
Synthesis and Characterization of an Analgesic Potential Conotoxin Lv32.1


Abstract
:1. Introduction
2. Results
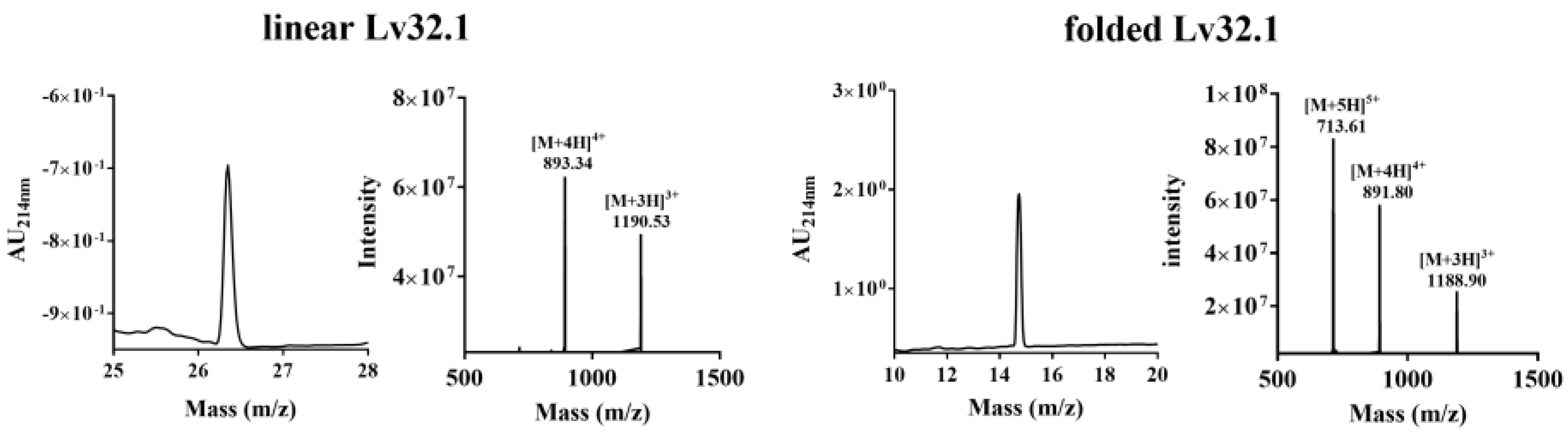
2.1. Peptide Synthesis and Oxidation Optimization
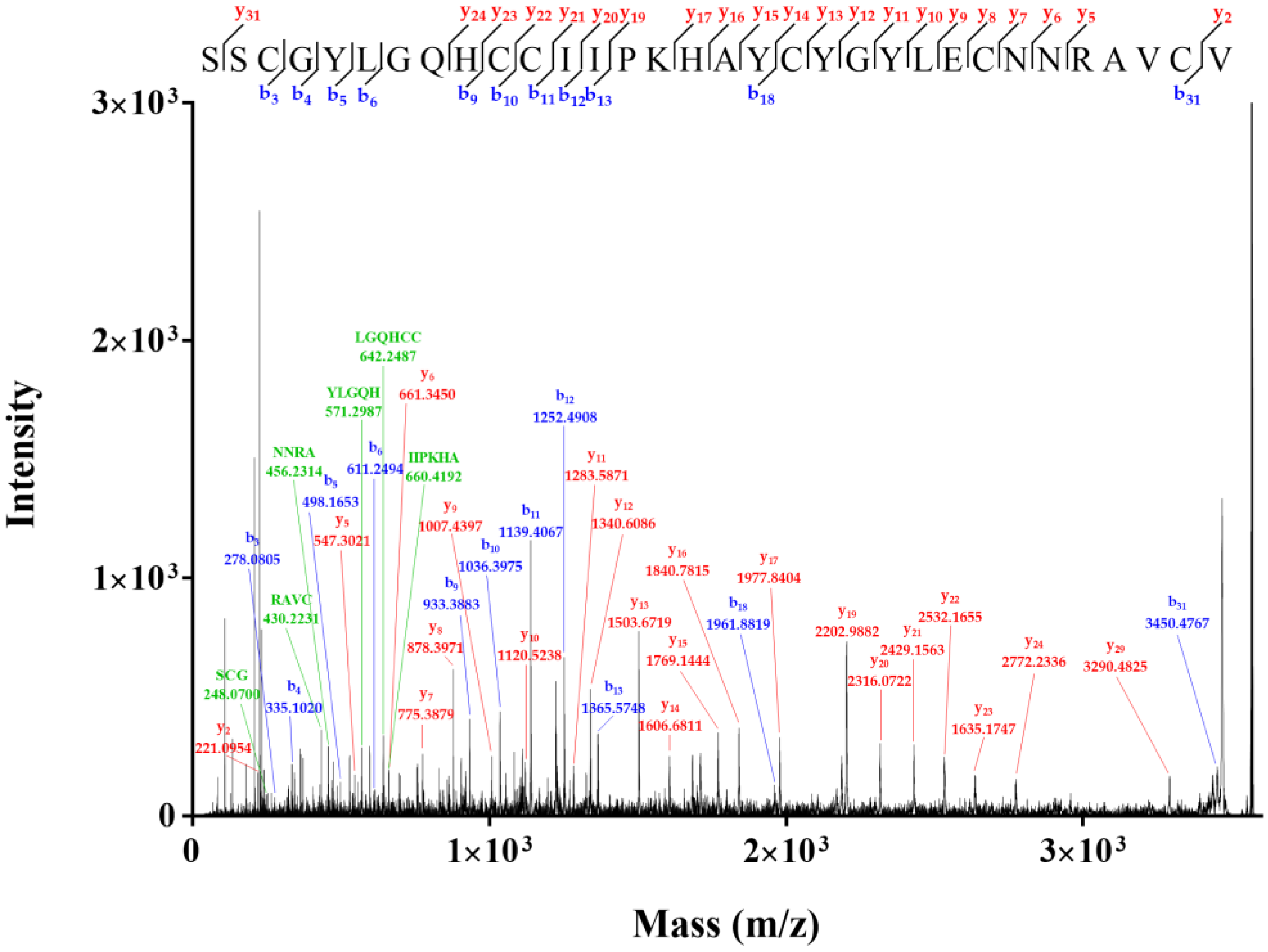
2.2. Novel Disulfide Connectivity of I–III, II–VI, IV–V
2.3. Lv32.1 Exhibited Analgesic Potency
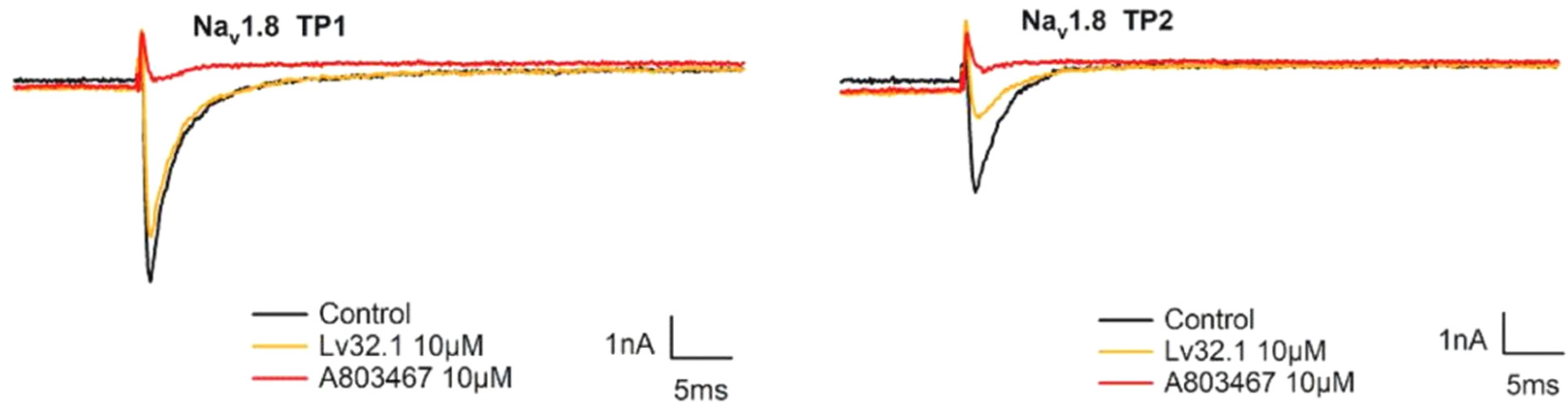
2.4. Weak Inhibition on Nav1.8 Channel and α9α10 nAChR
3. Discussion
4. Materials and Methods
4.1. Peptide Synthesis and Oxidation Optimization
4.2. Identification of Disulfide Connectivity
4.3. Analgesic Evaluation by Hot Plate Test in Mice
4.3.1. Animals
4.3.2. Intracerebroventricular (i.c.v.) Surgery
4.3.3. Pain Threshold Measurement by Hot Plate Test
4.3.4. Statistical Analysis
4.4. Whole-Cell Patch Clamp Measurement on Nav1.8 Channel
4.5. Two-Electrode Voltage Patch Clamp Recording on α9α10 nAChR
4.6. Data Analysis of Electrophysiological Assay on α9α10 nAChR and Nav1.8 Channel
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
Abbreviations
References
- Treede, R.D.; Rief, W.; Barke, A.; Aziz, Q.; Bennett, M.I.; Benoliel, R.; Cohen, M.; Evers, S.; Finnerup, N.B.; First, M.B.; et al. Chronic pain as a symptom or a disease: The IASP Classification of Chronic Pain for the International Classification of Diseases (ICD-11). Pain 2019, 160, 19–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mills, S.E.E.; Nicolson, K.P.; Smith, B.H. Chronic pain: A review of its epidemiology and associated factors in population-based studies. Br. J. Anaesth. 2019, 123, e273–e283. [Google Scholar] [CrossRef] [PubMed]
- Treede, R.D.; Rief, W.; Barke, A.; Aziz, Q.; Bennett, M.I.; Benoliel, R.; Cohen, M.; Evers, S.; Finnerup, N.B.; First, M.B.; et al. A classification of chronic pain for ICD-11. Pain 2015, 156, 1003–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsson, C.; Hansson, E.E.; Sundquist, K.; Jakobsson, U. Chronic pain in older adults: Prevalence, incidence, and risk factors. Scand J. Rheumatol. 2017, 46, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Gobina, I.; Villberg, J.; Valimaa, R.; Tynjala, J.; Whitehead, R.; Cosma, A.; Brooks, F.; Cavallo, F.; Ng, K.; de Matos, M.G.; et al. Prevalence of selfreported chronic pain among adolescents: Evidence from 42 countries and regions. Eur. J. Pain 2019, 23, 316–326. [Google Scholar] [CrossRef] [Green Version]
