
Metabolic and Proteomic Analysis of Chlorella sorokiniana, Chloroidium saccharofilum, and Chlorella vulgaris Cells Cultured in Autotrophic, Photoheterotrophic, and Mixotrophic Cultivation Modes


Abstract
:1. Introduction
2. Results
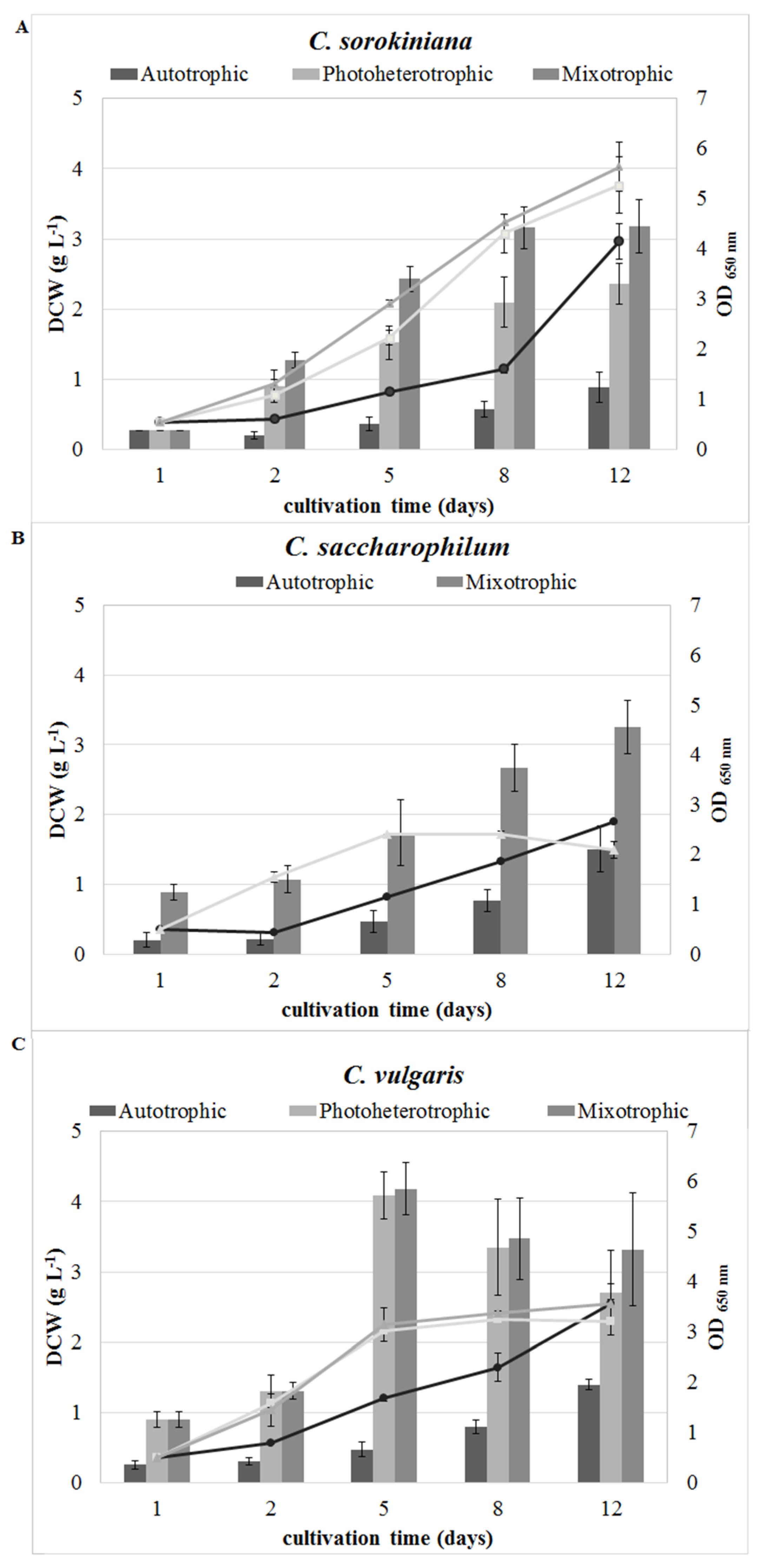
2.1. Effect of the Cultivation Mode on Growth Characteristics
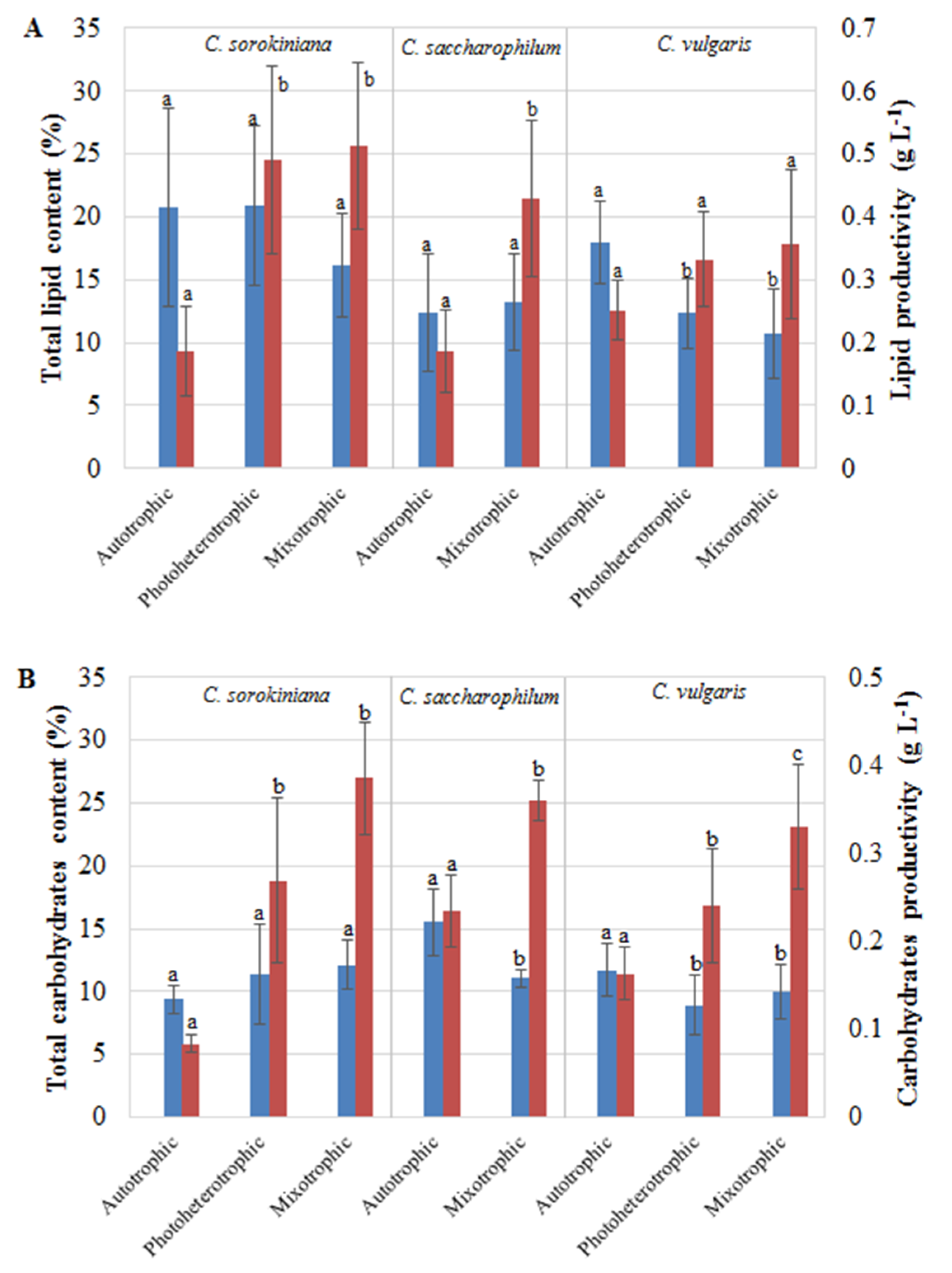
2.2. Characterization of Lipids and Carbohydrates
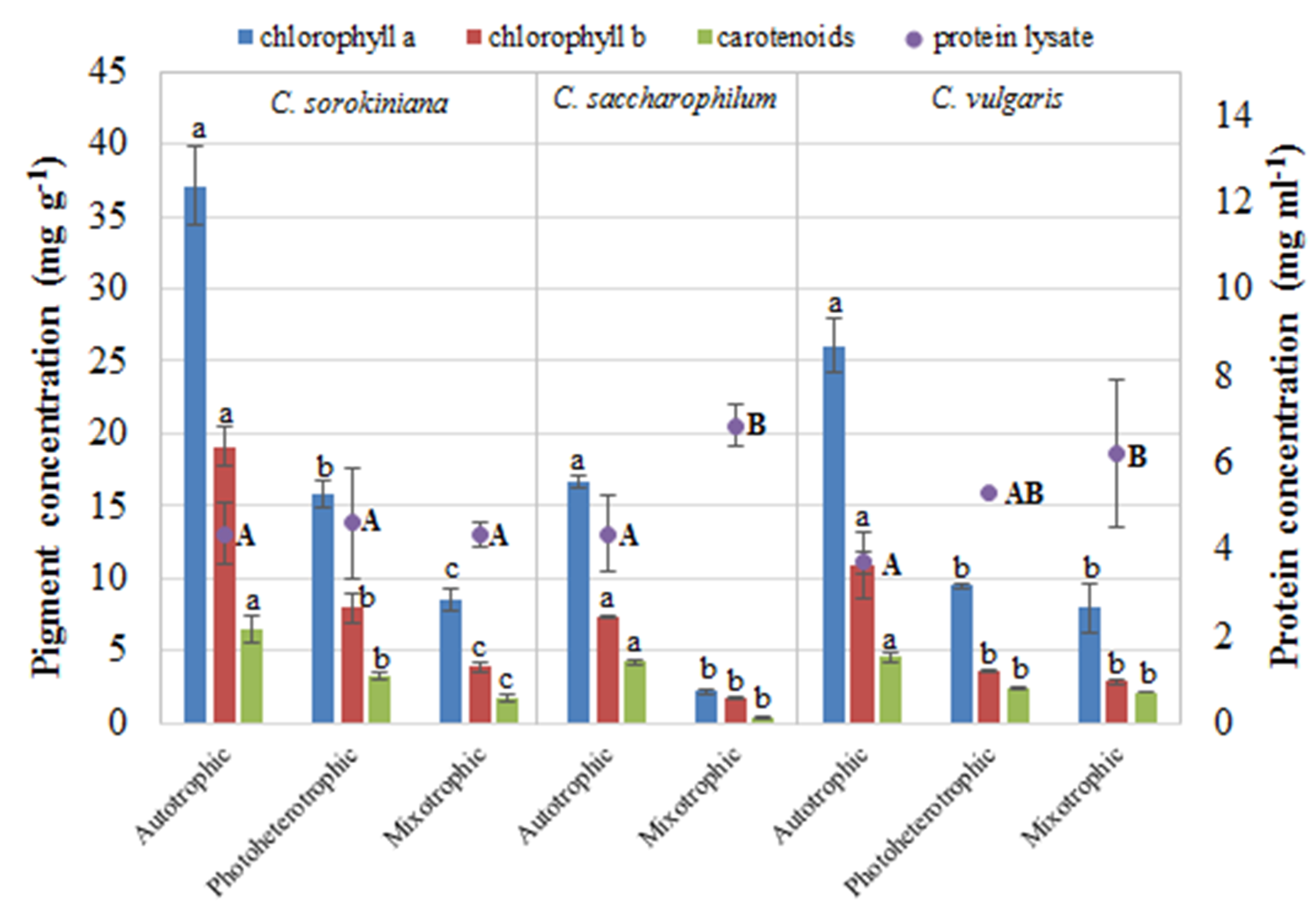
2.3. Characterization of Protein and Pigments
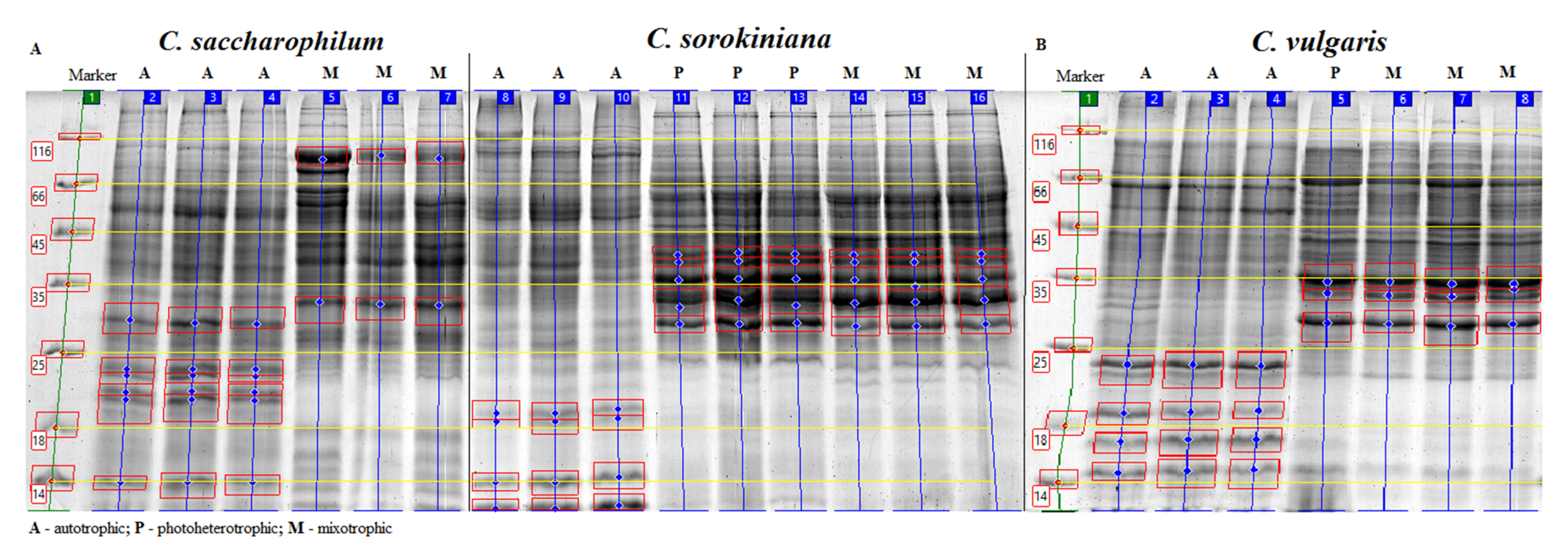
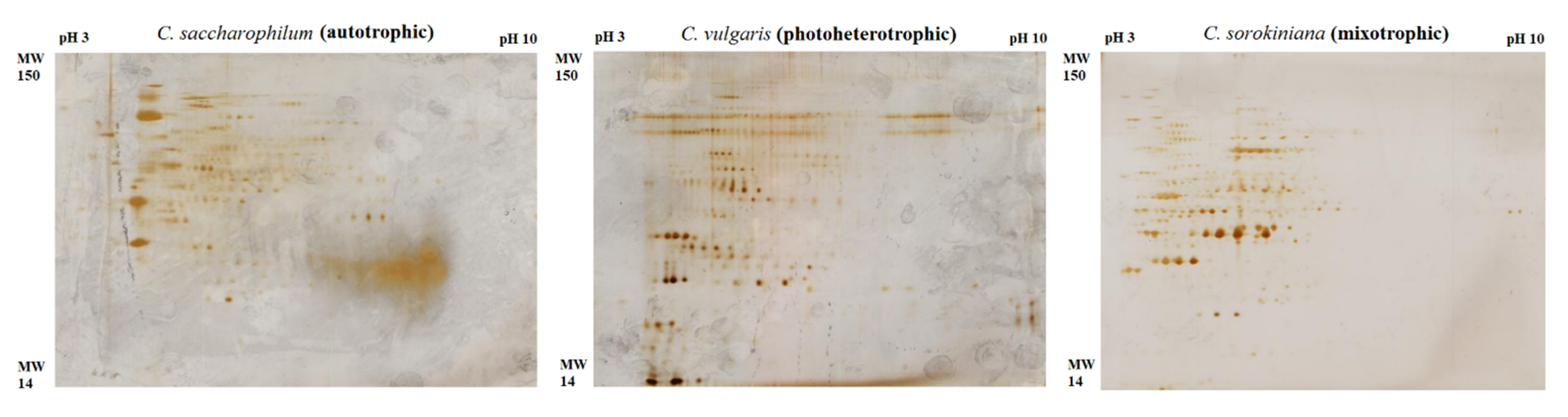
2.4. Electrophoretic Protein Profiles
2.5. Up-Regulated Protein in Photoheterotrophic and Mixotrophic Chlorella Cultures
3. Discussion
4. Materials and Methods
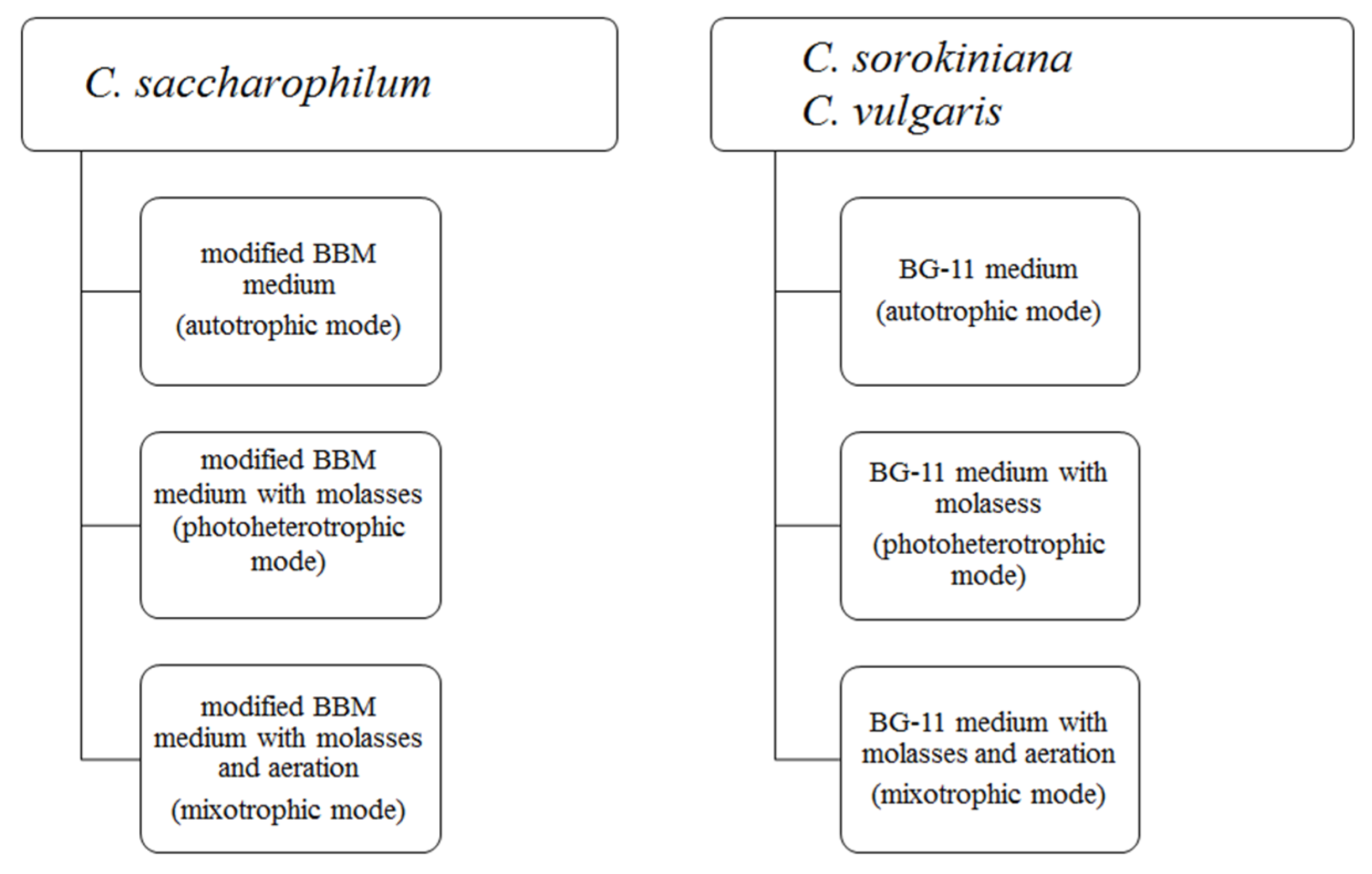
4.1. Algal Strains and Culture Conditions
4.2. Growth Characteristics
4.3. Determination of Lipids and Carbohydrates
4.4. Measurement of Photosynthetic Pigments
4.5. Protein Extraction, Determination and SDS-PAGE Electrophoresis
4.6. 2-DE Electrophoresis and MALDI/TOF Analysis
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Ismail, B.P.; Senaratne-Lenagala, L.; Stube, A.; Brackenridge, A. Protein demand: Review of plant and animal proteins used in alternative protein product development and production. Anim. Front. 2020, 10, 53–63. [Google Scholar] [CrossRef]
- Janssen, M.; Wijffels, R.H.; Barbosa, M.J. Microalgae based production of single-cell protein. Curr. Opin. Biotechnol. 2022, 75, 102705. [Google Scholar] [CrossRef]
- Montenegro-Herrera, C.A.; Vera-López Portillo, F.; Hernández-Chávez, G.T.; Martinez, A. Single-cell protein production potential with the extremophilic red microalgae Galdieria sulphuraria: Growth and biochemical characterization. J. Appl. Phycol. 2022, 34, 1341–1352. [Google Scholar] [CrossRef]
- Irvani, N.; Carna, A.; Agyei, D.; Oey, I. Algae as an Alternative Source of Protein. In Alternative Proteins: Safety and Food Security Considerations, 1st ed.; Bekhit, A.E.-D.A., Riley, W.W., Hussain, M.A., Eds.; CRC Press: Boca Raton, FL, USA, 2022. [Google Scholar]
