

Glucosinolates of Sisymbrium officinale and S. orientale
 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Glucosinolates and Volatile Constituents
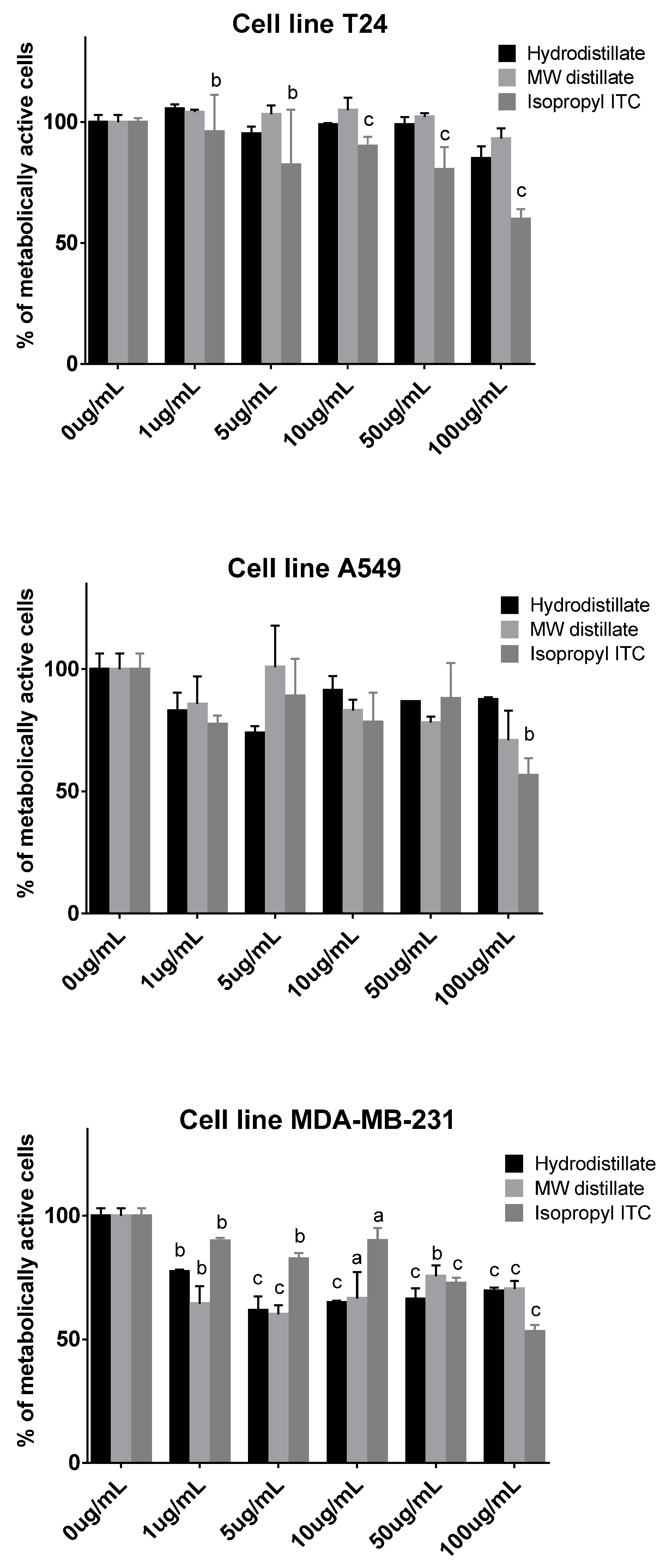
2.2. Antiproliferative Activity
3. Materials and Methods
3.1. Materials and Reagents
3.2. Isolation and Chemical Analysis
3.2.1. Isolation of Desulfoglucosinolates
3.2.2. UHPLC-MS/MS Analysis
3.2.3. Isolation of Volatiles
3.2.4. GC-MS Analysis
3.3. Cell Viability Assay (MTT)
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Blažević, I.; Montaut, S.; Burčul, F.; Olsen, C.E.; Burow, M.; Rollin, P.; Agerbirk, N. Glucosinolate structural diversity, identification, chemical synthesis and metabolism in plants. Phytochemistry 2020, 169, 112100. [Google Scholar] [CrossRef] [PubMed]
- Montaut, S.; Read, S.; Blažević, I.; Nuzillard, J.-M.; Roje, M.; Harakat, D.; Rollin, P. Investigation of the glucosinolates in Hesperis matronalis L. and Hesperis laciniata All.: Unveiling 4′-O-β-D-apiofuranosylglucomatronalin. Carbohydr. Res. 2020, 488, 107898. [Google Scholar] [CrossRef] [PubMed]
- Trabelcy, B.; Chinkov, N.; Samuni-Blank, M.; Merav, M.; Izhaki, I.; Carmeli, S.; Gerchman, Y. Investigation of glucosinolates in the desert plant Ochradenus baccatus (Brassicales: Resedaceae). Unveiling glucoochradenin, a new arabinosylated glucosinolate. Phytochemistry 2021, 187, 112760. [Google Scholar] [CrossRef]
- Sonderby, I.E.; Geuflores, F.; Halkier, B.A. Biosynthesis of glucosinolates—Gene discovery and beyond. Trends Plant Sci. 2010, 15, 283–290. [Google Scholar] [CrossRef]
- Harun, S.; Abdullah-Zawawi, M.-R.; Goh, H.-H.; Mohamed-Hussein, Z.-A. A comprehensive gene inventory for glucosinolate biosynthetic pathway in Arabidopsis thaliana. J. Agric. Food Chem. 2020, 68, 7281–7297. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wu, J.; Sun, S.; Liu, B.; Cheng, F.; Sun, R.; Wang, X. Glucosinolate biosynthetic genes in Brassica rapa. Gene 2011, 487, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Liu, Y.; Yang, X.; Tong, C.; Edwards, D.; Parkin, I.A.; Zhao, M.; Ma, J.; Yu, J.; Huang, S.; et al. The Brassica oleracea genome reveals the asymmetrical evolution of polyploidy genomes. Nat. Commun. 2014, 5, 3930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, L. The biosynthesis of glucosinolates: Insights, inconsistencies, and unknowns. Ann. Plant Rev. Online 2019, 2, 969–1000. [Google Scholar] [CrossRef]
