphenols Bioavailability
There is no relation between the concentration of polyphenols in food and their bioavailability in the human body. The polyphenols, after ingestion, pass through the gastrointestinal epithelium and enter the circulatory vessels to reach the site of action. In food, polyphenols can exist as aglycon, glycosides, esters, or polymers.
The polyphenols’ chemical structure limits the rate, absorption, and metabolites circulating in the plasma. The polyphenolic compounds with hydroxyl groups can be modified by methylation, glucuronidation, or sulfation enzymatic reactions. The 5–10% of total polyphenolic compounds may be metabolized in the small intestine. The rest of the polyphenols accumulate in the large intestine and are evacuated in the feces [23]. The conjugated polyphenols must be hydrolyzed by colonic microflora or intestinal enzymes (i.e., β-glucosidases and lactase-phlorizin hydrolase) before absorption [24]. During the absorption process, they are transformed into oligomeric phenols by gastric acid in the stomach, and glycosidic polyphenols are cleaved by cytosolic glucosidase and lactase in the small intestine into aglycon and glycoside(s) (e.g., glucose, xylose, and galactose) radicals [25]. Finally, intestinal bacterial enzymes can metabolize the remaining aglycone fraction. In the intestinal and colonic epithelium, polyphenols can be involved in conjugation reactions with methyl, glucuronide, or sulfate groups, making the identification of the metabolites in the blood and tissue complex [26]. The glycosides of quercetin and the isoflavones (genistein and daidzein) are not recovered in plasma or urine [27,28,29]. Instead, anthocyanins glycosides are the most representative circulating forms [30,31]. Experimental studies showed that quercetin, without glycosides, is absorbed at the gastric level [32], anthocyanins in the stomach [33,34], and proanthocyanidins [33,35] and hydroxycinnamic acids are absorbed by the small intestine [36]. The remaining polyphenols are hydrolyzed in the colon by microflora enzymes into aglycones that are metabolized into benzoic acid derivatives [37,38].
The polyphenols’ digestibility affects their biological properties [39]. Soluble polyphenols have more evident responses during gastrointestinal digestion since the cell wall does not protect them. Unfortunately, human enzymes cannot digest some cell wall materials.
The flavonoids linked to other macromolecules cannot exert their beneficial actions [37]. The heat and pressure application (processing parameters) can facilitate the disruption of the cell wall and their release improving their bioactivity [38].
The pH and number of OH groups in benzene rings can affect phenolic stability. Conjugated nonphenolic aromatic acids, such as trans-cinnamic acid, are stable at high pH. The aromatic acids with a single OH group (e.g., ferulic acid) are stable at high pH because they do not form quinone oxidation products. The aromatic acids, with two phenolic OH groups (e.g., caffeic acid) or three (e.g., gallic acid), are unstable at pH 7–11. The changes are ascribable to the two adjacent phenolic OH groups attached to the benzene ring. Flavonoid molecules (e.g., rutin) that have a wholly conjugated aromatic structure are influenced by pH because the spatial arrangement between the π-electron system and an OH group controls the extent of π-orbital overlap and susceptibilities to the chemical change. The flavonoids in which the first benzene ring is located in the meta-position (e.g., catechin, epicatechin) do not have planar structures. Therefore, the π-electrons of the two benzene rings cannot cooperate via conjugation and are less susceptible at high pH [40,41].
In plant-based food, polyphenols and cell wall polysaccharides co-exist, and their affinity may influence foods’ physicochemical and nutritional properties during processing and digestion. The affinity of cell wall polysaccharides with polyphenols depends on their structures, concentrations, temperature, pH, ionic strength, and the presence of proteins [42].
The enzyme concentrations, solubility, pH, digestion time [43], and processing methods (e.g., washing, refrigerating, fermentation, grain milling, roasting, juicing, blanching, and thermal processing) impact polyphenols’ bioaccessibility and absorption [9].
Cooking and freezing processes positively impact the polyphenols’ bioaccessibility since they soften the cell wall. The cooking medium also influences their bioaccessibility [44,45].
Pasteurization affects the polyphenols’ bioaccessibility in the function of the heat treatment intensity, steps involved in processing, and type of food, decreasing the adverse processing effects on small bioactive compounds and even increasing polyphenols content [46]. Pasteurization can enhance food polyphenols extraction since the temperature softens the cell wall [47].
Finally, the interactions between macronutrients, micronutrients, and other phytochemicals, in finished products may also impact polyphenols’ stability [48].
4. Polyphenols & Microbiota
Gut microbiota can break the flavonoid C-ring in different positions, producing simple phenolics from the A and B rings. Most of these metabolites are acid or aldehyde phenolics with 1, 2, and (or) 3 hydroxyl and methyl ester radicals. Non-flavonoid phenolics (e.g., hydrolyzable tannins, stilbenes, lignans, and hydroxy-benzoic acid derivatives) are absorbed in the small intestine based on their chemical complexity. The gut bacteria can hydrolyze the ester bonds in tannins, dehydroxylate, and decarboxylate, the gallic acid [49], and reduce the resveratrol and its precursors [50].
The role of polyphenols and their metabolites on the gut microbiota is not elucidated. They probably have a prebiotic-like effect [51], since they can modulate the gut microbial profile. [52,53].
The polyphenols’ prebiotic effect is associated mainly with the promotion of probiotics (e.g., Bifidobacteriaceae and Lactobacillaceae) or the inhibition of pathogenic bacteria (i.e., E. coli, Clostridium perfringens, and Helicobacter pylori [52]) resulting in reduced proinflammatory immune response and decreased risk of colon cancer, gastroenteritis, inflammatory bowel disease, and metabolic syndrome [54,55].
Some polyphenols prevent bacterial growth, binding the cell membranes in a concentration-dependent manner. Catechins can change the microbial (i.e., Bordetella bronchiseptica, Klebsiella pneumonie, E. coli, Pseudomonas aeruginosa, Serratia marcescens, Bacillus subtilis Salmonella choleraesis, and Staphylococcus aureus) cell membrane permeability by producing H2O2 [56]. In Gram-positive bacteria, polyphenolic compounds can delay the oligopeptides autoinducers that sense the bacterial quorum sensing. In Gram-negative bacteria, they can prevent the bacteria-acylated homoserine lactones autoinducers [57].
5. Effects of the Food-Processing Techniques on Polyphenol Levels and Bioavailability
The heat treatments (e.g., boiling, steaming, frying, stewing, baking, roasting, ovens, steam, and microwave) and the transformation food processing (e.g., roasting, toasting, drying, pasteurization, canning, and sterilization), can affect the polyphenols’ bioavailability. The heat breaks cell walls, mobilizes the phenolic compounds, improves their availability, enhances their oxidation processes, and degrades them based on their thermostability. Domestic cooking and industrial thermal processes can cause losses in polyphenols, with significant variability depending on the nature of food matrices [58].
Boiling produces the most harmful polyphenol composition changes. Instead, steaming and frying can preserve them since the polar media (water) can extract higher levels of polyphenol than nonpolar media (oil) [59]. During boiling, heat decomposes the tissues, and the phenolics leak into the water [60]. Water volume can impact the polyphenol alteration during the heat process: small water volumes produce lower phenolic extraction than larger volumes [61]. Diverse boiling times produce different polyphenol profiles in foods, and a long time can cause more severe damage than a short one [62]. The type of heat treatment affects the polyphenol bioavailability. Steaming is the best heat method to preserve phenolic fractions since they are indirectly exposed to water [63]. The form in which phenolics are present also affects bioavailability [64].
Canning, a process employed to produce sterilized and microbiologically safe food products by applying heat treatment, can decrease phenolic compound levels [65] because they migrate into the surrounding medium [66].
Drying, the preservation process that aims to decrease the moisture content of food by using heat and mass transfer, can affect the phenolic levels in the function of the temperature regime. Freeze-drying is the most efficient method to preserve phenolic content, while hot-air-drying is the least. The vast variety of chemical polyphenol classes also influences the variability in the effects caused by drying [67]. Oven-dried processes produce higher levels of bioaccessible phenolics than other drying processes [68,69,70]. Slow freezing enhances the bioavailability of the phenolic compounds since it forms ice crystals that favor the polyphenols extraction, oxidation, and degradation, during digestion [71].
Peeling fruits and vegetables determines the loss of high amounts of bioactive compounds since they are contained in the peel and external parts of the plants at higher levels than other parts [72]. Grinding, the technique that reduces the size of solid particles using mechanical forces, enhances the polyphenols extractions as a function of the particle size [73]. The ultrasound treatments pulsed electric field, high-pressure, and pulsed-light processing enhance polyphenols digestion, bioaccessibility, and bioavailability [74].
6. Polyphenol Biological Activities
Epidemiological studies have shown an inverse association between a polyphenolic-rich diet and the risk of chronic human diseases. Polyphenolic compound-rich foods and beverages can have antioxidant, anti-inflammatory, anticancer, and anti-aging properties and reduce the risk of degenerative diseases such as cardiovascular, diabetes mellitus, and neuronal diseases.
6.1. Antioxidant Activity
Experimental evidence showed that polyphenols protect cell constituents against oxidative damage and degenerative diseases associated with oxidative stress [75]. The polyphenol-rich foods can improve plasma antioxidant capacity by scavenging radical species (e.g., ROS, RNS) or repressing radicals’ formation by inhibiting the activities of the oxido-reductive enzymes’ and/or chelating the metals that intercept free radical production. Their phenolic groups can accept an electron to form phenoxy radicals, interrupting chain oxidation reactions, and conjugated aromatic systems can delocalize an unpaired electron [76]. Polyphenols reduce the oxidation of lipids and other molecules by donating hydrogen to radicals (R). The resonance makes PO· (phenoxy radical) relatively stable (new chain reactions are not started) and acts as terminators of the propagation route when reacting with other free radicals (Figure 2).
The reduction activity of phenolic acids and their derivatives depends on the free hydroxyl groups in the molecule [77]. The hydroxycinnamic acids show better antioxidant activity than hydroxybenzoic acid equivalents due to the aryloxy-radical stabilizing effect of the –CH=CH–COOH linked to the phenyl ring by resonance [78]. The phenolic acids’ antioxidant activity of free, esterified, glycosylated, and nonglycosylated phenolics is mainly ascribed to radical scavenging via the hydrogen atom donation mechanism [78,79].
The flavonoids’ radical scavenging depends on the ortho-dihydroxy structure in the B ring, which allows higher stability to the radical form and participates in electron delocalization of the 2,3-double bond with a 4-oxo function in the C ring [77].
The chemical electron deficiency of anthocyanins is particularly reactive toward ROS/RNS.
The polyphenolic compounds with dihydroxy groups prevent metals-induced free radical formation by conjugating the transition metals that interact with hydrogen peroxide (H2O2) through the Fenton reaction to form hydroxyl radicals (·OH).
Phenolic compounds with catecholate and gallate groups can stop metal-induced oxygen radicals by improving metal ion autoxidation or forming an inactive complex with weaker interaction [79].
The metal ions can attack the flavonoids into positions 3′ and 4′ (B ring), 3 and 4, 3 and 5, 4-keto and 3-hydroxy, and 4-keto and 5-hydroxy (C ring) [80].
Moreover, polyphenols can improve cellular antioxidant activity by regulating Nrf2, which controls some detoxifying enzymes (SOD, GSH, GPx1, NADP(H) quinone oxidoreductase, HO-1, and GST) [77]. Finally, polyphenols can influence microRNAs [81].
MicroRNAs are a class of small, endogenous, noncoding RNAs. Some microRNAs (i.e., miR-21, miR-125, and miR-146) are involved in vascular inflammation and diseases [82,83,84]. Dietary polyphenols can influence the microRNAs’ expression and biogenesis [30]. For example, curcuminoids can act as anti-atherosclerosis agents by upregulating miR-126 expression [85]. Resveratrol can act as a cardioprotective molecule by improving the mRNA activating SIRT1, and enhancing the SOD’ levels [86,87,88,89].
Gallic acid can decelerate atherosclerosis progression by upregulating miR-145 and downregulating miR-21 expression [90].
Under certain conditions, the polyphenolic compounds can initiate an autoxidation process and perform as prooxidants. In these cases, the phenoxy radicals can interact with oxygen to make quinones and superoxide anions [91]. pH, high concentrations of transition metal ions, and oxygen molecules can induce the autooxidation of polyphenols [92]. Quercetin and gallic acid can have prooxidant activity; instead, the hydrolyzable tannins have little or no prooxidant activity [93].
6.2. Anti-Inflammatory Activity
Plant polyphenols can decrease the effect of the cytokine, affecting their receptors or reducing their secretion processes [94].
Phenolic compounds can suppress the binding of proinflammatory mediators, control eicosanoid synthesis, prevent stimulated resistant units, and impede the activity of COX-2 and NO synthase, acting on NF-κB [95]. Some phenolic acids, such as rosmarinic acid and isosalvianolic acid, can reduce the production of IL-6, TNF-α, and IL-1β at the gene and protein levels [30].
6.3. Anticancer Activity
Cancer development consists of initiation, promotion, progression, invasion, and metastasis [100]. Genetic mutations occur when DNA damage is not repaired, and a clone of mutated cells is reproduced during mitosis. Tumor promotion is a reversible and long-term process in which a selective clonal expansion of the cells forms a population of aggressively proliferating multi-cellular cells (premalignant tumor). Clonal expansion determines the development of the premalignant cells into tumors (tumor progression phase). Finally, some tumor cells may be cut off from the primary tumor mass, migrate toward blood vessels or lymphatic vessels and produce a second lesion (invasion and metastasis phases). The natural phenolic compounds can induce cell cycle arrest at G1, S, S-G2, and G2 phases by down-regulating cyclins and cyclins-dependent kinases or producing the expression of p21, p53, and p27 genes [80].
Polyphenols can act against tumor initiation and promotion, changing the redox status and affecting essential cellular functions (i.e., cell cycle, apoptosis, angiogenesis, inflammation, invasion, and metastasis) [101]. Oxidative damage can cause cancer since ROSs can damage the DNA and affect cell replication and signal transduction [102].
The flavonoid anticancer effects are related to their antioxidant and pro-oxidant activities [103,104]. The flavonol kaempferol can induce apoptosis and arrest in the S-phase of cancerous cells by modulating ROS levels [105]. When it acts as pro-oxidants it decreases NF-κB levels and produces cyclooxygenase-2 (COX) overexpression, inducing apoptosis, and cell-cycle arrest [106].
