
Cloning, Expression, Purification, and Characterization of β-Galactosidase from Bifidobacterium longum and Bifidobacterium pseudocatenulatum

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
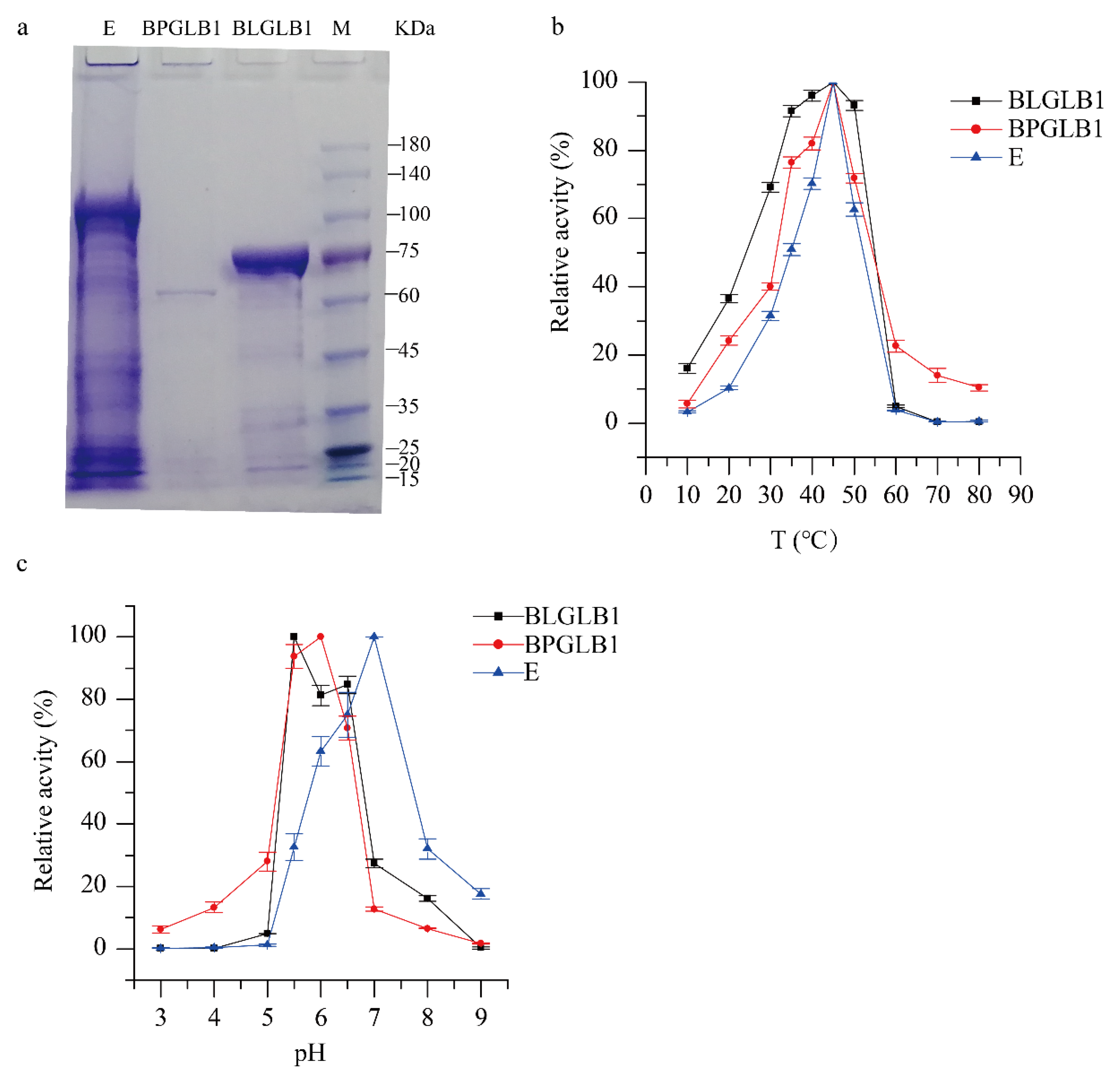
2.1. Characterization of BLGLB1 and BPGLB1
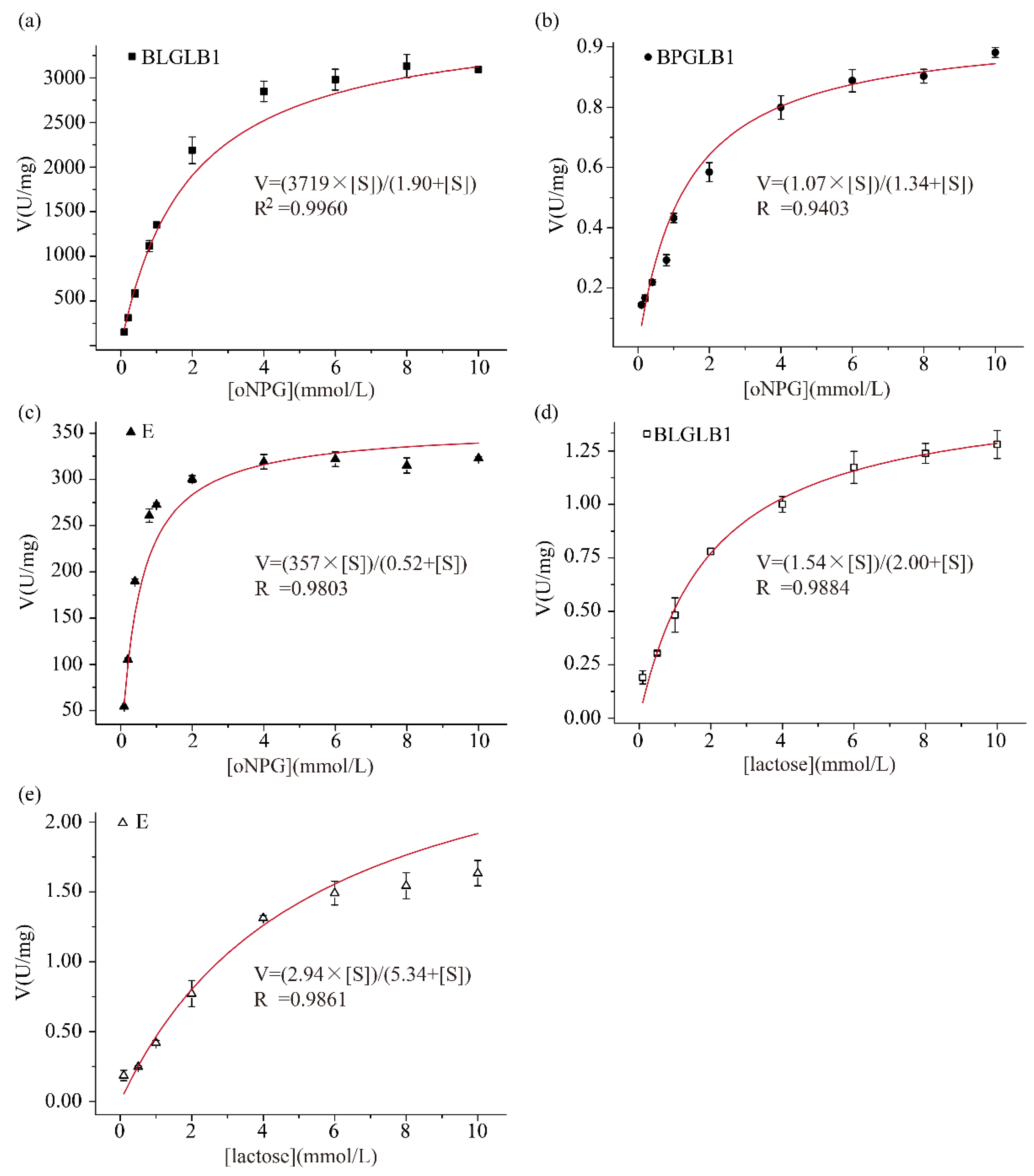
2.2. Determination of the Kinetic Parameters of BLGLB1 and BPGLB1

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Resource | Km (mM) | kcat | kcat/Km | Vmax (U∙mg−1) | Optimal Conditions | Reference |
|---|---|---|---|---|---|---|
| B. longum | 0.85 (oNPG) | - | - | 70.67 (oNPG) | pH = 7.0/50 °C | [25] |
| B. bifidum | 7.0 (oNPG) 24.0 (lactose) | 6900 (oNPG) 0.7 (lactose) | 985.71 (oNPG) 0.03 (lactose) | - | pH = 6.0/40 °C | [31] |
| B. adolescentis | 2.5 (oNPG) 3.7 (lactose) | - | - | 107 (oNPG) 22 (lactose) | pH = 7.0/37 °C | [32] |
| B. longum subsp. infantis | 16 ± 2 (lactose) | 97 ± 3 (lactose) | 6.1 ± 0.5 (lactose) | - | - | [33] |
| B. longum subsp. infantis | 44 ± 10 (lactose) | 8.6 ± 0.1 (lactose) | 0.19 ± 0.00 (lactose) | - | - | [33] |
| B. longum subsp. infantis | - | - | 0.08 ± 0.00 (lactose) | - | - | [33] |
| B. animalis subsp. lactis | 25.0 (lactose) | - | - | - | pH = 6.5/37 °C | [34] |
| Lactobacillus acidophilus | 3.84 (oNPG) 88.98 (lactose) | - | - | - | pH = 6.0/37 °C | [35] |
| infant feces | 20.95 ± 2.76 (oNPG) 140.2 ± 17.7 (lactose) | - | - | 5004.50± 318.8 (oNPG) 293.1 ± 14.7 (lactose) | pH = 6.5/50 °C | [36] |
| B. adolescentis | 60 (pNPG) | - | - | 1.129 (lactose) | pH = 6.0/50 °C | [37] |
| B. longum | 1.9 ± 0.1 (oNPG) | 1700 ± 40 (oNPG) | 870 (oNPG) | 3700 ± 100 (oNPG) | pH = 5.5/45 °C | This study (BLGLB1) |
| B. pseudocatenulatum | 1.3 ± 0.3 (oNPG) | 0.5 ± 0.02 (oNPG) | 0.36 (oNPG) | 1.1 ± 0.1 (oNPG) | pH = 6.0/45 °C | This study (BPGLB1) |
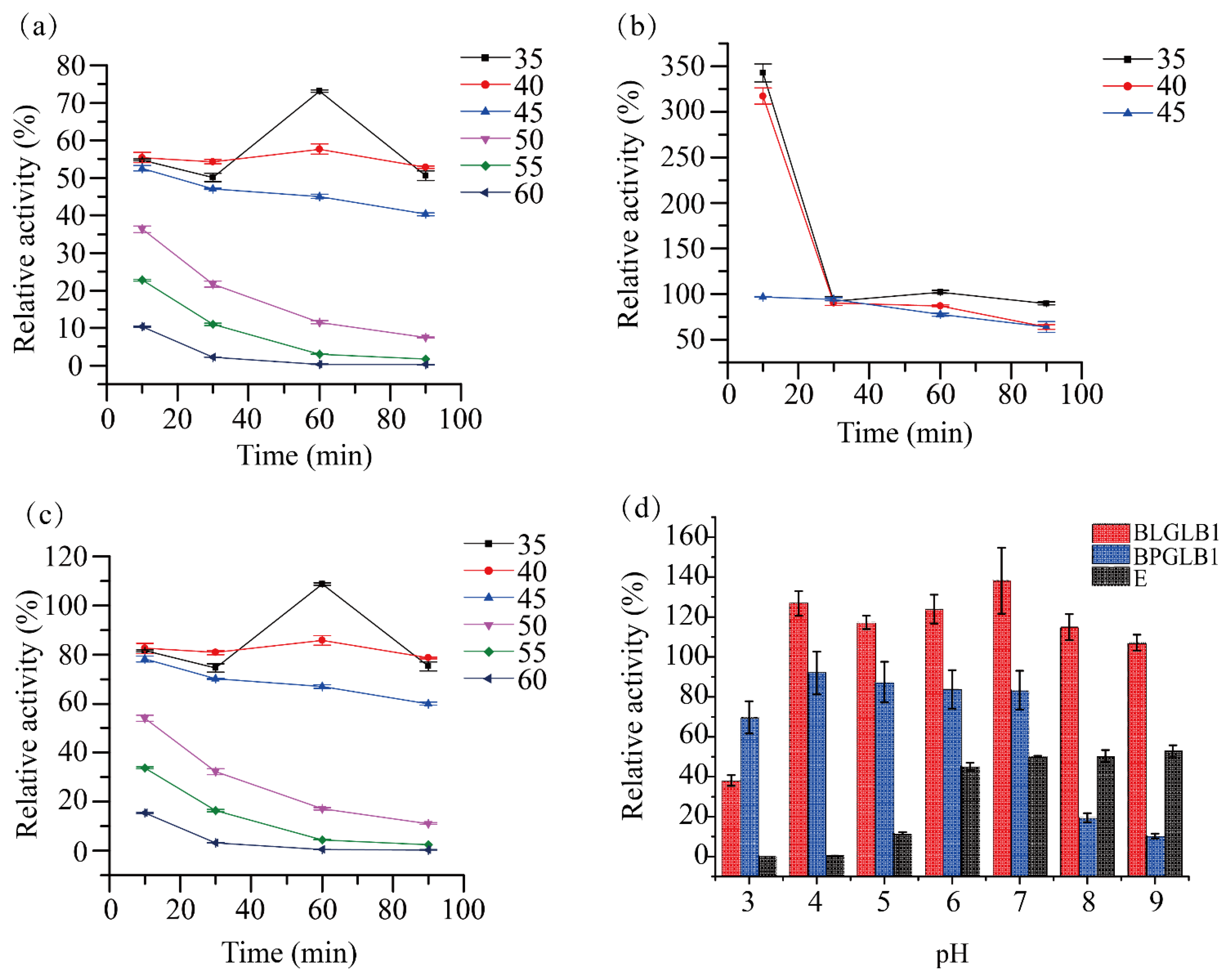
2.3. Stability of BLGLB1 and BPGLB1
2.4. Evaluation of the Hydrolytic Activity of BLGLB1 and BPGLB1
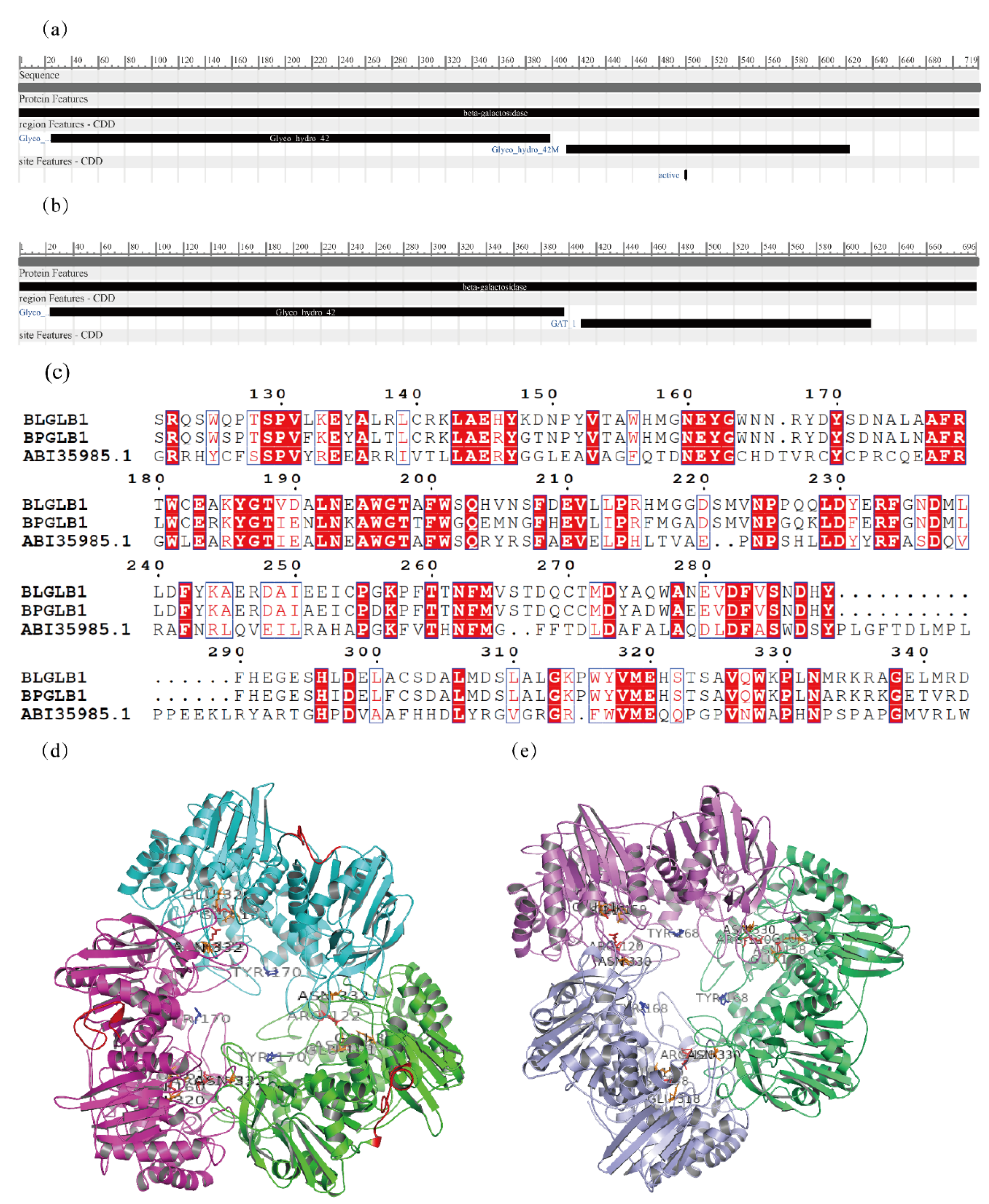