- Warwick, S.I.; Al-Shehbaz, I.A.; Price, R.A.; Sauder, C. Phylogeny of Sisymbrium (Brassicaceae) based on ITS sequences of nuclear ribosomal DNA. Can. J. Bot. 2002, 80, 1002–1017. [Google Scholar] [CrossRef]
- Nikolić, T. (Ed.) (2005-Onwards): Flora Croatica Database. Available online: http://hirc.botanic.hr/fcd (accessed on 16 October 2022).
- Žerdoner Čalasan, A.; German, D.A.; Hurka, H.; Neuffer, B. A story from the Miocene: Clock-dated phylogeny of Sisymbrium L. (Sisymbrieae, Brassicaceae). Ecol. Evol. 2021, 11, 2573–2595. [Google Scholar] [CrossRef]
- Griffiths, D.W.; Deighton, N.; Birch, A.N.E.; Patrian, B.; Baur, R.; Städler, E. Identification of glucosinolates on the leaf surface of plants from the Cruciferae and other closely related species. Phytochemistry 2001, 57, 693–700. [Google Scholar] [CrossRef] [PubMed]
- Di Sotto, A.; Vitalone, A.; Nicoletti, M.; Piccin, A.; Mazzanti, G. Pharmacological and phytochemical study on a Sisymbrium officinale Scop. extract. J. Ethnopharmacol. 2010, 127, 731–736. [Google Scholar] [CrossRef] [PubMed]
- Borgonovo, G.; Zimbaldi, N.; Guarise, M.; De Nisi, P.; De Petrocellis, L.; Moriello, S.A.; Bassoli, A. Isothiocyanates and glucosinolates from Sisymbrium officinale (L.) Scop. (“the Singers’ Plant”): Isolation and in vitro assays on the somatosensory and pain receptor TRPA1 channel. Molecules 2019, 24, 949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blažević, I.; Radonić, A.; Mastelić, J.; Zekić, M.; Skočibušić, M.; Maravić, A. Hedge mustard (Sisymbrium officinale): Chemical diversity of volatiles and their antimicrobial activity. Chem. Biodivers. 2010, 7, 2023–2034. [Google Scholar] [CrossRef]
- Daxenbichler, M.E.; Spencer, G.F.; Carlson, D.G.; Rose, G.B.; Brinker, A.M.; Powell, R.G. Glucosinolate composition of seeds from 297 species of wild plants. Phytochemistry 1991, 30, 2623. [Google Scholar] [CrossRef]
- Hasapis, X.; MacLeod, A.J.; Moreau, M. Glucosinolates of nine cruciferae and two capparaceae species. Phytochemistry 1981, 20, 2355–2358. [Google Scholar] [CrossRef]