Some flavonoids and resveratrol can affect the procarcinogens’ activation by impeding phase I metabolizing enzymes (e.g., cytochrome P450) [107,108,109,110]. They can help carcinogens’ detoxification and removal inducing the phase II metabolizing enzymes (e.g., glutathione S-transferase, UDP-glucuronyl-transferase, and NAD(P)H quinine oxidoreductase) [111].
The polyphenols can produce apoptosis-inducing cell cycle arrest inhibiting the extracellular regulated kinase, c-Jun N-terminal kinase, and P38 mitogen-activated protein kinase pathway, transcription factors, NF-κB, activator protein-1 (AP1), protein kinase C (PKC), and growth factor-mediated pathways. The apigenin inhibits the growth of human thyroid carcinoma cells, probably by decreasing the phosphorylation of MAPK and by activating the protein kinases, and scavenging H2O2 [112].
The 3,4 dihydroxybenzoic acid stimulates apoptosis, in human gastric carcinoma cells, by ROS overproduction which can activate JNK/p38 MAPKs [113].
The polyphenolic compounds can negatively affect some factors involved in the inflammatory processes, such as the NF-κB, proinflammatory cytokines release, COX-2, lipoxygenases, inducible nitric oxide synthase, and MAPK-mediated pathway [80]. For example, Epigallocatechin gallate can block NFκ B activation in human epithelial cells and downregulate the expression of inducible nitric oxide synthase and nitric oxide production in macrophages [114]. Finally, the kaempferol can counteract malignant cell invasion and metastasis, down-regulating the matrix metalloproteases (MMP-2 and MMP-9), urokinase-plasminogen activator (uPA), and uPA receptor expression [115,116].
6.4. Cardiovascular Protective Activity
Cardiovascular (CVD) pathologies are the primary cause of morbidity and mortality (ischemic heart disease and stroke contribute 85%) [117]. Oxidative stress and inflammatory processes are considered promoters of endothelial dysfunction [118,119]. Polyphenols have antioxidant, anti-inflammatory abilities and can modulate lipid metabolism [120]. They (mainly quercetin and resveratrol) can decrease LDL oxidation [121], cholesterol synthesis, improve LDL receptor expression and activity [122,123], and the cholesterol transporters expression [124]. The anthocyanins and resveratrol can improve fecal cholesterol elimination [125,126] and decrease the triglyceride plasma level, decreasing the apolipoprotein B48 and apolipoprotein B100 production in the liver and intestine [127] or the lipoprotein lipase expression [128]. Flavonoids can also reduce blood pressure ameliorating flow-mediated dilation in humans (by improving the NO synthase activity) [129,130] and influencing the renin-angiotensin system [131,132]. Moreover, they can prevent platelet aggregation, decreasing the activity of cyclooxygenase 1, and thromboxane A2 that act as a vasoconstrictor and platelet aggregation’s inducer, respectively [133]. Finally, the polyphenols’ prebiotic-like activity can account for the amelioration of markers of CVD [124].
6.5. Antidiabetic Activity
There are two main types of diabetes (diabetes-1 and diabetes-2). Diabetes type-2 or diabetes mellitus is due to damage in glucose metabolism and advancing insulin resistance that leads to hyperglycemia. The leading causes of hyperglycemia are dietary carbohydrates’ digestion and absorption, glycogen storage reduction, β-cell dysfunction, peripheral tissue insulin resistance, deficiency in insulin signaling pathways, and improved gluconeogenesis and production of hepatic glucose [134]. Polyphenols can decrease the intestinal absorption of carbohydrates, control the enzymes that regulate glucose metabolism, and increase the β-cell functionality, insulin secretion, and the anti-inflammatory and antioxidant properties of these components (Figure 3).
The phenolic acids, flavonoids, and tannins can regulate the key enzymes responsible for the digestion of carbohydrates (α-glucosidase and α-amylase) [135]. The catechin, epicatechins, and chlorogenic, caffeic, ferulic, and tannic acids can decrease the glucose transporters Na+-dependent (SGLT1 and SGLT2) [136]. The coffee phenols, anthocyanin, and curcumin can regulate postprandial glycemia and decrease the progression of glucose intolerance by a simplified insulin response and improved secretion of glucagon-like polypeptide-1 (GLP-1) and glucose-dependent insulinotropic polypeptide (GIP) [137]. Ferulic acid can decrease blood glucose by improving glucokinase activity and glycogen production in the liver [138]. Catechins and epicatechins can decrease hyperglycemia and hepatic glucose output, downregulating the expression of liver glucokinase, and upregulating the glucose-6-phosphatase and phosphoenolpyruvate carboxykinase [139].
6.6. Neurodegenerative Protection
Neurodegenerative diseases are due to the deterioration of neurons’ structure and/or function. Reactive oxygen and reactive nitrogen species can determine neuronal cell dysfunction and death. The phenolic compounds can interact with the amino acid residues of acetylcholinesterase’s (AChE) active site, making hydrogen bonds and hydrophobic and π–π interactions. Multiple hydroxyl groups can improve the inhibition of AChE, increasing the binding capacity [30]. Resveratrol can protect against microglia-dependent β-amyloid toxicity by decreasing the nuclear factor κB [140]. Some polyphenols protect against Parkinson’s disease by scavenging the neurotoxin N-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)-mediated radical formation [141] or decreasing free radicals’ formation by chelating iron [142].
6.7. Anti-Aging Action
Aging determines detrimental changes in the cells and tissues. The cosmetic industry constantly strives in product development and reformulation to meet consumers’ preferences. Today, nature-derived products are in demand on the market. Some botanical preparations that contain polyphenols (e.g., flavonoids, phenolic acids, and stilbenes) are employed in the composition of anti-aging products [143]. Free radicals and oxidative stress are the major contributors to aging damage. The phenolic hydroxyl groups on polyphenol molecules can scavenge ROS [144]. The polyphenolic compounds can regulate the production of oxidase enzymes (sodium oxide dismutase 1 in the cytosol, sodium oxide dismutase 2 in the mitochondria), and endogenous antioxidants [145,146,147,148,149] can improve the transcriptional factor Nrf2 DNA-binding activity and regulate protein expression [150,151,152]. Anti-aging formulations contain botanicals metabolites able to protect DNA, regulate the enzymes’ action, decrease inflammation, and alter hormone imbalance [143].
The epigallocatechin-3-gallate in green tea decreases the UVB-induced hydrogen peroxide release from normal epidermal keratinocytes, MAPK phosphorylation, and inflammation by activating NFkB. The flavins in black tea decrease UVB-induced AP-1 induction, prevent UVB-induced phosphatidyl-inositol 3-kinase activation, decrease the amount of ROS in the skin, and offer photoprotection by reducing local and systemic immunosuppression UVB-induced [153]. Resveratrol is employed to reduce hydrogen peroxide, improve lipid peroxidation, and decrease the levels of COX-2 and ornithine decarboxylase. Moreover, it can decrease UVA-induced oxidative stress in human keratinocytes since it controls the Keap1-a protein that acts on Nrf2 [153]. Curcuminoids (found in Turmeric spice) can decrease inflammation by inhibiting the MAPK and NFkB signaling pathways and decreasing nitric oxide levels and COX2. Moreover, in keratinocytes and fibroblasts, curcuminoids can decrease UVB-induced TNF mRNA expression and matrix metalloproteinase-1 expression [154].
6.8. Antiallergic Action
Allergic diseases happen when an organism becomes sensitive to an innocuous allergen and releases many allergy-related intermediaries. Polyphenols limit the production of IgE, the release of allergic mediators, and allergy symptoms. Polyphenols can control hypersensitivity by regulating oxidation and interacting with inflammatory mediators [155]. Catechins can decrease Th2 cytokine production and T cell activation and proliferation. Caffeic, chlorogenic, and ferulic acids can irreversibly bind peanut allergens (Ara h1 and Ara h2), reducing their allergenicity [156]. Punicalagin, phloridzin, and rutin can improve the growth of probiotics such as Lactobacillus and Bifidobacterium, which positively impact food allergies [157].
6.9. Antiosteoporotic Action
Osteoporosis causes the loss of bone mineral density, decreased bone mass, and microstructural deterioration. Flavonoids and stilbenes can improve osteogenesis by controlling the bone morphogenetic protein, NF-κB, IGF, and MAPK, and can inhibit the osteoclastogenesis pathways through epigenetic regulations. They can activate SIRT-1 (histone deacetylase) and modify the NAD+/NADH ratio [158,159,160].
6.10. Antimicrobial Action
Some plant extracts rich in polyphenols can decrease the growth of fungi and bacteria (i.e., Listeria monocytogenes, Salmonella spp., and Escherichia coli) [161,162], minimize the exposure of humans to resistant bacteria [163], and can have a synergic action with other antimicrobials. These findings have suggested a potential use of polyphenol-rich extracts as food preservatives and in the pharmaceutical industry to improve efficacy and decrease antibiotic side effects, such as repressing antibiotic-resistant bacteria [164]. The polyphenol-rich extracts can be placed on the food surface by spraying, dipping, brushing, or mixing with other ingredients [165]. Unfortunately, in some cases, the interaction with food components can cause a lack of antimicrobial efficacy. Therefore, it was thought to encapsulate them in carriers to increase their distribution in the food and reduce contact with food matrix molecules that reduce their effectiveness [166]. The mechanisms of antibacterial action are not yet entirely deciphered. However, it is known that many sites of action at the cellular level are involved. Polyphenols can modify the cell membrane permeability, destroy the cell wall integrity and change intracellular functions by binding some enzymes [167].
7. Polyphenols Potentialities in the Nutraceutical Era
Today, consumers include a high level of bioactive compounds in their standard diet, preferably derived from natural sources such as plants and fruits, in the hope of giving more life to the years by preventing disabling pathologies that decrease the skills that allow living life in all its manifestations [168]. Food and pharmaceutical companies develop nutraceutical foods and supplements that contain botanical extracts and metabolites alone or combined with other ingredients [169,170,171].
Numerous studies strongly suggested that including polyphenols or polyphenol-rich extracts in supplements or foods may protect the body tissues against oxidative stress and aging [172,173].
The primary issue in using plant extracts is that fungi, which can produce toxins, can contaminate the extracts (e.g., Aspergillus section Nigri that produces Ochratoxin A, a carcinogenic, teratogenic, nephrotoxic, neurotoxic, and immunotoxic toxin) [174]. The research should implement efforts to develop analytical controls that safeguard consumer safety.
8. Polyphenols Extraction
Extraction plays a pivotal role in the purification of polyphenols from foodstuffs. Extraction techniques can employ traditional or “conventional” such as percolation, maceration, and Soxhlet extraction, and modern methods, such as ultrasound or microwave, the latter is most extensively used (Figure 4). In both cases, extraction efficiency depends on various factors such as the nature of the solvent, solvent–solid ratio, temperature, and particle size. Polyphenol can be extracted from fresh, frozen, or dried plant samples. The extracts can be added to an organic solvent, such as methanol or ethanol, with low viscosity to accelerate mass transfer [175]. Before extraction, the pretreatment of the plant matrices (e.g., cleaning, washing, milling, grinding, drying, homogenizing) is crucial.
Percolation extraction uses water as a solvent. It takes a long time to obtain the pure extract to be concentrated [176].
The maceration is a solid-liquid extraction method using different solvents depending on the target compounds’ physical and chemical properties. It has low efficiency and extraction yield and employs a large volume of solvents compared to non-conventional techniques such as ultrasound-assisted extraction (UAE) [177]. A higher ratio of solid/solvent increases polyphenols recovery [178].
Decoction extracts plant materials by boiling. It is inefficient for heat and light-sensitive compounds [179]. The decoction of Citrus fruits produces by-products of Citrus peels with high concentration levels of polyphenol fraction [180].
Heat reflux extraction is a solid–liquid extraction method performed with repeated solvent evaporation and condensation at a constant temperature. It requires less extraction time and solvent than percolation or maceration and allows for a greater extraction yield [181]. Polyphenols from wastes of Vitis vinifera were extracted by Moldovan et al. by using heat reflux extraction [182].
The solid-phase extraction (SPE) method is considered quick and easy to extract polyphenols from vegetable oils. According to the experimental needs, different stationary phases were used (e.g., C8 cartridges, octadecyl C18, diol-bonded phase cartridges, amino-phase cartridges, and octadecyl C18EC) [6]. The non-conventional techniques employ supercritical fluid, high-voltage electric discharge, and enzyme-assisted extraction. It is preferable to extract the bound polyphenols, also referred to as non-extractable polyphenols (NEP), from plant sources using one or more combinations of modern technology rather than conventional methods. A comparison between traditional and SFE extraction performed on black tea leftovers showed that SPE technique gives the best performance in extraction of phenolic compounds (SPE gives 521 mg GAE/g; traditional gives 283 mg GAE/g) [183].
The extraction with supercritical fluids such as CO2, propane, argon, and SF6 allows for an easy penetration inside plant materials and high solvents. Power-pulsed electric fields (PEF) perform a gentle extraction due to the electroporation of cell membranes. It has been applied to exotic fruits, grapes, and oil crop components [184]. Microwave-assisted extraction (MAE) is an eco-friendly technique with higher efficiency in the recovery of polyphenols from waste products if compared to that of extracts prepared by ultrasound-assisted extraction (UAE) and conventional methods such as maceration [185].
9. Polyphenol Nano Delivery Systems
The potential of antioxidative and repair pathways decreases with age, causing several adverse effects such as the risk of neurodegenerative diseases such as Parkinson’s disease, memory loss, Alzheimer’s disease, atherosclerosis, and cancers due to the accumulation of reactive oxygen species. Polyphenols, due to their oxidizing ability, can protect from the damaging effects of ROS. Therefore, they can be used as active compounds in several formulations preventing oxidative stress [197,198]. The loading of polyphenols into lipid nanocarriers (NCs) is an essential tool for increasing bioavailability, reducing degradation, and protecting antioxidant polyphenols’ activity. The NCs are biodegradable and have no significant toxicity. The nanoemulsion, liposome, phytosome, solid lipid nanoparticles (SLNs), nanostructured lipid carrier (NLCs), and lipid-polymer hybrid nanoparticles (LPHNs) can encapsulate polyphenols to improve their biophysiological target [199]. Nanoparticles (NPs) have diameters as small as 1–100 nm. They can enhance polyphenols delivery and promote their absorption and bioavailability [200]. The transcellular pathway is a route for the NPs transportation (via endocytosis or macropinocytosis mechanisms) and subjected to the degradative microenvironment of the cellular lysosome environment in the acidic endosomal lumen [201]. Carbohydrate-based delivery systems, such as mono, oligo, and polysaccharide, are employed to encapsulate polyphenols due to their abundance and low cost; for example, curcumin was encapsulated in chitosan using an injection-gelation method to increase its bioavailability, antioxidant properties, and to improve stability and effects on tumor cells [202].