2.5. Bioinformatic Analysis
3. Materials and Methods
3.1. Materials
3.2. Bacterial Strains
3.3. Whole Genome Sequencing and β-Galactosidase Gene Mining
3.4. Cloning and Expression of BLGLB1 and BPGLB1
3.5. Purification of BLGLB1 and BPGLB1
3.6. Quantification of β-Galactosidase Activity
3.7. Effect of Temperature and pH on BLGLB1 and BPGLB1 Activity
3.8. Effect of Metallic Cations on BLGLB1 and BPGLB1 Activity
3.9. Determination of the Kinetic Parameters of BLGLB1 and BPGLB1
3.10. Stability
3.11. Evaluation of the Hydrolytic Activity of BLGLB1 and BPGLB1
3.12. Bioinformatic Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Kamran, A.; Bibi, Z.; Aman, A.; Qader, S.A.U. Lactose hydrolysis approach: Isolation and production of β-galactosidase from newly isolated Bacillus strain B-2. Biocatal. Agric. Biotechnol. 2016, 5, 99–103. [Google Scholar] [CrossRef]
- Nguyen, T.-T.; Nguyen, H.-M.; Geiger, B.; Mathiesen, G.; Eijsink, V.; Peterbauer, C.K.; Haltrich, D.; Nguyen, T.-H. Heterologous expression of a recombinant lactobacillal β-galactosidase in Lactobacillus plantarum: Effect of different parameters on the sakacin P-based expression system. Microb. Cell Factories 2015, 14, 30. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, C.; Guimarães, P.M.R.; Domingues, L. Recombinant microbial systems for improved β-galactosidase production and biotechnological applications. Biotechnol. Adv. 2011, 29, 600–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, J.; Romantsov, T.; Engel, K.; Doxey, A.C.; Rose, D.R.; Neufeld, J.D.; Charles, T.C. Functional metagenomics reveals novel β-galactosidases not predictable from gene sequences. PLoS ONE 2017, 12, e0172545. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.Z.K.; Chen, X.D. Effects of temperature and pH on the catalytic activity of the immobilized β-galactosidase from Kluyveromyces lactis. Biochem. Eng. J. 2001, 9, 33–40. [Google Scholar] [CrossRef]