- Fahey, J.W.; Zalcmann, A.T.; Talalay, P. The chemical diversity and distribution of glucosinolates and isothiocyanates among plants. Phytochemistry 2001, 56, 5–51. [Google Scholar] [CrossRef]
- Olsen, C.E.; Huang, X.-C.; Hansen, C.I.C.; Cipollini, D.; Ørgaard, M.; Matthes, A.; Geu-Flores, F.; Koch, M.A.; Agerbirk, N. Glucosinolate diversity within a phylogenetic framework of the tribe Cardamineae (Brassicaceae) unraveled with HPLC-MS/MS and NMR-based analytical distinction of 70 desulfoglucosinolates. Phytochemistry 2016, 132, 33–56. [Google Scholar] [CrossRef]
- Pfalz, M.; Vogel, H.; Kroymann, J. The gene controlling the indole glucosinolate modifier1 quantitative trait locus alters indole glucosinolate structures and aphid resistance in Arabidopsis. Plant Cell 2009, 21, 985–999. [Google Scholar] [CrossRef] [Green Version]
- Pfalz, M.; Mikkelsen, M.D.; Bednarek, P.; Olsen, C.E.; Halkier, B.A.; Kroymann, J. Metabolic engineering in Nicotiana benthamiana reveals key enzyme functions in Arabidopsis indole glucosinolate modification. Plant Cell 2011, 23, 716–729. [Google Scholar] [CrossRef]
- Pfalz, M.; Mukhaimar, M.; Perreau, F.; Kirk, J.; Hansen, C.I.; Olsen, C.E.; Agerbirk, N.; Kroymann, J. Methyl transfer in glucosinolate biosynthesis mediated by indole glucosinolate O-methyltransferase 5. Plant Physiol. 2016, 172, 2190–2203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acheson, R.M. 1-Hydroxypyrroles, 1-hydroxyindoles and 9-hydroxycarbazoles. Adv. Heterocycl. Chem. 1990, 51, 105–175. [Google Scholar] [CrossRef]
- Acheson, R.M.; Littlewood, D.M.; Rosenberg, H.E. Synthesis of 1-methoxyindoles. J. Chem. Soc. Chem. Commun. 1974, 16, 671. [Google Scholar] [CrossRef]
- Somei, M. 1-Hydroxyindoles. Heterocycles 1999, 50, 1157–1211. [Google Scholar] [CrossRef]
- Blažević, I.; Đulović, A.; Čikeš Čulić, V.; Popović, M.; Guillot, X.; Burčul, F.; Rollin, P. Microwave-assisted versus conventional isolation of glucosinolate degradation products from Lunaria annua L. and their cytotoxic activity. Biomolecules 2020, 10, 215. [Google Scholar] [CrossRef] [Green Version]
- Di Sotto, A.; Di Giacomo, S.; Vitalone, A.; Nicoletti, M.; Mazzanti, G. Antimutagenic thio compounds from Sisymbrium officinale. J. Nat. Prod. 2012, 75, 2062–2068. [Google Scholar] [CrossRef]