Protein-based systems have been employed to prepare nanoparticles for carrying polyphenols. The proteins act as “host” and polyphenols as “guest” molecules. The polyphenols bind specific regions on the protein surfaces through hydrogen and/or hydrophobic non-covalent bounds. The β-lactoglobulin nano delivery was used to increase the water solubility 3-fold at pH 7 of the epigallocathechin-3-gallate and naringenin [203]. Finally, polyphenols are used to design polyphenol-based nanomaterials for biomedical applications. For example, the polyphenol-grafted polymers are used as antidiabetic agents [204], the curcumin encapsulated in chitosan and polyglycolic acid (PGA) particles for wound healing [205], and the polyphenol-loaded electrospun nanofibers to improve the remineralization and regeneration of bone [206].
10. Polyphenols in Active Packaging
In recent years, packaging technology has evolved, including intelligent or smart packaging. Biodegradable, active, and bioactive packaging are new trends in food packaging research. Food contact materials (FCM) are engineered to protect foods, and improve their shelf-life [207]. Plastic packaging materials, such as polycarbonate, polyethylene, and polyethylene terephthalate, widely used in food packaging, are nonbiodegradable and disadvantageous to the environment and human health [208,209,210]. To date, natural and biodegradable biopolymer-based packaging films and edible coatings represent the alternative to plastic packaging materials [211]. Natural biopolymers, including proteins, polysaccharides, and lipids, have been used in packaging manufacturing [212]. Fruit industrial manufacture generates large amounts of waste that harm the environment and causes considerable treatment expense [213]. Nevertheless, these by-products are rich in bioactive compounds [214], some of which can be incorporated into biodegradable plastics for food packaging to protect the polymeric matrix against thermal [162,213], photo-induced degradation, and preserve the food freshness and quality [215]. The solvent-based impregnation of biodegradable polymers with extracts of Cistus linnaeus is used to improve the polymers’ thermal stability [216]. Chemically-synthesized and biomass-derived biodegradable polymers have been used as matrices to protect food during transportation, storage, and sale. Polyphenols are employed in the so-called “leaching systems” that are active-releasing systems able to release actives by direct contact between food and packaging material. For this purpose, the propolis is mixed with biopolymers, plasticizers, and reinforcing agents to produce active packaging and edible coatings [213,216].
Biopolymers can be dissolved in solvents depending on their hydrophilicity. Hydrophilic biopolymers, such as gelatin, κ-carrageenan, alginate, and agar in water, chitosan, can be dissolved in acidic solutions [217], while organic solvents, chloroform, and ethyl acetate can be employed for hydrophobic biopolymer such as polylactic acid (PLA) [218]. For example, the ethanolic propolis extract, and propolis in powder, together with plasticizers such as glycerol and (PEG) polyethylene glycol, have been added to biopolymeric solutions [219]. Tea polyphenols (TP) are employed as an active component in biopolymer materials for active food packaging. Shao et al. have incorporated TP into pullulan-carboxymethylcellulose sodium (Pul-CMC) solutions on electrospun nanofiber films [220].
11. Conclusions
Polyphenols are secondary plant metabolites that can benefit human health and preserve food. The use of polyphenols as supplements, antibiotic drugs, cosmetics, and natural food preservatives is a promising trend in the industry because of the growing interest in natural products and the multiple biological activities of these products.
The great demand for polyphenols and the small quantity produced by the plants have determined the need to use extraction techniques that allow exhaustive extraction even when it is necessary to recover them from non-traditional sources such as organic waste. For this purpose, unconventional techniques such as supercritical fluid, high-voltage electric discharge, and enzyme-assisted extraction must be optimized.
The polyphenols’ low oral bioavailability and interactions with other molecules negatively impact the possible industrial application. Therefore, different nanocarriers have been developed to protect, improve bioavailability, ensure achievement to the active site, and improve their effectiveness. It is essential to underline that polyphenols are commonly present in plant-based foods such as fruits and vegetables.
Guidelines for their consumption and supplementation should be provided by regulatory bodies to make consumers safe and informed.
Author Contributions
Conceptualization, writing—original draft preparation I.D. and L.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Li, Y.; Chemat, F. (Eds.) Food, Cosmetics and Drugs. In Plant Based “Green Chemistry 2.0”: Moving from Evolutionary to Revolutionary; Springer: Singapore, 2019; pp. 197–235. [Google Scholar]
- Becker, L.; Zaiter, A.; Petit, J.; Karam, M.-C.; Sudol, M.; Baudelaire, E.; Scher, J.; Dicko, A. How do grinding and sieving impact on physicochemical properties, polyphenol content, and antioxidant activity of Hieracium pilosella L. powders? J. Funct. Foods 2017, 35, 666–672. [Google Scholar] [CrossRef]
- Olszowy, M. What is responsible for antioxidant properties of polyphenolic compounds from plants? Plant Physiol. Biochem. 2019, 144, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Dini, I.; Izzo, L.; Graziani, G.; Ritieni, A. The Nutraceutical Properties of “Pizza Napoletana Marinara TSG” a Traditional Food Rich in Bioaccessible Antioxidants. Antioxidants 2021, 10, 495. [Google Scholar] [CrossRef] [PubMed]
- Dini, I. Spices and herbs as therapeutic foods. In Food Quality: Balancing Health and Disease; Holban, A.M., Grumezescu, A.M., Eds.; Academic Press Elservier: London, UK, 2018; pp. 433–469. [Google Scholar]
- Dini, I.; Seccia, S.; Senatore, A.; Coppola, D.; Morelli, E. Development and Validation of an Analytical Method for Total Polyphenols Quantification in Extra Virgin Olive Oils. Food Anal. Methods 2020, 13, 457–464. [Google Scholar] [CrossRef]
- Bertelli, A.; Biagi, M.; Corsini, M.; Baini, G.; Cappellucci, G.; Miraldi, E. Polyphenols: From Theory to Practice. Foods 2021, 10, 2595. [Google Scholar] [CrossRef] [PubMed]
- Pasinetti, G.M.; Singh, R.; Westfall, S.; Herman, F.; Faith, J.; Ho, L. The Role of the Gut Microbiota in the Metabolism of Polyphenols as Characterized by Gnotobiotic Mice. J. Alzheimers Dis. 2018, 63, 409–421. [Google Scholar] [CrossRef]
- Wojtunik-Kulesza, K.; Oniszczuk, A.; Oniszczuk, T.; Combrzyński, M.; Nowakowska, D.; Matwijczuk, A. Influence of In Vitro Digestion on Composition, Bioaccessibility and Antioxidant Activity of Food Polyphenols-A Non-Systematic Review. Nutrients 2020, 12, 1401. [Google Scholar] [CrossRef]
- Debelo, H.; Li, M.; Ferruzzi, M.G. Processing influences on food polyphenol profiles and biological activity. Curr. Opin. Food Sci. 2020, 32, 90–102. [Google Scholar] [CrossRef]
- Milinčić, D.D.; Popović, D.A.; Lević, S.M.; Kostić, A.; Tešić, Ž.L.; Nedović, V.A.; Pešić, M.B. Application of Polyphenol-Loaded Nanoparticles in Food Industry. Nanomaterials 2019, 9, 1629. [Google Scholar] [CrossRef] [Green Version]
- Hu, B.; Liu, X.; Zhang, C.; Zeng, X. Food macromolecule based nanodelivery systems for enhancing the bioavailability of polyphenols. J. Food Drug Anal. 2017, 25, 3–15. [Google Scholar] [CrossRef]
- Hu, B.; Ting, Y.; Zeng, X.; Huang, Q. Cellular uptake and cytotoxicity of chitosan-caseinophosphopeptides nanocomplexes loaded with epigallocatechin gallate. Carbohydr. Polym. 2012, 89, 362–370. [Google Scholar] [CrossRef] [PubMed]
- Homayouni, H.; Kavoosi, G.; Nassiri, S.M. Physicochemical, antioxidant and antibacterial properties of dispersion made from tapioca and gelatinized tapioca starch incorporated with carvacrol. LWT 2017, 77, 503–509. [Google Scholar] [CrossRef]
- Serpa Guerra, A.M.; Gómez Hoyos, C.; Velásquez-Cock, J.A.; Vélez Acosta, L.; Gañán Rojo, P.; Velásquez Giraldo, A.M.; Zuluaga Gallego, R. The nanotech potential of turmeric (Curcuma longa L.) in food technology: A review. Crit. Rev. Food Sci. Nutr. 2020, 60, 1842–1854. [Google Scholar] [CrossRef]
- Fereidoon, S.; Vamadevan, V.; Won Young, O.; Han, P. Phenolic compounds in agri-food by-products, their bioavailability and health effects. J. Food Bioact. 2019, 5, 57–119. [Google Scholar] [CrossRef] [Green Version]
- Tzima, K.; Brunton, N.P.; Lyng, J.G.; Frontuto, D.; Rai, D.K. The effect of Pulsed Electric Field as a pretreatment step in Ultrasound Assisted Extraction of phenolic compounds from fresh rosemary and thyme by-products. Innov. Food Sci. Emerg. Technol. 2021, 69, 102644. [Google Scholar] [CrossRef]
- Zhang, B.; Zhang, Y.; Li, H.; Deng, Z.; Tsao, R. A review on insoluble-bound phenolics in plant-based food matrix and their contribution to human health with future perspectives. Trends Food Sci. Technol. 2020, 105, 347–362. [Google Scholar] [CrossRef]
- Pino, C.; Sepúlveda, B.; Tapia, F.; Saavedra, J.; García-González, D.L.; Romero, N. The Impact of Mild Frost Occurring at Different Harvesting Times on the Volatile and Phenolic Composition of Virgin Olive Oil. Antioxidants 2022, 11, 852. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Wu, J.; Ma, Y.; Wang, P.; Gu, Z.; Yang, R. Biosynthesis, metabolic regulation and bioactivity of phenolic acids in plant food materials. Shipin Kexue/Food Sci. 2018, 39, 321–328. [Google Scholar]
- Petersen, M.; Hans, J.; Matern, U. Biosynthesis of Phenylpropanoids and Related Compounds. In Annual Plant Reviews Volume 40: Biochemistry of Plant Secondary Metabolism; Wiley-Blackwell: Hoboken, JN, USA, 2010; pp. 182–257. [Google Scholar]
- Liu, W.; Feng, Y.; Yu, S.; Fan, Z.; Li, X.; Li, J.; Yin, H. The Flavonoid Biosynthesis Network in Plants. Int. J. Mol. Sci. 2021, 22, 12824. [Google Scholar] [CrossRef]
- Chen, L.; Cao, H.; Xiao, J. Polyphenols: Absorption, Bioavailability, and Metabolomics; Woodhead Publishing: Cambridge, UK, 2018; pp. 45–67. [Google Scholar]
- Brown, N.M.; Lydeking-Olsen Grgić, J.; Šelo, G.; Planinić, M.; Tišma, M.; Bucić-Kojić, A. Role of the Encapsulation in Bioavailability of Phenolic Compounds. Antioxidants 2020, 9, 923. [Google Scholar] [CrossRef]
- Del Rio, D.; Rodriguez-Mateos, A.; Spencer, J.P.; Tognolini, M.; Borges, G.; Crozier, A. Dietary (poly)phenolics in human health: Structures, bioavailability, and evidence of protective effects against chronic diseases. Antioxid Redox Signal 2013, 18, 1818–1892. [Google Scholar] [CrossRef] [PubMed]
- Cipolletti, M.; Solar Fernandez, V.; Montalesi, E.; Marino, M.; Fiocchetti, M. Beyond the Antioxidant Activity of Dietary Polyphenols in Cancer: The Modulation of Estrogen Receptors (ERs) Signaling. Int. J. Mol. Sci. 2018, 19, 2624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sesink, A.L.; O’Leary, K.A.; Hollman, P.C. Quercetin glucuronides but not glucosides are present in human plasma after consumption of quercetin-3-glucoside or quercetin-4′-glucoside. J. Nutr. 2001, 131, 1938–1941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Setchell, K.D.; Brown, N.M.; Lydeking-Olsen, E. The clinical importance of the metabolite equol-a clue to the Effectiveness of soy and its isoflavones. J. Nutr. 2002, 132, 3577–3584. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.M.; Rahaman, M.S.; Islam, M.R.; Rahman, F.; Mithi, F.M.; Alqahtani, T.; Almikhlafi, M.A.; Alghamdi, S.Q.; Alruwaili, A.S.; Hossain, M.S.; et al. Role of Phenolic Compounds in Human Disease: Current Knowledge and Future Prospects. Molecules 2021, 27, 233. [Google Scholar] [CrossRef]