- Aburto, C.; Castillo, C.; Cornejo, F.; Arenas-Salinas, M.; Vásquez, C.; Guerrero, C.; Arenas, F.; Illanes, A.; Vera, C. β-Galactosidase from Exiguobacterium acetylicum: Cloning, expression, purification and characterization. Bioresour. Technol. 2019, 277, 211–215. [Google Scholar] [CrossRef]
- Simović, M.; Milivojević, A.; Ćorović, M.; Banjanac, K.; Bezbradica, D. Whey valorization using transgalactosylation activity of immobilized β-galactosidase. Int. J. Food Sci. Technol. 2019, 54, 3074–3082. [Google Scholar] [CrossRef]
- Kokkiligadda, A.; Beniwal, A.; Saini, P.; Vij, S. Utilization of Cheese Whey Using Synergistic Immobilization of beta-Galactosidase and Saccharomyces cerevisiae Cells in Dual Matrices. Appl. Biochem. Biotechnol. 2016, 179, 1469–1484. [Google Scholar] [CrossRef]
- Hegar, B.; Widodo, A. Lactose intolerance in Indonesian children. Asia Pac. J. Clin. Nutr. 2015, 24 (Suppl. 1), S31–S40. [Google Scholar] [CrossRef]
- Lukito, W.; Malik, S.G.; Surono, I.S.; Wahlqvist, M.L. From ‘lactose intolerance’ to ‘lactose nutrition’. Asia Pac. J. Clin. Nutr. 2015, 24 (Suppl. 1), S1–S8. [Google Scholar] [CrossRef]
- Mattar, R.; de Campos Mazo, D.F.; Carrilho, F.J. Lactose intolerance: Diagnosis, genetic, and clinical factors. Clin. Exp. Gastroenterol. 2012, 5, 113–121. [Google Scholar] [CrossRef] [Green Version]
- Yao, C.; Sun, J.; Wang, W.; Zhuang, Z.; Liu, J.; Hao, J. A novel cold-adapted β-galactosidase from Alteromonas sp. ML117 cleaves milk lactose effectively at low temperature. Process Biochem. 2019, 82, 94–101. [Google Scholar] [CrossRef]
- Aburto, C.; Guerrero, C.; Vera, C.; Illanes, A. Improvement in the yield and selectivity of lactulose synthesis with Bacillus circulans β-galactosidase. LWT 2020, 118, 108746. [Google Scholar] [CrossRef]
- Saqib, S.; Akram, A.; Halim, S.A.; Tassaduq, R. Sources of β-galactosidase and its applications in food industry. 3 Biotech 2017, 7, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chanalia, P.; Gandhi, D.; Attri, P.; Dhanda, S. Purification and characterization of β-galactosidase from probiotic Pediococcus acidilactici and its use in milk lactose hydrolysis and galactooligosaccharide synthesis. Bioorg. Chem. 2018, 77, 176–189. [Google Scholar] [CrossRef]