- Dahlhamer, J.; Lucas, J.; Zelaya, C.; Nahin, R.; Mackey, S.; DeBar, L.; Kerns, R.; Von Korff, M.; Porter, L.; Helmick, C. Prevalence of chronic pain and high-impact chronic pain among adults-United States, 2016. MMWR Morb. Mortal. Wkly. Rep. 2018, 67, 1001–1006. [Google Scholar] [CrossRef] [PubMed]
- Fayaz, A.; Croft, P.; Langford, R.M.; Donaldson, L.J.; Jones, G.T. Prevalence of chronic pain in the UK: A systematic review and meta-analysis of population studies. BMJ Open 2016, 6, e010364. [Google Scholar] [CrossRef]
- Hylands-White, N.; Duarte, R.V.; Raphael, J.H. An overview of treatment approaches for chronic pain management. Rheumatol. Int. 2017, 37, 29–42. [Google Scholar] [CrossRef] [Green Version]
- Schwan, J.; Sclafani, J.; Tawfik, V.L. Chronic Pain Management in the Elderly. Anesthesiol. Clin. 2019, 37, 547–560. [Google Scholar] [CrossRef] [PubMed]
- Kalso, E.; Edwards, J.E.; Moore, A.R.; McQuay, H.J. Opioids in chronic non-cancer pain: Systematic review of efficacy and safety. Pain 2004, 112, 372–380. [Google Scholar] [CrossRef]
- Eccleston, C.; Fisher, E.; Thomas, K.H.; Hearn, L.; Derry, S.; Stannard, C.; Knaggs, R.; Moore, R.A. Interventions for the reduction of prescribed opioid use in chronic non-cancer pain. Cochrane Database Syst. Rev. 2017, 11, CD010323. [Google Scholar]
- Ballantyne, J.C.; Mao, J. Opioid therapy for chronic pain. N. Engl. J. Med. 2003, 349, 1943–1953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yekkirala, A.S.; Roberson, D.P.; Bean, B.P.; Woolf, C.J. Breaking barriers to novel analgesic drug development. Nat. Rev. Drug Discov. 2017, 16, 545–564. [Google Scholar] [CrossRef] [Green Version]
- Obeng, S.; Hiranita, T.; Leon, F.; McMahon, L.R.; McCurdy, C.R. Novel approaches, drug candidates, and targets in pain drug discovery. J. Med. Chem. 2021, 64, 6523–6548. [Google Scholar] [CrossRef]
- Vetter, I.J.; Lewis, R. Therapeutic potential of cone snail venom peptides (conopeptides). Curr. Top Med. Chem. 2012, 12, 1546–1552. [Google Scholar] [CrossRef] [PubMed]
- Jin, A.H.; Muttenthaler, M.; Dutertre, S.; Himaya, S.W.A.; Kaas, Q.; Craik, D.J.; Lewis, R.J.; Alewood, P.F. Conotoxins: Chemistry and biology. Chem. Rev. 2019, 119, 11510–11549. [Google Scholar] [CrossRef]