- Felgines, C.; Talavera, S.; Gonthier, M.P.; Texier, O.; Scalbert, A.; Lamaison, J.L.; Remesy, C. Strawberry anthocyanins are recovered in urine as glucuro- and sulfoconjugates in humans. J. Nutr. 2003, 133, 1296–1301. [Google Scholar] [CrossRef] [Green Version]
- Crespy, V.; Morand, C.; Besson, C.; Manach, C.; Demigne, C.; Remesy, C. Quercetin, but not Its Glycosides, Is Absorbed from the Rat Stomach. J. Agric. Food Chem. 2002, 50, 618–621. [Google Scholar] [CrossRef]
- Filesi, C.; Giovannini, C.; Masella, R. Polyphenols, dietary sources and bioavailability. Ann. Ist. Super. Sanita 2007, 43, 348–361. [Google Scholar]
- Passamonti, S.; Vrhovsek, U.; Vanzo, A.; Mattivi, F. Fast Access of Some Grape Pigments to the Brain. J. Agric. Food Chem. 2005, 53, 7029–7034. [Google Scholar] [CrossRef]
- Halliwell, B.; Zhao, K.; Whiteman, M. The gastrointestinal tract: A major site of antioxidant action? Free Radic. Res. 2000, 33, 819–830. [Google Scholar] [CrossRef] [PubMed]
- Clifford, M.N. Chlorogenic acids and other cinnamates–nature, occurrence and dietary burden. J. Sci. Food Agric. 1999, 79, 362–372. [Google Scholar] [CrossRef]
- Kuhnau, J. Flavonoids. A class of semi-essential food components: Their role in human nutrition. World Rev. Nutr. Diet. 1976, 24, 117–191. [Google Scholar] [PubMed]
- Nunes, M.C.; Graça, C.; Vlaisavljević, S.; Tenreiro, A.; Sousa, I.; Raymundo, A. Microalgal cell disruption: Effect on the bioactivity and rheology of wheat bread. Algal Res. 2020, 45, 101749. [Google Scholar] [CrossRef]
- Bohn, T.; McDougall, G.J.; Alegría, A.; Alminger, M.; Arrigoni, E.; Aura, A.M.; Brito, C.; Cilla, A.; El, S.N.; Karakaya, S.; et al. Mind the gap-deficits in our knowledge of aspects impacting the bioavailability of phytochemicals and their metabolites—A position paper focusing on carotenoids and polyphenols. Mol. Nutr. Food Res. 2015, 59, 1307–1323. [Google Scholar] [CrossRef] [Green Version]
- Friedman, M.; Jürgens, H.S. Effect of pH on the stability of plant phenolic compounds. J. Agric. Food Chem. 2000, 48, 2101–2110. [Google Scholar] [CrossRef]
- Williams, A.W.; Boileau, T.W.M.; Erdman, J.W. Factors Influencing the Uptake and Absorption of Carotenoids. Proc. Soc. Exp. Biol. Med. 1998, 218, 106–108. [Google Scholar] [CrossRef]
- Zhu, F. Interactions between cell wall polysaccharides and polyphenols. Crit. Rev. Food Sci. Nutr. 2018, 58, 1808–1831. [Google Scholar] [CrossRef]
- Diep, T.T.; Yoo, M.J.Y.; Rush, E. Effect of In Vitro Gastrointestinal Digestion on Amino Acids, Polyphenols and Antioxidant Capacity of Tamarillo Yoghurts. Int. J. Mol. Sci. 2022, 23, 2526. [Google Scholar] [CrossRef]
- Palermo, M.; Pellegrini, N.; Fogliano, V. The effect of cooking on the phytochemical content of vegetables. J. Sci. Food Agric. 2014, 94, 1057–1070. [Google Scholar] [CrossRef]
- Dalmau, M.E.; Llabrés, P.J.; Eim, V.S.; Rosselló, C.; Simal, S. Influence of freezing on the bioaccessibility of beetroot (Beta vulgaris) bioactive compounds during in vitro gastric digestion. Sci. Food Agric. J. 2019, 99, 1055–1065. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, J.M.; Estevez, M.; Barba, F.J.; Thirumdas, R.; Franco, D.; Munekata, P.E.S. Polyphenols: Bioaccessibility and Bioavailability of Bioactive Components; Woodhead Publishing Series in Food Science; Elsevier: Amsterdam, The Netherlands, 2019; pp. 309–332. [Google Scholar]
- Barba, F.J.; Terefe, N.S.; Buckow, R.; Knorr, D.; Orlien, V. New opportunities and perspectives of high pressure treatment to improve health and safety attributes of foods. A review. Food Res. Int. 2015, 77, 725–742. [Google Scholar] [CrossRef]
- Ho, K.; Redan, B.W. Impact of thermal processing on the nutrients, phytochemicals, and metal contaminants in edible algae. Crit. Rev. Food Sci. Nutr. 2022, 62, 508–526. [Google Scholar] [CrossRef] [PubMed]
- Tomás-Barberán, F.A.; Gonzalez-Sarrias, A.; García-Villalba, R.; Núñez-Sánchez, M.Á.; Selma, M.V.; Garcia-Conesa, M.T.; Espín, J.C. Urolithins, the rescue of “old” metabolites to understand a “new” concept: Metabotypes as a nexus among phenolic metabolism, microbiota dysbiosis, and host health status. Mol. Nutr. Food Res. 2017, 61, 1500901. [Google Scholar] [CrossRef]
- Bode, L.M.; Bunzel, D.; Huch, M.; Cho, G.S.; Ruhland, D.; Bunzel, M.; Bub, A.; Franz, C.M.; Kulling, S.E. In vivo and in vitro metabolism of trans-resveratrol by human gut microbiota. Am. J. Clin. Nutr. 2013, 97, 295–309. [Google Scholar] [CrossRef] [Green Version]
- Filosa, S.; Di Meo, F.; Crispi, S. Polyphenols-gut microbiota interplay and brain neuromodulation. Neural Regen. Res. 2018, 13, 2055–2059. [Google Scholar] [CrossRef]
- Alves-Santos, A.M.; Sugizaki, C.S.A.; Lima, G.C.; Naves, M.M.V. Prebiotic effect of dietary polyphenols: A systematic review. J. Funct. Foods 2020, 74, 104169. [Google Scholar] [CrossRef]
- Moorthy, M.; Chaiyakunapruk, N.; Jacob, S.A.; Palanisamy, U.D. Prebiotic potential of polyphenols, its effect on gut microbiota and anthropometric/clinical markers: A systematic review of randomised controlled trials. Trends Food Sci. Technol. 2020, 99, 634–649. [Google Scholar] [CrossRef]
- Roberfroid, M.; Gibson, G.R.; Hoyles, L.; McCartney, A.L.; Rastall, R.; Rowland, I.; Wolvers, D.; Watzl, B.; Szajewska, H.; Stahl, B.; et al. Prebiotic effects: Metabolic and health benefits. Br. J. Nutr. 2010, 104 (Suppl. S2), S1–S63. [Google Scholar] [CrossRef] [Green Version]
- Morais, C.A.; de Rosso, V.V.; Estadella, D.; Pisani, L.P. Anthocyanins as inflammatory modulators and the role of the gut microbiota. J. Nutr. Biochem. 2016, 33, 1–7. [Google Scholar] [CrossRef]
- Wu, M.; Brown, A.C. Applications of Catechins in the Treatment of Bacterial Infections. Pathogens 2021, 10, 546. [Google Scholar] [CrossRef] [PubMed]
- Bouyahya, A.; Dakka, N.; Et-Touys, A.; Abrini, J.; Bakri, Y. Medicinal plant products targeting quorum sensing for combating bacterial infections. Asian Pac. J. Trop. Med. 2017, 10, 729–743. [Google Scholar] [CrossRef] [PubMed]
- Rothwell, J.A.; Medina-Remón, A.; Pérez-Jiménez, J.; Neveu, V.; Knaze, V.; Slimani, N.; Scalbert, A. Effects of food processing on polyphenol contents: A systematic analysis using Phenol-Explorer data. Mol. Nutr. Food Res. 2015, 59, 160–170. [Google Scholar] [CrossRef] [PubMed]
- Farooq, S.; Abdullah; Zhang, H.; Weiss, J. A comprehensive review on polarity, partitioning, and interactions of phenolic antioxidants at oil-water interface of food emulsions. Compr. Rev. Food Sci. Food Saf. 2021, 20, 4250–4277. [Google Scholar] [CrossRef] [PubMed]
- Igor Otavio, M.; Cristine Vanz, B.; Maria Izabela, F.; Hector Alonzo Gomez, G.; Chung-Yen Oliver, C.; Giuseppina Pace Pereira, L. Phenolic Compounds: Functional Properties, Impact of Processing and Bioavailability. In Phenolic Compounds; Marcos, S.-H., Mariana, P.-T., Maria del Rosario, G.-M., Eds.; IntechOpen: Rijeka, Croatia, 2017; p. Ch. 1. [Google Scholar]
- Gil-Martín, E.; Forbes-Hernández, T.; Romero, A.; Cianciosi, D.; Giampieri, F.; Battino, M. Influence of the extraction method on the recovery of bioactive phenolic compounds from food industry by-products. Food Chem. 2022, 378, 131918. [Google Scholar] [CrossRef]
- Wołosiak, R.; Drużyńska, B.; Piecyk, M.; Majewska, E.; Worobiej, E. Effect of Sterilization Process and Storage on the Antioxidative Properties of Runner Bean. Molecules 2018, 23, 1409. [Google Scholar] [CrossRef] [Green Version]
- de Lima, A.C.S.; da Rocha Viana, J.D.; de Sousa Sabino, L.B.; da Silva, L.M.R.; da Silva, N.K.V.; de Sousa, P.H.M. Processing of three different cooking methods of cassava: Effects on in vitro bioaccessibility of phenolic compounds and antioxidant activity. LWT—Food Sci. Technol. 2017, 76, 253–258. [Google Scholar] [CrossRef]
- Kurilich, A.C.; Clevidence, B.A.; Britz, S.J.; Simon, P.W.; Novotny, J.A. Plasma and urine responses are lower for acylated vs nonacylated anthocyanins from raw and cooked purple carrots. J. Agric. Food Chem. 2005, 53, 6537–6542. [Google Scholar] [CrossRef]
- Wani, S.M.; Masoodi, F.A.; Yousuf, S.; Dar, B.N.; Rather, S.A. Phenolic compounds and antiproliferative activity of apricots: Influence of canning, freezing, and drying. J. Food Process. Preserv. 2020, 44, e14887. [Google Scholar] [CrossRef]
- Chaovanalikit, A.; Wrolstad, R.E. Anthocyanin and Polyphenolic Composition of Fresh and Processed Cherries. J. Food Sci. 2004, 69, FCT73–FCT83. [Google Scholar] [CrossRef]
- Arfaoui, L. Dietary Plant Polyphenols: Effects of Food Processing on Their Content and Bioavailability. Molecules 2021, 26, 2959. [Google Scholar] [CrossRef] [PubMed]
- Kamiloglu, S.; Demirci, M.; Selen, S.; Toydemir, G.; Boyacioglu, D.; Capanoglu, E. Home processing of tomatoes (Solanum lycopersicum): Effects on in vitro bioaccessibility of total lycopene, phenolics, flavonoids, and antioxidant capacity. J. Sci. Food Agric. 2014, 94, 2225–2233. [Google Scholar] [CrossRef] [PubMed]
- Kamiloglu, S.; Capanoglu, E. Investigating the in vitro bioaccessibility of polyphenols in fresh and sun-dried figs (Ficus carica L.). Int. J. Food Sci. Technol. 2013, 48, 2621–2629. [Google Scholar] [CrossRef]
- Aydin, E.; Gocmen, D. The influences of drying method and metabisulfite pretreatment on the color, functional properties and phenolic acids contents and bioaccessibility of pumpkin flour. LWT—Food Sci. Technol. 2015, 60, 385–392. [Google Scholar] [CrossRef]
- Yanat, M.; Baysal, T. Effect of freezing rate and storage time on quality parameters of strawberry frozen in modified and home type freezer. Hrvat. Časopis Za Prehrambenu Tehnol. Biotehnol. I Nutr. 2018, 13, 154–158. [Google Scholar] [CrossRef] [Green Version]
- Rafiq, S.; Kaul, R.; Sofi, S.A.; Bashir, N.; Nazir, F.; Ahmad Nayik, G. Citrus peel as a source of functional ingredient: A review. J. Saudi Soc. Agric. Sci. 2018, 17, 351–358. [Google Scholar] [CrossRef] [Green Version]
- Derossi, A.; Ricci, I.; Caporizzi, R.; Fiore, A.; Severini, C. How grinding level and brewing method (Espresso, American, Turkish) could affect the antioxidant activity and bioactive compounds in a coffee cup. J. Sci. Food Agric. 2018, 98, 3198–3207. [Google Scholar] [CrossRef]
- Li, S.; Zhang, R.; Lei, D.; Huang, Y.; Cheng, S.; Zhu, Z.; Wu, Z.; Cravotto, G. Impact of ultrasound, microwaves and high-pressure processing on food components and their interactions. Trends Food Sci. Technol. 2021, 109, 1–15. [Google Scholar] [CrossRef]
- Leyane, T.S.; Jere, S.W.; Houreld, N.N. Oxidative Stress in Ageing and Chronic Degenerative Pathologies: Molecular Mechanisms Involved in Counteracting Oxidative Stress and Chronic Inflammation. Int. J. Mol. Sci. 2022, 23, 7273. [Google Scholar] [CrossRef]
- Gulcin, İ. Antioxidants and antioxidant methods: An updated overview. Arch. Toxicol. 2020, 94, 651–715. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Si, H.; Jia, Z.; Liu, D. Dietary Anti-Aging Polyphenols and Potential Mechanisms. Antioxidants 2021, 10, 283. [Google Scholar] [CrossRef] [PubMed]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free Radic. Biol. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef] [PubMed]
