
Vasorelaxant Effect of Trachelospermi caulis Extract on Rat Mesenteric Resistance Arteries


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
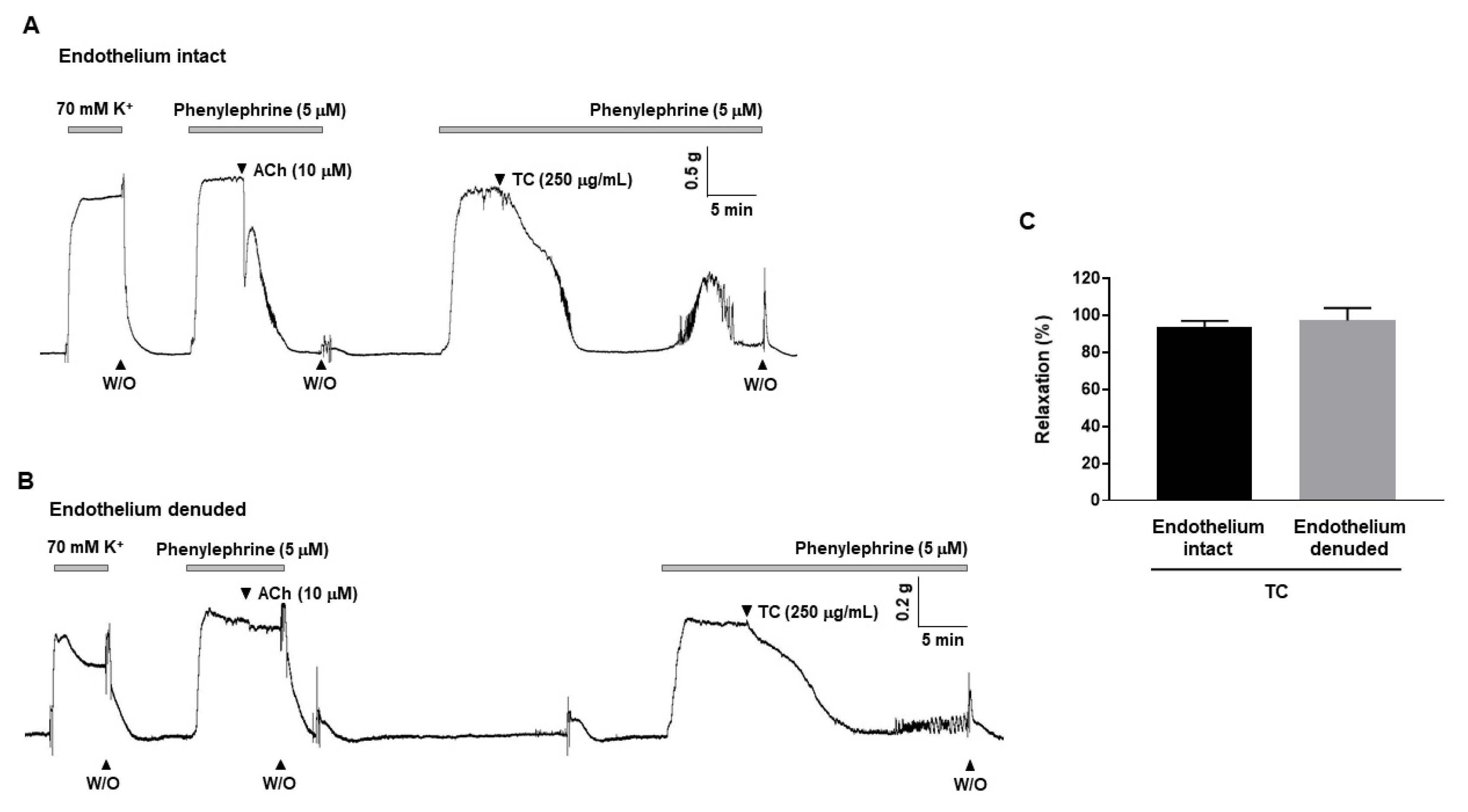
2.1. Effect of T. caulis on Contraction Induced by KCl or PE in Rat Mesenteric Arteries
2.2. T. caulis-Induced Endothelium-Independent Relaxation in Rat Mesenteric Arteries
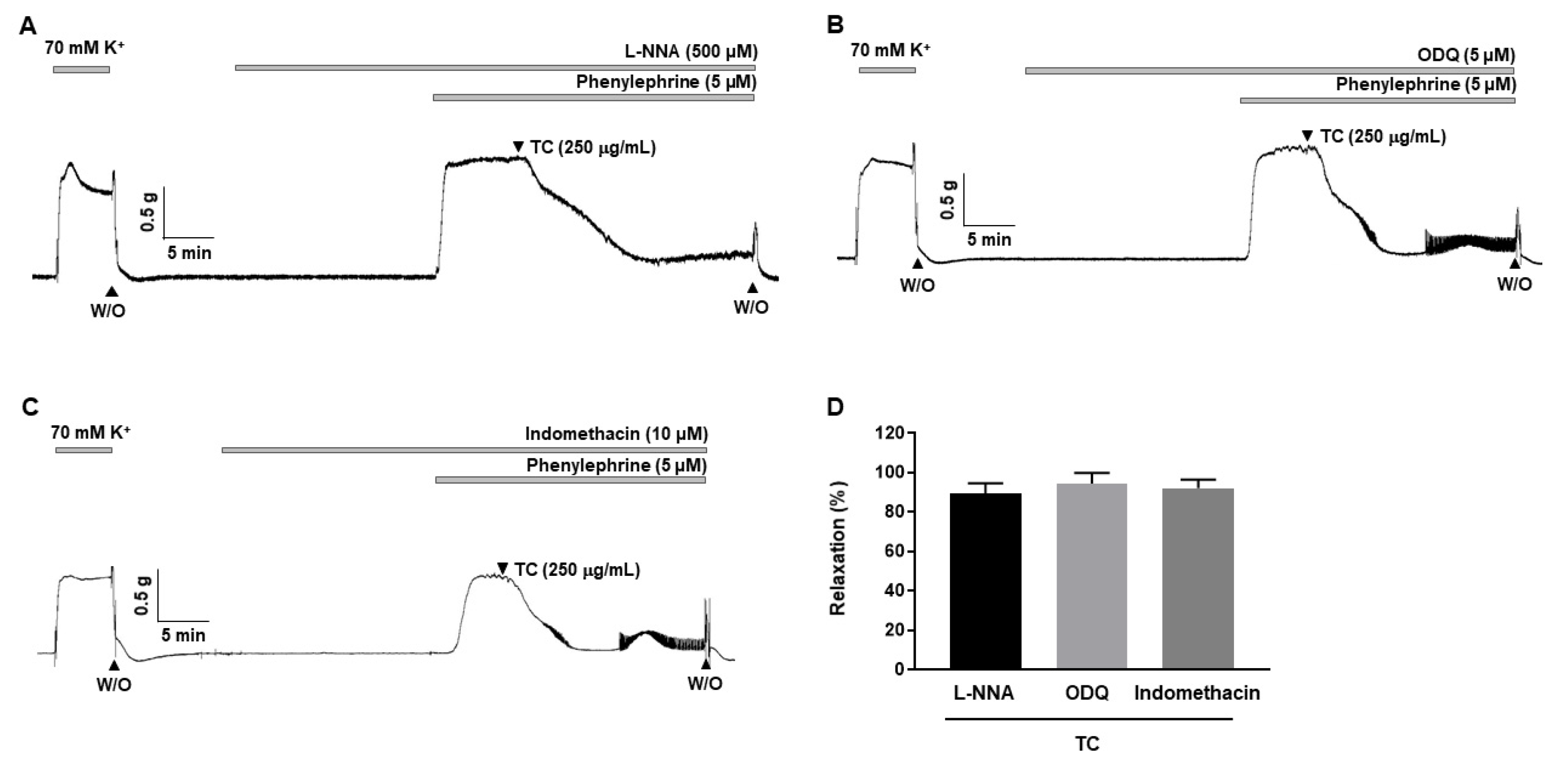
2.3. Effects of L-NNA, Indomethacin and ODQ on T. caulis-Induced Vasodilation
2.4. Effects of K+ Channel Blockers on T. caulis Extract-Induced Vascular Relaxation
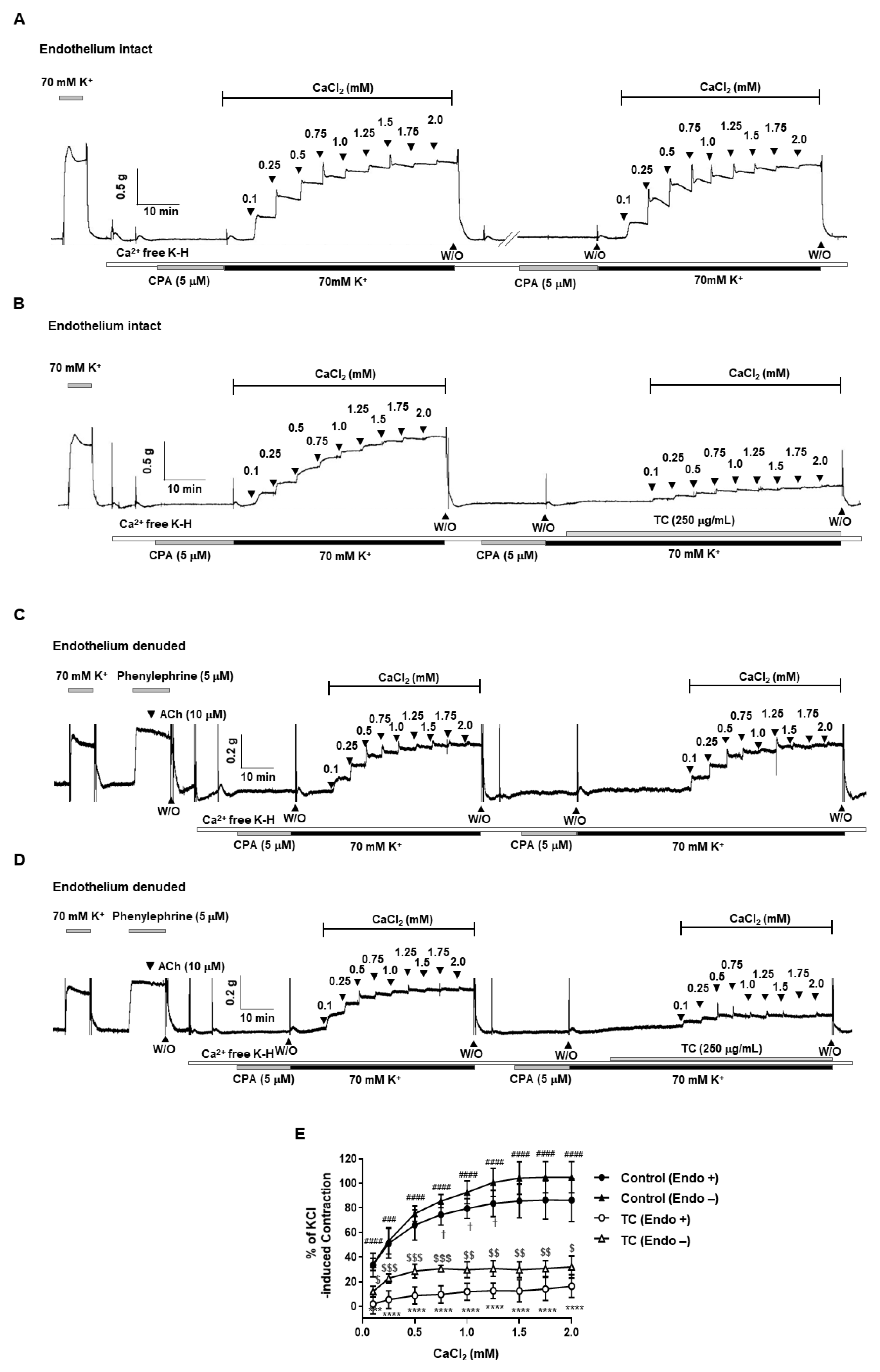
2.5. Effect of T. caulis on Extracellular Ca2+-Induced Contraction
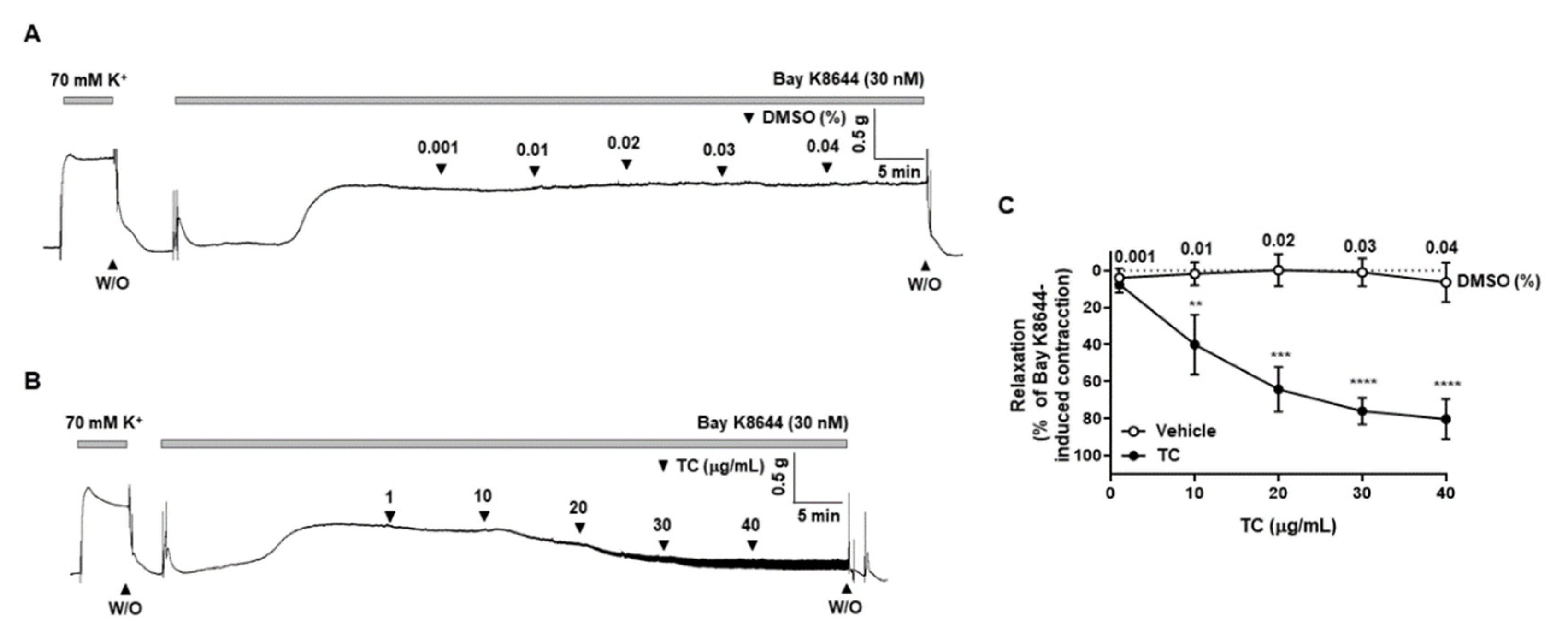
2.6. Effect of T. caulis on the BAY K8644-Induced Contraction
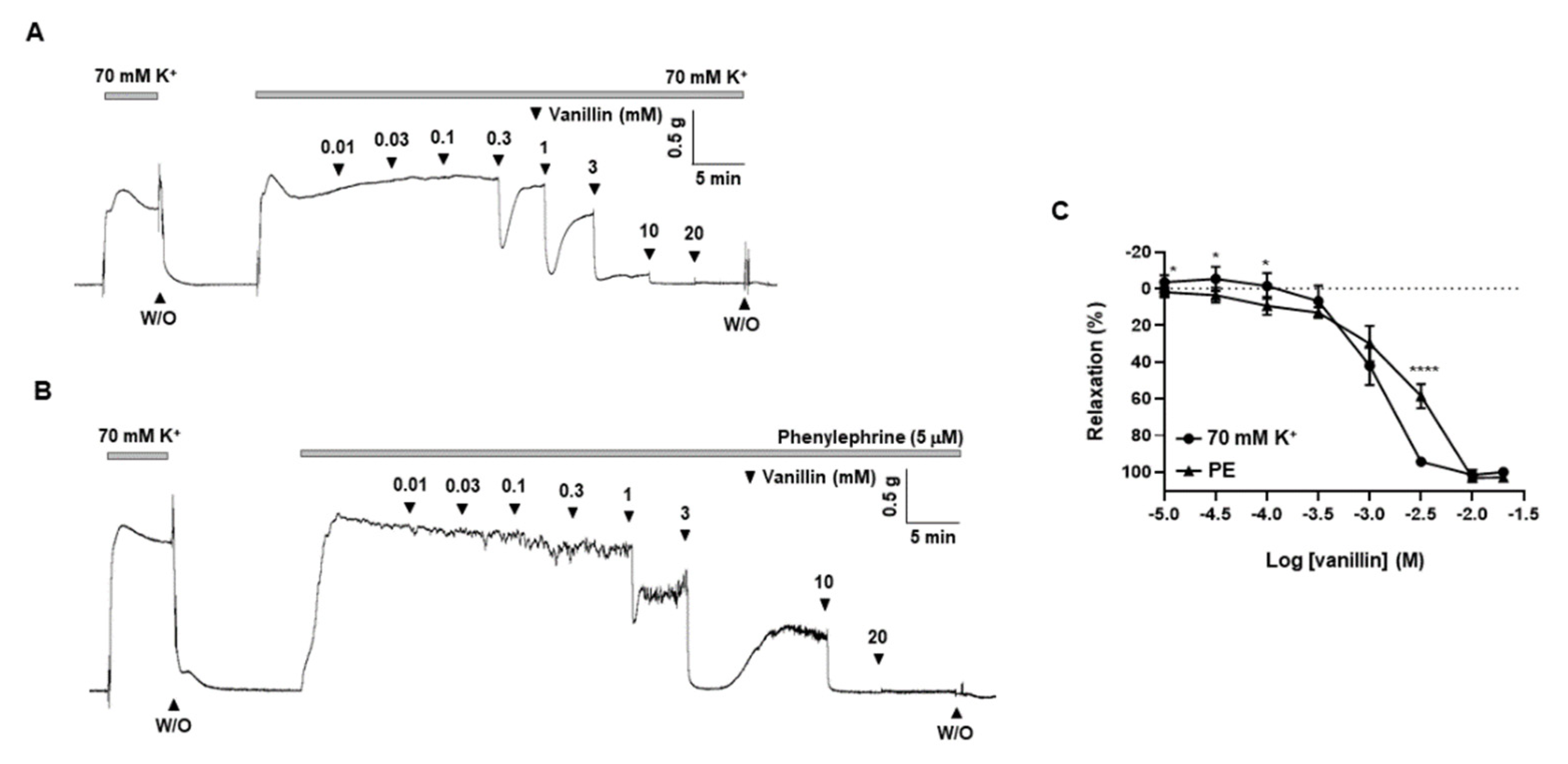
2.7. Effects of Vanillin, a Single Active Compound of T. caulis, on Mesenteric Arteries
3. Discussion
4. Materials and Methods
4.1. Animals and Tissue Preparation
4.2. Preparation of T. caulis Extract
4.3. Measurement of Isometric Tension in Mesenteric Arteries
4.4. Chemicals and Reagents
