
Screening and Characteristics of Marine Bacillus velezensis Z-1 Protease and Its Application of Enzymatic Hydrolysis of Mussels to Prepare Antioxidant Active Substances
 ,
,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Strain Screen and Identification
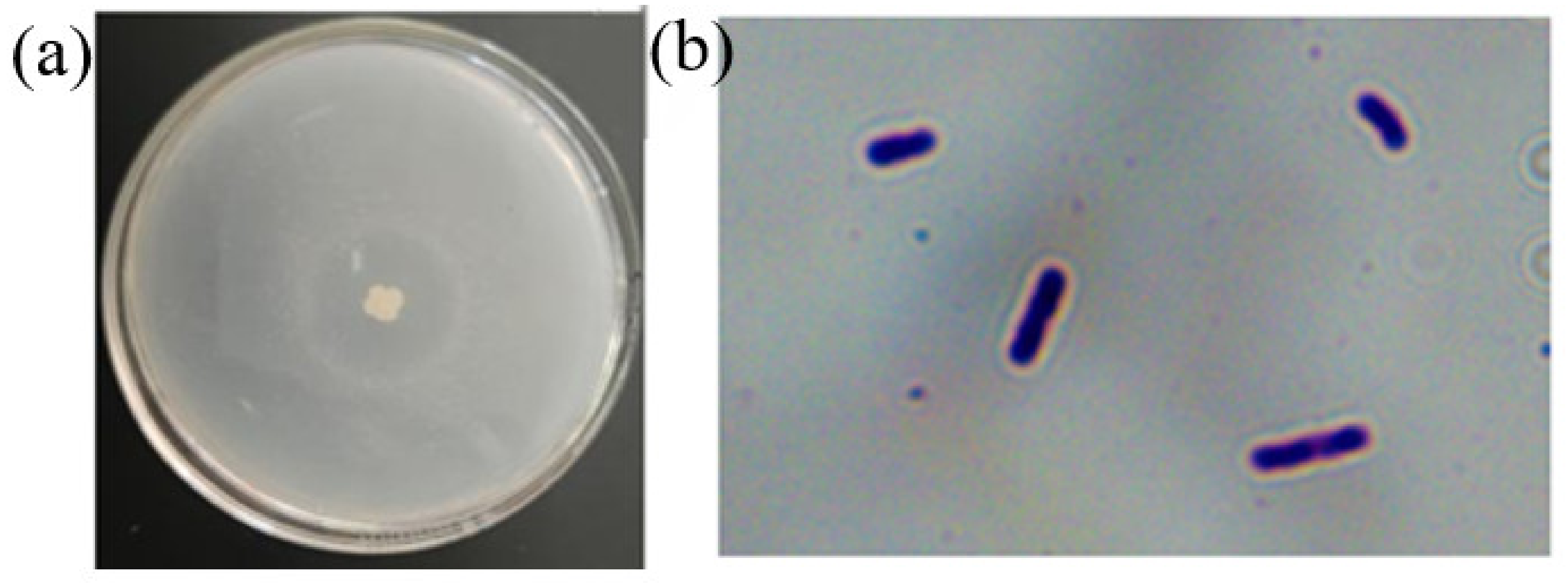
2.1.1. Screening of Strains
2.1.2. Morphological Biological Identification of Strain Z-1
2.1.3. Physiological and Biochemical Characteristics of Strain Z-1
2.1.4. Molecular Biological Identification of Strain Z-1
2.2. Optimization Analysis of Protease Production by Fermentation of Strain Z-1
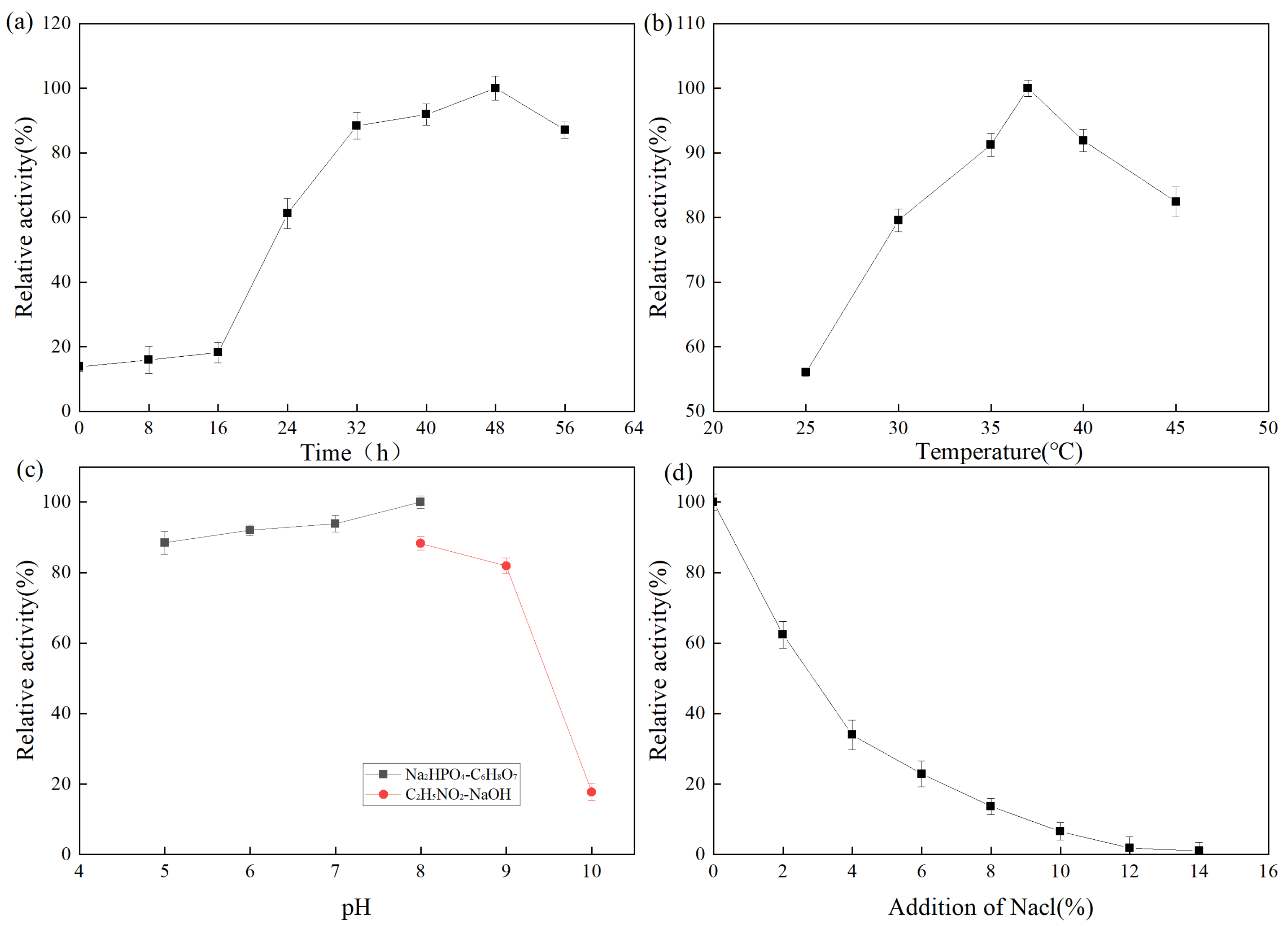
2.2.1. Analysis of Single Factor Experiment Results
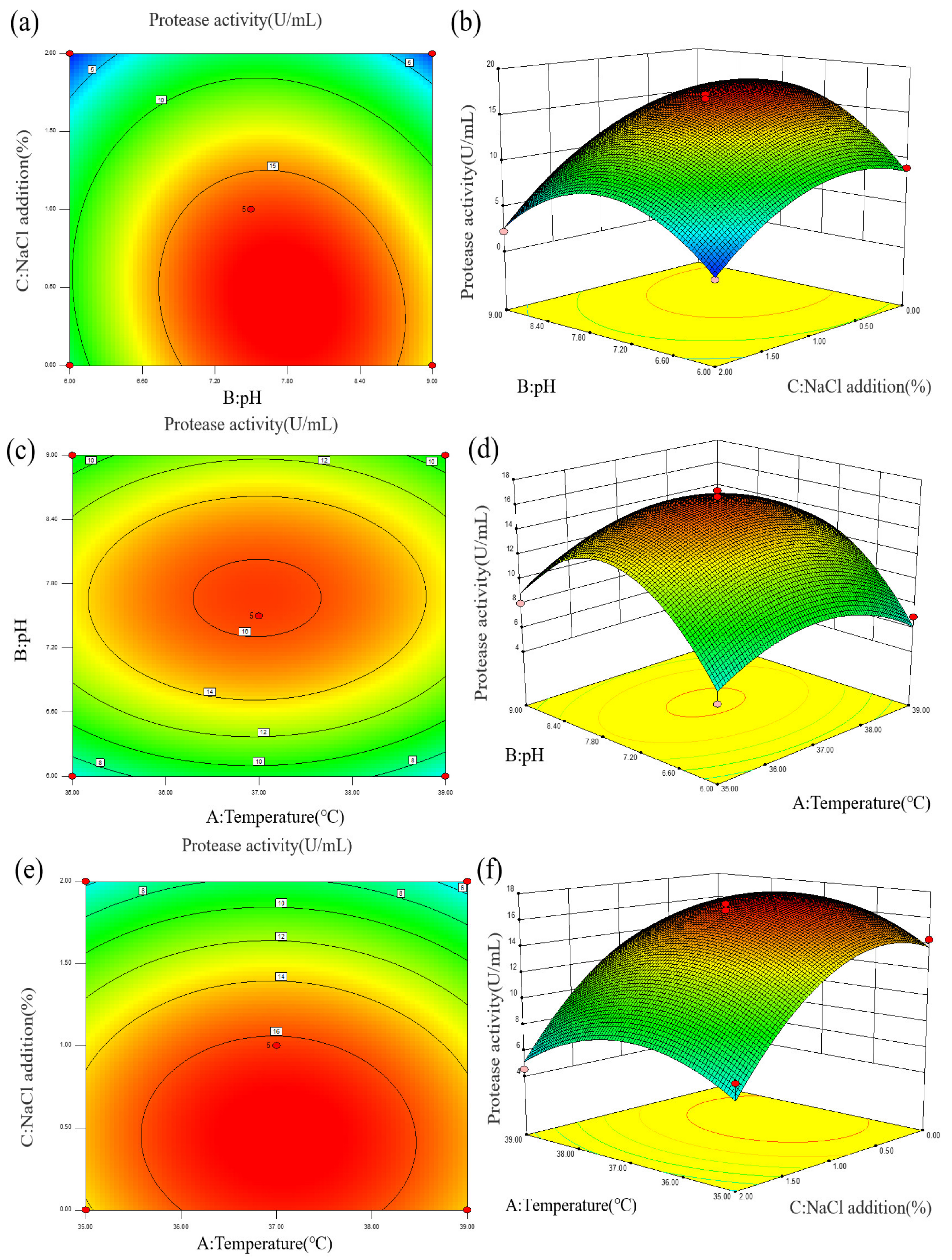
2.2.2. Analysis of Results of Enzyme Production Optimized by Response Surface Methodology
- (1)
- Model establishment and variance analysis
- (2)
- Response surface analysis
- (3)
- Verification experiment
2.3. Enzymatic Properties of Strain Z-1 Protease
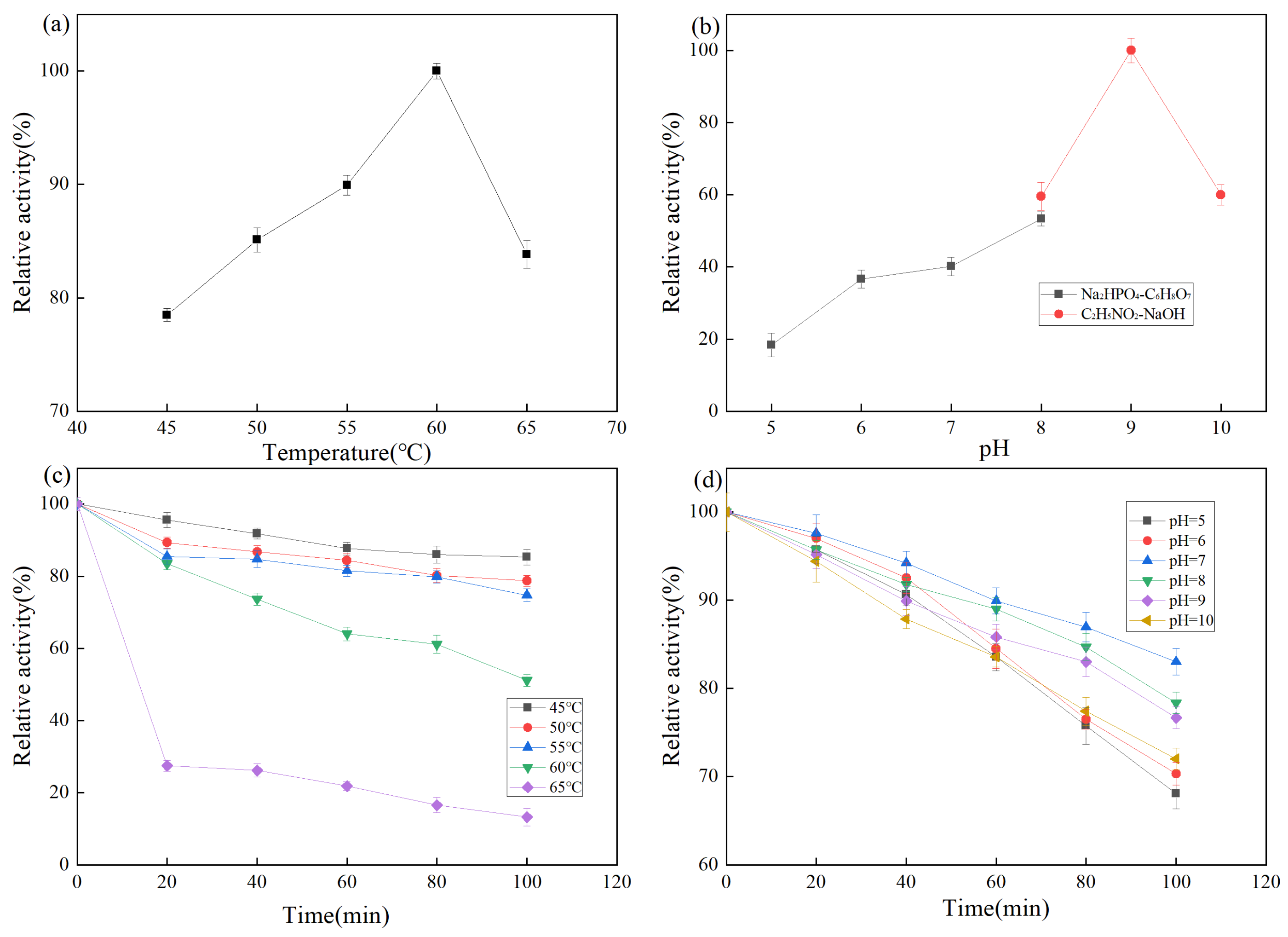
2.3.1. Effects of Temperature on Protease Activity of Strain Z-1
2.3.2. Effects of pH on Protease Activity of Strain Z-1
2.3.3. Effects of Metal Ions on Protease Activity of Strain Z-1
2.3.4. Effects of Reducing Agent, Denaturant, Organic Solvent and Other Compounds on Protease Activity
2.3.5. Zymography of B. velezensis Z-1 extracellular proteases
2.4. Oxidation Resistance Analysis of Mussel Enzymolysis Product Y1
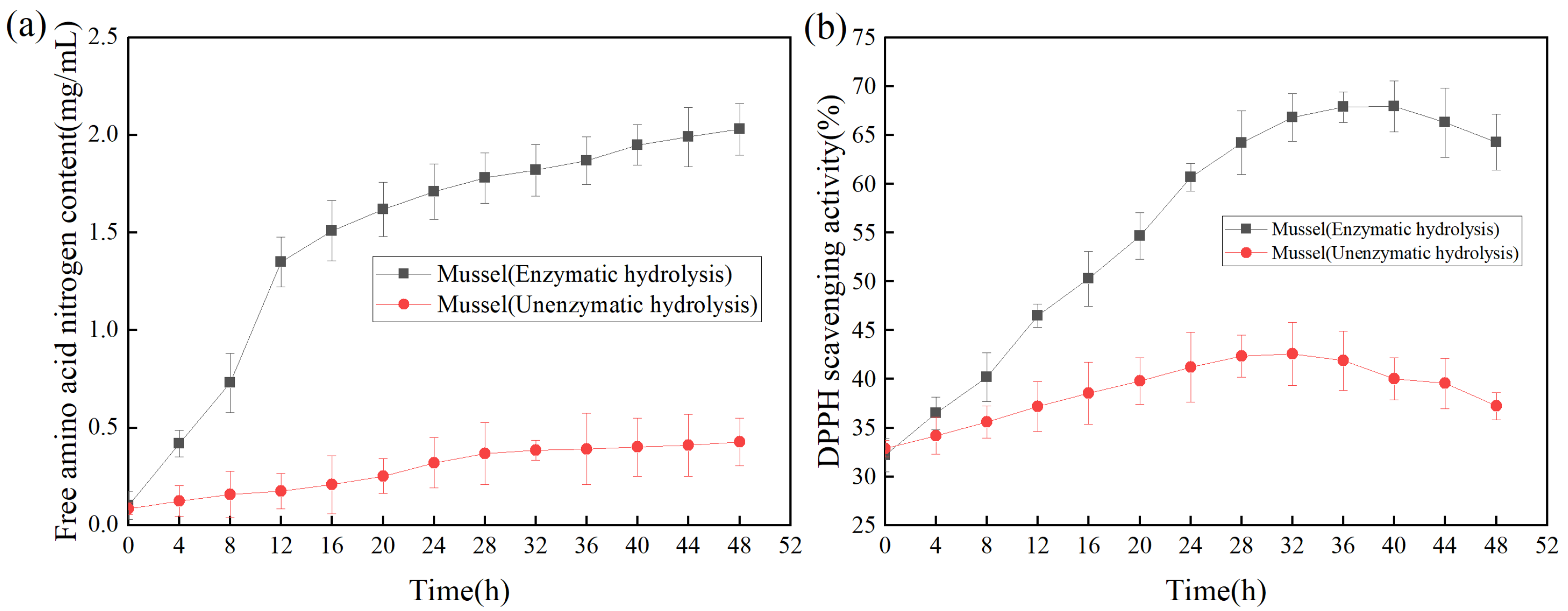
2.4.1. Effect of Hydrolysis Time on Antioxidant Activity of Mussels
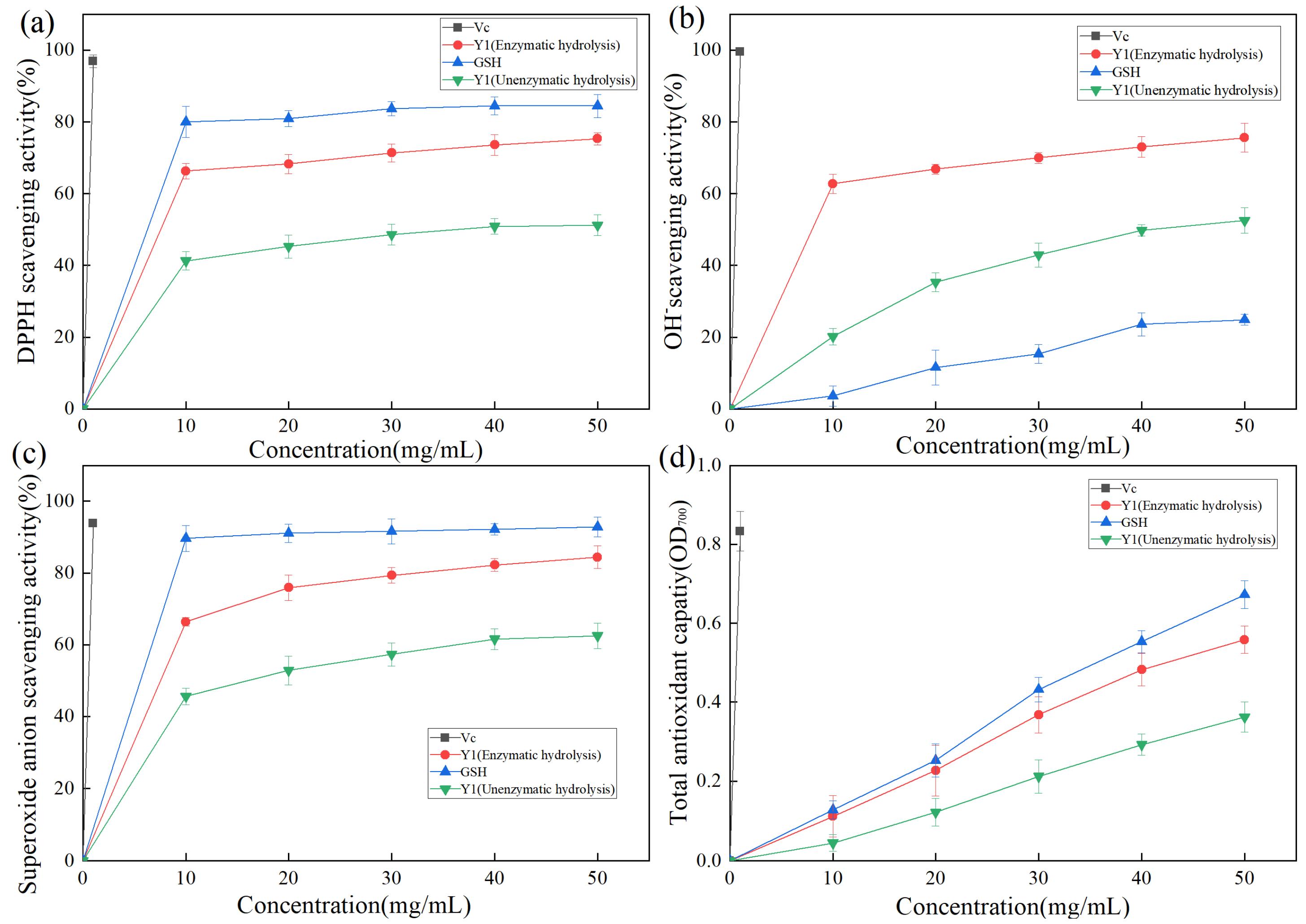
2.4.2. Analysis of DPPH Radical Scavenging Activity of Enzymolysis Products
2.4.3. Analysis of Hydroxyl Radical Scavenging Activity of Enzymolysis Products
2.4.4. Analysis of Superoxide Anion Radical Scavenging Activity of Enzymolysis Products
2.4.5. Reduction Force Analysis of Enzymolysis Products
2.4.6. Comprehensive Application Analysis of Antioxidant Activity of Mussel Enzymolysis Products
3. Materials and Methods
3.1. Samples and Medium
3.2. Screening of Protease Producing Strains
3.3. Determination of Protease Activity
3.4. Biomorphological Identification of B. velezensis Z-1