- Chalas, J.; Claise, C.; Edeas, M.; Messaoudi, C.; Vergnes, L.; Abella, A.; Lindenbaum, A. Effect of ethyl esterification of phenolic acids on low-density lipoprotein oxidation. Biomed. Pharmacother. 2001, 55, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Mumper, R.J. Plant Phenolics: Extraction, Analysis and Their Antioxidant and Anticancer Properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef] [PubMed]
- Gandhy, S.U.; Kim, K.; Larsen, L.; Rosengren, R.J.; Safe, S. Curcumin and synthetic analogs induce reactive oxygen species and decreases specificity protein (Sp) transcription factors by targeting microRNAs. BMC Cancer 2012, 12, 564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urbich, C.; Kuehbacher, A.; Dimmeler, S. Role of microRNAs in vascular diseases, inflammation, and angiogenesis. Cardiovasc. Res. 2008, 79, 581–588. [Google Scholar] [CrossRef] [Green Version]
- Cheng, H.S.; Sivachandran, N.; Lau, A.; Boudreau, E.; Zhao, J.L.; Baltimore, D.; Delgado-Olguin, P.; Cybulsky, M.; Fish, J.E. MicroRNA-146 represses endothelial activation by inhibiting pro-inflammatory pathways. EMBO Mol. Med. 2013, 5, 1017–1034. [Google Scholar] [CrossRef]
- Li, D.; Yang, P.; Xiong, Q.; Song, X.; Yang, X.; Liu, L.; Yuan, W.; Rui, Y.C. MicroRNA-125a/b-5p inhibits endothelin-1 expression in vascular endothelial cells. J. Hypertens. 2010, 28, 1646–1654. [Google Scholar] [CrossRef]
- Cione, E.; La Torre, C.; Cannataro, R.; Caroleo, M.C.; Plastina, P.; Gallelli, L. Quercetin, Epigallocatechin Gallate, Curcumin, and Resveratrol: From Dietary Sources to Human MicroRNA Modulation. Molecules 2019, 25, 63. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Tian, L.; Sun, D.; Yin, D. Curcumin ameliorates atherosclerosis through upregulation of miR-126. J. Cell. Physiol. 2019, 234, 21049–21059. [Google Scholar] [CrossRef]
- Tanno, M.; Kuno, A.; Yano, T.; Miura, T.; Hisahara, S.; Ishikawa, S.; Shimamoto, K.; Horio, Y. Induction of manganese superoxide dismutase by nuclear translocation and activation of SIRT1 promotes cell survival in chronic heart failure. J. Biol. Chem. 2010, 285, 8375–8382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.P.; Hsu, S.C.; Li, D.E.; Chen, K.H.; Kuo, C.Y.; Hung, L.M. Resveratrol Mitigates High-Fat Diet-Induced Vascular Dysfunction by Activating the Akt/eNOS/NO and Sirt1/ER Pathway. J. Cardiovasc. Pharmacol. 2018, 72, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Fourny, N.; Lan, C.; Sérée, E.; Bernard, M.; Desrois, M. Protective Effect of Resveratrol against Ischemia-Reperfusion Injury via Enhanced High Energy Compounds and eNOS-SIRT1 Expression in Type 2 Diabetic Female Rat Heart. Nutrients 2019, 11, 105. [Google Scholar] [CrossRef] [Green Version]
- Chung, D.J.; Wu, Y.L.; Yang, M.Y.; Chan, K.C.; Lee, H.J.; Wang, C.J. Nelumbo nucifera leaf polyphenol extract and gallic acid inhibit TNF-α-induced vascular smooth muscle cell proliferation and migration involving the regulation of miR-21, miR-143 and miR-145. Food Funct. 2020, 11, 8602–8611. [Google Scholar] [CrossRef] [PubMed]
- Shubina, V.S.; Kozina, V.I.; Shatalin, Y.V. Comparison of Antioxidant Properties of a Conjugate of Taxifolin with Glyoxylic Acid and Selected Flavonoids. Antioxidants 2021, 10, 1262. [Google Scholar] [CrossRef]
- Eghbaliferiz, S.; Iranshahi, M. Prooxidant Activity of Polyphenols, Flavonoids, Anthocyanins and Carotenoids: Updated Review of Mechanisms and Catalyzing Metals. Phytother Res. 2016, 30, 1379–1391. [Google Scholar] [CrossRef]
- Velderrain-Rodríguez, G.R.; Torres-Moreno, H.; Villegas-Ochoa, M.A.; Ayala-Zavala, J.F.; Robles-Zepeda, R.E.; Wall-Medrano, A.; González-Aguilar, G.A. Gallic Acid Content and an Antioxidant Mechanism Are Responsible for the Antiproliferative Activity of ‘Ataulfo’ Mango Peel on LS180 Cells. Molecules 2018, 23, 695. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Li, X.; Sang, S.; McClements, D.J.; Chen, L.; Long, J.; Jiao, A.; Jin, Z.; Qiu, C. Polyphenols as Plant-Based Nutraceuticals: Health Effects, Encapsulation, Nano-Delivery, and Application. Foods 2022, 11, 2189. [Google Scholar] [CrossRef]
- de la Lastra, C.A.; Villegas, I. Resveratrol as an anti-inflammatory and anti-aging agent: Mechanisms and clinical implications. Mol. Nutr. Food Res. 2005, 49, 405–430. [Google Scholar] [CrossRef]
- Lobo, A.; Liu, Y.; Song, Y.; Liu, S.; Zhang, R.; Liang, H.; Xin, H. Effect of procyanidins on lipid metabolism and inflammation in rats exposed to alcohol and iron. Heliyon 2020, 6, e04847. [Google Scholar] [CrossRef]
- Zhang, P.; Mak, J.C.; Man, R.Y.; Leung, S.W. Flavonoids reduces lipopolysaccharide-induced release of inflammatory mediators in human bronchial epithelial cells: Structure-activity relationship. Eur. J. Pharmacol. 2019, 865, 172731. [Google Scholar] [CrossRef]
- Liu, H.; Ma, S.; Xia, H.; Lou, H.; Zhu, F.; Sun, L. Anti-inflammatory activities and potential mechanisms of phenolic acids isolated from Salvia miltiorrhiza f. alba roots in THP-1 macrophages. J. Ethnopharmacol. 2018, 222, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.Y.; Li, Q.; Bi, K.S. Bioactive flavonoids in medicinal plants: Structure, activity and biological fate. Asian J. Pharm. Sci. 2018, 13, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Asif, P.J.; Longobardi, C.; Hahne, M.; Medema, J.P. The Role of Cancer-Associated Fibroblasts in Cancer Invasion and Metastasis. Cancers 2021, 13, 4720. [Google Scholar] [CrossRef]
- Kampa, M.; Nifli, A.P.; Notas, G.; Castanas, E. Polyphenols and cancer cell growth. Rev. Physiol. Biochem. Pharmacol. 2007, 159, 79–113. [Google Scholar] [CrossRef] [PubMed]
- Vaidya, F.U.; Chhipa, A.S.; Sagar, N.; Pathak, C. Oxidative Stress and Inflammation Can Fuel Cancer. In Role of Oxidative Stress in Pathophysiology of Diseases; Maurya, P.K., Dua, K., Eds.; Springer: Singapore, 2020; pp. 229–258. [Google Scholar]
- Oliveira-Marques, V.; Marinho, H.S.; Cyrne, L.; Antunes, F. Modulation of NF-kappaB-dependent gene expression by H2O2: A major role for a simple chemical process in a complex biological response. Antioxid. Redox Signal. 2009, 11, 2043–2053. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Norris, L.E.; Collene, A.L.; Asp, M.L.; Hsu, J.C.; Liu, L.F.; Richardson, J.R.; Li, D.; Bell, D.; Osei, K.; Jackson, R.D.; et al. Comparison of dietary conjugated linoleic acid with safflower oil on body composition in obese postmenopausal women with type 2 diabetes mellitus. Am. J. Clin. Nutr. 2009, 90, 468–476. [Google Scholar] [CrossRef] [Green Version]
- Shirakami, Y.; Sakai, H.; Kochi, T.; Seishima, M.; Shimizu, M. Catechins and Its Role in Chronic Diseases. Adv. Exp. Med. Biol. 2016, 929, 67–90. [Google Scholar]
- Miron, A.; Aprotosoaie, A.C.; Trifan, A.; Xiao, J. Flavonoids as modulators of metabolic enzymes and drug transporters. Ann. NY Acad. Sci. 2017, 1398, 152–167. [Google Scholar] [CrossRef]
- Guthrie, A.R.; Chow, H.H.S.; Martinez, J.A. Effects of resveratrol on drug- and carcinogen-metabolizing enzymes, implications for cancer prevention. Pharmacol. Res. Perspect. 2017, 5, e00294. [Google Scholar] [CrossRef]
- Truong, V.-L.; Jeong, W.-S. Cellular Defensive Mechanisms of Tea Polyphenols: Structure-Activity Relationship. Int. J. Mol. Sci. 2021, 22, 9109. [Google Scholar] [CrossRef] [PubMed]
- Abulnaja, K.; Bakkar, A.; Kannan, K.; Al-Manzlawi, A.M.; Kumosani, T.; Qari, M.; Moselhy, S. Olive leaf (Olea europaea L. folium) extract influences liver microsomal detoxifying enzymes in rats orally exposed to 2-amino-l-methyI-6-phenyI-imidazo pyridine (PhIP). Environ. Sci. Pollut. Res. 2022, 2022, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Majidinia, M.; Bishayee, A.; Yousefi, B. Polyphenols: Major regulators of key components of DNA damage response in cancer. DNA Repair 2019, 82, 102679. [Google Scholar] [CrossRef] [PubMed]
- Yin, F.; Giuliano, A.E.; Van Herle, A.J. Signal pathways involved in apigenin inhibition of growth and induction of apoptosis of human anaplastic thyroid cancer cells (ARO). Anticancer Res. 1999, 19, 4297–4303. [Google Scholar]
- Chen, C.; Yu, R.; Owuor, E.D.; Kong, A.N. Activation of antioxidant-response element (ARE), mitogen-activated protein kinases (MAPKs) and caspases by major green tea polyphenol components during cell survival and death. Arch. Pharmacal Res. 2000, 23, 605–612. [Google Scholar] [CrossRef]
- Yahfoufi, N.; Alsadi, N.; Jambi, M.; Matar, C. The Immunomodulatory and Anti-Inflammatory Role of Polyphenols. Nutrients 2018, 10, 1618. [Google Scholar] [CrossRef] [Green Version]
- Talib, W.H.; Abuawad, A.; Thiab, S.; Alshweiat, A.; Mahmod, A.I. Flavonoid-based nanomedicines to target tumor microenvironment. OpenNano 2022, 8, 100081. [Google Scholar] [CrossRef]
- Chen, H.J.; Lin, C.M.; Lee, C.Y.; Shih, N.C.; Peng, S.F.; Tsuzuki, M.; Amagaya, S.; Huang, W.W.; Yang, J.S. Kaempferol suppresses cell metastasis via inhibition of the ERK-p38-JNK and AP-1 signaling pathways in U-2 OS human osteosarcoma cells. Oncol. Rep. 2013, 30, 925–932. [Google Scholar] [CrossRef] [Green Version]
- Ke, C.; Gupta, R.; Xavier, D.; Prabhakaran, D.; Mathur, P.; Kalkonde, Y.V.; Kolpak, P.; Suraweera, W.; Jha, P. Divergent trends in ischaemic heart disease and stroke mortality in India from 2000 to 2015: A nationally representative mortality study. Lancet Glob. Health 2018, 6, e914–e923. [Google Scholar] [CrossRef] [Green Version]
- Koenig, W. Inflammation Revisited: Atherosclerosis in The Post-CANTOS Era. Eur. Cardiol. 2017, 12, 89–91. [Google Scholar] [CrossRef] [PubMed]
- Hurtubise, J.; McLellan, K.; Durr, K.; Onasanya, O.; Nwabuko, D.; Ndisang, J.F. The Different Facets of Dyslipidemia and Hypertension in Atherosclerosis. Curr. Atheroscler. Rep. 2016, 18, 82. [Google Scholar] [CrossRef] [PubMed]
- Zanotti, I.; Dall’Asta, M.; Mena, P.; Mele, L.; Bruni, R.; Ray, S.; Del Rio, D. Atheroprotective effects of (poly)phenols: A focus on cell cholesterol metabolism. Food Funct. 2015, 6, 13–31. [Google Scholar] [CrossRef] [PubMed]
- Elejalde, E.; Villarán, M.C.; Alonso, R.M. Grape polyphenols supplementation for exercise-induced oxidative stress. J. Int. Soc. Sport. Nutr. 2021, 18, 3. [Google Scholar] [CrossRef] [PubMed]
- Chambers, K.F.; Day, P.E.; Aboufarrag, H.T.; Kroon, P.A. Polyphenol Effects on Cholesterol Metabolism via Bile Acid Biosynthesis, CYP7A1: A Review. Nutrients 2019, 11, 2588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, D.; Wang, T.; Long, M.; Li, P. Quercetin: Its main pharmacological activity and potential application in clinical medicine. Oxidative Med. Cell. Longev. 2020, 2020, 8825387. [Google Scholar] [CrossRef]
- Potì, F.; Santi, D.; Spaggiari, G.; Zimetti, F.; Zanotti, I. Polyphenol Health Effects on Cardiovascular and Neurodegenerative Disorders: A Review and Meta-Analysis. Int. J. Mol. Sci. 2019, 20, 351. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, S. The Effect of Polyphenols on Hypercholesterolemia through Inhibiting the Transport and Expression of Niemann-Pick C1-Like 1. Int. J. Mol. Sci. 2019, 20, 4939. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Hao, Q.; Wang, Q.; Wang, Y.; Wan, X.; Zhou, Y. Supplementation with green tea extract affects lipid metabolism and egg yolk lipid composition in laying hens. J. Appl. Poult. Res. 2019, 28, 881–891. [Google Scholar] [CrossRef]
- Pang, J.; Xu, H.; Wang, X.; Chen, X.; Li, Q.; Liu, Q.; You, Y.; Zhang, H.; Xu, Z.; Zhao, Y.; et al. Resveratrol enhances trans-intestinal cholesterol excretion through selective activation of intestinal liver X receptor alpha. Biochem. Pharmacol. 2021, 186, 114481. [Google Scholar] [CrossRef]