- Piasecka, A.; Nawrocka, A.; Wiącek, D.; Krzemińska, I. Agro-industrial by-product in photoheterotrophic and mixotrophic culture of Tetradesmus obliquus: Production of ω3 and ω6 essential fatty acids with biotechnological importance. Sci. Rep. 2020, 10, 6411. [Google Scholar] [CrossRef] [PubMed]
- Tejano, L.A.; Peralta, J.P.; Yap, E.E.S.; Panjaitan, F.C.A.; Chang, Y.-W. Prediction of Bioactive Peptides from Chlorella sorokiniana Proteins Using Proteomic Techniques in Combination with Bioinformatics Analyses. Int. J. Mol. Sci. 2019, 20, 1786. [Google Scholar] [CrossRef] [Green Version]
- Waghmare, A.G.; Salve, M.K.; LeBlanc, J.G.; Arya, S.S. Concentration and characterization of microalgae proteins from Chlorella pyrenoidosa. Bioresour. Bioprocess. 2016, 3, 16. [Google Scholar] [CrossRef] [Green Version]
- Muys, M.; Sui, Y.; Schwaiger, B.; Lesueur, C.; Vandenheuvel, D.; Vermeir, P.; Vlaeminck, S.E. High variability in nutritional value and safety of commercially available Chlorella and Spirulina biomass indicates the need for smart production strategies. Bioresour. Technol. 2019, 275, 247–257. [Google Scholar] [CrossRef]
- Babich, O.; Sukhikh, S.; Larina, V.; Kalashnikova, O.; Kashirskikh, E.; Prosekov, A.; Noskova, S.; Ivanova, S.; Fendri, I.; Smaoui, S.; et al. Algae: Study of Edible and Biologically Active Fractions, Their Properties and Applications. Plants 2022, 11, 780. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Miao, X.; Wu, Q. High quality biodiesel production from a microalga Chlorella protothecoides by heterotrophic growth in fermenters. J. Biotechnol. 2006, 126, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Zhan, J.; Rong, J.; Wang, Q. Mixotrophic cultivation, a preferable microalgae cultivation mode for biomass/bioenergy production, and bioremediation, advances and prospect. Int. J. Hydrog. Energy 2017, 42, 8505–8517. [Google Scholar] [CrossRef]