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Virani, S.S.; Alonso, A.; Benjamin, E.J.; Bittencourt, M.S.; Callaway, C.W.; Carson, A.P.; Chamberlain, A.M.; Chang, A.R.; Cheng, S.; Delling, F.N.; et al. Heart disease and stroke statistics-2020 update: A report from the american heart association. Circulation 2020, 141, e139–e596. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Xi, B.; Zhao, M.; Wang, L.; Veeranki, S.P. Uncontrolled hypertension increases risk of all-cause and cardiovascular disease mortality in us adults: The nhanes iii linked mortality study. Sci. Rep. 2018, 8, 9418. [Google Scholar] [CrossRef] [PubMed]
- Masaebi, F.; Salehi, M.; Kazemi, M.; Vahabi, N.; Looha, M.A.; Zayeri, F. Trend analysis of disability adjusted life years due to cardiovascular diseases: Results from the global burden of disease study 2019. BMC Public Health 2021, 21, 1268. [Google Scholar] [CrossRef] [PubMed]
- de Lima, R.S.; Silva, J.C.S.; Lima, C.T.; de Souza, L.E.; da Silva, M.B.; Baladi, M.G.; Irigoyen, M.C.; Lacchini, S. Proinflammatory role of angiotensin ii in the aorta of normotensive mice. Biomed. Res. Int. 2019, 2019, 9326896. [Google Scholar] [PubMed]
- Christensen, K.L.; Mulvany, M.J. Location of resistance arteries. J. Vasc. Res. 2001, 38, 1–12. [Google Scholar] [CrossRef]