3.5. Optimization of Protease Production by Fermentation of B. velezensis Z-1
3.6. Characterization of Enzymatic Properties of B. velezensis Protease
3.7. Preparation and Separation of Mussel Enzymolysis Products by B. velezensis Protease
3.7.1. Determination of Free Amino Acid Nitrogen
3.7.2. Effect of Enzymolysis Time on Antioxidant Activity
3.7.3. Preparation and Separation of Antioxidants
3.8. Evaluation of Antioxidant Activity of Enzymolysis Products by B. velezensis Protease
3.9. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Isildak, O.; Yildiz, I.; Genc, N. A new potentiometric PVC membrane sensor for the determination of DPPH radical scavenging activity of plant extracts. Food Chem. 2022, 373, 131420. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Jing, X.; Chen, Z.; Wang, X. Purification and identification of antioxidant peptides from millet gliadin treated with high hydrostatic pressure. LWT 2022, 164, 113654. [Google Scholar] [CrossRef]
- Lv, R.; Dong, Y.; Bao, Z.; Zhang, S.; Lin, S.; Sun, N. Advances in the activity evaluation and cellular regulation pathways of food-derived antioxidant peptides. Trends Food Sci. Technol. 2022, 122, 171–186. [Google Scholar] [CrossRef]
- Davis, C.K.; Vemuganti, R. Antioxidant therapies in traumatic brain injury. Neurochem. Int. 2022, 152, 105255. [Google Scholar] [CrossRef] [PubMed]
- Obulesu, M. Chapter 3—Antioxidants in Alzheimer’s Therapy. In Alzheimer’s Disease Theranostics; Obulesu, M., Ed.; Academic Press: San Francisco, CA, USA, 2019; pp. 13–18. [Google Scholar]
- Cunha, S.A.; Pintado, M.E. Bioactive peptides derived from marine sources: Biological and functional properties. Trends Food Sci. Technol. 2022, 119, 348–370. [Google Scholar] [CrossRef]
- Park, J.S.; Shin, S.K.; Wu, H.; Yarish, C.; Yoo, H.I.; Kim, J.K. Evaluation of nutrient bioextraction by seaweed and shellfish aquaculture in Korea. J. World Aquacult. Soc. 2021, 52, 1118–1134. [Google Scholar] [CrossRef]
- Kang, X.; Zhao, Y.; Tan, Z.; Ning, J.; Zhai, Y.; Zheng, G. Evaluation of multivariate data analysis for marine mussels Mytilus edulis authentication in China: Based on stable isotope ratio and compositions of C, N, O and H. J. Food Compos. Anal. 2022, 111, 1–9. [Google Scholar] [CrossRef]
