

Natural Compounds in the Battle against Microorganisms—Linalool

Abstract
:1. Introduction
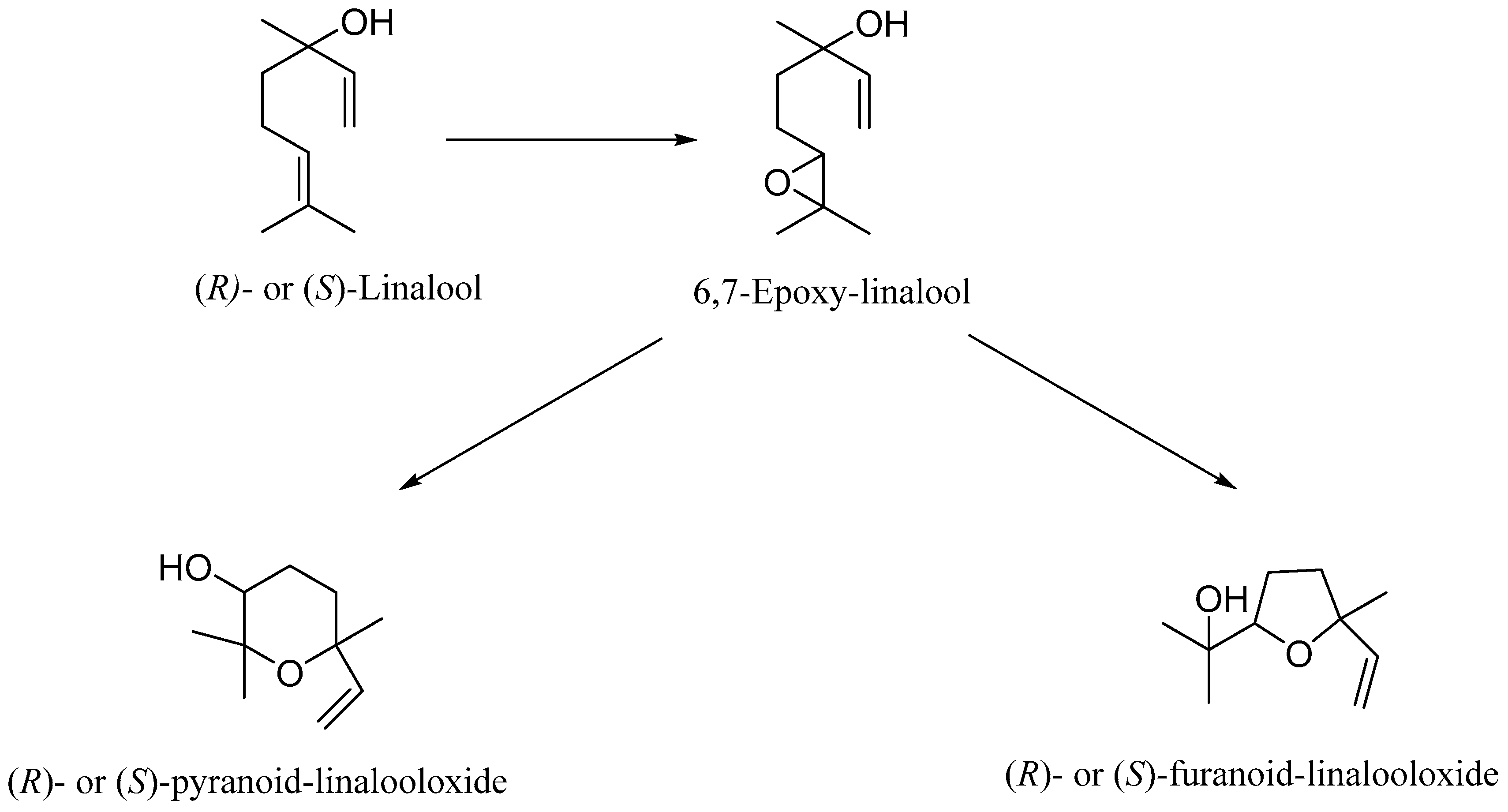
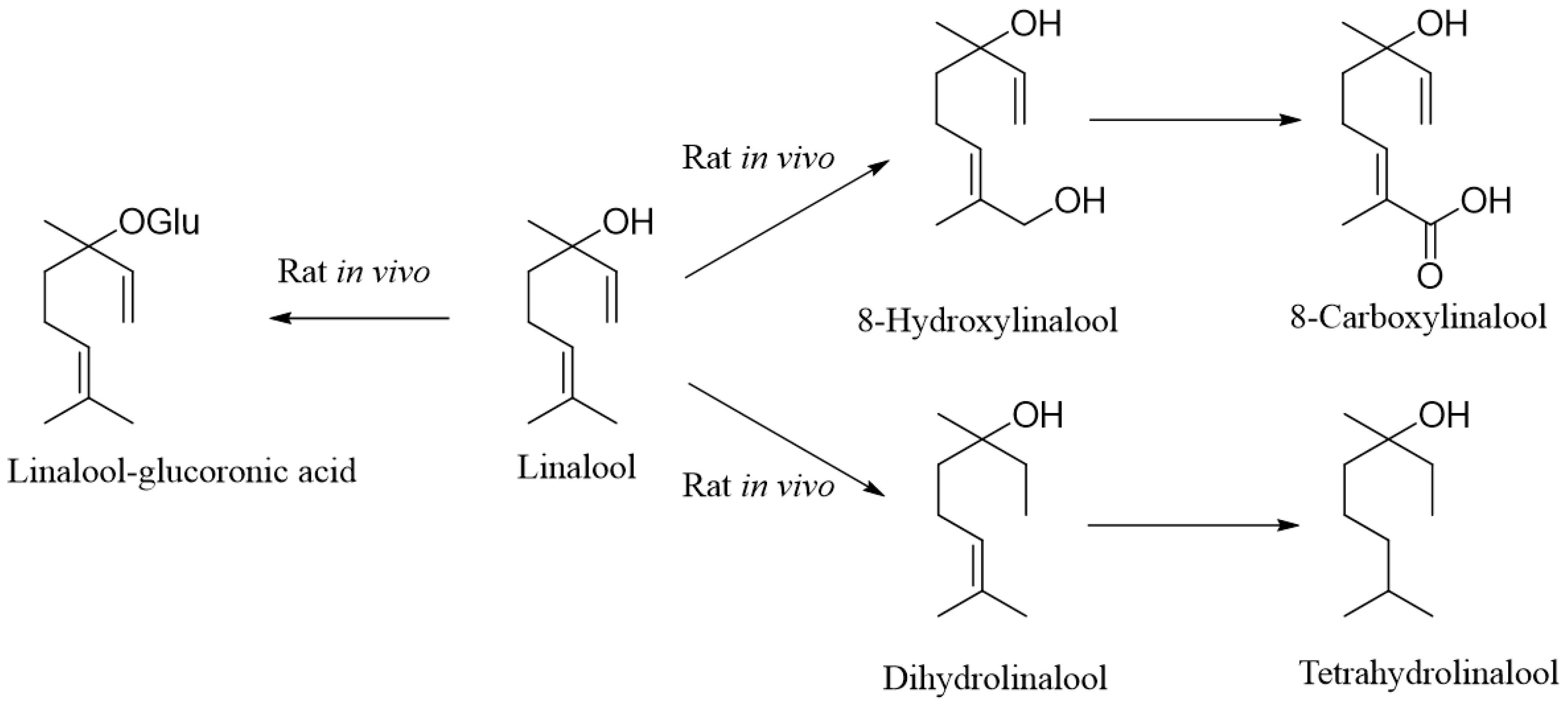
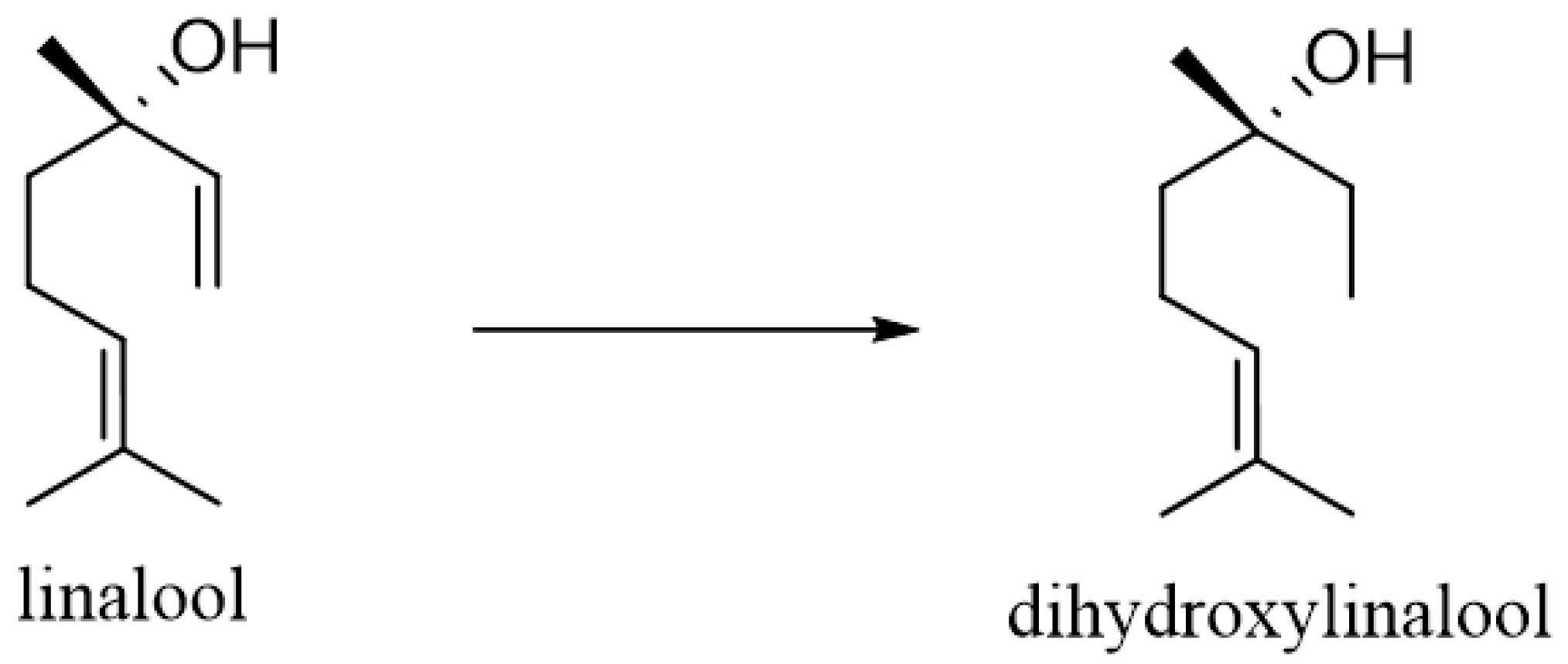
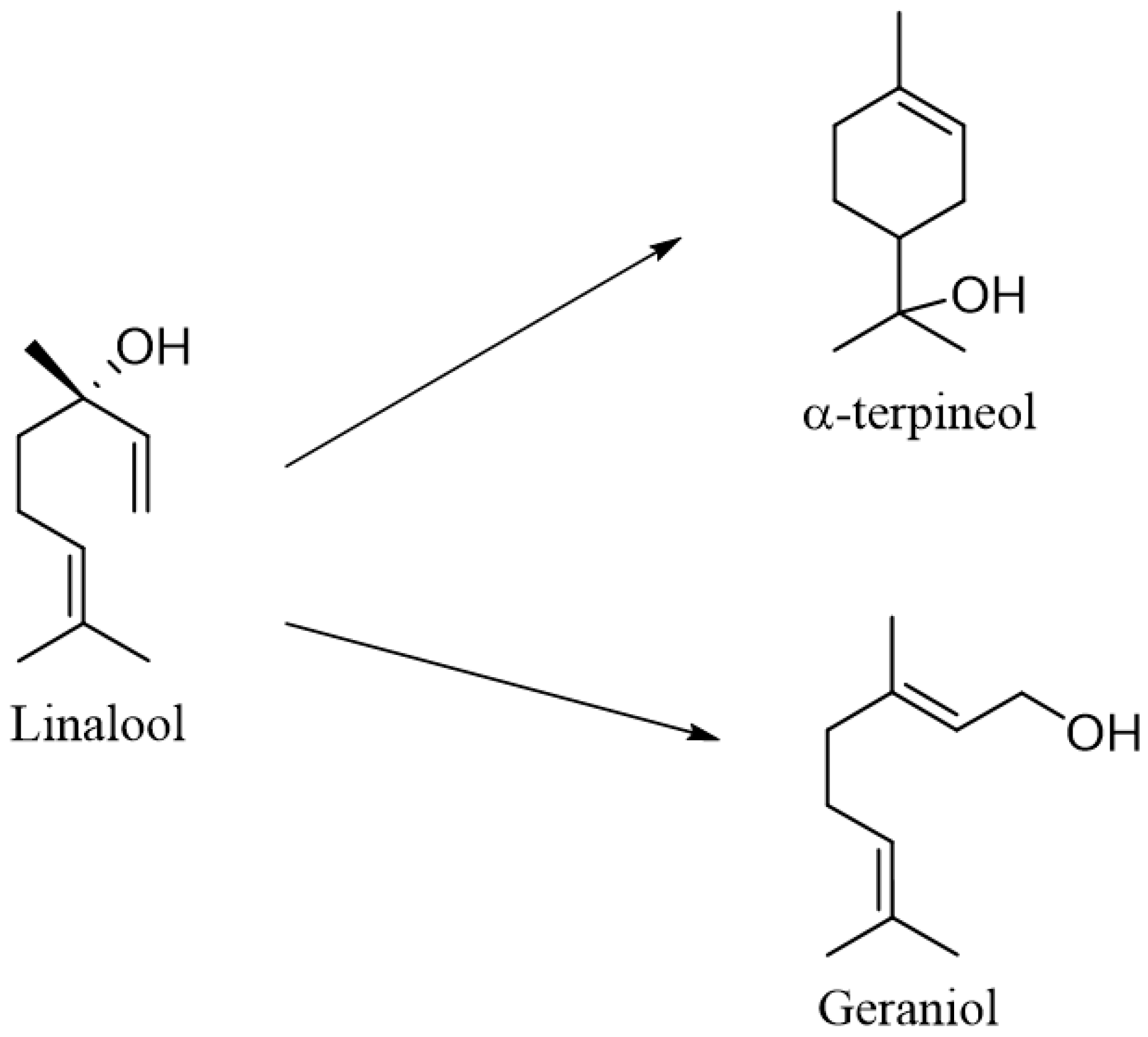
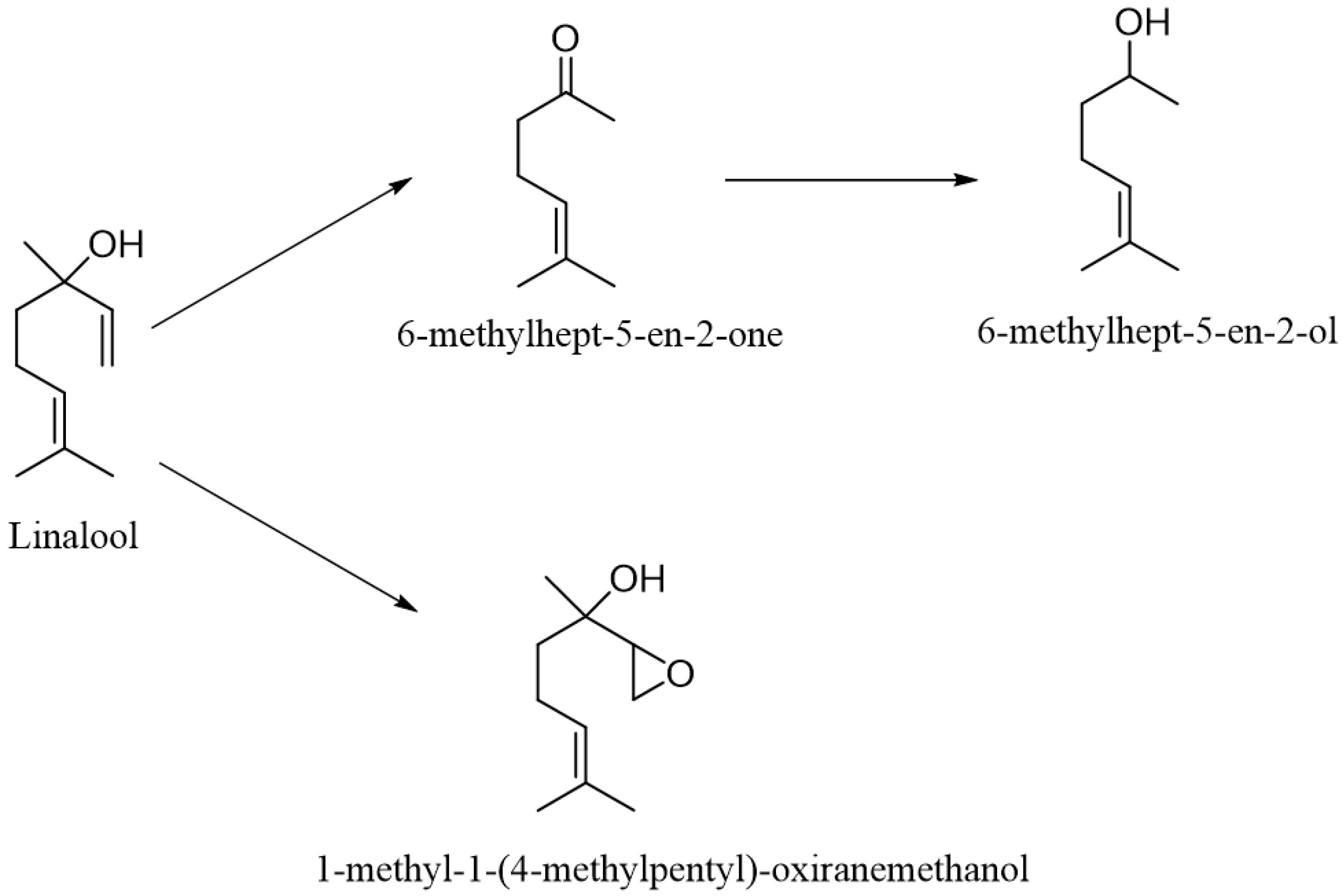
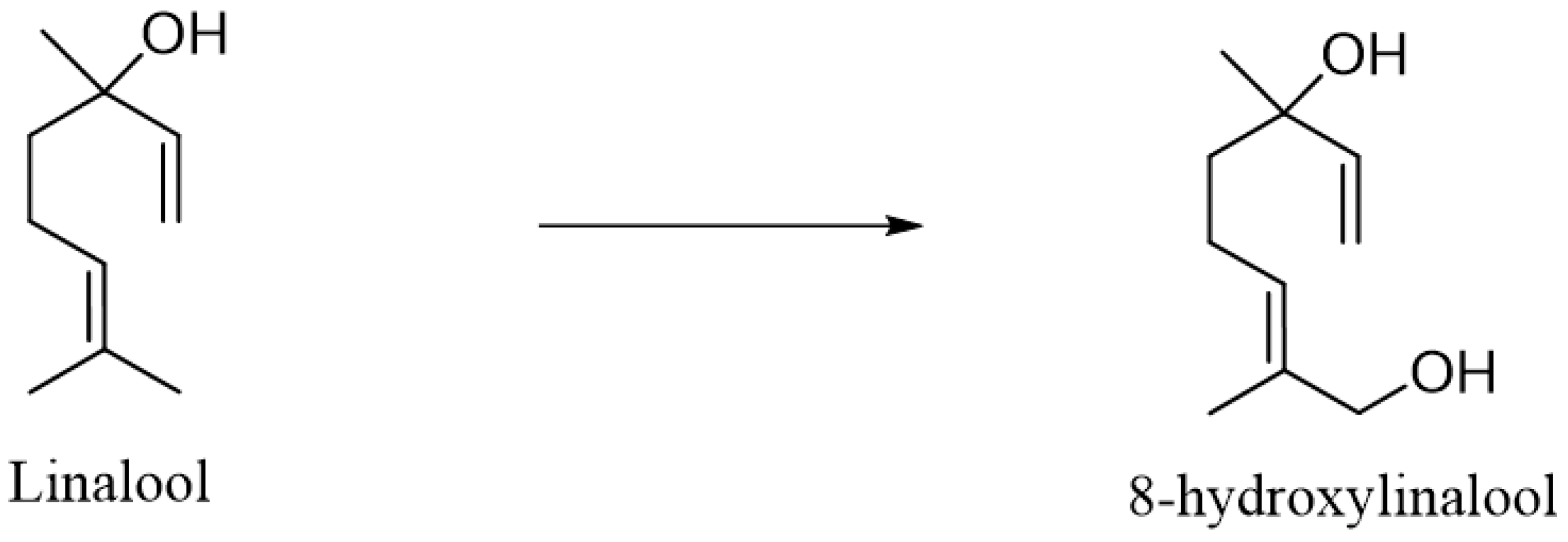
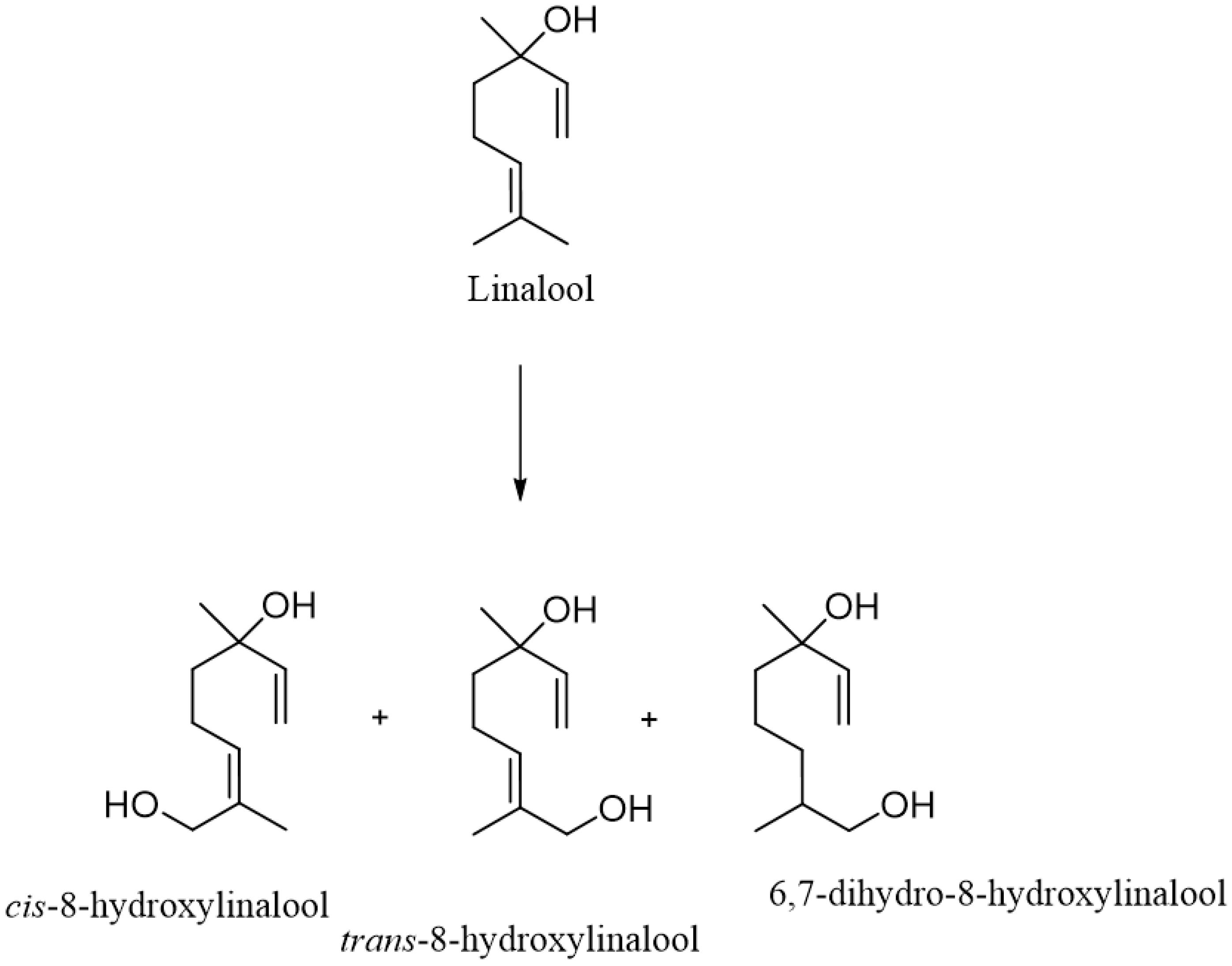
2. Metabolism of Linalool
3. Antimicrobial Activity
3.1. Mechanism of the Antibacterial Action of Linalool
3.1.1. Alteration of Amino Acid Metabolism
3.1.2. Change in Carbohydrate Metabolism
3.1.3. Changes in Lipid Metabolism
3.2. Antifungal Properties of Linalool
4. Microbial Biotransformation of Linalool
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Ferraz, C.A.; Leferink, N.G.; Kosov, I.; Scrutton, N.S. Isopentenol utilization pathway for the production of linalool in Escherichia coli using an improved bacterial linalool/nerolidol synthase. ChemBioChem 2021, 22, 2325–2334. [Google Scholar] [CrossRef] [PubMed]
- Caputi, L.; Aprea, E. Use of terpenoids as natural flavouring compounds in food industry. Recent Pat. Food Nutr. Agric. 2011, 3, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Derbassi, N.B.; Pedrosa, M.C.; Heleno, S.A.; Carocho, M.; Ferreira, I.C.; Barros, L. Plant volatiles: Using scented molecules as food additives. Trends Food Sci. Technol. 2022, 122, 97–103. [Google Scholar] [CrossRef]
- Rinaldi, M.A.; Tait, S.; Toogood, H.S.; Scrutton, N.S. Bioproduction of Linalool From Paper Mill Waste. Front. Bioeng. Biotechnol. 2022, 10, 892896. [Google Scholar] [CrossRef] [PubMed]
- de Alvarenga, J.F.R.; Genaro, B.; Costa, B.L.; Purgatto, E.; Manach, C.; Fiamoncini, J. Monoterpenes: Current knowledge on food source, metabolism, and health effects. Crit. Rev. Food Sci. Nutr. 2021, 2, 1–38. [Google Scholar] [CrossRef] [PubMed]
- Orth, A.M.; Poplacean, I.; Fastowski, O.; Engel, K.H. Assessment of dietary exposure to flavouring substances via consumption of flavoured teas. Part II: Transfer rates of linalool and linalyl esters into Earl Grey tea infusions. Food Addit. Contam. Part A 2014, 31, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Chantraine, J.M.; Dhénin, J.M.; Moretti, C. Chemical variability of rosewood (Aniba rosaeodora Ducke) essential oil in French Guiana. J. Essent. Oil Res. 2009, 21, 486–495. [Google Scholar] [CrossRef]
- Frizzo, C.D.; Santos, A.C.; Paroul, N.; Serafini, L.A.; Dellacassa, E.; Lorenzo, D.; Moyna, P. Essential oils of camphor tree (Cinnamomum camphora Nees & Eberm) cultivated in Southern Brazil. Braz. Arch. Biol. Technol. 2000, 43, 313–316. [Google Scholar]
- Kofidis, G.; Bosabalidis, A.; Kokkini, S. Seasonal variation of essential oils in a linalool-rich chemotype of Mentha spicata grown wild in Greece. J. Essent. Oil Res. 2004, 16, 469–472. [Google Scholar] [CrossRef]
- Cheng, B.H.; Lin, C.Y.; Yeh, T.F.; Cheng, S.S.; Chang, S.T. Potential source of S-(+)-linalool from Cinnamomum osmophloeum ct. linalool leaf: Essential oil profile and enantiomeric purity. J. Agric. Food Chem. 2012, 60, 7623–7628. [Google Scholar] [CrossRef]
- Prosea Foundation. Plant Resources of South-East Asia, 19, Essentialoil Plants; Oyen, L.P.A., Dung, N.X., Eds.; Prosea Foundation: Bogor, Indonesia, 1999. [Google Scholar]
- Eikani, M.H.; Golmohammad, F.; Rowshanzamir, S. Subcritical water extraction of essential oils from coriander seeds (Coriandrum sativum L.). J. Food Eng. 2007, 80, 735–740. [Google Scholar] [CrossRef]