- Lavergne, V.; Harliwong, I.; Jones, A.; Miller, D.; Taft, R.J.; Alewood, P.F. Optimized deep-targeted proteotranscriptomic profiling reveals unexplored Conus toxin diversity and novel cysteine frameworks. Proc. Natl. Acad. Sci. USA 2015, 112, E3782–E3791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, J.; Jones, A.; Lewis, R.J. Remarkable inter- and intra-species complexity of conotoxins revealed by LC/MS. Peptides 2009, 30, 1222–1227. [Google Scholar] [CrossRef] [PubMed]
- Sudewi, A.A.R.; Susilawathi, N.M.; Mahardika, B.K.; Mahendra, A.N.; Pharmawati, M.; Phuong, M.A.; Mahardika, G.N. Selecting potential neuronal drug leads from conotoxins of various venomous marine cone snails in Bali, Indonesia. ACS Omega 2019, 4, 19483–19490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajaian Pushpabai, R.; Wilson Alphonse, C.R.; Mani, R.; Arun Apte, D.; Franklin, J.B. Diversity of conopeptides and conoenzymes from the venom duct of the marine cone snail Conus bayani as determined from transcriptomic and proteomic analyses. Mar. Drugs 2021, 19, 202. [Google Scholar] [CrossRef]
- Li, X.; Chen, W.; Zhangsun, D.; Luo, S. Diversity of conopeptides and their precursor genes of Conus litteratus. Mar. Drugs 2020, 18, 464. [Google Scholar] [CrossRef] [PubMed]
- Kancherla, A.K.; Meesala, S.; Jorwal, P.; Palanisamy, R.; Sikdar, S.K.; Sarma, S.P. A disulfide stabilized beta-sandwich defines the structure of a new cysteine framework M-superfamily conotoxin. ACS Chem. Biol. 2015, 10, 1847–1860. [Google Scholar] [CrossRef] [PubMed]
- Yao, G.; Peng, C.; Zhu, Y.; Fan, C.; Jiang, H.; Chen, J.; Cao, Y.; Shi, Q. High-throughput identification and analysis of novel conotoxins from three vermivorous cone snails by transcriptome sequencing. Mar. Drugs 2019, 17, 193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, C.; Yao, G.; Gao, B.M.; Fan, C.X.; Bian, C.; Wang, J.; Cao, Y.; Wen, B.; Zhu, Y.; Ruan, Z.; et al. High-throughput identification of novel conotoxins from the Chinese tubular cone snail (Conus betulinus) by multi-transcriptome sequencing. Gigascience 2016, 5, 17–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Xu, N.; Hu, J.; Zhao, C.; Yu, Z.; Dai, Q. Identification of novel I-superfamily conopeptides from several clades of Conus species found in the South China Sea. Peptides 2009, 30, 1782–1787. [Google Scholar] [CrossRef]