- Abreu, A.P.; Morais, C.R.; Teixeira, J.A.; Nunes, J. A comparison between microalgal autotrophic growth and metabolite accumulation with heterotrophic, mixotrophic and photoheterotrophic cultivation modes. Renew. Sustain. Energy Rev. 2022, 159, 112247. [Google Scholar] [CrossRef]
- Pereira, I.; Rangel, A.; Chagas, B.; de Moura, B.; Urbano, S.; Sassi, R.; Camara, F.; Castro, C. Microalgae Growth under Mixotrophic Condition Using Agro-Industrial Waste: A Review. In Biotechnological Applications of Biomass; Basso, T.P., Basso, T.O., Basso, L.C., Eds.; IntechOpen: London, UK, 2021. [Google Scholar]
- Chakdar, H.; Hasan, M.; Pabbi, S.; Nevalainen, H.; and Shukla, P. High-throughput Proteomics and Metabolomic Studies Guide Re-engineering of Metabolic Pathways in Eukaryotic Microalgae: A Review. Bioresour. Technol. 2021, 321, 124495. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xu, H.; Han, F.; Mu, J.; Chen, D.; Feng, B.; Zeng, H. Regulation of lipid metabolism in the green microalga Chlorella protothecoides by heterotrophy-photoinduction cultivation regime. Bioresour. Technol. 2015, 192, 781–791. [Google Scholar] [CrossRef]
- Pego, J.V.; Kortstee, A.J.; Huijser, C.; Smeekens, S.C.M. Photosynthesis, sugars and the regulation of gene expression. J. Exp. Bot. 2000, 51, 407–416. [Google Scholar] [CrossRef] [Green Version]
- Arora, N.; Philippidis, G.P. Insights into the physiology of Chlorella vulgaris cultivated in sweet sorghum bagasse hydrolysate for sustainable algal biomass and lipid production. Sci. Rep. 2021, 11, 6779. [Google Scholar] [CrossRef]
- León-Vaz, A.; León, R.; Díaz-Santos, E.; Vigara, J.; Raposo, S. Using agro-industrial wastes for mixotrophic growth and lipids production by the green microalga Chlorella sorokiniana. N. Biotechnol. 2019, 51, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Piasecka, A.; Krzemińska, I.; Tys, J. Enrichment of Parachlorella kessleri biomass with bioproducts: Oil and protein by utilization of beet molasses, J. Appl. Phycol. 2017, 29, 1735–1743. [Google Scholar] [CrossRef] [Green Version]