- Popović, M.; Maravić, A.; Čikeš Čulić, V.; Đulović, A.; Burčul, F.; Blažević, I. Biological effects of glucosinolate degradation products from horseradish: A horse that wins the race. Biomolecules 2020, 10, 343. [Google Scholar] [CrossRef] [Green Version]
- Clarke, D.B. Glucosinolates, structures and analysis in food. Anal. Methods 2010, 2, 310–325. [Google Scholar] [CrossRef]
- Al-Gendy, A.A.; Nematallah, K.A.; Zaghloul, S.S.; Ayoub, N.A. Glucosinolates profile, volatile constituents, antimicrobial, and cytotoxic activities of Lobularia libyca. Pharm. Biol. 2016, 54, 3257–3263. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
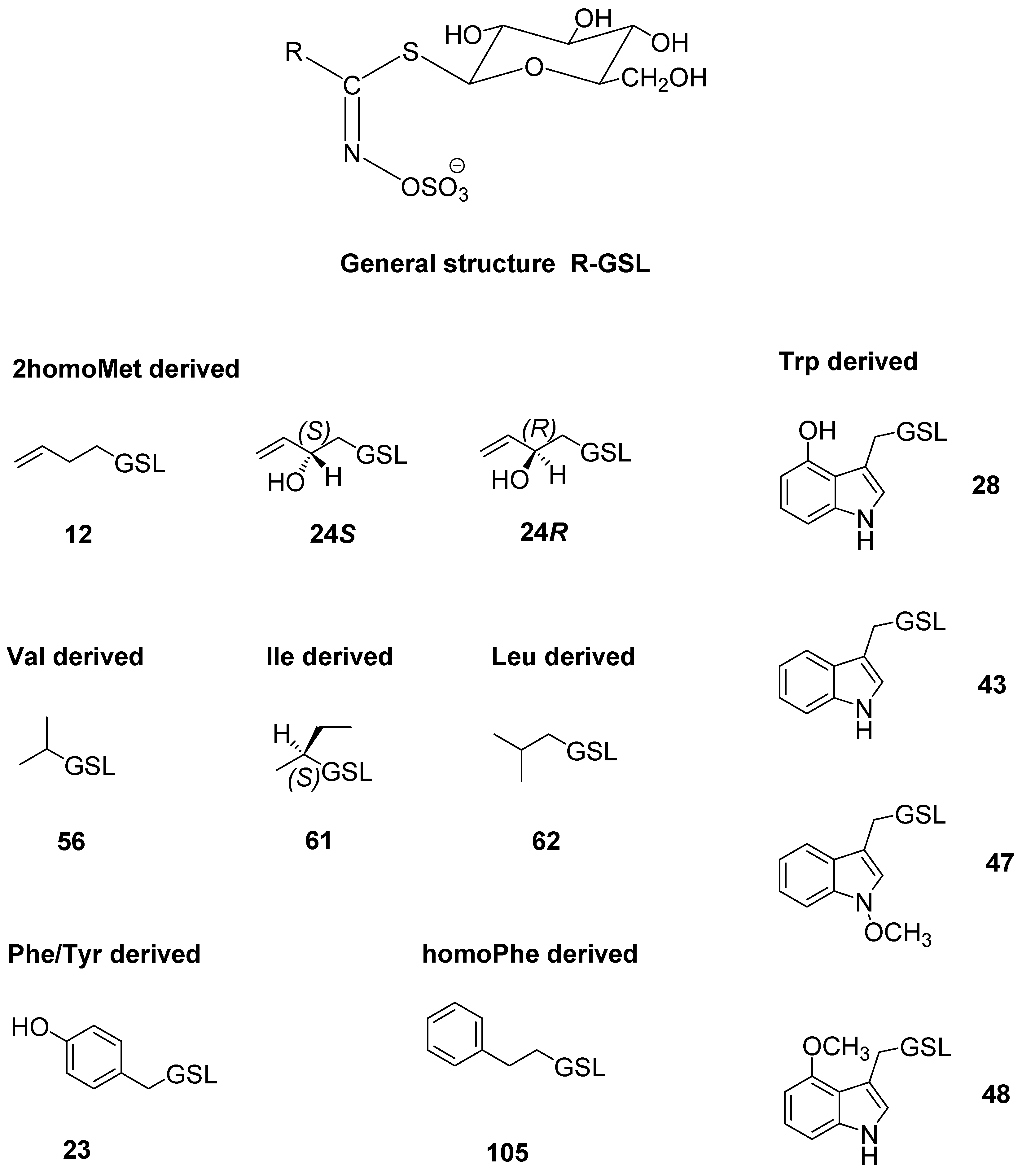
| No. * | Glucosinolate (GSL) (Trivial Name) | Glucosinolates (µmol/g DW) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Split | Krka | ||||||||||
| tR (min) | [M + Na]+ | Root | Stem | Leaf | Flower | Root | Stem | Leaf | Flower | ||
| Val-derived | |||||||||||
| 56 | Isopropyl GSL (glucoputranjivin) | 4.12 | 304 | 1.60 ± 0.21 | 13.54 ± 0.35 | 10.62 ± 0.11 | 11.66 ± 0.18 | tr | 2.72 ± 0.05 | 2.88 ± 0.10 | 7.40 ± 0.22 |
| Leu-derived | |||||||||||
| 62 | Isobutyl GSL | 5.30 | 318 | n.d. | tr | tr | tr | n.d. | n.d. | n.d. | tr |
| Ile-derived | |||||||||||
| 61 | sec-Butyl GSL (glucocochlearin) | 5.64 | 318 | tr | 0.61 ± 0.03 | tr | tr | n.d. | 0.24 ± 0.06 | tr | tr |
| Phe/Tyr-derived | |||||||||||
| 23 | 4-Hydroxybenzyl GSL (glucosinalbin) a | 4.51 | 368 | 0.58 ± 0.08 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Trp-derived | |||||||||||
| 28 | 4-Hydroxyindol-3-ylmethyl GSL (4-hydroxy-glucobrassicin) a | 5.42 | 407 | n.d. | n.d. | tr | tr | n.d. | n.d. | 1.41 ± 0.14 | n.d. |
| 43 | Indol-3-ylmethyl GSL (glucobrassicin) a | 7.21 | 391 | 0.36 ± 0.04 | 1.70 ± 0.15 | 3.81 ± 0.06 | 3.01 ± 0.10 | 0.99 ± 0.13 | 0.74 ± 0.06 | 5.72 ± 0.21 | 4.33 ± 0.17 |
| 48 | 4-Methoxyindol-3-ylmethyl GSL (4-methoxyglucobrassicin) a | 8.01 | 421 | 0.46 ± 0.00 | 0.07 ± 0.00 | n.d. | n.d. | 0.77 ± 0.02 | n.d. | n.d. | n.d. |
| 47 | N-Methoxyindol-3-ylmethyl GSL (neoglucobrassicin) a | 9.34 | 421 | 3.96 ± 0.12 | n.d. | n.d. | n.d. | 5.18 ± 0.18 | 0.02 ± 0.00 | n.d. | 0.04 ± 0.00 |
| Total (µmol/g DW) | 6.96 ± 0.45 | 15.92 ± 1.06 | 14.43 ± 0.17 | 14.67 ± 0.28 | 6.94 ± 0.33 | 3.72 ± 0.17 | 10.01 ± 0.45 | 11.77 ± 0.39 | |||