- Ju, S.; Zhang, Y.; Guo, X.; Yan, Q.; Liu, S.; Ma, B.; Zhang, M.; Bao, J.; Luo, S.; Fu, Y. Anti-ovarian cancer conotoxins identified from Conus venom. Molecules 2022, 27, 6609. [Google Scholar] [CrossRef] [PubMed]
- Ye, M.; Khoo, K.K.; Xu, S.; Zhou, M.; Boonyalai, N.; Perugini, M.A.; Shao, X.; Chi, C.; Galea, C.A.; Wang, C.; et al. A helical conotoxin from Conus imperialis has a novel cysteine framework and defines a new superfamily. J. Biol. Chem. 2012, 287, 14973–14983. [Google Scholar] [CrossRef] [Green Version]
- Kaas, Q.; Yu, R.; Jin, A.H.; Dutertre, S.; Craik, D.J. ConoServer: Updated content, knowledge, and discovery tools in the conopeptide database. Nucleic Acids Res. 2012, 40, D325–D330. [Google Scholar] [CrossRef]
- Favreau, P.; Stocklin, R.; Kamerling, J.P.; Boelens, R. Structure of the O-glycosylated conopeptide CcTx from Conus consors venom. Chem.-Eur. J. 2013, 19, 870–879. [Google Scholar]
- Akondi, K.B.; Muttenthaler, M.; Dutertre, S.; Kaas, Q.; Craik, D.J.; Lewis, R.J.; Alewood, P.F. Discovery, synthesis, and structureactivity relationships of conotoxins. Chem. Rev. 2014, 114, 5815–5847. [Google Scholar] [CrossRef]
- Kaas, Q.; Westermann, J.C.; Craik, D.J. Conopeptide characterization and classifications: An analysis using ConoServer. Toxicon 2010, 55, 1491–1509. [Google Scholar] [CrossRef]
- Buczek, O.; Wei, D.; Babon, J.J.; Yang, X.; Fiedler, B.; Chen, P.; Yoshikami, D.; Olivera, B.M.; Bulaj, G.; Norton, R.S. Structure and sodium channel activity of an excitatory I1-superfamily conotoxin. Biochemistry 2007, 46, 9929–9940. [Google Scholar] [CrossRef] [PubMed]
- Du, W.H.; Han, Y.H.; Huang, F.J.; Li, J.; Chi, C.W.; Fang, W.H. Solution structure of an M-1 conotoxin with a novel disulfide linkage. FEBS J. 2007, 274, 2596–2602. [Google Scholar] [CrossRef]
- Himaya, S.W.; Jin, A.H.; Dutertre, S.; Giacomotto, J.; Mohialdeen, H.; Vetter, I.; Alewood, P.F.; Lewis, R.J. Comparative Venomics Reveals the Complex Prey Capture Strategy of the Piscivorous Cone Snail Conus catus. J. Proteome Res. 2015, 14, 4372–4381. [Google Scholar] [CrossRef]
- Franco, A.; Dovell, S.; Moller, C.; Grandal, M.; Clark, E.; Mari, F. Structural plasticity of mini-M conotoxins-expression of all miniM subtypes by Conus regius. FEBS J. 2018, 285, 887–902. [Google Scholar] [CrossRef] [Green Version]