- López, G.; Yate, C.; Ramos, F.; Cala, M.P.; Restrepo, S.; Baena, S. Production of Polyunsaturated Fatty Acids and Lipids from Autotrophic, Mixotrophic and Heterotrophic cultivation of Galdieria sp. strain USBA-GBX-832. Sci. Rep. 2019, 9, 10791. [Google Scholar] [CrossRef] [PubMed]
- Behera, B.; Unpaprom, T.; Ramaraj, R.; Maniam, G.P.; Govindan, N.; Paramasivan, B. Integrated biomolecular and bioprocess engineering strategies for enhancing the lipid yield from microalgae. Renew. Sustain. Energy Rev. 2021, 148, 111270. [Google Scholar] [CrossRef]
- Shi, T.Q.; Wang, L.R.; Zhang, Z.X.; Sun, X.M.; Huang, H. Stresses as first-line tools for enhancing lipid and carotenoid production in microalgae. Front. Bioeng. Biotechnol. 2020, 8, 610. [Google Scholar] [CrossRef]
- Abreu, A.P.; Fernandes, B.; Vicente, A.A.; Teixeira, J.; Dragone, G. Mixotrophic cultivation of Chlorella vulgaris using industrial dairy waste as organic carbon source. Bioresour. Technol. 2012, 118, 61–66. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Liu, S.F.; Qin, Z.H.; Balamurugan, S.; Li, H.Y.; Lin, C.S.K. Sustainable and stepwise waste-based utilisation strategy for the production of biomass and biofuels by engineered microalgae. Environ. Pollut. 2020, 265, 114854. [Google Scholar] [CrossRef] [PubMed]
- Lau, K.Y.; Pleissner, D.; Lin, C.S.K. Recycling of food waste as nutrients in Chlorella vulgaris cultivation. Bioresour. Technol. 2014, 170, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Yew, G.Y.; Puah, B.K.; Chew, K.W.; Teng, S.Y.; Show, P.L.; Nguyen, T.H.P. Chlorella vulgaris FSP-E cultivation in waste molasses: Photo-to-property estimation by artificial intelligence. Chem. Eng. J. 2020, 402, 126230. [Google Scholar] [CrossRef]
- Gao, B.; Wang, F.; Huang, L.; Liu, H.; Zhong, Y.; Zhang, C. Biomass, lipid accumulation kinetics, and the transcriptome of heterotrophic oleaginous microalga Tetradesmus bernardii under different carbon and nitrogen sources. Biotechnol. Biofuels 2021, 14, 4. [Google Scholar] [CrossRef]