- National Institute of Industrial Research (NIIR) Board. The Complete Technology Book of Essential Oils (Aromatic Chemicals); Asia Pacific Business Press: Delhi, India, 2003. [Google Scholar]
- Galata, M.; Sarker, L.S.; Mahmoud, S.S. Transcriptome profiling, and cloning and characterization of the main monoterpene synthases of Coriandrum sativum L. Phytochemistry 2014, 102, 64–73. [Google Scholar] [CrossRef] [Green Version]
- Jain, N.; Srivastava, S.K.; Aggarwal, K.K.; Ramesh, S.; Kumar, S. Essential oil composition of Zanthoxylum alatum seeds from northern India. Flavour. Fragr J. 2001, 16, 408–410. [Google Scholar] [CrossRef]
- Sourmaghi, M.H.S.; Kiaee, G.; Golfakhrabadi, F.; Jamalifar, H.; Khanavi, M. Comparison of essential oil composition and antimicrobial activity of Coriandrum sativum L. extracted by hydrodistillation and microwave-assisted hydrodistillation. J. Food Sci. Technol. 2015, 52, 2452–2457. [Google Scholar] [CrossRef] [Green Version]
- Carrasco, A.; Martinez-Gutierrez, R.; Tomas, V.; Tudela, J. Lavandula angustifolia and Lavandula latifolia essential oils from Spain: Aromatic profile and bioactivities. Planta Med. 2016, 82, 163–170. [Google Scholar]
- Tomazoni, E.Z.; Pansera, M.R.; Pauletti, G.F.; Moura, S.; Ribeiro, R.T.; Schwambach, J. In vitro antifungal activity of four chemotypes of Lippia alba (Verbenaceae) essential oils against Alternaria solani (Pleosporeaceae) isolates. An. Acad. Bras. Cienc. 2016, 88, 999–1010. [Google Scholar] [CrossRef] [Green Version]
- Venditti, A.; Bianco, A.; Frezza, C.; Conti, F.; Bini, L.M.; Giuliani, C.; Bramucci, M.; Quassinti, L.; Damiano, S.; Lupidi, G.; et al. Essential oil composition, polar compounds, glandular trichomes and biological activity of Hyssopus officinalis subsp. aristatus (Godr.) Nyman from central Italy. Ind Crops Prod. 2015, 77, 353–363. [Google Scholar] [CrossRef]
- Stashenko, E.E.; Martínez, J.R. Sampling flower scent for chromatographic analysis. J. Sep. Sci. 2008, 31, 2022–2031. [Google Scholar] [CrossRef]
- Aprotosoaie, A.C.; Hăncianu, M.; Costache, I.-I.; Miron, A. Linalool: A review on a key odorant molecule with valuable biological properties. Flavour Fragr. J. 2014, 29, 193–219. [Google Scholar] [CrossRef]
- Cseke, L.; Dudareva, N.; Pichersky, E. Structure and evolution of linalool synthase. Mol. Biol. Evol. 1998, 15, 1491–1498. [Google Scholar] [CrossRef] [Green Version]
- Nitta, N.; Tajima, Y.; Yamamoto, Y.; Moriya, M.; Matsudaira, A.; Hoshino, Y.; Usuda, Y. Fermentative production of enantiopure (S)-linalool using a metabolically engineered Pantoea ananatis. Microb. Cell Fact. 2021, 20, 1–14. [Google Scholar] [CrossRef]
- Peana, A.T.; D’Aquila, P.S.; Panin, F.; Serra, G.; Pippia, P.; Moretti, M.D.L. Anti-inflammatory activity of linalool and linalyl acetate constituents of essential oils. Phytomed 2002, 9, 721–726. [Google Scholar] [CrossRef]
- Downer, E.J. Anti-inflammatory potential of terpenes present in Cannabis sativa L. ACS Chem. Neurosci. 2020, 11, 659–662. [Google Scholar] [CrossRef]
- Liu, K.; Chen, Q.; Liu, Y.; Zhou, X.; Wang, X. Isolation and biological activities of decanal, linalool, valencene, and octanal from sweet orange oil. J. Food Sci. 2012, 77, C1156–C1161. [Google Scholar] [CrossRef]