- Naik, A.S.; Hayes, M. Bioprocessing of mussel by-products for value added ingredients. Trends Food Sci. Technol. 2019, 92, 111–121. [Google Scholar] [CrossRef]
- Breternitz, N.R.; Bolini, H.M.A.; Hubinger, M.D. Sensory acceptance evaluation of a new food flavoring produced by microencapsulation of a mussel (Perna perna) protein hydrolysate. LWT 2017, 83, 141–149. [Google Scholar] [CrossRef]
- Zhang, L.; Song, C.; Chang, J.; Wang, Z.; Meng, X. Optimization of protein hydrolysates production from defatted peanut meal based on physicochemical characteristics and sensory analysis. LWT 2022, 163, 113572. [Google Scholar] [CrossRef]
- Pan, F.-D.; Liu, S.; Xu, Q.-M.; Chen, X.-Y.; Cheng, J.-S. Bioconversion of kitchen waste to surfactin via simultaneous enzymolysis and fermentation using mixed-culture of enzyme- producing fungi and Bacillus amyloliquefaciens HM618. Biochem. Eng. J. 2021, 172, 108036. [Google Scholar] [CrossRef]
- Rajapakse, N.; Mendis, E.; Jung, W.-K.; Je, J.-Y.; Kim, S.-K. Purification of a radical scavenging peptide from fermented mussel sauce and its antioxidant properties. Food Res. Int. 2005, 38, 175–182. [Google Scholar] [CrossRef]
- Zhi, T.; Li, X.; Sadiq, F.A.; Mao, K.; Gao, J.; Mi, S.; Liu, X.; Deng, W.; Chitrakar, B.; Sang, Y. Novel antioxidant peptides from protein hydrolysates of scallop (Argopecten irradians) mantle using enzymatic and microbial methods: Preparation, purification, identification and characterization. LWT 2022, 164, 113636. [Google Scholar] [CrossRef]
- Li, J.; Wu, S.; Tian, X.; Li, X. Fabrication of a multifunctional nanomaterial from a mussel-derived peptide for multimodal synergistic cancer therapy. Chem. Eng. J. 2022, 446, 136837. [Google Scholar] [CrossRef]
- Zeng, Z.; Wang, Y.; Anwar, M.; Hu, Z.; Wang, C.; Lou, S.; Li, H. Molecular cloning and expression analysis of mytilin-like antimicrobial peptides from Asian green mussel Perna viridis. Fish Shellfish Immunol. 2022, 121, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Novoa, B.; Estepa, A.; Figueras, A. Antimicrobial peptides from mussel Mytilus galloprovincialis: The myticins. Fish Shellfish Immunol. 2016, 53, 1–8. [Google Scholar] [CrossRef]