- Lima, P.C.; Gazoni, I.; de Carvalho, A.M.G.; Bresolin, D.; Cavalheiro, D.; de Oliveira, D.; Rigo, E. β-galactosidase from Kluyveromyces lactis in genipin-activated chitosan: An investigation on immobilization, stability, and application in diluted UHT milk. Food Chem. 2021, 349, 129050. [Google Scholar] [CrossRef]
- Zerva, A.; Limnaios, A.; Kritikou, A.S.; Thomaidis, N.S.; Taoukis, P.; Topakas, E. A novel thermophile β-galactosidase from Thermothielavioides terrestris producing galactooligosaccharides from acid whey. New Biotechnol. 2021, 63, 45–53. [Google Scholar] [CrossRef]
- Nguyen, T.-H.; Splechtna, B.; Krasteva, S.; Kneifel, W.; Kulbe, K.D.; Divne, C.; Haltrich, D. Characterization and molecular cloning of a heterodimeric β-galactosidase from the probiotic strain Lactobacillus acidophilus R22. FEMS Microbiol. Lett. 2007, 269, 136–144. [Google Scholar] [CrossRef] [Green Version]
- Hung, M.N.; Lee, B.H. Purification and characterization of a recombinant beta-galactosidase with transgalactosylation activity from Bifidobacterium infantis HL96. Appl. Microbiol. Biotechnol. 2002, 58, 439–445. [Google Scholar] [CrossRef]
- Ji, D.; Oey, I.; Agyei, D. Purification, characterization and thermal inactivation kinetics of β-galactosidase from Lactobacillus leichmannii 313. LWT 2019, 116, 108545. [Google Scholar] [CrossRef]
- Park, M.J.; Park, M.S.; Ji, G.E. Cloning and Heterologous Expression of the β-Galactosidase Gene from Bifidobacterium longum RD47 in B. bifidum BGN4. J. Microbiol. Biotechnol. 2019, 29, 1717–1728. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Li, W.; Rui, X.; Sun, X.; Dong, M. Lactobacillus plantarum 70810 from Chinese paocai as a potential source of β-galactosidase for prebiotic galactooligosaccharides synthesis. Eur. Food Res. Technol. 2013, 236, 817–826. [Google Scholar] [CrossRef]