- Corpuz, G.P.; Jacobsen, R.B.; Jimenez, E.C.; Watkins, M.; Walker, C.; Colledge, C.; Garrett, J.E.; McDougal, O.; Li, W.; Gray, W.R.; et al. Definition of the M-conotoxin superfamily: Characterization of novel peptides from molluscivorous Conus venoms. Biochemistry 2005, 44, 8176–8186. [Google Scholar] [CrossRef]
- Karl, T.; Pabst, R.; von Horsten, S. Behavioral phenotyping of mice in pharmacological and toxicological research. Exp. Toxicol. Pathol. 2003, 55, 69–83. [Google Scholar] [CrossRef]
- Luo, S.; Zhangsun, D.; Harvey, P.J.; Kaas, Q.; Wu, Y.; Zhu, X.; Hu, Y.; Li, X.; Tsetlin, V.I.; Christensen, S.; et al. Cloning, synthesis, and characterization of alphaO-conotoxin GeXIVA, a potent alpha9alpha10 nicotinic acetylcholine receptor antagonist. Proc. Natl. Acad. Sci. USA 2015, 112, E4026–E4035. [Google Scholar] [CrossRef] [Green Version]
- Xue, Y.; Chidiac, C.; Herault, Y.; Gaveriaux-Ruff, C. Pain behavior in SCN9A (Nav1.7) and SCN10A (Nav1.8) mutant rodent models. Neurosci. Lett. 2021, 753, 135844. [Google Scholar] [CrossRef]
- Ekberg, J.; Jayamanne, A.; Vaughan, C.W.; Aslan, S.; Thomas, L.; Mould, J.; Drinkwater, R.; Baker, M.D.; Abrahamsen, B.; Wood, J.N.; et al. μO-conotoxin MrVIB selectively blocks Nav1.8 sensory neuron specific sodium channels and chronic pain behavior without motor deficits. Proc. Natl. Acad. Sci. USA 2006, 103, 17030–17035. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Li, X.; Zhangsun, D.; Yu, G.; Su, R.; Luo, S. The alpha9alpha10 nicotinic acetylcholine receptor antagonist alphaO-Conotoxin GeXIVA [1,2] alleviates and reverses chemotherapy- Induced neuropathic pain. Mar. Drugs 2019, 17, 265. [Google Scholar] [CrossRef] [Green Version]
- Heimer, P.; Tietze, A.A.; Bäuml, C.A.; Resemann, A.; Mayer, F.J.; Suckau, D.; Ohlenschläger, O.; Tietze, D.; Imhof, D. Conformational μ-conotoxin PIIIA isomers revisited: Impact of cysteine pairing on disulfide-bond assignment and structure elucidation. Anal. Chem. 2018, 90, 3321–3327. [Google Scholar] [CrossRef]
- Zimmermann, M. Ethical guidelines for investigations of experimental pain in conscious animals. Pain 1983, 16, 109–110. [Google Scholar] [CrossRef]