- Azorín-Ortuño, M.; Yáñez-Gascón, M.J.; González-Sarrías, A.; Larrosa, M.; Vallejo, F.; Pallarés, F.J.; Lucas, R.; Morales, J.C.; Tomás-Barberán, F.A.; García-Conesa, M.T.; et al. Effects of long-term consumption of low doses of resveratrol on diet-induced mild hypercholesterolemia in pigs: A transcriptomic approach to disease prevention. J. Nutr. Biochem. 2012, 23, 829–837. [Google Scholar] [CrossRef] [PubMed]
- Mozos, I.; Flangea, C.; Vlad, D.C.; Gug, C.; Mozos, C.; Stoian, D.; Luca, C.T.; Horbańczuk, J.O.; Horbańczuk, O.K.; Atanasov, A.G. Effects of Anthocyanins on Vascular Health. Biomolecules 2021, 11, 811. [Google Scholar] [CrossRef] [PubMed]
- Sultana, R.; Alashi, A.M.; Islam, K.; Saifullah, M.; Haque, C.E.; Aluko, R.E. Inhibitory Activities of Polyphenolic Extracts of Bangladeshi Vegetables against α-Amylase, α-Glucosidase, Pancreatic Lipase, Renin, and Angiotensin-Converting Enzyme. Foods 2020, 9, 844. [Google Scholar] [CrossRef] [PubMed]
- Abbaszadeh, H.; Keikhaei, B.; Mottaghi, S. A review of molecular mechanisms involved in anticancer and antiangiogenic effects of natural polyphenolic compounds. Phytother. Res. 2019, 33, 2002–2014. [Google Scholar] [CrossRef]
- Yang, T.; Chen, Y.Y.; Liu, J.R.; Zhao, H.; Vaziri, N.D.; Guo, Y.; Zhao, Y.Y. Natural products against renin-angiotensin system for antifibrosis therapy. Eur. J. Med. Chem. 2019, 179, 623–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, T.; Xiao, D.; Muhammed, A.; Deng, J.; Chen, L.; He, J. Anti-Inflammatory Action and Mechanisms of Resveratrol. Molecules 2021, 26, 229. [Google Scholar] [CrossRef] [PubMed]
- DeFronzo, R.A.; Ferrannini, E.; Groop, L.; Henry, R.R.; Herman, W.H.; Holst, J.J.; Hu, F.B.; Kahn, C.R.; Raz, I.; Shulman, G.I.; et al. Type 2 diabetes mellitus. Nat. Rev. Dis. Prim. 2015, 1, 15019. [Google Scholar] [CrossRef] [PubMed]
- Bahadoran, Z.; Mirmiran, P.; Azizi, F. Dietary polyphenols as potential nutraceuticals in management of diabetes: A review. J Diabetes Metab Disord 2013, 12, 43. [Google Scholar] [CrossRef] [Green Version]
- Hanhineva, K.; Törrönen, R.; Bondia-Pons, I.; Pekkinen, J.; Kolehmainen, M.; Mykkänen, H.; Poutanen, K. Impact of Dietary Polyphenols on Carbohydrate Metabolism. Int. J. Mol. Sci. 2010, 11, 1365–1402. [Google Scholar] [CrossRef] [Green Version]
- Dao, T.M.; Waget, A.; Klopp, P.; Serino, M.; Vachoux, C.; Pechere, L.; Drucker, D.J.; Champion, S.; Barthélemy, S.; Barra, Y.; et al. Resveratrol increases glucose induced GLP-1 secretion in mice: A mechanism which contributes to the glycemic control. PLoS ONE 2011, 6, e20700. [Google Scholar] [CrossRef]
- Wang, Y.; Alkhalidy, H.; Liu, D. The Emerging Role of Polyphenols in the Management of Type 2 Diabetes. Molecules 2021, 26, 703. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wu, J.; Xu, F.; Chu, C.; Li, X.; Shi, X.; Zheng, W.; Wang, Z.; Jia, Y.; Xiao, W. Use of Ferulic Acid in the Management of Diabetes Mellitus and Its Complications. Molecules 2022, 27, 6010. [Google Scholar] [CrossRef] [PubMed]
- Markus, M.A.; Morris, B.J. Resveratrol in prevention and treatment of common clinical conditions of aging. Clin. Interv. Aging 2008, 3, 331–339. [Google Scholar]
- Aquilano, K.; Baldelli, S.; Rotilio, G.; Ciriolo, M.R. Role of nitric oxide synthases in Parkinson’s disease: A review on the antioxidant and anti-inflammatory activity of polyphenols. Neurochem. Res. 2008, 33, 2416–2426. [Google Scholar] [CrossRef]
- Nebrisi, E.E. Neuroprotective Activities of Curcumin in Parkinson’s Disease: A Review of the Literature. Int. J. Mol. Sci. 2021, 22, 11248. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, M.S.; Magalhães, M.C.; Oliveira, R.; Sousa-Lobo, J.M.; Almeida, I.F. Trends in the Use of Botanicals in Anti-Aging Cosmetics. Molecules 2021, 26, 3584. [Google Scholar] [CrossRef]
- Gasmi, A.; Mujawdiya, P.K.; Noor, S.; Lysiuk, R.; Darmohray, R.; Piscopo, S.; Lenchyk, L.; Antonyak, H.; Dehtiarova, K.; Shanaida, M.; et al. Polyphenols in Metabolic Diseases. Molecules 2022, 27, 6280. [Google Scholar] [CrossRef]
- Yang, X.H.; Li, L.; Xue, Y.B.; Zhou, X.X.; Tang, J.H. Flavonoids from Epimedium pubescens: Extraction and mechanism, antioxidant capacity and effects on CAT and GSH-Px of Drosophila melanogaster. PeerJ 2020, 8, e8361. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Liu, Y.; Ge, J.; Wang, X.; Liu, L.; Bu, Z.; Liu, P. Resveratrol protects human lens epithelial cells against H2O2-induced oxidative stress by increasing catalase, SOD-1, and HO-1 expression. Mol. Vis. 2010, 16, 1467–1474. [Google Scholar]
- Yang, Y.; Wu, Z.Z.; Cheng, Y.L.; Lin, W.; Qu, C. Resveratrol protects against oxidative damage of retinal pigment epithelium cells by modulating SOD/MDA activity and activating Bcl-2 expression. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 378–388. [Google Scholar] [CrossRef]
- Uygur, R.; Yagmurca, M.; Alkoc, O.A.; Genc, A.; Songur, A.; Ucok, K.; Ozen, O.A. Effects of quercetin and fish n-3 fatty acids on testicular injury induced by ethanol in rats. Andrologia 2014, 46, 356–369. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Sannigrahi, S.; Vaddepalli, R.P.; Ghosh, B.; Pusp, P. A novel combination of methotrexate and epigallocatechin attenuates the overexpression of proinflammatory cartilage cytokines and modulates antioxidant status in adjuvant arthritic rats. Inflammation 2012, 35, 1435–1447. [Google Scholar] [CrossRef] [PubMed]
- Hussain, Y.; Khan, H.; Alsharif, K.F.; Hayat Khan, A.; Aschner, M.; Saso, L. The Therapeutic Potential of Kaemferol and Other Naturally Occurring Polyphenols Might Be Modulated by Nrf2-ARE Signaling Pathway: Current Status and Future Direction. Molecules 2022, 27, 4145. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Yamamoto, M. Molecular mechanisms activating the Nrf2-Keap1 pathway of antioxidant gene regulation. Antioxid. Redox Signal. 2005, 7, 385–394. [Google Scholar] [CrossRef]
- Kim, J.Y.; Park, Y.K.; Lee, K.P.; Lee, S.M.; Kang, T.W.; Kim, H.J.; Dho, S.H.; Kim, S.Y.; Kwon, K.S. Genome-wide profiling of the microRNA-mRNA regulatory network in skeletal muscle with aging. Aging 2014, 6, 524–544. [Google Scholar] [CrossRef] [Green Version]
- Hoang, H.T.; Moon, J.-Y.; Lee, Y.-C. Natural Antioxidants from Plant Extracts in Skincare Cosmetics: Recent Applications, Challenges and Perspectives. Cosmetics 2021, 8, 106. [Google Scholar] [CrossRef]
- Xue, N.; Liu, Y.; Jin, J.; Ji, M.; Chen, X. Chlorogenic Acid Prevents UVA-Induced Skin Photoaging through Regulating Collagen Metabolism and Apoptosis in Human Dermal Fibroblasts. Int. J. Mol. Sci. 2022, 23, 6941. [Google Scholar] [CrossRef]
- Oršolić, N. Allergic Inflammation: Effect of Propolis and Its Flavonoids. Molecules 2022, 27, 6694. [Google Scholar] [CrossRef]
- Shakoor, H.; Feehan, J.; Apostolopoulos, V.; Platat, C.; Al Dhaheri, A.S.; Ali, H.I.; Ismail, L.C.; Bosevski, M.; Stojanovska, L. Immunomodulatory Effects of Dietary Polyphenols. Nutrients 2021, 13, 728. [Google Scholar] [CrossRef]
- Loo, Y.T.; Howell, K.; Chan, M.; Zhang, P.; Ng, K. Modulation of the human gut microbiota by phenolics and phenolic fiber-rich foods. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1268–1298. [Google Scholar] [CrossRef]
- Lee, J.S.; Lee, M.S.; An, S.; Yang, K.; Lee, K.; Yang, H.S.; Lee, H.; Cho, S.-W. Plant Flavonoid-Mediated Multifunctional Surface Modification Chemistry: Catechin Coating for Enhanced Osteogenesis of Human Stem Cells. Chem. Mater. 2017, 29, 4375–4384. [Google Scholar] [CrossRef]
- Yu, T.; Wang, Z.; You, X.; Zhou, H.; He, W.; Li, B.; Xia, J.; Zhu, H.; Zhao, Y.; Yu, G.; et al. Resveratrol promotes osteogenesis and alleviates osteoporosis by inhibiting p53. Aging 2020, 12, 10359–10369. [Google Scholar] [CrossRef] [PubMed]
- Bellavia, D.; Caradonna, F.; Dimarco, E.; Costa, V.; Carina, V.; De Luca, A.; Raimondi, L.; Fini, M.; Gentile, C.; Giavaresi, G. Non-flavonoid polyphenols in osteoporosis: Preclinical evidence. Trends Endocrinol. Metab. 2021, 32, 515–529. [Google Scholar] [CrossRef] [PubMed]
- Tomadoni, B.; Viacava, G.; Cassani, L.; Moreira, M.R.; Ponce, A. Novel biopreservatives to enhance the safety and quality of strawberry juice. J. Food Sci. Technol. 2016, 53, 281–292. [Google Scholar] [CrossRef] [Green Version]
- Dini, I. Chapter 14—Use of Essential Oils in Food Packaging. In Essential Oils in Food Preservation, Flavor and Safety; Preedy, V.R., Ed.; Academic Press: San Diego, CA, USA, 2016; pp. 139–147. [Google Scholar]
- Gupta, P.D.; Birdi, T.J. Development of botanicals to combat antibiotic resistance. J. Ayurveda Integr. Med. 2017, 8, 266–275. [Google Scholar] [CrossRef]
- Olszewska, M.A.; Gędas, A.; Simões, M. Antimicrobial polyphenol-rich extracts: Applications and limitations in the food industry. Food Res. Int. 2020, 134, 109214. [Google Scholar] [CrossRef]
- Villalobos-Delgado, L.H.; Nevárez-Moorillon, G.V.; Caro, I.; Quinto, E.J.; Mateo, J. 4—Natural antimicrobial agents to improve foods shelf life. In Food Quality and Shelf Life; Galanakis, C.M., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 125–157. [Google Scholar]
- Martinengo, P.; Arunachalam, K.; Shi, C. Polyphenolic Antibacterials for Food Preservation: Review, Challenges, and Current Applications. Foods 2021, 10, 2469. [Google Scholar] [CrossRef]
- Bouarab-Chibane, L.; Forquet, V.; Lantéri, P.; Clément, Y.; Léonard-Akkari, L.; Oulahal, N.; Degraeve, P.; Bordes, C. Antibacterial Properties of Polyphenols: Characterization and QSAR (Quantitative Structure-Activity Relationship) Models. Front. Microbiol. 2019, 10, 829. [Google Scholar] [CrossRef] [Green Version]
- Dini, I.; Laneri, S. Spices, Condiments, Extra Virgin Olive Oil and Aromas as Not Only Flavorings, but Precious Allies for Our Wellbeing. Antioxidants 2021, 10, 868. [Google Scholar] [CrossRef]
- Dini, I. Contribution of Nanoscience Research in Antioxidants Delivery Used in Nutricosmetic Sector. Antioxidants 2022, 11, 563. [Google Scholar] [CrossRef]
- Dini, I. The Potential of Dietary Antioxidants. Antioxidants 2021, 10, 1752. [Google Scholar] [CrossRef]
- Cavallo, P.; Dini, I.; Sepe, I.; Galasso, G.; Fedele, F.L.; Sicari, A.; Bolletti Censi, S.; Gaspari, A.; Ritieni, A.; Lorito, M.; et al. An Innovative Olive Pâté with Nutraceutical Properties. Antioxidants 2020, 9, 581. [Google Scholar] [CrossRef] [PubMed]
- Dini, I.; Laneri, S. The New Challenge of Green Cosmetics: Natural Food Ingredients for Cosmetic Formulations. Molecules 2021, 26, 3921. [Google Scholar] [CrossRef] [PubMed]
- Dini, I.; Laneri, S. Nutricosmetics: A brief overview. Phytother. Res. 2019, 33, 3054–3063.Dini, I. Bio Discarded from Waste to Resource. Foods 2021, 10, 2652. [Google Scholar] [CrossRef]
- Fanelli, F.; Cozzi, G.; Raiola, A.; Dini, I.; Mulè, G.; Logrieco, A.F.; Ritieni, A. Raisins and Currants as Conventional Nutraceuticals in Italian Market: Natural Occurrence of Ochratoxin A. J. Food Sci. 2017, 82, 2306–2312. [Google Scholar] [CrossRef]
- Ajila, C.M.; Brar, S.K.; Verma, M.; Tyagi, R.D.; Godbout, S.; Valéro, J.R. Extraction and analysis of polyphenols: Recent trends. Crit. Rev. Biotechnol. 2011, 31, 227–249. [Google Scholar] [CrossRef]
- Sridhar, A.; Ponnuchamy, M.; Kumar, P.S.; Kapoor, A.; Vo, D.V.N.; Prabhakar, S. Techniques and Modeling of Polyphenol Extraction from Food: A Review. Environ. Chem. Lett. 2021, 19, 3409–3443. [Google Scholar] [CrossRef]
- Safdar, M.N.; Kausar, T.; Jabbar, S.; Mumtaz, A.; Ahad, K.; Saddozai, A.A. Extraction and quantification of polyphenols from kinnow (Citrus reticulate L.) peel using ultrasound and maceration techniques. J. Food Drug Anal. 2017, 25, 488–500. [Google Scholar] [CrossRef] [Green Version]