- Kim, J.W.; Rajagopal, S.N. Isolation and characterization of β-galactosidase fromLactobacillus crispatus. Folia Microbiol. 2000, 45, 29–34. [Google Scholar] [CrossRef]
- Ibrahim, S.A.; Alazzeh, A.Y.; Awaisheh, S.S.; Song, D.; Shahbazi, A.; AbuGhazaleh, A.A. Enhancement of α- and β-Galactosidase Activity in Lactobacillus reuteri by Different Metal Ions. Biol. Trace Elem. Res. 2010, 136, 106–116. [Google Scholar] [CrossRef]
- Hsu, C.-A.; Yu, R.-C.; Chou, C.-C. Purification and characterization of a sodium-stimulated β-galactosidasefrom Bifidobacterium longum CCRC 15708. World J. Microbiol. Biotechnol. 2006, 22, 355–361. [Google Scholar] [CrossRef]
- Budriene, S.; Gorochovceva, N.; Romaskevic, T.; Yugova, L.; Miezeliene, A.; Dienys, G.; Zubriene, A. β-Galactosidase from Penicillium canescens. Properties and immobilization. Open Chem. 2005, 3, 95–105. [Google Scholar] [CrossRef]
- Vidya, B.; Palaniswamy, M.; Angayarkanni, J.; Nawaz, K.A.; Thandeeswaran, M.; Chaithanya, K.K.; Tekluu, B.; Muthusamy, K.; Gopalakrishnan, V.K. Purification and characterization of β-galactosidase from newly isolated Aspergillus terreus (KUBCF1306) and evaluating its efficacy on breast cancer cell line (MCF-7). Bioorg. Chem. 2020, 94, 103442. [Google Scholar] [CrossRef]
- Liao, F.; Tian, K.C.; Yang, X.; Zhou, Q.X.; Zeng, Z.C.; Zuo, Y.P. Kinetic substrate quantification by fitting the enzyme reaction curve to the integrated Michaelis-Menten equation. Anal. Bioanal. Chem. 2003, 375, 756–762. [Google Scholar] [CrossRef]
- Hildebrandt, P.; Wanarska, M.; Kur, J. A new cold-adapted beta-D-galactosidase from the Antarctic Arthrobacter sp. 32c—gene cloning, overexpression, purification and properties. BMC Microbiol. 2009, 9, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turroni, F.; Bottacini, F.; Foroni, E.; Mulder, I.; Kim, J.H.; Zomer, A.; Sánchez, B.; Bidossi, A.; Ferrarini, A.; Giubellini, V.; et al. Genome analysis of Bifidobacterium bifidum PRL2010 reveals metabolic pathways for host-derived glycan foraging. Proc. Natl. Acad. Sci. USA 2010, 107, 19514–19519. [Google Scholar] [CrossRef] [Green Version]