- Touyz, R.M.; Alves-Lopes, R.; Rios, F.J.; Camargo, L.L.; Anagnostopoulou, A.; Arner, A.; Montezano, A.C. Vascular smooth muscle contraction in hypertension. Cardiovasc. Res. 2018, 114, 529–539. [Google Scholar] [CrossRef]
- Dunn, W.R.; Gardiner, S.M. Structural and functional properties of isolated, pressurized, mesenteric resistance arteries from a vasopressin-deficient rat model of genetic hypertension. Hypertension 1995, 26, 390–396. [Google Scholar] [CrossRef]
- Schiffrin, E.L. Reactivity of small blood-vessels in hypertension—relation with structural-changes—state-of-the-art lecture. Hypertension 1992, 19, 1–9. [Google Scholar] [CrossRef]
- Maruhashi, T.; Kihara, Y.; Higashi, Y. Assessment of endothelium-independent vasodilation: From methodology to clinical perspectives. J. Hypertens. 2018, 36, 1460–1467. [Google Scholar] [CrossRef]
- Maruhashi, T.; Soga, J.; Fujimura, N.; Idei, N.; Mikami, S.; Iwamoto, Y.; Kajikawa, M.; Matsumoto, T.; Hidaka, T.; Kihara, Y.; et al. Nitroglycerine-induced vasodilation for assessment of vascular function: A comparison with flow-mediated vasodilation. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 1401–1408. [Google Scholar] [CrossRef]