- Ćujić, N.; Šavikin, K.; Janković, T.; Pljevljakušić, D.; Zdunić, G.; Ibrić, S. Optimization of polyphenols extraction from dried chokeberry using maceration as traditional technique. Food Chem. 2016, 194, 135–142. [Google Scholar] [CrossRef]
- Sharma, K.; Ko, E.Y.; Assefa, A.D.; Ha, S.; Nile, S.H.; Lee, E.T.; Park, S.W. Temperature-dependent studies on the total phenolics, flavonoids, antioxidant activities, and sugar content in six onion varieties. J. Food Drug Anal. 2015, 23, 243–252. [Google Scholar] [CrossRef] [Green Version]
- Reynoso-Camacho, R.; Rodríguez-Villanueva, L.D.; Sotelo-González, A.M.; Ramos-Gómez, M.; Pérez-Ramírez, I.F. Citrus decoction by-product represents a rich source of carotenoid, phytosterol, extractable and non-extractable polyphenols. Food Chem. 2021, 350, 129239. [Google Scholar] [CrossRef] [PubMed]
- Moldovan, M.L.; Iurian, S.; Puscas, C.; Silaghi-Dumitrescu, R.; Hanganu, D.; Bogdan, C.; Vlase, L.; Oniga, I.; Benedec, D. A Design of Experiments Strategy to Enhance the Recovery of Polyphenolic Compounds from Vitis vinifera By-Products through Heat Reflux Extraction. Biomolecules 2019, 9, 529. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.W.; Lin, L.G.; Ye, W.C. Techniques for extraction and isolation of natural products: A comprehensive review. Chin. Med. 2018, 13, 20. [Google Scholar] [CrossRef] [Green Version]
- Mushtaq, M.; Sultana, B.; Akram, S.; Anwar, F.; Adnan, A.; Rizvi, S.S.H. Enzyme-assisted supercritical fluid extraction: An alternative and green technology for non-extractable polyphenols. Anal. Bioanal. Chem. 2017, 409, 3645–3655. [Google Scholar] [CrossRef] [PubMed]
- Siddeeg, A.; Faisal Manzoor, M.; Haseeb Ahmad, M.; Ahmad, N.; Ahmed, Z.; Kashif Iqbal Khan, M.; Aslam Maan, A.; Mahr-Un-Nisa; Zeng, X.-A.; Ammar, A.-F. Pulsed Electric Field-Assisted Ethanolic Extraction of Date Palm Fruits: Bioactive Compounds, Antioxidant Activity and Physicochemical Properties. Processes 2019, 7, 585. [Google Scholar] [CrossRef] [Green Version]
- da Rosa, G.S.; Vanga, S.K.; Gariepy, Y.; Raghavan, V. Comparison of microwave, ultrasonic and conventional techniques for extraction of bioactive compounds from olive leaves (Olea europaea L.). Innov. Food Sci. Emerg. Technol. 2019, 58, 102234. [Google Scholar] [CrossRef]
- Zagklis, D.P.; Paraskeva, C.A. Isolation of organic compounds with high added values from agro-industrial solid wastes. J. Environ. Manag. 2018, 216, 183–191. [Google Scholar] [CrossRef]
- Tapia-Quirós, P.; Montenegro-Landívar, M.F.; Reig, M.; Vecino, X.; Saurina, J.; Granados, M.; Cortina, J.L. Integration of membrane processes for the recovery and separation of polyphenols from winery and olive mill wastes using green solvent-based processing. J. Environ. Manag. 2022, 307, 114555. [Google Scholar] [CrossRef]
- Alara, O.R.; Abdurahman, N.H.; Ukaegbu, C.I. Soxhlet extraction pf phenolic compounds from Vernonia cinereal leaves and its antioxidant activity. J. Appl. Res. Med. Aromat. Plants 2018, 11, 12–17. [Google Scholar]
- Ma, Y.; Meng, A.; Liu, P.; Chen, Y.; Yuan, A.; Dai, Y.; Ye, K.; Yang, Y.; Wang, Y.; Li, Z. Reflux Extraction Optimization and Antioxidant Activity of Phenolic Compounds from Pleioblastus amarus (Keng) Shell. Molecules 2022, 27, 362. [Google Scholar] [CrossRef]
- Rodríguez, Ó.; Bona, S.; Stäbler, A.; Rodríguez-Turienzo, L. Ultrasound-Assisted Extraction of Polyphenols from Olive Pomace: Scale up from Laboratory to Pilot Scenario. Processes 2022, 10, 2481. [Google Scholar] [CrossRef]
- Alchera, F.; Ginepro, M.; Giacalone, G. Microwave-Assisted Extraction of Polyphenols from Blackcurrant By-Products and Possible Uses of the Extracts in Active Packaging. Foods 2022, 11, 2727. [Google Scholar] [CrossRef] [PubMed]
- Farías-Campomanes, A.M.; Rostagno, M.A.; Coaquira-Quispe, J.J.; Meireles, M.A.A. Supercritical fluid extraction of polyphenols from lees: Overall extraction curve, kinetic data and composition of the extracts. Bioresour. Bioprocess. 2015, 2, 45. [Google Scholar] [CrossRef] [Green Version]
- Yong, D.; Ting, J.; Jun, X. Circulating Polyphenols Extraction System with High-Voltage Electrical Discharge: Design and Performance Evaluation. Sustain. Chem. Eng. 2018, 6, 15402–15410. [Google Scholar]
- Lakka, A.; Bozinou, E.; Makris, D.P.; Lalas, S.I. Evaluation of Pulsed Electric Field Polyphenol Extraction from Vitis vinifera, Sideritis scardica and Crocus sativus. ChemEngineering 2021, 5, 25. [Google Scholar] [CrossRef]
- Heemann, A.C.W.; Heemann, R.; Kalegari, P.; Spier, M.R.; Santin, E. Enzyme-assisted extraction of polyphenols from green yerba mate. Braz. J. Food Technol. 2019, 22, e2017222. [Google Scholar] [CrossRef] [Green Version]
- Meini, M.-R.; Cabezudo, I.; Boschetti, C.E.; Romanini, D. Recovery of phenolic antioxidants from Syrah grape pomace through the optimization of an enzymatic extraction process. Food Chem. 2019, 283, 257–264. [Google Scholar] [CrossRef]
- Ciccone, L.; Piragine, E.; Brogi, S.; Camodeca, C.; Fucci, R.; Calderone, V.; Nencetti, S.; Martelli, A.; Orlandini, E. Resveratrol-like Compounds as SIRT1 Activators. Int. J. Mol. Sci. 2022, 23, 15105. [Google Scholar] [CrossRef]
- Rudrapal, M.; Khairnar, S.J.; Khan, J.; Dukhyil, A.B.; Ansari, M.A.; Alomary, M.N.; Alshabrmi, F.M.; Palai, S.; Deb, P.K.; Devi, R. Dietary Polyphenols and Their Role in Oxidative Stress-Induced Human Diseases: Insights into Protective Effects, Antioxidant Potentials and Mechanism(s) of Action. Front. Pharm. 2022, 13, 806470. [Google Scholar] [CrossRef]
- Pimentel-Moral, S.; Verardo, V.; Robert, P.; Segura-Carretero, A.; Martínez-Férez, A. 13—Nanoencapsulation strategies applied to maximize target delivery of intact polyphenols. In Encapsulations; Grumezescu, A.M., Ed.; Academic Press: Cambridge, MA, USA, 2016; pp. 559–595. [Google Scholar]
- Ivanov, M.; Novović, K.; Malešević, M.; Dinić, M.; Stojković, D.; Jovčić, B.; Soković, M. Polyphenols as Inhibitors of Antibiotic Resistant Bacteria-Mechanisms Underlying Rutin Interference with Bacterial Virulence. Pharmaceuticals 2022, 15, 385. [Google Scholar] [CrossRef]
- Acosta, E. Bioavailability of nanoparticles in nutrient and nutraceutical delivery. Curr. Opin. Colloid Interface Sci. 2009, 14, 3–15. [Google Scholar] [CrossRef]
- Wang, J.; Byrne, J.D.; Napier, M.E.; DeSimone, J.M. More Effective Nanomedicines through Particle Design. Small 2011, 7, 1919–1931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammadian, M.; Salami, M.; Momen, S.; Alavi, F.; Emam-Djomeh, Z. Fabrication of curcumin-loaded whey protein microgels: Structural properties, antioxidant activity, and in vitro release behavior. LWT 2019, 103, 94–100. [Google Scholar] [CrossRef]
- Caballero, S.; Li, Y.O.; McClements, D.J.; Davidov-Pardo, G. Encapsulation and delivery of bioactive citrus pomace polyphenols: A review. Crit. Rev. Food Sci. Nutr. 2022, 62, 8028–8044. [Google Scholar] [CrossRef] [PubMed]
- Pandey, K.B.; Rizvi, S.I. Role of red grape polyphenols as antidiabetic agents. Integr. Med. Res. 2014, 3, 119–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.-H.; Lin, J.-H.; Hong, Y.-S. Development of chitosan/poly-γ-glutamic acid/pluronic/curcumin nanoparticles in chitosan dressings for wound regeneration. J. Biomed. Mater. Res. Part B Appl. Biomater. 2017, 105, 81–90. [Google Scholar] [CrossRef]
- Raja, I.S.; Preeth, D.R.; Vedhanayagam, M.; Hyon, S.-H.; Lim, D.; Kim, B.; Rajalakshmi, S.; Han, D.-W. Polyphenols-loaded electrospun nanofibers in bone tissue engineering and regeneration. Biomater. Res. 2021, 25, 29. [Google Scholar] [CrossRef]
- Drago, E.; Campardelli, R.; Pettinato, M.; Perego, P. Innovations in Smart Packaging Concepts for Food: An Extensive Review. Foods 2020, 9, 1628. [Google Scholar] [CrossRef]
- García Ibarra, V.; Rodríguez Bernaldo de Quirós, A.; Paseiro Losada, P.; Sendón, R. Non-target analysis of intentionally and non intentionally added substances from plastic packaging materials and their migration into food simulants. Food Packag. Shelf Life 2019, 21, 100325. [Google Scholar] [CrossRef]
- Pilevar, Z.; Bahrami, A.; Beikzadeh, S.; Hosseini, H.; Jafari, S.M. Migration of styrene monomer from polystyrene packaging materials into foods: Characterization and safety evaluation. Trends Food Sci. Technol. 2019, 91, 248–261. [Google Scholar] [CrossRef]
- Yildirim, S.; Röcker, B.; Pettersen, M.K.; Nilsen-Nygaard, J.; Ayhan, Z.; Rutkaite, R.; Radusin, T.; Suminska, P.; Marcos, B.; Coma, V. Active Packaging Applications for Food. Compr. Rev. Food Sci. Food Saf. 2018, 17, 165–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, B.; Chatha, S.A.S.; Hussain, A.I.; Zia, K.M.; Akhtar, N. Recent advances on polysaccharides, lipids and protein based edible films and coatings: A review. Int. J. Biol. Macromol. 2018, 109, 1095–1107. [Google Scholar] [CrossRef] [PubMed]
- Fierascu, R.C.; Sieniawska, E.; Ortan, A.; Fierascu, I.; Xiao, J. Fruits By-Products—A Source of Valuable Active Principles. A Short Review. Front. Bioeng. Biotechnol. 2020, 8, 319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaouch, M.A.; Benvenuti, S. The Role of Fruit by-Products as Bioactive Compounds for Intestinal Health. Foods 2020, 9, 1716. [Google Scholar] [CrossRef]
- Vasile, C.; Baican, M. Progresses in Food Packaging, Food Quality, and Safety-Controlled-Release Antioxidant and/or Antimicrobial Packaging. Molecules 2021, 26, 1263. [Google Scholar] [CrossRef]
- Yong, H.; Liu, J. Active packaging films and edible coatings based on polyphenol-rich propolis extract: A review. Compr. Rev. Food Sci. Food Saf. 2021, 20, 2106–2145. [Google Scholar] [CrossRef]
- Latos-Brozio, M.; Masek, A. Effect of Impregnation of Biodegradable Polyesters with Polyphenols from Cistus linnaeus and Juglans regia Linnaeus Walnut Green Husk. Polymers 2019, 11, 669. [Google Scholar] [CrossRef] [Green Version]
- Shahbazi, Y.; Shavisi, N. A novel active food packaging film for shelf-life extension of minced beef meat. J. Food Saf. 2018, 38, e12569. [Google Scholar] [CrossRef]
- Mascheroni, E.; Guillard, V.; Nalin, F.; Mora, L.; Piergiovanni, L. Diffusivity of propolis compounds in Polylactic acid polymer for the development of anti-microbial packaging films. J. Food Eng. 2010, 98, 294–301. [Google Scholar] [CrossRef]
- Shao, P.; Niu, B.; Chen, H.; Sun, P. Fabrication and characterization of tea polyphenols loaded pullulan-CMC electrospun nanofiber for fruit preservation. Int. J. Boil. Macromol. 2018, 107, 1908–1914. [Google Scholar] [CrossRef]
Figure 1.
Chemical structures of the different polyphenol classes. The colors indicate the sub-class. The numbers indicate the positions in the nomenclature.
Figure 1.
Chemical structures of the different polyphenol classes. The colors indicate the sub-class. The numbers indicate the positions in the nomenclature.

Figure 2.
Reactions between lipids and phenols.

Figure 3.
The polyphenols’ effects on glucose homeostasis and insulin resistance.

Figure 4.
The polyphenols’ traditional and modern extraction methods. Blue arrows: traditional extraction methods; red arrows: modern methods; in the orange circle: techniques used to improve polyphenol management.
Figure 4.
The polyphenols’ traditional and modern extraction methods. Blue arrows: traditional extraction methods; red arrows: modern methods; in the orange circle: techniques used to improve polyphenol management.