- Godoy, A.S.; Camilo, C.M.; Kadowaki, M.A.; Muniz, H.D.; Santo, M.E.; Murakami, M.T.; Nascimento, A.S.; Polikarpov, I. Crystal structure of β-1,6-galactosidase from Bifidobacterium bifidum S17: Trimeric architecture, molecular determinants of the enzymatic activity and its inhibition by α-galactose. FEBS J. 2016, 283, 4097–4112. [Google Scholar] [CrossRef]
- Mulualem, D.M.; Agbavwe, C.; Ogilvie, L.A.; Jones, B.V.; Kilcoyne, M.; O’Byrne, C.; Boyd, A. Metagenomic identification, purification and characterisation of the Bifidobacterium adolescentis BgaC β-galactosidase. Appl. Microbiol. Biotechnol. 2021, 105, 1063–1078. [Google Scholar] [CrossRef]
- Viborg, A.H.; Katayama, T.; Abou Hachem, M.; Andersen, M.C.; Nishimoto, M.; Clausen, M.H.; Urashima, T.; Svensson, B.; Kitaoka, M. Distinct substrate specificities of three glycoside hydrolase family 42 β-galactosidases from Bifidobacterium longum subsp. infantis ATCC 15697. Glycobiology 2014, 24, 208–216. [Google Scholar] [CrossRef] [Green Version]
- Viborg, A.H.; Fredslund, F.; Katayama, T.; Nielsen, S.K.; Svensson, B.; Kitaoka, M.; Lo Leggio, L.; Abou Hachem, M. A β1-6/β1-3 galactosidase from Bifidobacterium animalis subsp. lactis Bl-04 gives insight into sub-specificities of β-galactoside catabolism within Bifidobacterium. Mol. Microbiol. 2014, 94, 1024–1040. [Google Scholar] [CrossRef]
- Pan, Q.; Zhu, J.; Liu, L.; Cong, Y.; Hu, F.; Li, J.; Yu, X. Functional identification of a putative beta-galactosidase gene in the special lac gene cluster of Lactobacillus acidophilus. Curr. Microbiol. 2010, 60, 172–178. [Google Scholar] [CrossRef]
- Xin, Y.; Guo, T.; Zhang, Y.; Wu, J.; Kong, J. A new β-galactosidase extracted from the infant feces with high hydrolytic and transgalactosylation activity. Appl. Microbiol. Biotechnol. 2019, 103, 8439–8448. [Google Scholar] [CrossRef]