- Adams, M.R.; Robinson, J.; McCredie, R.; Seale, J.P.; Sorensen, K.E.; Deanfield, J.E.; Celermajer, D.S. Smooth muscle dysfunction occurs independently of impaired endothelium-dependent dilation in adults at risk of atherosclerosis. J. Am. Coll. Cardiol. 1998, 32, 123–127. [Google Scholar] [CrossRef]
- Handler, J. Quality of life and antihypertensive drug therapy. J. Clin. Hypertens. 2005, 7, 274–285. [Google Scholar] [CrossRef] [PubMed]
- Alaerts, G.; Merino-Arevalo, M.; Dumarey, M.; Dejaegher, B.; Noppe, N.; Matthijs, N.; Smeyers-Verbeke, J.; Vander Heyden, Y. Exploratory analysis of chromatographic fingerprints to distinguish rhizoma chuanxiong and rhizoma ligustici. J. Chromatogr. A 2010, 1217, 7706–7716. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [PubMed]
- Xia, M.; Qian, L.; Zhou, X.; Gao, Q.; Bruce, I.C.; Xia, Q. Endothelium-independent relaxation and contraction of rat aorta induced by ethyl acetate extract from leaves of morus alba (l.). J. Ethnopharmacol. 2008, 120, 442–446. [Google Scholar] [CrossRef]
- Ibarra-Alvarado, C.; Rojas, A.; Mendoza, S.; Bah, M.; Gutierrez, D.M.; Hernandez-Sandoval, L.; Martinez, M. Vasoactive and antioxidant activities of plants used in mexican traditional medicine for the treatment of cardiovascular diseases. Pharm. Biol. 2010, 48, 732–739. [Google Scholar] [CrossRef]