- de Melo, R.G.; de Andrade, A.F.; Bezerra, R.P.; Correia, D.S.; de Souza, V.C.; Brasileiro-Vidal, A.C.; Marques, D.A.V.; Porto, A.L.F. Chlorella vulgaris mixotrophic growth enhanced biomass productivity and reduced toxicity from agro-industrial by-products. Chemosphere 2018, 204, 344–350. [Google Scholar] [CrossRef]
- Adams, C.; Godfrey, V.; Wahlen, B.; Seefeldt, L.; Bugbee, B. Understanding precision nitrogen stress to optimize the growth and lipid content trade off in oleaginous green microalgae. Bioresour. Technol. 2013, 131, 188–194. [Google Scholar] [CrossRef] [Green Version]
- Markou, G.; Diamantis, A.; Korozi, E.; Tsagou, V.; Kefalogianni, I.; Chatzipavlidis, I. Effects of Monochromatic Illumination with LEDs Lights on the Growth and Photosynthetic Performance of Auxenochlorella protothecoides in Photo- and Mixotrophic Conditions. Plants 2021, 10, 799. [Google Scholar] [CrossRef] [PubMed]
- Cecchin, M.; Benfatto, S.; Griggio, F.; Mori, A.; Cazzaniga, S.; Vitulo, N.; Delledonne, M.; Ballottari, M. Molecular basis of autotrophic vs mixotrophic growth in Chlorella sorokiniana. Sci. Rep. 2018, 8, 6465. [Google Scholar] [CrossRef]
- Afify, A.E.M.R.; El Baroty, G.S.; El Baz, F.K.; Abd El Baky, H.H.; Murad, S.A. Scenedesmus obliquus: Antioxidant and antiviral activity of proteins hydrolyzed by three enzymes. J. Genet. Eng. Biotechnol. 2018, 16, 399–408. [Google Scholar] [CrossRef]
- Atkinson, N.; Leitão, N.; Orr, D.J.; Meyer, M.T.; Carmo-Silva, E.; Griffiths, H.; Smith, A.M.; McCormick, A.J. Rubisco small subunits from the unicellular green alga Chlamydomonas complement Rubisco-deficient mutants of Arabidopsis. New Phytol. 2017, 214, 655–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henard, C.A.; Guarnieri, M.T.; Knoshaug, E.P. The Chlorella vulgaris S-Nitrosoproteome under Nitrogen-Replete and -Deplete Conditions. Front. Bioeng. Biotechnol. 