- Grienke, U.; Silke, J.; Tasdemir, D. Bioactive compounds from marine mussels and their effects on human health. Food Chem. 2014, 142, 48–60. [Google Scholar] [CrossRef]
- Je, J.Y.; Park, P.J.; Byun, H.G.; Jung, W.K.; Kim, S.K. Angiotensin I converting enzyme (ACE) inhibitory peptide derived from the sauce of fermented blue mussel, Mytilus edulis. Bioresour. Technol. 2005, 96, 1624–1629. [Google Scholar] [CrossRef] [PubMed]
- Ge, Z.; Kuang, Z.; Chen, J.; Chen, J.; Liu, T.; She, Z.; Lu, Y. Comparative genomics analysis of Bacillus velezensis LOH112 isolated from a nonagenarian provides insights into its biocontrol and probiotic traits. Gene 2022, 1, 146644. [Google Scholar] [CrossRef]
- Liu, G.; Kong, Y.; Fan, Y.; Geng, C.; Peng, D.; Sun, M. Whole-genome sequencing of Bacillus velezensis LS69, a strain with a broad inhibitory spectrum against pathogenic bacteria. J. Biotechnol. 2017, 249, 20–24. [Google Scholar] [CrossRef]
- Li, X.; Gao, X.; Zhang, S.; Jiang, Z.; Yang, H.; Liu, X.; Jiang, Q.; Zhang, X. Characterization of a Bacillus velezensis with antibacterial activity and inhibitory effect on common aquatic pathogens. Aquaculture 2020, 523, 735165. [Google Scholar] [CrossRef]
- Jiang, C.-H.; Liao, M.-J.; Wang, H.-K.; Zheng, M.-Z.; Xu, J.-J.; Guo, J.-H. Bacillus velezensis, a potential and efficient biocontrol agent in control of pepper gray mold caused by Botrytis cinerea. Biological. Control. 2018, 126, 147–157. [Google Scholar] [CrossRef]
- Liu, Y.; Guo, H.; Gu, J.; Qin, W. Optimize purification of a cellulase from Bacillus velezensis A4 by aqueous two-phase system (ATPS) using response surface methodology. Process Biochem. 2019, 87, 196–203. [Google Scholar] [CrossRef]
- Bhatt, K.; Lal, S.; Srinivasan, R.; Joshi, B. Molecular analysis of Bacillus velezensis KB 2216, purification and biochemical characterization of alpha-amylase. Int. J. Biol. Macromol. 2020, 164, 3332–3339. [Google Scholar] [CrossRef]
- Lekshmi, R.; Arif Nisha, S.; Kaleeswaran, B.; Alfarhan, A.H. Pomegranate peel is a low-cost substrate for the production of tannase by Bacillus velezensis TA3 under solid state fermentation. J. King Saud Univ.-Sci. 2020, 32, 1831–1837. [Google Scholar] [CrossRef]