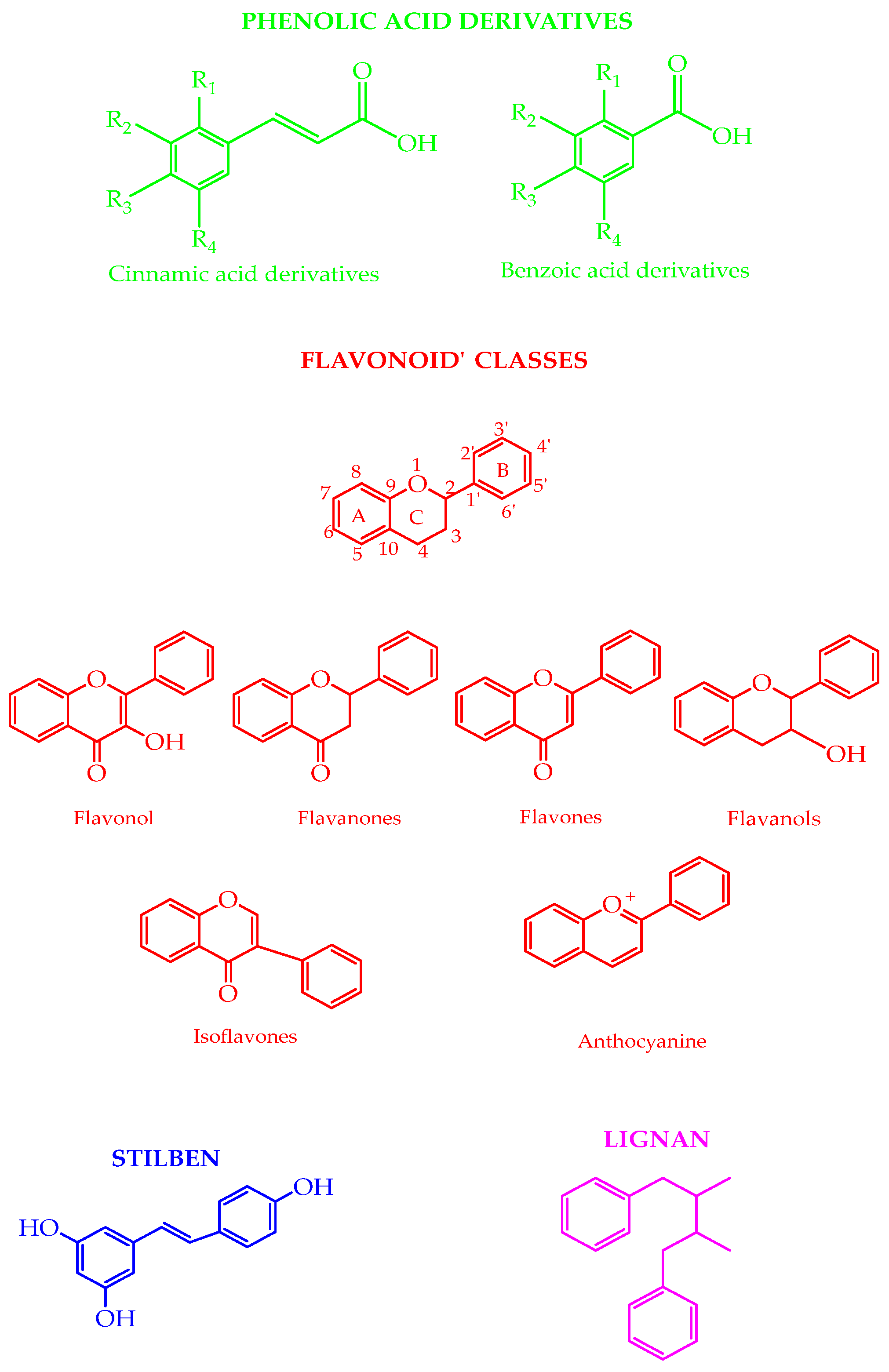{kind=link}
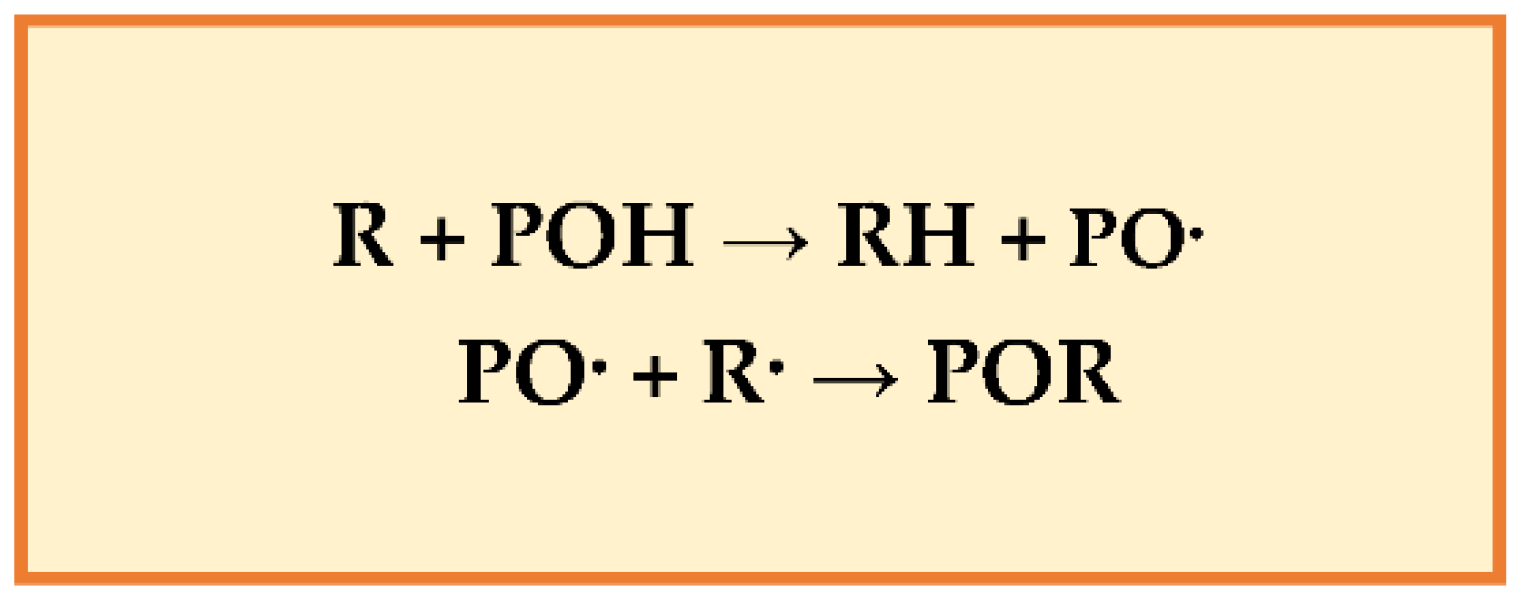{kind=link}
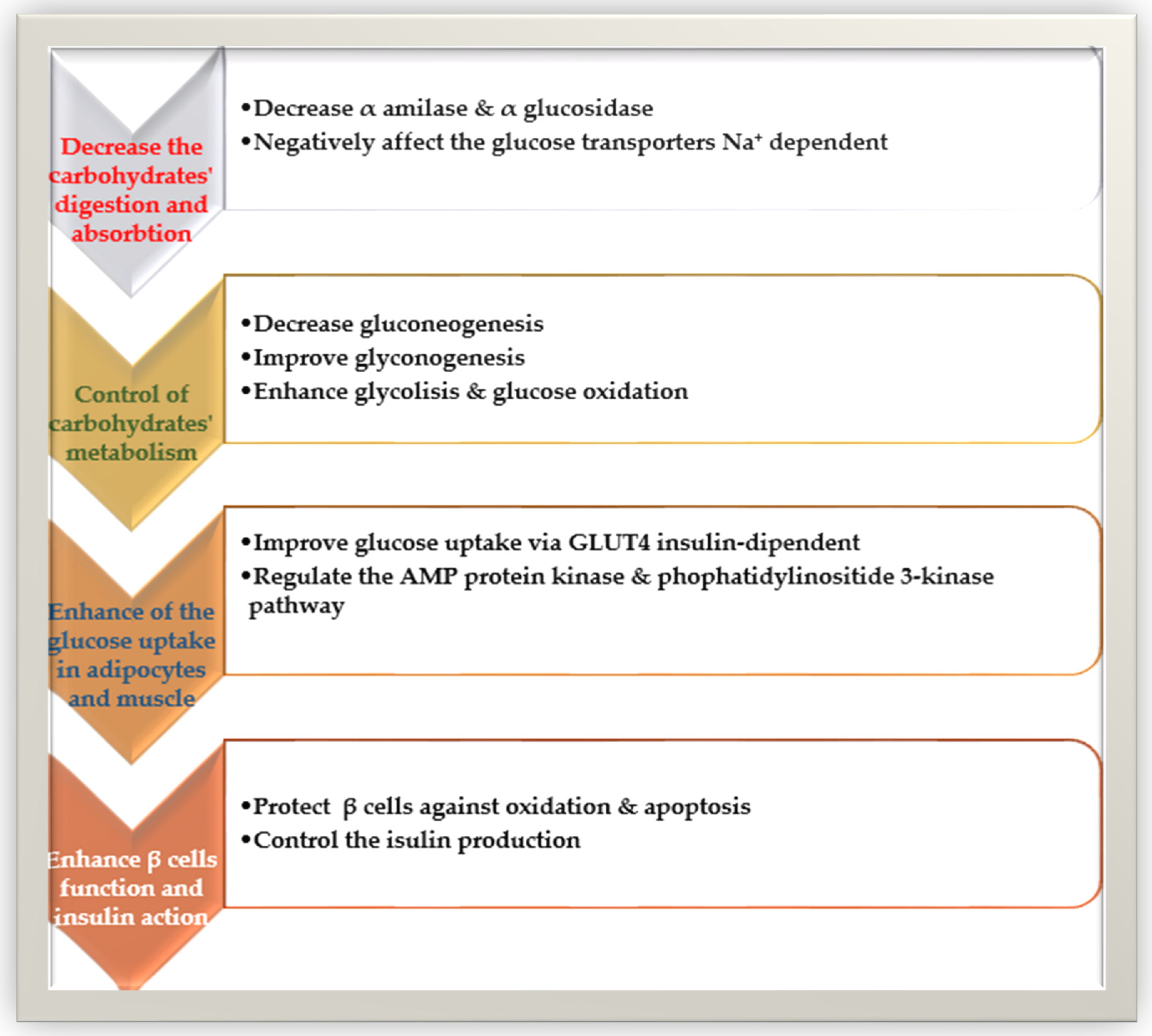{kind=link}
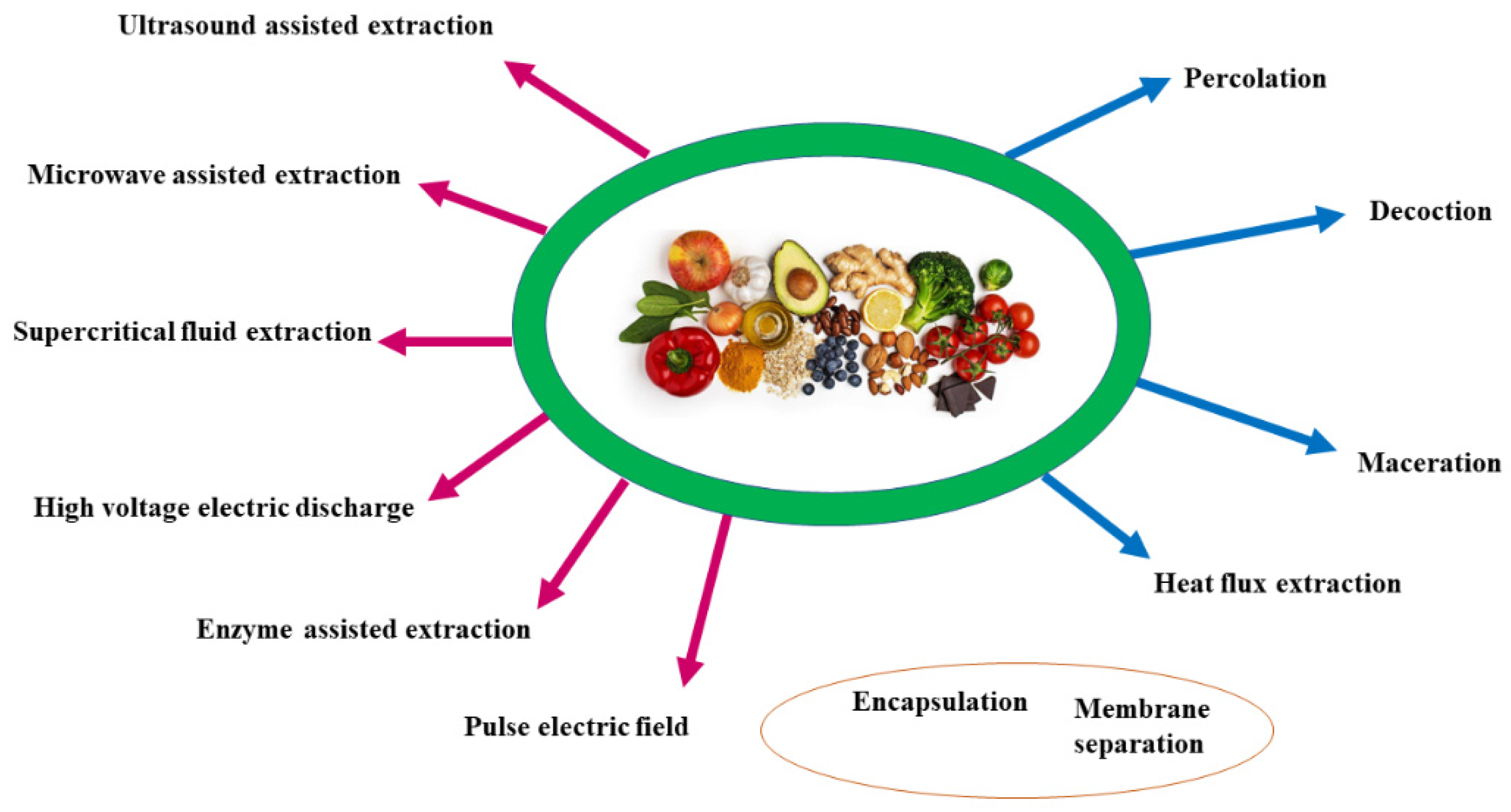{kind=link}
Table 1.
Extraction of free and bound polyphenols.
| Conventional Extraction Methods | Matrix | Extraction Solvent/Membrane Separation Type | Higher % of Recovery Compared to Traditional Method | Reference |
|---|---|---|---|---|
| Percolation | Vernonia cinerea leaves | Ethanol 60% | [188] | |
| Decoction | Citrus peels | Ethanol 75% | [180] | |
| Heat reflux extraction | Pleioblastus amarus | Ethanol 75% | [189] | |
| Maceration | Citrus peel | Ethanol 80% | [180] | |
| Non- Conventional methods | ||||
| Ultrasound assisted | Olive pomace | water | 58% | [190] |
| Microwave assisted | Blackcurrant By-Products | water | 25% | [191] |
| Supercritical fluid | Lees Vitis vinifera grapes | CO2 | −35% | [192] |
| High voltage electric discharge | Spent coffee grounds | 24% ethanol | 20.03% | [193] |
| Pulse electric field | Vitis vinifera, Sideritis scardica and Crocus sativus | water | 44.36–49.15% | [194] |
| Enzyme assisted | Green yerba mate | water | 38.67–52.08% | [195] |
| Membrane assisted pre-purification | Winery and olive mill wastes | NF270 membrane | 95% for polyphenol compounds removal | [196] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Dini, I.; Grumetto, L. Recent Advances in Natural Polyphenol Research. Molecules 2022, 27, 8777. https://doi.org/10.3390/molecules27248777
AMA Style
Dini I, Grumetto L. Recent Advances in Natural Polyphenol Research. Molecules. 2022; 27(24):8777. https://doi.org/10.3390/molecules27248777
Chicago/Turabian StyleDini, Irene, and Lucia Grumetto. 2022. "Recent Advances in Natural Polyphenol Research" Molecules 27, no. 24: 8777. https://doi.org/10.3390/molecules27248777