2017, 4, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Mu, J.; Chen, D.; Xu, H.; Han, F.; Feng, B.; Zeng, H. Proteomics analysis for enhanced lipid accumulation in oleaginous Chlorella vulgaris under a heterotrophic-Na+ induction two-step regime. Biotechnol. Lett. 2015, 37, 1021–1030. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Singh, G.P.; Sharma, V.K. Effects of culture conditions on growth and biochemical profile of Chlorella vulgaris. J. Plant Pathol. Microbiol. 2012, 3, 1000131. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.; Zhang, H.; Zhao, J.; Xu, Y.; Ge, F. Mechanisms of cetyltrimethyl ammonium chloride-induced toxicity to photosystem II oxygen evolution complex of Chlorella vulgaris F1068. J. Hazard. Mater. 2020, 383, 121063. [Google Scholar] [CrossRef]
- Li, T.; Yang, F.; Xu, J.; Wu, H.; Mo, J.; Dai, L.; Xiang, W. Evaluating differences in growth, photosynthetic efficiency, and transcriptome of asterarcys sp. SCS-1881 under autotrophic, mixotrophic, and heterotrophic culturing conditions. Algal Res. 2020, 45, 101753. [Google Scholar] [CrossRef]
- Morales-Sánchez, D.; Tinoco-Valencia, R.; Kyndt, J.; Martinez, A. Heterotrophic growth of Neochloris oleoabundans using glucose as a carbon source. Biotechnol. Biofuels 2013, 6, 100. [Google Scholar] [CrossRef] [PubMed]
- León-Vaz, A.; Romero, L.C.; Gotor, C.; León, R.; Vigara, J. Effect of cadmium in the microalga Chlorella sorokiniana: A proteomic study. Ecotoxicol. Environ. Saf. 2021, 207, 111301. [Google Scholar] [CrossRef]
- Vidotti, A.D.; Riaño-Pachón, D.M.; Mattiello, L.; Giraldi, L.A.; Winck, F.V.; Franco, T.T. Analysis of autotrophic, mixotrophic and heterotrophic phenotypes in the microalgae Chlorella vulgaris using time-resolved proteomics and transcriptomics approaches. Algal Res. 2020, 51, 102060. [Google Scholar] [CrossRef]
- Krzemińska, I.; Oleszek, M.; Wiącek, D. Liquid Anaerobic Digestate as a Source of Nutrients for Lipid and Fatty Acid Accumulation by Auxenochlorella Protothecoides. Molecules 2019, 24, 3582. [Google Scholar] [CrossRef] [Green Version]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Phys. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Piasecka, A.; Krzemińska, I.; Tys, J. Physical methods of microalgal biomass pretreatment. Int. Agrophys. 2014, 28, 341–348. [Google Scholar] [CrossRef] [Green Version]