- Luna-Vazquez, F.J.; Ibarra-Alvarado, C.; Rojas-Molina, A.; Rojas-Molina, I.; Zavala-Sanchez, M.A. Vasodilator compounds derived from plants and their mechanisms of action. Molecules 2013, 18, 5814–5857. [Google Scholar] [CrossRef]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxidative Med. Cell. Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef]
- Shaito, A.; Thuan, D.T.B.; Phu, H.T.; Nguyen, T.H.D.; Hasan, H.; Halabi, S.; Abdelhady, S.; Nasrallah, G.K.; Eid, A.H.; Pintus, G. Herbal medicine for cardiovascular diseases: Efficacy, mechanisms, and safety. Front. Pharmacol. 2020, 11, 422. [Google Scholar] [CrossRef]
- Liu, C.; Huang, Y. Chinese herbal medicine on cardiovascular diseases and the mechanisms of action. Front. Pharmacol. 2016, 7, 469. [Google Scholar] [CrossRef]
- Shin, H.S.; Bae, M.J.; Jung, S.Y.; See, H.J.; Kim, Y.T.; Do, J.R.; Back, S.Y.; Choi, S.W.; Shon, D.H. Enhancing effect of trachelogenin from trachelospermi caulis extract on intestinal barrier function. Biol. Pharm. Bull. 2015, 38, 1707–1713. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.T.; Wang, X.G.; Yang, Y.; Xu, R.; Meng, F.H.; Yu, N.J.; Zhao, Y.M. Qualitative and quantitative analysis of lignan constituents in caulis trachelospermi by hplc-qtof-ms and hplc-uv. Molecules 2015, 20, 8107–8124. [Google Scholar] [CrossRef] [PubMed]