- Lee, S.C.; Wang, S.Y.; Li, C.C.; Liu, C.T. Anti-inflammatory effect of cinnamaldehyde and linalool from the leaf essential oil of Cinnamomum osmophloeum Kanehira in endotoxin-induced mice. J. Food Drug Anal. 2018, 26, 211–220. [Google Scholar] [CrossRef] [Green Version]
- Deepa, B.; Venkatraman Anuradha, C. Effects of linalool on inflammation, matrix accumulation and podocyte loss in kidney of streptozotocin-induced diabetic rats. Toxicol. Mechan. Methods. 2013, 23, 223–234. [Google Scholar] [CrossRef]
- Cho, S.Y.; Jun, H.J.; Lee, J.H.; Jia, Y.; Kim, K.H.; Lee, S.J. Linalool reduces the expression of 3-hydroxy-3-methylglutaryl CoA reductase via sterol regulatory element binding protein-2- and ubiquitin-dependent mechanisms. FEBS Lett. 2011, 585, 3289–3296. [Google Scholar] [CrossRef] [Green Version]
- Peana, A.T.; Marzocco, S.; Popolo, A.; Pinto, A. (−)-Linalool inhibits in vitro NO formation: Probable involvement in the antinociceptive activity of this monoterpene compound. Life Sci. 2006, 78, 719–723. [Google Scholar] [CrossRef]
- de Lucena, J.D.; Gadelha-Filho, C.V.J.; da Costa, R.O.; de Araújo, D.P.; Lima, F.A.V.; Neves, K.R.T.; de Barros Viana, G.S. L-linalool exerts a neuroprotective action on hemiparkinsonian rats. Naunyn Schmiedebergs Arch. Pharmacol. 2020, 393, 1077–1088. [Google Scholar] [CrossRef]
- Aoe, M.; Ueno-Iio, T.; Shibakura, M.; Shinohata, R.; Usui, S.; Arao, Y.; Ikeda, S.; Miyahara, N.; Tanimoto, M.; Kataoka, M. Lavender essential oil and its main constituents inhibit the expression of TNF-α-induced cell adhesion molecules in endothelial cells. Acta Med. Okayama 2017, 71, 493–503. [Google Scholar]
- Li, Y.; Lv, O.; Zhou, F.; Li, Q.; Wu, Z.; Zheng, Y. Linalool inhibits LPS-induced inflammation in BV2 microglia cells by activating Nrf2. Neurochem. Res. 2015, 40, 1520–1525. [Google Scholar] [CrossRef] [PubMed]
- Weston-Green, K.; Clunas, H.; Jimenez Naranjo, C. A review of the potential use of pinene and linalool as terpene-based medicines for brain health: Discovering novel therapeutics in the flavours and fragrances of cannabis. Front. Psychiatry 2021, 12, 583211. [Google Scholar] [CrossRef] [PubMed]
- Sabogal-Guáqueta, A.M.; Osorio, E.; Cardona-Gómez, G.P. Linalool reverses neuropathological and behavioral impairments in old triple transgenic Alzheimer’s mice. Neuropharmacology 2016, 102, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Pereira, I.; Severino, P.; Santos, A.C.; Silva, A.M.; Souto, E.B. Linalool bioactive properties and potential applicability in drug delivery systems. Colloids Surf. B Biointerfaces 2018, 171, 566–578. [Google Scholar] [CrossRef]
- Elbe, H.; Ozturk, F.; Yigitturk, G.; Baygar, T.; Cavusoglu, T. Anticancer activity of linalool: Comparative investigation of ultrastructural changes and apoptosis in breast cancer cells. Ultrastruct. Pathol. 2022, 46, 1–11. [Google Scholar] [CrossRef]
- Silva, B.I.; Nascimento, E.A.; Silva, C.J.; Silva, T.G.; Aguiar, J.S. Anticancer activity of monoterpenes: A systematic review. Mol. Biol. Rep. 2021, 48, 5775–5785. [Google Scholar] [CrossRef]
- Kasper, S.; Gastpar, M.; Muller, W.E.; Volz, H.P.; Moller, H.J.; Dienel, A.; Schlafke, S. Efficacy and safety of silexan, a new, orally administered lavender oil preparation, in subthreshold anxiety disorder-evidence from clinical trials. Wien. Med. Wochenschr. 2010, 160, 547–556. [Google Scholar] [CrossRef]