- Trevelyan, W.; Forrest, R.; Harrison, J. Determination of Yeast Carbohydrates with the Anthrone Reagent. Nature 1952, 170, 626–627. [Google Scholar] [CrossRef] [PubMed]
- Kalinowska, R.; Pawlik-Skowrońska, B. Response of two terrestrial green microalgae (Chlorophyta, Trebouxiophyceae) isolated from Cu-rich and unpolluted soils to copper stress. Environ. Pollut. 2010, 158, 2778–2785. [Google Scholar] [CrossRef] [PubMed]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Cid, C.; Garcia-Descalzo, L.; Casado-Lafuente, V.; Amils, R.; Aguilera, A. Proteomic analysis of the response of an acidophilic strain of Chlamydomonas sp. (Chlorophyta) to natural metal-rich water. Proteomics 2010, 10, 2026–2036. [Google Scholar] [CrossRef]
- Jia, Y.L.; Chen, H.; Zhang, C.; Gao, L.J.; Wang, X.C.; Qiu, L.L.; Wu, J.F. Proteomic analysis of halotolerant proteins under high and low salt stress in Dunaliella salina using two-dimensional differential in-gel electrophoresis. Genet. Mol. Biol. 2016, 39, 239–247. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | C. sorokiniana | C. saccharophilum | C. vulgaris | |||||
|---|---|---|---|---|---|---|---|---|
| Conditions | Autotrophic | Photoheterotrophic | Mixotrophic | Autotrophic | Mixotrophic | Autotrophic | Photoheterotrophic | Mixotrophic |
| Specific growth rate µ (d−1) | 0.19 ± 0.01 a | 0.37 ± 0.02 b | 0.44 ± 0.01 c | 0.21 ± 0.01 a | 0.39 ± 0.00 b | 0.30 ± 0.01 a | 0.45 ± 0.00 b | 0.46 ± 0.03 b |
| Doubling time (h) | 90.01 ± 5.76 a | 44.63 ± 1.81 b | 37.81 ± 0.57 c | 81.10 ± 2.67 a | 42.45 ± 0.13 b | 54.98 ± 1.37 a | 36.92 ± 0.21 b | 36.47 ± 2.94 b |
| Biomass productivity (mg L−1 d−1) | 74.38 ± 18.05 a | 196.53 ± 24.07 b | 264.58 ± 31.71 c | 125.56 ± 27.07 a | 271.04 ± 31.72 b | 116.67 ± 6.80 a | 225.00 ± 50.00 b | 276.39 ± 67.27 b |