- Hempen, C.H.; Fischer, T. A Materia Medica for Chinese Medicine: Plants, Minerals, and Animal Products; Churchill Livingstone: London, UK, 2001; p. 374. [Google Scholar]
- Lee, M.H.; Lee, J.M.; Jun, S.H.; Ha, C.G.; Lee, S.H.; Kim, N.W.; Lee, J.H.; Ko, N.Y.; Mun, S.H.; Park, S.H.; et al. In-vitro and in-vivo anti-inflammatory action of the ethanol extract of trachelospermi caulis. J. Pharm. Pharmacol. 2007, 59, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, P.; Rengarajan, T.; Thangavel, J.; Nishigaki, Y.; Sakthisekaran, D.; Sethi, G.; Nishigaki, I. The vascular endothelium and human diseases. Int. J. Biol. Sci. 2013, 9, 1057–1069. [Google Scholar] [CrossRef]
- Rubanyi, G.M. Endothelium-derived relaxing and contracting factors. J. Cell. Biochem. 1991, 46, 27–36. [Google Scholar] [CrossRef]
- Munzel, T.; Daiber, A.; Ullrich, V.; Mulsch, A. Vascular consequences of endothelial nitric oxide synthase uncoupling for the activity and expression of the soluble guanylyl cyclase and the cgmp-dependent protein kinase. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 1551–1557. [Google Scholar] [CrossRef]