- Yoshida, K.; Yamamoto, N.; Fujiwara, S.; Kamei, A.; Abe, K.; Nakamura, A. Inhalation of a racemic mixture (R,S)-linalool by rats experiencing restraint stress alters neuropeptide and MHC class I gene expression in the hypothalamus. Neurosci. Lett. 2017, 653, 314–319. [Google Scholar] [CrossRef]
- Mohamed, M.E.; Abduldaium, M.S.; Younis, N.S. Cardioprotective Effect of Linalool against isoproterenol-induced myocardial infarction. Life 2021, 11, 120. [Google Scholar] [CrossRef]
- Liang, Y.; Zhong, Y.; Li, X.; Xiao, Y.; Wu, Y.; Xie, P. Biological evaluation of linalool on the function of blood vessels. Mol. Med. Rep. 2021, 24, 1–9. [Google Scholar] [CrossRef]
- Buchbauer, G.; Jirovetz, L. Percutaneous absorption of lavender oil from a massage oil. J. Soc. Cosmet. Chem. 1992, 43, 49–54. [Google Scholar]
- Friedl, S.M.; Oedendorfer, K.; Kitzer, S.; Reznicek, G.; Sladek, G.; Heuberger, E. Comparison of liquid-liquid partition, HS-SPME and static HS GC/MS analysis for the quantification of (−)-linalool in human whole blood samples. Nat. Prod. Commun. 2010, 5, 1447–1452. [Google Scholar] [CrossRef]
- Cal, K. How does the type of vehicle influence the in vitro skin absorption and elimination kinetics of terpenes? Arch. Derm. Res. 2006, 297, 311–315. [Google Scholar] [CrossRef]
- Bradley, B.F.; Brown, S.L.; Chu, S.; Lea, R.W. Effects of orally administered lavender essential oil on responses to anxiety-provoking film clips. Hum. Psychopharmacol. 2009, 24, 319–330. [Google Scholar] [CrossRef]
- Meesters, R.J.W.; Duisken, M.; Hollender, J. Study on the cytochrome P450-mediated oxidative metabolism of the terpene alcohol linalool: Indication of biological epoxidation. Xenobiotica 2007, 37, 604–617. [Google Scholar] [CrossRef]
- Roffey, S.J.; Walker, R.; Gibson, G.G. Hepatic peroxisomal and microsomal enzyme induction by citral and linalool in rats. Food Chem Toxicol 1990, 28, 403–408. [Google Scholar] [CrossRef]
- Parke, D.V.; Rahman, K.M.Q.; Walker, R. The absorption, distribution and excretion of linalool in the rat. Biochem. Soc. Trans. 1974, 2, 612–615. [Google Scholar] [CrossRef]
- Parke, D.V.; Rahman, K.M.Q.; Walker, R. Effect of linalool on hepatic drug-metabolizing enzymes in the rat. Biochem Soc. Trans. 1974, 2, 615–618. [Google Scholar] [CrossRef]
- Chadha, A.; Madyastha, K.M. Metabolism of geraniol and linalool in the rat and effects on liver and lung microsomal enzymes. Xenobiotica 1984, 14, 365–374. [Google Scholar] [CrossRef]
- Bickers, D.; Calow, P.; Greim, H.; Hanifin, J.M.; Rogers, A.E.; Saurat, J.H.; Sipes, I.G.; Smith, R.L.; Tagami, H. A toxicologic and dermatologic assessment of linalool and related esters when used as fragrance ingredients. Food Chem. Toxicol. 2003, 41, 919–942. [Google Scholar] [CrossRef]
- Nosková, K.; Dovrtělová, G.; Zendulka, O.; Řemínek, R.; Juřica, J. The effect of (–)-linalool on the metabolic activity of liver CYP enzymes in rats. Physiol. Res. 2016, 65 (Suppl. 4), S499–S504. [Google Scholar] [CrossRef]
- Zehetner, P.; Höferl, M.; Buchbauer, G. Essential oil components and cytochrome P450 enzymes: A review. Flavour Fragr. J. 2019, 34, 223–240. [Google Scholar] [CrossRef]
- Zengin, H.; Baysal, A.H. Antibacterial and Antioxidant Activity of Essential Oil Terpenes against Pathogenic and Spoilage-Forming Bacteria and Cell Structure-Activity Relationships Evaluated by SEM Microscopy. Molecules 2014, 19, 17773–17798. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Cai, J.; Chen, H.; Zhong, Q.; Hou, Y.; Chen, W.; Chen, W. Antibacterial activity and mechanism of linalool against Pseudomonas aeruginosa. Microb. Pathog. 2020, 141, 103980. [Google Scholar] [CrossRef]