- Weiser, T. Comparison of the effects of four Na+ channel analgesics on TTX-resistant Na+ currents in rat sensory neurons and recombinant Nav1.2 channels. Neurosci. Lett. 2006, 395, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Zhang, Y.; Ju, S.; Ma, B.; Huang, W.; Luo, S. Isolation and characterization of five novel disulfide-poor conopeptides from Conus marmoreus venom. J. Venom. Anim. Toxins Incl. Trop. Dis. 2022, 28, e20210116. [Google Scholar] [CrossRef]
- Xu, P.; Kaas, Q.; Wu, Y.; Zhu, X.; Li, X.; Harvey, P.J.; Zhangsun, D.; Craik, D.J.; Luo, S. Structure and Activity Studies of Disulfide-Deficient Analogues of alphaO-Conotoxin GeXIVA. J. Med. Chem. 2020, 63, 1564–1575. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Source | Sequence | Reference |
|---|---|---|---|
| Lv32.1/ QGV13653.1 | C. lividus | SSCGYLGQHCCIIPKHAYCYGYLECNNRAVCV | This work [19] |
| Ba32.1 | C. bayani | DCGERDEPCCVNSSGVKYCESPWSCMHTTLLCEQN | [20] |
| Lt32.1 | C. litteratus | SSCGYLGQQCCIVPKRAYCHGDLECNPVAMCVA | [21] |
| Mo3964 | C. monile | DGECGDKDEPCCGRPDGAKVCNDPWVCILTSSRCENP | [22] |
| Qc32.1 | C. quercinus | SSCGYLGQPCCVVPRRAYCHGDLECNDVTMCV | [23] |
| AMP44674.1 | C. betulinus | SSCGYVGQPCCIVPRRAYCHGDLNCNNVAMCV | [24] |
| ACV87167.1 | C. eburneus | SGCGYLGEPCCISPKRAYCHGDLECNNVAMCV | [25] |
| ACV87166.1 | C. marmoreus | SGCGYLGEPCCVAPKRAYCHGDLECNNVAMCV | [25] |
| ACV87168.1 | C. marmoreus | SGCGYLGEPCCVAPKRAYCHGDLECNSVAMCV | [25] |
| ACV87169.1 | C. litteratus | SGCGYLGEPCCVAPKRAYCHGDLECNNIAMCV | [25] |
| Disulfide Connectivity | Cysteine Framework | Reference |
|---|---|---|
| 1–2, 3–4, 5–6 | XXIII | [27] |
| 1–3, 2–5, 4–6 | IV, XXVII, XXXII | [22,28,29] |
| 1–4, 2–5, 3–6 | III, VI/VII | [30,31,32] |
| 1–6, 2–4, 3–5 | III | [33] |
| 1–5, 2–3, 4–6 | IV | [34] |
| 1–5, 2–4, 3–6 | III | [35,36] |
| 1–3, 2–6, 4–5 | XXXII | this work |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.; Li, C.; You, S.; Yan, Q.; Luo, S.; Fu, Y. Synthesis and Characterization of an Analgesic Potential Conotoxin Lv32.1. Molecules 2022, 27, 8617. https://doi.org/10.3390/molecules27238617
Liu S, Li C, You S, Yan Q, Luo S, Fu Y. Synthesis and Characterization of an Analgesic Potential Conotoxin Lv32.1. Molecules. 2022; 27(23):8617. https://doi.org/10.3390/molecules27238617
Chicago/Turabian StyleLiu, Siyi, Cheng Li, Shen You, Qinghui Yan, Sulan Luo, and Ying Fu. 2022. "Synthesis and Characterization of an Analgesic Potential Conotoxin Lv32.1" Molecules 27, no. 23: 8617. https://doi.org/10.3390/molecules27238617