- Ghosh, A.; Chandra, A.; Dhar, A.; Shukla, P.; Baishya, D. Multi-efficient thermostable endoxylanase from Bacillus velezensis AG20 and its production of xylooligosaccharides as efficient prebiotics with anticancer activity. Process Biochem. 2021, 109, 59–71. [Google Scholar] [CrossRef]
- Zeng, J.; Huang, W.; Tian, X.; Hu, X.; Wu, Z. Brewer’s spent grain fermentation improves its soluble sugar and protein as well as enzymatic activities using Bacillus velezensis. Process Biochem. 2021, 111, 12–20. [Google Scholar] [CrossRef]
- Li, Y.; Ruan, S.; Zhou, A.; Xie, P.; Azam, S.M.R.; Ma, H. Ultrasonic modification on fermentation characteristics of Bacillus varieties: Impact on protease activity, peptide content and its correlation coefficient. LWT 2022, 154, 1–7. [Google Scholar] [CrossRef]
- Qiu, Y.; Yan, H.-H.; Sun, S.-M.; Wang, Y.-Q.; Zhao, X.-R.; Wang, H.-Y. Use of Bacillus velezensis SDTB022 against tobacco black shank (TBS) and the biochemical mechanism involved. Biol. Control 2022, 165, 104785. [Google Scholar] [CrossRef]
- Chen, W.; Li, L.; Ye, C.; Zhao, Z.; Huang, K.; Zou, D.; Wei, X. Efficient production of extracellular alkaline protease in Bacillus amyloliquefaciens by host strain construction. LWT 2022, 163, 113620. [Google Scholar] [CrossRef]
- Mukhia, S.; Kumar, A.; Kumar, R. Generation of antioxidant peptides from soy protein isolate through psychrotrophic Chryseobacterium sp. derived alkaline broad temperature active protease. LWT 2021, 143, 111152. [Google Scholar] [CrossRef]
- Kumar, A.; Mukhia, S.; Kumar, N.; Acharya, V.; Kumar, S.; Kumar, R. A Broad Temperature Active Lipase Purified From a Psychrotrophic Bacterium of Sikkim Himalaya With Potential Application in Detergent Formulation. Front. Bioeng. Biotechnol. 2020, 8, 642. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, K.S.; Ertan, H.; Poljak, A.; Bridge, W.J. Evaluating Enzymatic Productivity-The Missing Link to Enzyme Utility. Int. J. Mol. Sci. 2022, 23, 6908. [Google Scholar] [CrossRef] [PubMed]
- Nikhita, R.; Sachindra, N.M. Optimization of chemical and enzymatic hydrolysis for production of chicken blood protein hydrolysate rich in angiotensin-I converting enzyme inhibitory and antioxidant activity. Poult. Sci. 2021, 100, 101047. [Google Scholar] [CrossRef]