- Sellers, R.S.; Radi, Z.A.; Khan, N.K. Pathophysiology of cyclooxygenases in cardiovascular homeostasis. Vet. Pathol. 2010, 47, 601–613. [Google Scholar] [CrossRef]
- Leloup, A.J.A.; Van Hove, C.E.; De Moudt, S.; De Meyer, G.R.Y.; De Keulenaer, G.W.; Fransen, P. Vascular smooth muscle cell contraction and relaxation in the isolated aorta: A critical regulator of large artery compliance. Physiol. Rep. 2019, 7, e13934. [Google Scholar] [CrossRef]
- Gollasch, M.; Ried, C.; Bychkov, R.; Luft, F.C.; Haller, H. K+ currents in human coronary artery vascular smooth muscle cells. Circ. Res. 1996, 78, 676–688. [Google Scholar] [CrossRef]
- Kwon, Y.; Haam, C.E.; Byeon, S.; Choi, S.J.; Shin, D.H.; Choi, S.K.; Lee, Y.H. Vasodilatory Effect of Phellinus linteus Extract in Rat Mesenteric Arteries. Molecules 2020, 25, 3160. [Google Scholar] [CrossRef]
- Raffai, G.; Khang, G.; Vanhoutte, P.M. Vanillin and vanillin analogs relax porcine coronary and basilar arteries by inhibiting l-type ca2+ channels. J. Pharmacol. Exp. Ther. 2015, 352, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Hwang, I.K.; Won, M.H. Vanillin, 4-hydroxybenzyl aldehyde and 4-hydroxybenzyl alcohol prevent hippocampal ca1 cell death following global ischemia. Brain Res. 2007, 