- Ghosh, T.; Srivastava, S.K.; Gaurav, A.; Kumar, A.; Kumar, P.; Yadav, A.S.; Pathania, R.; Navani, N.K. A combination of linalool, vitamin C, and copper synergistically triggers reactive oxygen species and DNA damage and inhibits Salmonella enterica subsp. enterica serovar Typhi and Vibrio fluvialis. Appl. Environ. Microbiol. 2019, 85, e02487-18. [Google Scholar] [CrossRef] [Green Version]
- Prakash, A.; Vadivel, V.; Rubini, D.; Nithyanand, P. Antibacterial and antibiofilm activities of linalool nanoemulsions against Salmonella Typhimurium. Food Biosci. 2019, 28, 57–65. [Google Scholar] [CrossRef]
- Duarte, A.; Luís, Â.; Oleastro, M.; Domingues, F.C. Antioxidant properties of coriander essential oil and linalool and their potential to control Campylobacter spp. Food Control. 2016, 61, 115–122. [Google Scholar] [CrossRef]
- Park, S.N.; Lim, Y.K.; Freire, M.O.; Cho, E.; Jin, D.; Kook, J.K. Antimicrobial effect of linalool and α-terpineol against periodontopathic and cariogenic bacteria. Anaerobe 2012, 18, 369–372. [Google Scholar] [CrossRef]
- Soković, M.; Glamočlija, J.; Marin, P.D.; Brkić, D.; Griensven, L.J.L.D.V. Antibacterial Effects of the Essential Oils of Commonly Consumed Medicinal Herbs Using an In Vitro Model. Molecules 2010, 15, 7532–7546. [Google Scholar] [CrossRef] [Green Version]
- Prakash, A.; Vadivel, V. Citral and linalool nanoemulsions: Impact of synergism and ripening inhibitors on the stability and antibacterial activity against Listeria monocytogenes. J. Food Sci. Technol. 2020, 57, 1495–1504. [Google Scholar] [CrossRef]
- Herman, A.; Tambor, K.; Herman, A. Linalool Affects the Antimicrobial Efficacy of Essential Oils. Curr. Microbiol. 2016, 72, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Chen, Q.; Liang, Q.; Zhang, M.; Chen, W.; Chen, H.; Yun, Y.; Zhong, Q.; Chen, W. Antimicrobial Activity and Proposed Action Mechanism of Linalool Against Pseudomonas fluorescens. Front. Microbiol. 2021, 12, 562094. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Liang, Q.; Zhang, M.; Chen, W.; Chen, H.; Yun, Y.; Zhong, Q.; Chen, W. Antibacterial Activity and Mechanism of Linalool against Shewanella putrefaciens. Molecules 2021, 26, 245. [Google Scholar] [CrossRef] [PubMed]
- Máté, G.; Kovács, D.; Gazdag, Z.; Pesti, M.; Szántó, Á. Linalool-induced oxidative stress processes in the human pathogen Candida albicans. Acta Biol. Hung. 2017, 68, 220–231. [Google Scholar] [CrossRef]
- De Oliveira Lima, M.I.; Araújo de Medeiros, A.C.; Souza Silva, K.V.; Cardoso, G.N.; de Oliveira Lima, E.; de Oliveira Pereira, F. Investigation of the antifungal potential of linalool against clinical isolates of fluconazole resistant Trichophyton rubrum. J. Mycol. Med. 2017, 27, 195–202. [Google Scholar] [CrossRef]
- Li, S.; Zhang, S.; Lv, Y.; Zhai, H.; Hu, Y.; Cai, J. Heptanal inhibits the growth of Aspergillus flavus through disturbance of plasma membrane integrity, mitochondrial function and antioxidant enzyme activity. LWT-Food Sci. Technol. 2022, 154, 112655. [Google Scholar] [CrossRef]