- Babini, E.; Tagliazucchi, D.; Martini, S.; Dei Piu, L.; Gianotti, A. LC-ESI-QTOF-MS identification of novel antioxidant peptides obtained by enzymatic and microbial hydrolysis of vegetable proteins. Food Chem. 2017, 228, 186–196. [Google Scholar] [CrossRef]
- Chen, X.; Liang, L.; Han, C. Borate suppresses the scavenging activity of gallic acid and plant polyphenol extracts on DPPH radical: A potential interference to DPPH assay. LWT 2020, 131, 109769. [Google Scholar] [CrossRef]
- Guo, X.; Dong, Z.; Li, Q.; Wan, D.; Zhong, J.; Dongzhi, D.; Huang, M. Flavonoids from Rhododendron nivale Hook. f delay aging via modulation of gut microbiota and glutathione metabolism. Phytomedicine 2022, 104, 154270. [Google Scholar] [CrossRef]
- Wu, S. Glutathione suppresses lipid oxidation of Clannish bilineata larvae meat during frozen storage. Entomol. Res. 2018, 48, 451–454. [Google Scholar] [CrossRef]
- Karunakaran, U.; Elumalai, S.; Moon, J.S.; Won, K.C. c-Abl Tyrosine Kinase Inhibition Attenuate Oxidative Stress-Induced Pancreatic beta-Cell Dysfunction via Glutathione Antioxidant System. Transl. Res. 2022, 6, 1–7. [Google Scholar] [CrossRef]
- Leung, R.; Venus, C.; Zeng, T.; Tsopmo, A. Structure-function relationships of hydroxyl radical scavenging and chromium-VI reducing cysteine-tripeptides derived from rye secalin. Food Chem. 2018, 254, 165–169. [Google Scholar] [CrossRef]
- Lv, J.-Y.; Nawaz, M.A.H.; Liu, N.; Zhou, H.-P.; Hussain, E.; Wen, X.; Gou, X.-Y.; Jin, X.; Yu, C. A Nile red-based near-infrared fluorescent probe for the detection of superoxide radical anion in living cells. Chin. J. Anal. Chem. 2022, 50, 1–9. [Google Scholar] [CrossRef]
- Wang, B.; Li, L.; Chi, C.F.; Ma, J.H.; Luo, H.Y.; Xu, Y.F. Purification and characterisation of a novel antioxidant peptide derived from blue mussel (Mytilus edulis) protein hydrolysate. Food Chem. 2013, 138, 1713–1719. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Feng, Y.; Lai, X.; Deng, T.; Liu, X.; Lyu, M.; Wang, S. Virgibacillus halodenitrificans ST-1 for fermentation of shrimp paste and hydrolysates of its protease. Food Sci. Nutr. 2020, 8, 5352–5361. [Google Scholar] [CrossRef] [PubMed]