1181, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Li, C.; Liao, G.; Huang, Y.; Lv, X.; Liu, X.; Chen, W.; Zhang, L. Vanillin attenuates proinflammatory factors in a tmcao mouse model via inhibition of tlr4/nf-kb signaling pathway. Neuroscience 2022, 491, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Sultana, S.; Asif, H.M. Review: Medicinal plants combating against hypertension: A green antihypertensive approach. Pak. J. Pharm. Sci. 2017, 30, 2311–2319. [Google Scholar] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haam, C.E.; Byeon, S.; Choi, S.; Oh, E.Y.; Choi, S.-K.; Lee, Y.-H. Vasorelaxant Effect of Trachelospermi caulis Extract on Rat Mesenteric Resistance Arteries. Molecules 2022, 27, 5300. https://doi.org/10.3390/molecules27165300
Haam CE, Byeon S, Choi S, Oh EY, Choi S-K, Lee Y-H. Vasorelaxant Effect of Trachelospermi caulis Extract on Rat Mesenteric Resistance Arteries. Molecules. 2022; 27(16):5300. https://doi.org/10.3390/molecules27165300
Chicago/Turabian StyleHaam, Chae Eun, Seonhee Byeon, Sooyeon Choi, Eun Yi Oh, Soo-Kyoung Choi, and Young-Ho Lee. 2022. "Vasorelaxant Effect of Trachelospermi caulis Extract on Rat Mesenteric Resistance Arteries" Molecules 27, no. 16: 5300. https://doi.org/10.3390/molecules27165300