- Khan, A.; Ahmad, A.; Manzoor, N.; Khan, L.A. Antifungal activities of Ocimum sanctum essential oil and its lead molecules. Nat. Prod. Commun. 2010, 5, 345–349. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, N.N.; Alviano, C.S.; Blank, A.F.; Romanos, M.T.V.; Fonseca, B.B.; Rozental, S.; Rodrigues, I.A.; Alviano, D.S. Synergism effect of the essential oil from Ocimum basilicum var. Maria Bonita and its major components with fluconazole and its influence on ergosterol biosynthesis. Evid.-based Complement. Altern. Med. 2016, 2016, 5647182. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Z.; Zhang, B.; Wang, Y.; Li, F.; Jin, Z.; Lü, X.; Zhang, H.; Zhang, J.; Zhao, J. Transcriptomic Analysis Reveals That Exogenous Indole-3-Butyric Acid Affects the Rooting Process During Stem Segment Culturing of Cinnamomum camphora Linalool Type. Plant. Mol. Biol. Rep. 2022, 1–13. [Google Scholar] [CrossRef]
- Sabogal-Guáqueta, A.M.; Posada-Duque, R.; Cortes, N.C.; Arias-Londoño, J.D.; Cardona-Gómez, G.P. Changes in the hippocampal and peripheral phospholipid profiles are associated with neurodegeneration hallmarks in a long-term global cerebral ischemia model: Attenuation by Linalool. Neuropharmacology 2018, 135, 555–571. [Google Scholar] [CrossRef]
- Hsu, C.C.; Lai, W.L.; Chuang, K.C.; Lee, M.H.; Tsai, Y.C. The inhibitory activity of linalool against the filamentous growth and biofilm formation in Candida albicans. Med. Mycol. 2013, 51, 473–482. [Google Scholar] [CrossRef] [Green Version]
- Souza, C.M.; Pereira Junior, S.A.; Moraes Tda, S.; Damasceno, J.L.; Amorim Mendes, S.; Dias, H.J.; Stefani, R.; Tavares, D.C.; Martins, C.H.; Crotti, A.E.; et al. Antifungal activity of plant-derived essential oils on Candida tropicalis planktonic and biofilms cells. Med. Mycol. 2016, 54, 515–523. [Google Scholar] [CrossRef] [Green Version]
- Silva, K.V.S.; Lima, M.I.O.; Cardoso, G.N.; Santos, A.S.; Silva, G.S.; Pereira, F.O. Inibitory effects of linalool on fungal pathogenicity of clinical isolates of Microsporum canis and Microsporum gypseum. Mycoses 2017, 60, 387–393. [Google Scholar] [CrossRef]
- Mirata, M.-A.; Wust, M.; Mosandl, A.; Schrader, J. Fungal Biotransformation of (±)-Linalool. J. Agric. Food Chem. 2008, 56, 3287–3296. [Google Scholar] [CrossRef]
- Demyttenaere, J.C.R.; Willemen, H.M. Biotransformation of linalool to furanoid and pyranoid linalool oxides by Aspergillus Niger. Phytochem. 1998, 47, 1029–1036. [Google Scholar] [CrossRef]
- Demyttenaere, J.C.; Adams, A.; Vanoverschelde, J.; De Kimpe, N. Biotransformation of (S)-(+)-linalool by Aspergillus niger: An investigation of the culture conditions. J. Agric. Food Chem. 2001, 49, 5895–5901. [Google Scholar] [CrossRef]
- Abraham, W.-R.; Stumpf, B.; Arfmann, H.-A. Chiral intermediates by microbial epoxidations. J. Essent. Oil Res. 1990, 2, 251–257. [Google Scholar] [CrossRef]
- Bormann, S.; Etschmann, M.M.; Mirata, M.A.; Schrader, J. Integrated bioprocess for the stereospecific production of linalool oxides from linalool with Corynespora cassiicola DSM 62475. J. Ind. Microbiol. Biotechnol. 2012, 39, 1761–1769. [Google Scholar] [CrossRef]
- Sales, A.; Afonso, L.F.; Americo, J.A.; de Freitas Rebelo, M.; Pastore, G.M.; Bicas, J.L. Monoterpene biotransformation by Colletotrichum species. Biotechnol. Lett. 2018, 40, 561–567. [Google Scholar] [CrossRef]
- Ghasemi, Y.; Mohagheghzadeh, A.; Moshavash, M.; Ostovan, Z.; Rasoul-Amini, S.; Morowvat, M.H.; Ghoshoon, M.B.; Raee, M.J.; Mosavi-Azam, S.B. Biotransformation of monoterpenes by Oocystis pusilla. World J. Microbiol. Biotechnol. 2009, 25, 1301–1304. [Google Scholar] [CrossRef]