- Hinz, S.W.A.; Van Den Broek, L.A.M.; Beldman, G.; Vincken, J.-P.; Voragen, A.G.J. β-Galactosidase from Bifidobacterium adolescentis DSM20083 prefers β(1,4)-galactosides over lactose. Appl. Microbiol. Biotechnol. 2004, 66, 276–284. [Google Scholar] [CrossRef]
- Fernandez-Lopez, L.; Pedrero, S.G.; Lopez-Carrobles, N.; Gorines, B.C.; Virgen-Ortiz, J.J.; Fernandez-Lafuente, R. Effect of protein load on stability of immobilized enzymes. Enzym. Microb. Technol. 2017, 98, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Hidaka, M.; Fushinobu, S.; Ohtsu, N.; Motoshima, H.; Matsuzawa, H.; Shoun, H.; Wakagi, T. Trimeric crystal structure of the glycoside hydrolase family 42 beta-galactosidase from Thermus thermophilus A4 and the structure of its complex with galactose. J. Mol. Biol. 2002, 322, 79–91. [Google Scholar] [CrossRef]
- Yang, S.; Xie, X.; Ma, J.; He, X.; Li, Y.; Du, M.; Li, L.; Yang, L.; Wu, Q.; Chen, W.; et al. Selective Isolation of Bifidobacterium From Human Faeces Using Pangenomics, Metagenomics, and Enzymology. Front. Microbiol. 2021, 12, 649698. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Cane, D.E. Stereospecific Formation of Z-Trisubstituted Double Bonds by the Successive Action of Ketoreductase and Dehydratase Domains from trans-AT Polyketide Synthases. Biochemistry 2018, 57, 3126–3129. [Google Scholar] [CrossRef]
- Herlet, J.; Kornberger, P.; Roessler, B.; Glanz, J.; Schwarz, W.H.; Liebl, W.; Zverlov, V.V. A new method to evaluate temperature vs. pH activity profiles for biotechnological relevant enzymes. Biotechnol. Biofuels 2017, 10, 234. [Google Scholar] [CrossRef] [Green Version]
- Nowak, P.; Zgirski, A. Effects of metal ions on activity of plasmin. Biol. Trace Elem. Res. 2003, 93, 87–94. [Google Scholar] [CrossRef]
- Deshavath, N.N.; Mukherjee, G.; Goud, V.V.; Veeranki, V.D.; Sastri, C.V. Pitfalls in the 3,5-dinitrosalicylic acid (DNS) assay for the reducing sugars: Interference of furfural and 5-hydroxymethylfurfural. Int. J. Biol. Macromol. 2020, 156, 180–185. [Google Scholar] [CrossRef] [PubMed]