- Hassabo, A.A.; Selim, M.H.; Saad, M.M.; Abdelraof, M. Optimization of l-methioninase and l-arginase production by newly isolated marine yeast using response surface methodology. Biocatal. Agric. Biotechnol. 2022, 42, 1–9. [Google Scholar] [CrossRef]
- Zhu, X.; Hua, Y.; Li, X.; Kong, X.; Zhang, C.; Chen, Y. Isolation and characterization of an activator-dependent protease from Aspergillus ochraceus screened from low denatured defatted soybean meal and the proteolysis of soy proteins. LWT 2021, 150, 112026. [Google Scholar] [CrossRef]
- Olena, Z.; Yang, Y.; TingTing, Y.; XiaoTao, Y.; HaiLian, R.; Xun, X.; Dong, X.; CuiLing, W.; HaiLun, H. Simultaneous preparation of antioxidant peptides and lipids from microalgae by pretreatment with bacterial proteases. Bioresour. Technol. 2022, 348, 126759. [Google Scholar] [CrossRef]
- Mudgil, P.; Baba, W.N.; Kamal, H.; FitzGerald, R.J.; Hassan, H.M.; Ayoub, M.A.; Gan, C.Y.; Maqsood, S. A comparative investigation into novel cholesterol esterase and pancreatic lipase inhibitory peptides from cow and camel casein hydrolysates generated upon enzymatic hydrolysis and in-vitro digestion. Food Chem. 2022, 367, 130661. [Google Scholar] [CrossRef]
- Hu, J.; Tian, X.; Wei, T.; Wu, H.; Lu, J.; Lyu, M.; Wang, S. Anti-Helicobacter pylori Activity of a Lactobacillus sp. PW-7 Exopolysaccharide. Foods 2021, 10, 2453. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Project | Result | Project | Result |
|---|---|---|---|
| Gram stain | + | Urea | + |
| 7% NaCl salt tolerance | + | Glucose | + |
| PH 5.7 growth | + | Sucrose | + |
| V-P | + | Maltose | − |
| Gelatin liquefaction | + | Cellobiose | − |
| Amylase hydrolysis | + | Lactose | − |
| Nitrate reduction | + | Gum sugar | + |
| H2S gas production | − | Xylose | + |
| Methyl red | − | Aescin | + |
| Propionate | − | Salicin | − |
| Mannitol | + | Ornithine decarboxylase | − |
| Sorbitol Indole | + − | Lysine decarboxylase | − |
| Metal Ion | Relative Enzyme Activity (%) (5 mM) | Relative Enzyme Activity (%) (25 mM) |
|---|---|---|
| Blank | 100 ± 0.43 | 100 ± 0.64 |
| Ca2+ | 102.44 ± 1.62 | 91.43 ± 2.72 |
| K+ | 82.58 ± 1.32 | 93.30 ± 3.20 |