- Rasoul-Amini, S.; Fotooh-Abadi, E.; Ghasemi, Y. Biotransformation of monoterpenes by immobilized microalgae. J. Appl. Phycol. 2011, 23, 975–981. [Google Scholar] [CrossRef]
- Molina, G.; Pinheiro, D.M.; Pimentel, M.R.; dos Ssanros, R.; Pastore, G.M. Monoterpene bioconversion for the production of aroma compounds by fungi isolated from Brazilian fruits. Food Sci. Biotechnol. 2013, 22, 999–1006. [Google Scholar] [CrossRef]
- King, A.; Richard Dickinson, J. Biotransformation of monoterpene alcohols by Saccharomyces cerevisiae, Torulaspora delbrueckii and Kluyveromyces lactis. Yeast 2000, 16, 499–506. [Google Scholar] [CrossRef]
- Moreno Rueda, M.G.; Guerrini, A.; Giovannini, P.P.; Medici, A.; Grandini, A.; Sacchetti, G.; Pedrini, P. Biotransformations of Terpenes by Fungi from Amazonian Citrus Plants. Chem. Biodivers. 2013, 10, 1909–1919. [Google Scholar] [CrossRef]
- Bell, S.G.; Dale, A.; Rees, N.H.; Wong, L.-L. A cytochrome P450 class I electron transfer system from Novosphingobium aromaticivorans. Appl. Microbiol. Biotechnol. 2010, 86, 163–175. [Google Scholar] [CrossRef]
- Bock, G.; Benda, I.; Schreier, P. Biotransformation of linalool by Botrytis cinerea. J. Food Sci. 1986, 51, 659–662. [Google Scholar] [CrossRef]
- Toshifumi, H.; Tadashi, A.; Yoshio, H.; Takashi, I.; Takayuki, S. The biotransformation of foreign substrates by tissue cultures. I. The hydroxylation of linalool and its related compounds with the suspension cells of Nicotiana tabacum. Bull. Chem. Soc. Jpn. 1981, 54, 3527–3529. [Google Scholar]
- Mathieu, S.; Wirth, J.; Sauvage, F.-X.; Lepoutre, J.-P.; Baumes, R.; Gunata, Z. Biotransformation of C13-norisoprenoids and monoterpenes by a cell suspension culture of cv. Gamay (Vitis vinifera). Plant. Cell Tiss Organ. Cult. 2009, 97, 203–221. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant | Plant Part | Percentage [%] | Ref. |
|---|---|---|---|
| Aniba rosaeodora Ducke | Wood | 100 | [7] |
| Cinnamomum camphora Nees & Eberm var. linaloolifera | Leaf | 95 | [8] |
| Mentha spicata | Leaf | 94 | [9] |
| Cinnamomum osmophloeum ct. | Leaf | 94 | [10] |
| Cinnamomum camphora Nees & Eberm var. linaloolifera | Wood | 90 | [11] |
| Coriandrum sativum L. | Seeds | 83 | [12] |
| Orthodon linalooliferum Fujita | Aerial parts | 82 | [13] |
| Coriandrum sativum L. | Mericarp | 79 | [14] |
| Zanthoxylum alatum Roxb | Seeds | 71 | [15] |
| Coriandrum sativum L. | Seeds | 66 | [16] |
| Lavandula angustifolia | Whole plant | 54 | [17] |
| Lippia alba | Leaves | 53 | [18] |
| Hyssopus officinalis | Aerial parts | 51 | [19] |
| Lavandula latifolia | Whole plant | 51 | [14] |
| Strain | Activity | Reference |
|---|---|---|
| Gram-Positive Bacteria | ||
| Micrococcus flavus ATCC 9341 | 4 µg/mL | [61] |
| Bacillus subtilis ATCC 10707 | 4 µg/mL | [61] |
| Staphylococcus epidermidis ATCC 12228 | 4 µg/mL | [61] |
| Staphylococcus aureus ATCC 25923 | 5 µg/mL | [61] |
| Staphylococcus aureus NCTC 10788 | 13 mm | [63] |
| Listeria monocytogenes | 5 µg/mL | [61] |
| Listeria innocua NRRLB-33314 | 1% | [55] |
| Gram-Negative Bacteria | ||
| Salmonella enterica subsp. enterica serovar Enteritidis ATCC 13076 | 5 µg/mL | [61] |
| Salmonella enterica subsp. enterica serovar Typhimurium ATCC 13311 | 5 µg/mL | [61] |
| Salmonella enterica subsp. enterica serovar Typhimurium CCM 5445 | 0.7% | [55] |
| Vibrio fluvialis L-15318 | 32 mM | [57] |
| Salmonella enterica subsp. enterica serovar Typhi ATCC 14901 | 7.5 mM | [57] |
| E. coli ATCC 25922 | 212 µM | [57] |