| No. * | Parent Glucosinolate Identified Breakdown Compound | KI | HD (%) | MAD (%) |
|---|---|---|---|---|
| 56 | Glucoputranjivin | |||
| Isopropyl isothiocyanate a,b | 860 | 27.03 | 51.05 | |
| 61 | Glucocohlearin | |||
| sec-Butyl isothiocyanate b | 955 | 2.27 | 1.46 | |
| Other volatiles | ||||
| Nonanal b | 1114 | tr | tr | |
| β-Cyclocitral b | 1231 | tr | tr | |
| β-Ionone b | 1491 | 2.26 | tr | |
| 6,10,14-Trimethylpentadecan-2-one b | 1857 | 2.67 | 6.48 | |
| Hexadecanoic acid a,b | 2027 | 64.59 | 36.33 | |
| Total (%) | 98.82 | 95.32 | ||
| Yield (µg/g) | 24.55 | 8.90 |
| No.* | Glucosinolate (GSL) (Trivial Name) | Glucosinolates (µmol/g DW) | ||||
|---|---|---|---|---|---|---|
| tR (min) | [M + Na]+ | Root | Stem | Siliquae | ||
| 2homoMet-derived | ||||||
| 24R | (2R)-Hydroxybut-3-enyl GSL (progoitrin) a | 1.41 | 332 | tr | n.d. | 3.03 ± 0.24 |
| 24S | (2S)-Hydroxybut-3-enyl GSL (epiprogoitrin) a | 1.92 | 332 | 0.53 ± 0.13 | 0.25 ± 0.03 | 18.40 ± 0.22 |
| 12 | But-3-enyl GSL (gluconapin) a | 4.80 | 316 | 0.73 ± 0.08 | tr | 0.36 ± 0.09 |
| Phe/Tyr-derived | ||||||
| 23 | 4-Hydroxybenzyl GSL (glucosinalbin) a | 4.51 | 368 | tr | n.d. | n.d. |
| homoPhe derived | ||||||
| 105 | 2-Phenylethyl GSL (gluconasturtiin) a | 7.93 | 366 | 7.89 ± 0.15 | tr | 0.06 ± 0.00 |
| Trp-derived | ||||||
| 47 | N-Methoxyindol-3-ylmethyl GSL (neoglucobrassicin) a | 9.33 | 421 | 0.15 ± 0.00 | n.d. | n.d. |
| Total (µmol/g DW) | 9.15 ± 0.36 | 0.25 ± 0.03 | 21.79 ± 0.55 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Đulović, A.; Popović, M.; Burčul, F.; Čikeš Čulić, V.; Marijan, S.; Ruščić, M.; Anđelković, N.; Blažević, I. Glucosinolates of Sisymbrium officinale and S. orientale. Molecules 2022, 27, 8431. https://doi.org/10.3390/molecules27238431
Đulović A, Popović M, Burčul F, Čikeš Čulić V, Marijan S, Ruščić M, Anđelković N, Blažević I. Glucosinolates of Sisymbrium officinale and S. orientale. Molecules. 2022; 27(23):8431. https://doi.org/10.3390/molecules27238431
Chicago/Turabian StyleĐulović, Azra, Marijana Popović, Franko Burčul, Vedrana Čikeš Čulić, Sandra Marijan, Mirko Ruščić, Nikolina Anđelković, and Ivica Blažević. 2022. "Glucosinolates of Sisymbrium officinale and S. orientale" Molecules 27, no. 23: 8431. https://doi.org/10.3390/molecules27238431