| Na+ | 56.73 ± 2.63 | 99.39 ± 0.68 |
| Zn2+ | 51.83 ± 1.02 | 5.94 ± 0.84 |
| Mg2+ | 72.10 ± 0.26 | 106.45 ± 1.61 |
| Ni2+ | 71.67 ± 2.89 | 43.63 ± 2.63 |
| Ba2+ | 65.93 ± 2.36 | 104.47 ± 0.57 |
| Sr2+ | 74.22 ± 1.28 | 98.70 ± 2.40 |
| NH4+ | 71.88 ± 1.60 | 135.91 ± 1.39 |
| Li+ | 69.86 ± 0.57 | 103.73 ± 2.38 |
| Mn2+ | 89.12 ± 0.80 | 147.18 ± 0.93 |
| Reagent Type | Reagent Name | Final Concentration | Relative Enzyme Activity(%) |
|---|---|---|---|
| Control | Ultrapure water | - | 100 ± 0.68 |
| Reductant | Glutathione | 10 mM | 91.32 ± 1.58 |
| β-Mercaptoethanol | 10 mM | 89.41 ± 1.22 | |
| Cysteine | 10 mM | 98.63 ± 2.12 | |
| Denaturant | SDS | 0.10% | 71.00 ± 0.48 |
| Urea | 4 M | 78.32 ± 1.65 | |
| Protease Inhibitor | EDTA | 5 mM | 79.27 ± 2.56 |
| PMSF | 5 mM | 52.75 ± 0.78 | |
| Surfactant | TritonX-100 | 0.10% | 77.83 ± 1.50 |
| Tween80 | 0.10% | 119.89 ± 2.61 | |
| Organic solvent | Methyl alcohol | 10% | 113.24 ± 1.55 |
| Ethyl acetate | 10% | 45.43 ± 1.48 | |
| Dimethyl sulfoxide | 10% | 111.58 ± 2.87 | |
| Ethanol | 10% | 104.31 ± 0.45 | |
| Ethanol | 20% | 29.90 ± 1.23 | |
| Ethanol | 30% | 21.33 ± 0.84 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, J.; Zhao, Y.; Hu, R.; Cheng, Y.; Qin, J.; Yang, J.; Fang, Y.; Lyu, M.; Wang, S. Screening and Characteristics of Marine Bacillus velezensis Z-1 Protease and Its Application of Enzymatic Hydrolysis of Mussels to Prepare Antioxidant Active Substances. Molecules 2022, 27, 6570. https://doi.org/10.3390/molecules27196570
Lu J, Zhao Y, Hu R, Cheng Y, Qin J, Yang J, Fang Y, Lyu M, Wang S. Screening and Characteristics of Marine Bacillus velezensis Z-1 Protease and Its Application of Enzymatic Hydrolysis of Mussels to Prepare Antioxidant Active Substances. Molecules. 2022; 27(19):6570. https://doi.org/10.3390/molecules27196570
Chicago/Turabian StyleLu, Jing, Yu Zhao, Rong Hu, Yu Cheng, Junhuan Qin, Jie Yang, Yaowei Fang, Mingsheng Lyu, and Shujun Wang. 2022. "Screening and Characteristics of Marine Bacillus velezensis Z-1 Protease and Its Application of Enzymatic Hydrolysis of Mussels to Prepare Antioxidant Active Substances" Molecules 27, no. 19: 6570. https://doi.org/10.3390/molecules27196570