| E. coli NCTC 12923 | 21 mm | [63] |
| E. coli ATCC O157:H7 | 6 µg/mL (0.6%) | [55,61] |
| Pseudomonas aeruginosa NCTC 12924 | 8 mm | [63] |
| Pseudomonas aeruginosa ATCC 27853 | 7 µg/mL | [61] |
| Pseudomonas aeruginosa ATCC 9027 | 431 µg/mL | [56] |
| Pseudomonas fluorescens ATCC 13525 | 1.25 µg/mL | [64] |
| Enterobacter cloacae (h.i.) | 6 µg/mL | [61] |
| Proteus mirabilis (h.i.) | 6 µg/mL | [61] |
| Campylobacter jejuni ATCC 33560 | 0.5 µL/mL | [59] |
| Campylobacter coli ATCC 33559 | 0.5 µL/mL | [59] |
| Serratia liquefaciens NRRLB-951 | 1% | [55] |
| Porphyromonas gingivalis ATCC 33277 | 0.8 mg/mL | [60] |
| Porphyromonas gingivalis ATCC 49417, ATCC 53978 | 0.1 mg/mL | [60] |
| Prevotella intermedia ATCC 25611 | 0.2 mg/mL | [60] |
| Prevotella intermedia ATCC 49046 | 1.6 mg/mL | [60] |
| Prevotella nigrescens ATCC 25261, ATCC 33563 | 0.8 mg/mL | [60] |
| Fusobacterium nucleatum subsp. nucletum ATCC 25586 | 0.2 mg/mL | [60] |
| Fusobacterium nucleatum subsp. polymorphum ATCC 10953 | 0.2 mg/mL | [60] |
| Fusobacterium nucleatum subsp. vincenti ATCC 49046 | 0.1 mg/mL | [60] |
| Fusobacterium nucleatum subsp. fusiforme ATCC 51190 | 0.2 mg/mL | [60] |
| Fusobacterium nucleatum subsp. animals ATCC 51191 | 0.2 mg/mL | [60] |
| Aggregatibacter actinomycetemcomitans ATCC 33384, ATCC 43717, ATCC 43718 | 0.1 mg/mL | [60] |
| Parameter | Linalool | Effect |
|---|---|---|
| Alkaline phosphatase (ā) |  | Destruction of the integrity of the cell wall |
| Histidine and methionine |  | Damage to the structure of the biofilm |
| L-glutamate, L-glutamine, L-arginine, L-aspartate |  | Disturbance of the pH homeostasis of the bacterial cell |
| Parameter | linalool | Effect |
|---|---|---|
| Pyruvate, Citrate, Fumarate, Lactate, 2-Okoglutarate | | ATPase disturbance ≥ disturbance of energy production ≥ abnormal TCA cycle and glycolysis |
| Pyruvate kinase (ā) |  | Disrupted TCA cycle |
| Succinate dehydrogenase (ā) | | Disturbance of oxidative phosphorylation |
| Parameter | Linalool | Effect |
|---|---|---|
| fatty acids: linolenic, linoleic, oleic, arachidonic, myristic, octadecadiene, palmitic |  | Low fluidity of the cell membrane |
| Phosphatidylcholine, phosphatidylethanolamine |  binding | Damage to the structure of the cell membrane |
| Strain | Activity of Linalool | Referece |
|---|---|---|
| Candida albicans NCPF3179 | 12 mm | [63] |
| Candida albicans ATCC 14053 | 8 mM | [73] |
| Candida glabrata (c.s.) | 16 mM | [73] |
| Candida tropicalis ATCC 13803 | 125 µg/mL | [74] |
| Candida tropicalis (c.s.) | 16 mM | [73] |
| Trichophyton rubrum LM 305 | 256 µg/mL | [67] |
| Microsporum canis (c.s.—LM910, LM82, LM29) | 128 µg/mL | [75] |
| Microsporum gypseum (c.s.—LM512, LM130) | 128 µg/mL | [75] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mączka, W.; Duda-Madej, A.; Grabarczyk, M.; Wińska, K. Natural Compounds in the Battle against Microorganisms—Linalool. Molecules 2022, 27, 6928. https://doi.org/10.3390/molecules27206928
Mączka W, Duda-Madej A, Grabarczyk M, Wińska K. Natural Compounds in the Battle against Microorganisms—Linalool. Molecules. 2022; 27(20):6928. https://doi.org/10.3390/molecules27206928
Chicago/Turabian StyleMączka, Wanda, Anna Duda-Madej, Małgorzata Grabarczyk, and Katarzyna Wińska. 2022. "Natural Compounds in the Battle against Microorganisms—Linalool" Molecules 27, no. 20: 6928. https://doi.org/10.3390/molecules27206928