| Metallic Cations (5 mM) | BLGLB1 (%) | BPGLB1 (%) | E (%) |
|---|---|---|---|
| Untreated | 100 | 100 | 100 |
| Zn2+ | 103 ± 2.6 | 122 ± 2.5 | 147 ± 1.4 |
| Co2+ | 87 ± 4.2 | 186 ± 5.1 | 141 ± 1.0 |
| Al3+ | 44 ± 0.1 | 96 ±1.7 | 60 ± 3.9 |
| Ni2+ | 83 ± 3.6 | 212 ± 1.1 | 156 ± 3.7 |
| Fe2+ | 94 ± 0.5 | 104 ± 1.8 | 132 ± 5.6 |
| Mg2+ | 96 ± 2.5 | 154 ± 1.9 | 162 ± 6.2 |
| Na+ | 104 ± 0.9 | 167 ± 2.0 | 121 ± 3.2 |
| Ca2+ | 104 ± 0.3 | 111 ± 4.6 | 104 ± 9.7 |
| Mn2+ | 101 ± 0.6 | 101 ± 1.1 | 161 ± 2.0 |
| Li+ | 109 ± 1.1 | 129 ± 13.2 | 115 ± 6.3 |
| Enzyme | BLGLB1 (U/mg) | BPGLB1 (U/mg) | E (U/mg) |
|---|---|---|---|
| pNPG | 4600 ± 24 | 4.1 ± 0.4 | 54 ± 3 |
| oNPG | 2200 ± 15 | 0.58 ± 0.03 | 300 ± 3.6 |
| 4-nitrophenol-α-galactoside | 100 ± 10 | 0.4 ± 0.03 | 0.5 ± 0.4 |
| 2-nitrophenol-β-glucoside | 12 ± 3 | 0.07 ± 0.01 | 0 |
| Lactose | 1.3 ± 0.1 | - | 1.6 ± 0.1 |
| Milk | 2.3 ± 0.2 | - | 3.1 ± 0.5 |
| Acidic whey | 4.8 ± 0.8 | - | 11 ± 0.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, M.; Yang, S.; Jiang, T.; Liang, T.; Li, Y.; Cai, S.; Wu, Q.; Zhang, J.; Chen, W.; Xie, X. Cloning, Expression, Purification, and Characterization of β-Galactosidase from Bifidobacterium longum and Bifidobacterium pseudocatenulatum. Molecules 2022, 27, 4497. https://doi.org/10.3390/molecules27144497
Du M, Yang S, Jiang T, Liang T, Li Y, Cai S, Wu Q, Zhang J, Chen W, Xie X. Cloning, Expression, Purification, and Characterization of β-Galactosidase from Bifidobacterium longum and Bifidobacterium pseudocatenulatum. Molecules. 2022; 27(14):4497. https://doi.org/10.3390/molecules27144497
Chicago/Turabian StyleDu, Mingzhu, Shuanghong Yang, Tong Jiang, Tingting Liang, Ying Li, Shuzhen Cai, Qingping Wu, Jumei Zhang, Wei Chen, and Xinqiang Xie. 2022. "Cloning, Expression, Purification, and Characterization of β-Galactosidase from Bifidobacterium longum and Bifidobacterium pseudocatenulatum" Molecules 27, no. 14: 4497. https://doi.org/10.3390/molecules27144497