| No. | Up-Regulated Protein | Species/Culture Conditions | Theoretical MW/MW (kDa) | pI | Fold Change | Molecular Function/Localization |
|---|---|---|---|---|---|---|
| 1 | ATP synthase subunit beta | C. sorokiniana/mixotrophic | 42.1/39.6 | 5.3 | 3.2 | Energy metabolism/chloroplast |
| 2 | Chloroplast light-harvesting complex II | C. sorokiniana/mixotrophic | 24.0/26.1 | 5.0 | 2.9 | Protein synthesis/chloroplast |
| 3 | Luminal binding protein | C. saccharophilum/mixotrophic | 71.8/73 | 4.8 | 3.6 | Stress response/ER |
| 4 | Glucose-6-phosphate 1 DH precursor | C. saccharophilum/mixotrophic | 66.9/67 | 8.5 | 4.1 | Carbohydrate metabolism/chloroplast |
| 5 | Hsp70 | C. vulgaris/photoheterotrophic | 70.9/71 | 5.3 | 2.9 | Stress response/chloroplast |
| 6 | Hsp90 | C. vulgaris/photoheterotrophic | 80.7/80 | 4.9 | 2.7 | Stress response/chloroplast |
| 7 | Dynein | C. vulgaris/photoheterotrophic | 76/77 | 5.4 | 3.1 | Cytoskeleton protein |
| 8 | α-tubulin | C. vulgaris/photoheterotrophic | 49/50 | 5.0 | 2.2 | Cytoskeleton protein |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piasecka, A.; Baier, A. Metabolic and Proteomic Analysis of Chlorella sorokiniana, Chloroidium saccharofilum, and Chlorella vulgaris Cells Cultured in Autotrophic, Photoheterotrophic, and Mixotrophic Cultivation Modes. Molecules 2022, 27, 4817. https://doi.org/10.3390/molecules27154817
Piasecka A, Baier A. Metabolic and Proteomic Analysis of Chlorella sorokiniana, Chloroidium saccharofilum, and Chlorella vulgaris Cells Cultured in Autotrophic, Photoheterotrophic, and Mixotrophic Cultivation Modes. Molecules. 2022; 27(15):4817. https://doi.org/10.3390/molecules27154817
Chicago/Turabian StylePiasecka, Agata, and Andrea Baier. 2022. "Metabolic and Proteomic Analysis of Chlorella sorokiniana, Chloroidium saccharofilum, and Chlorella vulgaris Cells Cultured in Autotrophic, Photoheterotrophic, and Mixotrophic Cultivation Modes" Molecules 27, no. 15: 4817. https://doi.org/10.3390/molecules27154817